Influence of mulch and soil compaction on earthworm cast properties
Corinna Buck
∗, Marcus Langmaack, Stefan Schrader
Zoologisches Institut der TU Braunschweig, AG Bodenzoologie, Spielmannstrasse 8, D-38092 Braunschweig, Germany
Received 3 August 1999; received in revised form 23 February 2000; accepted 25 February 2000
Abstract
The effects of different mulch materials applied to compacted and uncompacted soil on the quantity and the quality of deposited earthworm casts were investigated. Biochemical properties and water stability of soil aggregates were compared with the corresponding properties of worm casts. This short-time experiment was conducted in the laboratory, simulating field conditions of mulch management in temperate agricultural systems. In microcosmsLumbricus terrestrisandOctolasion cyaneumwere inoculated separately. Barley, lupin, maize, or sugar-beet as straw or leaves were applied as mulch in amounts comparable to those usually found in the field. The soil was compacted artificially to a bulk density of 1.0 or 1.5 Mg m−3.
In general, plant material and to a lesser extent soil compaction influenced the dynamic processes in the soil affecting microbial activity and water stable aggregation. Higher values of phosphatase activity was measured in compacted soil, while the corresponding enzyme activities in the casts were less affected by compaction. The worm species and the nutritional quality of the food source were factors strongly influencing water stable aggregation. Mulch as well as soil compaction had consequences for the burrowing activity of the worms, which resulted in different rates of cast production depending on the species. © 2000 Elsevier Science B.V. All rights reserved.
Keywords:Mulch; Soil compaction; Earthworm cast production; Enzyme activity; Water stable aggregation
1. Introduction
Soil biota plays an important role in supporting nutrient cycling as well as creating and stabilizing soil structure. Due to their dominant position in the soil animal biomass and high contribution to rates of soil turnover, earthworms are of special importance for soil fertility. The cultivation effect of earthworms is both the result of their burrowing activity and the consequence of their casting and mucus secretion. Agriculture may profit by earthworm activity, espe-cially as humans are dealing with the problem of soil degradation worldwide today. Increasing degradation
∗Corresponding author. Tel.:+49-531-391-3237;
fax:+49-531-391-8198.
E-mail address:[email protected] (C. Buck)
of soil structure as a consequence of soil erosion and soil compaction has been a growing problem in agriculture in recent decades. Modern farming prac-tices rely on the use of large, heavy machinery and on more vehicular traffic than in the past. In com-pacted soil living conditions for plant roots and soil organisms have deteriorated: water infiltration and gas exchange decrease (Ehlers, 1983) and colonizable habitats are limited following reduced pore space (Aritajat et al., 1977). Though earthworms may coun-teract soil compaction through burrowing and casting activity (e.g. Whalley et al., 1995) soil compaction, mostly in combination with intensified soil tillage, damages earthworm populations (Langmaack, 1999). In contrast to these demecological investigations, the present study aims at the autecology of earthworms as influenced by soil compaction. The focus of this
study is the casts of earthworms as compared with the parent soil. The gut passage of soil involves a physical and biochemical modification of the ingested material. Martin and Marinissen (1993) reviewed the effects of gut transit leading, e.g. to an accumulation of available bioelements, a growth of microbial popu-lations and an alterated aggregation status in the cast compared to corresponding soil. The experiments in the present study were conducted in the laboratory, simulating field conditions of mulch management in different crops. The consequences of soil compaction combined with different mulch material for the qual-ity and quantqual-ity of cast production by earthworms belonging to different ecotypes were examined.
2. Material and methods
2.1. Experimental design
A laboratory experiment was conducted in order to obtain the casts of two earthworm species produced in soil treatments varying in mulch material and degree of soil compaction (1.0 and 1.5 Mg m−3). Two
earth-worm species were kept separately in microcosms in which the mulch material served as food.
Lumbricus terrestrisL. 1758 and Octolasion
cya-neum (Savigny, 1926) are both common species in
agroecosystems of Middle Europe (Graff, 1953). L.
terrestrisbelongs to the anecic group of earthworms,
while O. cyaneum was chosen as a representive of
endogeic earthworms.
The plant species used as mulch material represent a variety of typical crops cultivated in Middle Europe
and demonstrate a wide range ofC/N-ratios: Leaves
and straw were taken from barley (Hordeum vulgare, C/N: 87.8) as a representative cereal crop and from
lupine (Lupinus angustifolius,C/N: 15.6), a green
ma-nure. Only the leaves were taken from maize (Zea
mays,C/N: 11.3) and sugar-beet (Beta vulgaris, ssp.
altissima, C/N: 10.9) representing a C4-plant and a
root crop, respectively. The air-dried plant material was cut into 0.5–1.5 cm fragments.
The earthworms were kept in special microcosms, based on those described by Evans (1947). They con-sisted of two parallel glass sheets (350 mm×250 mm)
of defined distance apart according to the body width of the earthworms and were filled with sieved
1–2 mm soil aggregates. The soil was taken from the Ap-horizon of a cultivated soil in southern Lower Sax-ony. It is characterized as a Pseudogley-Parabraunerde (Gleyic Podzoluvisol, FAO) derived from loess with a texture of loamy silt. After sieving, the air-dried aggregates were remoistened until a water content between 19 and 21 wt.% was reached. Before the soil was filled into the microcosms, 2.5 g mulch was mixed into the soil for each microcosm, with 0.2 g of this mulch being littered on the soil surface. The microcosms of the treatments with high bulk density were filled within a frame compressing the soil with a stiff plate from one side. This procedure ensures a homogeneous bulk density in addition to a homo-geneous distribution of the soil aggregates. The soil was filled from above in the uncompacted treatments. Two adult earthworms were inoculated into each
microcosm and kept for about 4 weeks at 10◦C in
a dark climate room. To determine their change of biomass the worms were weighed directly before in-oculation and again after removal from the soil at the end of the experiment.
In all, there were 16 treatments in the experiment (n=5) varying in soil bulk density (1.0 or 1.5 Mg m−3),
mulch material (barley, lupin, maize, or sugar-beet), and earthworm species (L. terrestrisorO. cyaneum). Eight microcosms without worms and two without worms and mulch were also established as reference treatments.
2.2. Analyses
phos-phate available for them. Alkaline (APA) and acid phosphatase activity (SPA) were measured following
p-nitrophenol release from p-nitrophenyl-phosphate. This method was suggested by Tabatabai and Brem-ner (1969) and modified by Margesin (1993) for an
incubation temperature of 37◦C. The assays of the
phosphatases only differ in the choice of the pH-value of the buffer (6.5 and 11). All enzyme assays were repeated three times per microcosm.
The remaining casts and soil within the microcosms were air-dried for measurements of water stable ag-gregation (WSA in %). WSA of reference soil and cast aggregates of the same size was determined by a modified wet-sieving-method of Kemper and Rosenau (1986), calculated from the difference of the weight of the aggregates before and after the sieving proce-dure (n=5). A sieve with 0.2 mm mesh size was used,
rotating 38 rpm with a stroke of 40 mm for 3 min. The cast production data are presented as dry matter
per gram live weight of worms per day (mg DM g−1
LW per day).
2.3. Statistics
The Kolmogorov–Smirnov test confirmed that all the values were normally distributed. Effects of soil and cast aggregates, mulch type, and soil compaction on enzyme activities were analysed by three-factor analysis of variance (3-ANOVA) and the studentt-test was used to compare treatment means. Relationships between different parameters were investigated with regression analysis. All statistical analysis was done using the software package SPSS.
3. Results
3.1. Cast production (CP)
The cast production ofL. terrestrisranged from 280
to 431 mg DM g−1LW per day whereasO. cyaneum
deposited 2–3.5-fold (852–1305 mg DM g−1LW per
day) as much in the corresponding mulch types, as shown in Fig. 1. Except for barley,L. terrestris pro-duced more casts in compacted soil than in soil with low bulk density, but not significantly so.O. cyaneum
egested more casts in the uncompacted soils, except for the lupin treatments. The introducedL. terrestris
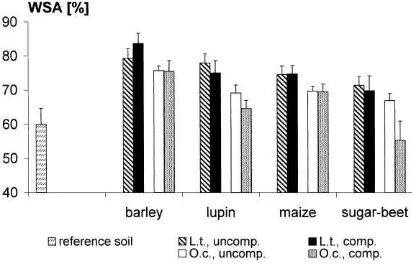
Fig. 1. Arithmetic means (±S.D.) of cast production (CP) by earthworms (L.t.=Lumbricus terrestris,O.c.=Octolasion cyaneum) in relation to soil bulk density (uncomp.=uncompacted soil, comp.=compacted soil) and mulch types (n=5).
weighed on average 4.77 g, the average weight of
O. cyaneumwas 2.29 g before inoculation. At the end of the experiment,L. terrestrishad an average weight
of 4.84 g, andO. cyaneumweighed 3.35 g.
3.2. Enzyme activities
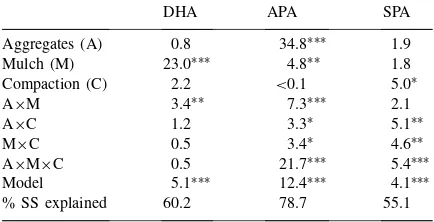
According to 3-ANOVA, mulch types strongly af-fected enzyme activity in the cases of DHA and APA (Table 1). The influence of soil and cast aggregates of
L. terrestrisandO. cyaneum was only significant in the case of APA. Significant effects of soil compaction were only found for SPA (p=0.028). All two-way and
three-way interactions were significant for APA and
Table 1
F-values from three-way analysis of variance (3-ANOVA) on the effects of aggregates (soil and earthworm casts), mulch type, and soil compaction on dehydrogenase (DHA), alkaline phosphatase (APA), and acid phosphatase (SPA) activity
DHA APA SPA
Aggregates (A) 0.8 34.8∗∗∗ 1.9
Mulch (M) 23.0∗∗∗ 4.8∗∗ 1.8
Compaction (C) 2.2 <0.1 5.0∗
A×M 3.4∗∗ 7.3∗∗∗ 2.1
A×C 1.2 3.3∗ 5.1∗∗
M×C 0.5 3.4∗ 4.6∗∗
A×M×C 0.5 21.7∗∗∗ 5.4∗∗∗
Model 5.1∗∗∗ 12.4∗∗∗ 4.1∗∗∗
% SS explained 60.2 78.7 55.1
∗p
<0.05.
∗∗p
<0.01.
∗∗∗p
Table 2
Arithmetic mean values (±S.D.) for three types of enzyme activity in worm casts (n=20) and in soil (n=12)a
DHA (mg TPF g−1 per day) APA (mg NP g−1h−1) SPA (mg NP g−1h−1)
Uncompacted soil
Soil aggregates 236.005ab±50.077 353.401c±30.445 237.56ab±25.483
L. terrestriscasts 255.722a±30.111 386.682bd±58.630 258.904ab±42.733
O. cyaneumcasts 245.527ab±33.293 424.094a±50.948 273.112a±38.864
Compacted soil
Soil aggregates 236.718ab±43.017 360.215cd±18.601 252.859ab±26.119
L. terrestriscasts 236.465b±27.630 386.412b±49.516 242.432b±35.897
O. cyaneumcasts 239.443ab±35.238 400.439ab±28.383 243.553b±28.973
aThe data for the various types of plant residues are pooled. Significant differences between means within columns based on
Student’s-t-test are indicated by different letters (p<0.05). DHA=dehydrogenase activity, APA=alkaline phosphatase activity, SPA=acid phosphatase activity.
SPA except for the aggregate×mulch interaction in the
case of SPA, while only the aggregate×mulch
inter-action was significant in the case of DHA.
The influence of soil bulk density on enzyme activ-ity in soil and earthworm casts is illustrated in Table 2. APA in the worm casts was significantly enhanced compared with soil. Both phosphatase activities were higher inO. cyaneumcasts than inL. terrestriscasts, but the differences were only significant for APA in the uncompacted treatments (n=20;p=0.042).
The DHA, APA and SPA values in casts deposited by O. cyaneum in compacted soil were lower than those for the uncompacted treatments, but were only significantly so for SPA (n=20;p=0.01). In the case
ofL. terrestriscasts, lower levels were also found in compacted soils, but this effect was only significant
for DHA (n=20; p=0.042). APA and DHA values
were similar, while SPA values were slightly increased in compacted soil aggregates in comparison with un-compacted soil but this difference was not statistically significant.
3.3. Water stable aggregation (WSA)
Fig. 2 shows significant differences for water sta-bility (WSA) between reference soil and cast aggre-gates, except for the casts produced byO. cyaneumin compacted soil with sugar-beet mulch. The WSA of
L. terrestris casts ranged from 70% with sugar-beet up to 84% with barley mulch, while the values for
O. cyaneumcasts were between 55 and 75% in corre-sponding treatments. Barley had the strongest
stabiliz-ing effect on worm caststabiliz-ings, sugar-beet had the lowest. Higher bulk density was associated with lower WSA values forO. cyaneumcasts (n=5;p=0.051). Except
for the barley treatments the same tendency was shown forL. terrestriscasts, but this was not significant.
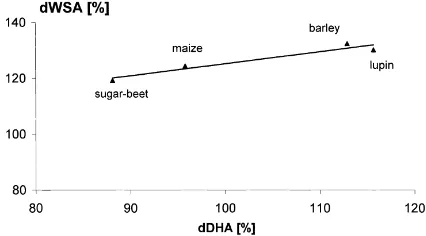
A positive correlation was found between WSA and enzyme activity in the worm casts, when analyzed within each compaction level. With increasing dDHA
(%) (r2=0.9252; n=5; p=0.0381) the dWSA (%)
of the cast aggregates produced by L. terrestriswas
significantly enhanced at low soil compaction (Fig. 3). This relationship was also found for the casts of
O. cyaneum to a lesser extent. There was a linear relationship between the dWSA (%) and the dAPA
Fig. 3. Correlation between relative dehydrogenase activity (dDHA (%)) and relative water stable aggregation (dWSA (%)) for casts of
Lumbricus terrestrisdeposited in uncompacted soil (n=5). dDHA (%) is the percentage of dehydrogenase activity in the casts in relation to the soil. dWSA (%) is the percentage of the water stable aggregation in the casts in relation to the soil. The data were calculated from the arithmetic means y=0.4286x+82.308;
r2=0.9252;p=0.0381.
(%) of cast aggregates produced by O. cyaneum in
compacted soil (r2=0.8391;p=0.084).
4. Discussion
4.1. Cast production
Earthworm cast production is highly dependent on the ecophysiology of the species.L. terrestrisis detri-tivorous and feeds on plant debris at the soil surface,
whereasO. cyaneumis geophagous and feeds on
min-eral horizon material which is poorer in organic mat-ter. This highly selective consumption explains why the detritivorous species needs less food to satisfy its energy requirement. To reach this required level the geophageous species has to ingest more material of lower nutritional quality, resulting in a 2–3.5-fold higher cast production compared toL. terrestris. Com-paring cast production byL. terrestriswith results of other authors (e.g. Shipitalo and Protz (1989) with 70–180 mg g−1per day) the values generally cited are
relatively low because most studies considered only the casts deposited on the soil surface. The results of Flegel et al. (1998), who also collected casts from the whole bulk soil, are similar to ours (228–461 mg g−1
per day). For O. cyaneum, Scheu (1987) found ca.
50% higher cast production (1880 mg g−1per day) in a beechwood soil.
Comparison of cast production at the two levels of soil bulk density suggests thatL. terrestrisis no longer able to push the soil particles aside, but is forced to ingest a higher amount of soil to construct its burrows ´ın compacted soil. These results agree with earlier ob-servations by Joschko et al. (1989), who carried out column experiments with the same species. Physiolog-ical investigations of radial and axial pressures of the earthworm muscle system correspond to these findings
(Keudel and Schrader, 1999).O. cyaneumproduced a
lower amount of castings in compacted soil as a con-sequence of reduced burrowing activity as indicated by the smaller burrow system that was observed in the microcosms.
4.2. Enzyme activity
The destiny of ingested microorganisms depends on their adaptation to the intestinal conditions of the earthworm (Moody et al., 1995). Two oppos-ing processes act duroppos-ing digestion. On the one hand favourable pH-value and increased nutrient and water supply in the gut increases the microbial population during gut passage (Barois and Lavelle, 1986), while on the other hand intestinal transit can reduce numbers of some species of bacteria by digestion (Devliegher and Verstraete, 1995).
casts produced in uncompacted soil compared with those from compacted soil could be that the closer grain packing of the compacted material makes the gut muscles less effective in grinding and mixing the gut content. The opposite is true for the uningested soil aggregates which showed higher SPA activity in the compaction treatment compared with uncom-pacted conditions. It may be that comuncom-pacted soil offers favourable conditions for colonization with decomposer microflora on the basis of closer contact of mineral and plant particles.
4.3. Water stable aggregation
These experiments reveal the stabilizing effect of organic residues during gut passage on soil aggregates. Flegel et al. (1998), who carried out comparable labo-ratory experiments, reported increase in water stability
of casts produced byL. terrestris of 90–290%,
com-pared with soil aggregates. The moulding processes in the gut destroys original linkages between the par-ticles to a great extent. But after egestion, microbial activity, age-hardening, and drying processes connect the mineral and organic compounds again and new bonds can be formed, leading to higher WSA than be-fore ingestion (Shipitalo and Protz, 1989). Fungal hy-phae are another binding agent, preferring substrates high in organic matter content, such as casts, because fungi are among the most important organisms in the decomposition process (Swift et al., 1979).
Differences in the stability of cast aggregates be-tween L. terrestris and O. cyaneum reflect different food selection by these species. Schrader and Zhang (1997) concluded, that during gut transit inL. terrestris
more plant material comes in contact with mineral par-ticles, because the percentage of organic matter in the food of detritivorous earthworms is much higher than for geophageous species.
The effect of casting on the stabilization of soil structure strongly depends on the material serving as food source to the worm. It is well established, that organic material has a stabilizing effect on WSA (Tisdall and Oades, 1982). In addition to the amount of organic matter, the efficiency on the stability of ag-gregates is determined by the decomposability of the organic compound in the substrate (Skinner, 1979). Readily available substrates, such as sugars (e.g. polysaccarides) increase WSA, but this effect is only
temporary because the glues are decomposed read-ily by rapidly growing microbial populations. This is true for sugar-beet and maize leaves, which have high proportions of readily available sucrose. Their high nutritional quality results from low C/N-values of 11 and relatively low fibre contents of 2.1% (sugar beet) and 28% (maize) (Brouwer, 1972). In contrast, the high mechanical strength based on high val-ues of silicic acid and 40% fibre content (Brouwer, 1972) makes barley less attractive for microbial at-tack and has a pronounced stabilizing effect on the casts. Microorganisms play a central role in aggre-gate stability (Anderson, 1991). They directly serve as binding agents or produce polysaccharides and gum material for the same purpose. The present pos-itive correlation between WSA and enzyme activity clearly demonstrates this relationship. Like enzyme
activities, WSA of cast aggregates of O. cyaneum
is particularly reduced in the compacted treatments. Furthermore, the compaction process causes stresses to the aggregates resulting in a loss of strength and an increase in failure zones in the aggregates (Horn, 1993). Watts et al. (1996), who compared WSA of soil aggregates collected prior and following to plough-ing, observed a destabilizing effect of tillage on soil aggregates.
5. Conclusions
impact of soil compaction is less important than mulch quality for cast properties.
Acknowledgements
We thank Prof. Dr. J.P. Curry, Prof. Dr. O. Larink and Prof. Dr. H.G. Wolff for valuable comments. We also thank Dr. Nortmeier (BBA, Braunschweig) and many collegues for technical advice. The study was supported by the DFG (German Reseach Foundation) Bonn, Germany.
References
Anderson, T.-H., 1991. Bedeutung der Mikroorganismen für die Bildung von Aggregaten im Boden. Z. Pflanzenernähr. Bodenkd. 154, 409–416.
Aritajat, U., Madge, D.S., Gooderham, P.T., 1977. The effects of compaction of agricultural soils on soil fauna. 1. Field investigations. Pedobiologia 17, 262–282.
Barois, I., Lavelle, P., 1986. Changes in respiration rate and some physicochemical properties of a tropical soil during transit throughPontoscolex corethrurus. Soil Biol. Biochem. 18, 539– 541.
Brouwer, W., 1972. Handbuch des speziellen Pflanzenbaus, Bd. 1 und Bd. 2. Paul Parey, Berlin, Hamburg.
Businelli, M., Perucci, P., Patumi, P., Guisqiuani, P.L., 1984. Chemical composition and enzymic activity of some worm casts. Plant and Soil 80, 417–422.
Casida, L.E., Klein, D.A., Santoro, T., 1964. Soil dehydrogenase activity. Soil Sci. 98, 371–376.
Devliegher, W., Verstraete, W., 1995. Lumbricus terrestris in a soil core experiment: nutrient-enrichment processes (NEP) and gut associated processes (GAP) and their effect on microbial biomass and microbial activity. Soil Biol. Biochem. 27, 1573– 1580.
Ehlers, W., 1983. Auswirkungen der Bodenbelastung mit schwerem Gerät und der Bodenbearbeitung auf das Bodengefüge und das Pflanzenwachstum, Kali-Briefe. Büntehof. 16, 499–516. Evans, A.C., 1947. A method of studying the burrowing activities
of earthworms. Ann. Mag. Nat. Hist. 11, 643–650.
Flegel, M., Schrader, S., Zhang, H., 1998. Influence of food quality on the physical and chemical properties of detrivorous earthworm casts. Appl. Soil Ecol. 9, 263–269.
Graff, O., 1953. Die Regenwürmer Deutschlands. Schriftenreihe der FAL. Braunschweig Völkenrode, Heft 7. Verlag Schaper, Hannover, 81 pp.
Horn, R., 1993. Mechanical properties of structured unsaturated soils. Soil Technol. 6, 47–75.
Joschko, M., Diestel, H., Larink, O., 1989. Assessment of earthworm burrowing efficiency in compacted soil with a combination of morphological and soil physical measurements. Biol. Fertil. Soils 8, 191–196.
Kemper, W.D., Rosenau, R.C., 1986. Aggregate stability and size distribution. In: Klute, A. (Ed.), Methods of Soil Analysis. Part 1. Physical and Mineralogical Methods. Agronomy Monograph Nr. 9, 2nd Edition, pp. 425–442.
Keudel, M., Schrader, S., 1999. Axial and radial pressure exerted by earthworms of different ecological groups. Biol. Fertil. Soils 29, 262–269.
Kristufek, V., Ravaz, K., Pizl, V., 1992. Changes in densities of bacteria and microfungi during gut transit inLumbricus rubellus
and Aporrectodea caliginosa, Oligochaeta, Lumbricidae. Soil Biol. Biochem. 24, 1499–1500.
Langmaack, M., 1999. Earthworm communities in arable land influenced by tillage, compaction, and soil. Z. Ökologie u. Naturschutz 8, 11–21.
Malkomes, H.D., 1993. Eine modifizierte Methode zur Erfassung der Dehydrogenaseaktivitat, TTC-Reduktion im Boden nach Herbizidanwendung. Nachrichtenbl. Deut. Pflanzenschutzd. 45, 180–185.
Margesin, R., 1993. Bestimmung der sauren und alkalischen Phosphomonesterase-Aktivität. In: Schinner, F., Kandeler, E., Öhlinger, R., Margesin, R. (Eds.), Bodenbiologische Arbeitsmethoden. Springer, Berlin, pp. 200–203.
Martin, A., Marinissen, J.C.Y., 1993. Biological and physico-chemical processes in excrements of soil animals. Geoderma 56, 331–347.
Moody, S.A., Briones, M.J.I., Piearce, T.G., Dighton, J., 1995. Selective consumption of decomposing wheat straw by earthworms. Soil Biol. Biochem. 27, 1209–1213.
Satchell, J.E., Martin, K., 1984. Phosphatase activity in earthworm faeces. Soil Biol. Biochem. 16, 191–194.
Scheu, S., 1987. The role of substrate feeding earthworms (Lumbricidae) for bioturbation in a beechwood soil. Oecologia 72, 192–196.
Schrader, S., Zhang, H., 1997. Earthworm casting: stabilization or destabilization of soil structure? Soil Biol. Biochem. 29, 469– 475.
Shipitalo, M.J., Protz, R., 1989. Chemistry and micromorphology of aggregation in earthworm casts. Geoderma 45, 357–374. Skinner, F.A., 1979. Rothamsted studies of soil structure. VII. The
effects of incubation on soil aggregate stability. J. Soil Sci. 30, 473–481.
Soane, B.D., van Ouwerkerk, C., 1994. Soil compaction problems in world agriculture. In: Soane, B.D., van Ouwerkerk, C. (Eds.), Soil Compaction in Crop Production. Elsevier, Amsterdam, pp. 1–21.
Swift, M.J., Heal, O.W., Anderson, J.M., 1979. Decomposition in Terrestrial Ecosystems. Studies in Ecology, Vol. 5. University of California Press, Berkley, Los Angeles.
Tabatabai, M.A., Bremner, J.M., 1969. Use of p -nitrophenyl-phosphate for assay of soil phosphatase activity. Soil Biol. Biochem. 1, 301–307.
Tisdall, J.M., Oades, J.M., 1982. Organic matter and water stable aggregates in soils. J. Soil Sci. 33, 141–163.
Watts, C.W., Dexter, A.R., Longstaff, D.J., 1996. An assessment of the vulnerability of soil structure to destabilisation during tillage. Part II. Field trials. Soil Till. Res. 37, 175–190. Whalley, W.R., Dumitru, E., Dexter, A.R., 1995. Biological effects