ABSTRAK
Peningkatan resistensi udang windu Penaeus monodon melalui transfer gen antivirus belum pernah dilaporkan. Penelitian ini bertujuan untuk mengetahui pola ekspresi gen antivirus PmAV pada embrio dan larva, serta performa udang windu transgenik F0 setelah ditantang dengan virus WSSV. Transfer gen antivirus ke embrio udang dilakukan dengan metode transfeksi menggunakan larutan jetPEI. Pengamatan ekspresi gen antivirus PmAV dilakukan pada 12, 18, 24, dan 30 jam setelah transfeksi (jst). Uji tantang dilakukan pada larva udang windu PL-25 metode perendaman dengan menggunakan isolat WSSV dari udang windu yang terinfeksi secara alami. Pengamatan kelangsungan hidup dan ekspresi gen antivirus PmAV dilakukan setelah uji tantang. Hasil penelitian menunjukkan bahwa penggunaan larutan transfeksi jetPEI dan gen antivirus PmAV tidak memberikan efek yang membahayakan bagi embrio udang. Ekspresi gen PmAV mulai terlihat pada pengamatan 12 jst dan selanjutnya meningkat sampai mencapai puncak pada 24 jst dan kemudian terjadi penurunan ekspresi pada 30 jst. Hasil penelitian juga menunjukkan bahwa ekspresi gen PmAV meningkat
(up-regulated) akibat infeksi virus WSSV. Selain itu, udang windu transgenik F0
memperlihatkan resistensi (kelangsungan hidup 95,6%) lebih tinggi (P<0,05) dibandingkan dengan udang windu kontrol non-transgenik (kelangsungan hidup 71,1%). Berat dan panjang total tubuh udang windu transgenik F0 umur 1,5 bulan tidak memperlihatkan perbedaan yang signifikan (P>0,05) dengan udang windu non-transgenik (kontrol).
Kata kunci: ekspresi gen, gen antivirus, transfeksi, uji tantang, WSSV, udang
EXPRESSION OF ANTIVIRAL GENE ON EMBRYO AND
LARVAE OF TIGER SHRIMP Penaeus monodon
ABSTRACT
No study on enhanced diseases resistance of tiger shrimp has been reported. The aim of this research was to determine the transient expression pattern of antiviral gene on embryo as well as larvae, and the performance of F0 transgenic larvae by challenged with WSSV. Transfer of antiviral gene into the embryo was done by transfection method using jetPEI reagent. Expression of
PmAV gene was observed at 12, 18, 24, and 30 hours after transfection (hat). The
challenged test to PL-25 larvae was conducted by water borne infection using the WSSV isolated from infected shrimp. Survival rate and PmAV antiviral gene were observed after challenge. The results indicated that the jetPEI reagent and antiviral PmAV gene did not show the toxicity to the shrimp larvae. The transient expression of PmAV gene was detected at 12 hat until reaching the peak within 24 hat, and then decreased slightly at 30 hat. The result also showed that PmAV expression was up-regulated during infection. When challenged with WSSV, the transgenic shrimp exhibited higher (P<0.05) resistance to the WSSV infection (95.6% survived) as compared to the control shrimp (71.1% survived). The body weight and length of 1.5 months transgenic shrimp did not show significantly difference (P>0.05) with the non-transgenic shrimp.
Keywords: gene expression, antiviral gene, transfection, challenge test, WSSV,
tiger shrimp.
PENDAHULUAN
Teknologi transgenesis memberikan peluang besar dalam mengembangkan strain yang memiliki resistensi yang tinggi terhadap patogen penyebab penyakit. Salah satu upaya potensial yang dapat dilakukan dalam peningkatan resistensi penyakit adalah produksi hewan akuatik transgenik yang mengandung gen peptida antibakteri. Penemuan protein antibakteri cecropin pada insekta oleh Steiner et al. (1981) merupakan awal dari penelitian antimikroba, yang selanjutnya diidentifikasinya beberapa antimikroba lainnya. Beberapa antimikroba telah diidentifikasi pada mamalia (Lehrer et al. 1993), ampibi (Bevins & Zasloff 1990), dan pada insekta (Hoffmann et al. 1996) dimana organisme yang mengandung
gen pengkode tersebut memperlihatkan resistensi yang lebih baik terhadap penyakit.
Pada krustase, keberhasilan kloning dan karakterisasi gen pengkode antimikroba memberikan harapan dalam mempelajari lebih dalam mengenai peranan gen antimikroba pada udang. Aktivitas antimikroba yang berperan dalam imunitas non-spesifik pada krustase telah dilaporkan oleh beberapa peneliti sebelumnya, misalnya penaeidin (Wang et al. 2006; Jiravanichpaisal et al. 2007; Perdomo-Morales et al. 2007; Ho & Song 2009), proPO (prophenoloxidase) (Destoumieux et al. 1997; 2000a; 2000b; Sritunyalucksana et al. 1999; Wang et al. 2006; Jiravanichpaisal et al. 2007; Wang et al. 2007; Ai et al. 2008; Wang & Zhang 2008; Yeh et al. 2009); serine protease (Amparyup et al. 2007; Shi et al. 2009), lisozim (Vega et al. 2006; Burge et al. 2007), dan lektin (Denis et al. 2003; Ma et al. 2007; Sun et al. 2007; Zhang et al. 2009). Studi-studi in vitro tersebut mengindikasikan bahwa penggunaan konstruksi gen antimikroba dapat meningkatkan resistensi terhadap patogen.
Anderson et al. (1996) pertama kali membuktikan secara in vivo bahwa
resistensi ikan rainbow trout Oncorhynchus mykiss dapat ditingkatkan melalui transfer gen coat protein virus. Introduksi konstruksi peptida cecropin-B meningkatkan resistensi terhadap bakteri sampai dengan empat kali lipat pada ikan channel catfish (Dunham et al. 2002). Hal yang sama juga ditunjukkan pada ikan medaka transgenik yang memiliki resistensi lebih tinggi dibandingkan dengan ikan non-transgenik terhadap bakteri Pseudomonas sp. dan Vibrio sp. (Sarmasik et al. 2002). Sementara itu, pendekatan rekombinan DNA khususnya vaksin DNA telah mulai diaplikasikan pada akuakultur. Penyuntikan salmon Atlantik dengan plasmid pengkode glikorotein IHNV dengan pengontrol promotor
pCMV menunjukkan proteksi yang signifikan dengan adanya pembentukan
antibodi penetral virus setelah imunisasi dan titernya meningkat setelah uji tantang (Traxler et al. 1999). Seperti halnya dengan antimikroba lainnya, gen lisozim telah dilaporkan merupakan salah satu gen pengkode ketahanan penyakit khususnya antimikroba yang tidak spesifik (Austin & Allen-Austin 1985). Dengan menggunakan konstruksi promoter AFP ocean pout dan gen lisozim yang
diintroduksi ke ikan salmon menunjukkan adanya kemampuan melawan berbagai jenis mikroba.
Pada krustase khususnya udang, peningkatan resistensi pada level molekular masih terbatas. Penemuan gen pengkode antimikroba penaeidin membuka peluang dalam peningkatan immunitas udang melawan serangan patogen. Aplikasi penaeidin telah memperlihatkan efek peningkatan resistensi pada udang vaname Litopenaeus vannamei (Destoumieux et al. 1997). Induksi imun pada udang melalui vaksinasi telah dilaporkan dengan penggunaan rekombinan protein WSSV pada udang Penaeus chinensis (Kim et al. 2004), dan RNA untai ganda (double-stranded RNA, dsRNA) pada udang L. vannamei (Robalino et al. 2004). Transfer gen antivirus baru dimulai pada udang vaname L.
vannamei melalui introduksi gen penyandi coat protein dari TSV (TSV-CP) (Sun et al. 2005). Lu & Sun (2005) melaporkan bahwa dengan introduksi gen TSV-CP,
udang vaname transgenik memperlihatkan kelangsungan hidup yang signifikan lebih tinggi dibandingkan dengan udang normal (non-transgenik). Berdasarkan hasil-hasil penelitian tersebut, maka upaya peningkatan resistensi udang windu memberikan harapan yang cerah untuk dapat dilakukan. Penelitian ini bertujuan untuk mendapatkan konstruksi gen antivirus PmAV, mengetahui pola ekspresi gen antivirus pada embrio dan larva, serta performa larva udang windu transgenik melalui uji tantang dengan virus WSSV.
BAHAN DAN METODE
Pembuatan Konstruksi Gen Antivirus PmAVKonstruksi gen antivirus pProAV-PmAV dibuat dengan menyambungkan atau meligasikan sekuen promoter ProAV (Parenrengi et al. 2009a) dan cDNA
PmAV (Parenrengi et al. 2009b). Promoter ProAV diligasi pada situs BamHI, di
bagian hulu atau upstream dari sekuen cDNA PmAV yang diligasi pada situs SalI dalam vektor pBlueskript-SK (Stratagen). Skema pembuatan konstruksi gen
pProAV-PmAV disajikan pada Lampiran 11. Uji orientasi ligasi ProAV dan PmAV
ccc aca ctc cat caa -3’dan PmAVSalI-R 5’- ttg tcg act cct tta gaa tat tta ttc ttg-3’. Reaksi PCR yang digunakan adalah 0,05 µL Taq Polimerase; 1 µL 10 X buffer; 0,8 µL dNTP mix; 0,8 µL MgCl2; 10 ρmol masing-masing primer; 1 µL templat
DNA; dan 4,35 µL SDW. Program PCR yang digunakan adalah: suhu pre-denaturasi 94oC selama 3 menit, 30 siklus untuk (denaturasi 94oC selama 30 detik, annealing 60oC selama 30 detik dan ekstensi 72oC selama 40 detik), serta final ekstensi 72oC selama 3 menit. Hasil PCR dielektroforesis pada gel agarosa 1,0% untuk melihat fragmen DNA yang terbentuk. Klon bakteri yang memperlihatkan hasil PCR dengan fragmen sekitar 1,2 kb menunjukkan arah ligasi yang sesuai.
Pematangan Induk Udang Windu
Pembenihan udang windu dilakukan berdasarkan prosedur standar operasional perbenihan udang windu. Prosedur pematangan induk udang windu dilakukan melalui beberapa tahapan seperti yang telah dijelaskan pada Bab IV.
Transfeksi Gen Antivirus
Bakteri yang mengandung plasmid pProAV-PmAV dikultur menggunakan media LB. Plasmid pProAV-PmAV diisolasi dari bakteri menggunakan GF-1
Plamid DNA Extraction Kit (Vivantis) dengan mengikuti prosedur manual kit.
Kultur bakteri dan isolasi plasmid dilakukan seperti yang dijelaskan pada Bab IV. Kuantitas dan kualitas isolat plasmid diukur dengan menggunakan UV-VIS spektrofotometer pada panjang gelombang 260 nm dan 280 nm. Kuantitas (konsentrasi) plasmid dihitung dengan mengacu pada rumus yang telah dikembangkan oleh Linacero et al. (1998), sedangkan kualitas (kemurnian) plasmid dihitung dari rasio absorpsi 260 nm dan 280 nm (OD260/OD280).
Koleksi telur udang windu yang baru memijah dan prosedur transfeksi menggunakan plasmid pProAV-PmAV mengacu pada standar protokol transfeksi konstruksi pProAV-EGFP yang telah diuraikan pada Bab IV.
Uji Tantang Larva dengan WSSV
Pengamatan performa udang windu hasil transfer gen antivirus PmAV dilakukan melalui uji tantang larva dengan WSSV, dimana larva non-transgenik digunakan sebagai pembanding atau kontrol. Larva udang windu transgenik dipelihara berdasarkan prosedur standar pemeliharaan larva sampai dengan mencapai stadia PL-25. Penyiapan virus WSSV berasal dari udang windu yang terinfeksi secara alamiah. Inokulasi virus tersebut diisolasi dengan cara cairan udang disentrifugasi pada kecepatan 6.000 rpm selama 15 menit dan supernatan disaring dengan kertas filter 0,45 µm.
Wadah percobaan disucihamakan dengan perendaman kaporit 30 ppm selama satu hari, kemudian dinetralkan dengan natrium thiosulfat 30 ppm. Wadah diisi dengan air laut yang telah difilter dengan membran filter sebanyak 2 liter per wadah. Larva udang windu PL-25 dimasukkan ke dalam wadah dengan kepadatan 15 ekor/wadah (lihat Lampiran 10). Inokulum WSSV diinfeksikan ke larva udang dengan konsentrasi 2 mL/liter mengacu pada nilai LC-50 yang telah dilakukan oleh Nurhidayah (2009). Perlakuan pada percobaan ini adalah uji tantang WSSV terhadap larva udang windu transgenik (A) dan non-transgenik (B), dan kontrol larva udang tanpa uji tantang (C) masing-masing 4 ulangan (1 ulangan untuk pengamatan ekspresi gen antivirus dan 3 ulangan untuk mengamatan kelangsungan hidup larva). Selama percobaan, udang uji diberi pakan larva berupa pelet secara ad libitum dengan pemberian 3 kali per hari (pagi, siang, dan sore) selama 5 hari pemeliharaan. Perkembangan laju infeksi diamati secara visual dengan memperhatikan perubahan penciri patofisiologisnya, serta pengamatan mortalitas dan pengambilan sampel hepatopankreas larva dilakukan pada 6 jam, 12 jam, 1 hari, 2 hari, 3 hari, 4 hari, dan 5 hari setelah uji tantang.
Analisis Ekspresi Gen PmAV
Pengamatan ekspresi gen antivirus PmAV dilakukan dengan teknik PCR semi-kuantitatif. Pada pengamatan ekspresi sementara gen antivirus PmAV, RNA total (dilanjutkan dengan sintesis cDNA) diekstraksi dengan menggunakan 50 sampel (pooled sample) untuk analisis ekspresi PmAV pada embrio dan larva
udang windu. Analisis ekspresi gen antivirus PmAV dilakukan dengan menggunakan teknik RT-PCR. Pengambilan sampel dilakukan dalam beberapa tahap pengamatan, yakni: 12, 18, 24, dan 30 jam setelah transfeksi (jst) Penghitungan derajat penetasan larva dilakukan setelah telur diinkubasi selama 24 jam. Gen antivirus PmAV dan ekspresinya dideteksi dengan teknik PCR semi-kuantitatif, dimana sampel telur tanpa transfeksi digunakan sebagai kontrol dalam penelitian ini.
Konfirmasi masuknya gen PmAV pada larva secara individu dilakukan dengan mengekstraksi DNA genom pada delapan ekor larva udang windu berukuran 0,15±0,05 g yang diambil secara acak untuk dua tahap pelaksanaan transfeksi. Primer yang digunakan adalah PmAV-F 5’-tag tgc atg cat atg ggt cat aca atc cta-3’ dan PmAV-R 5’-ctg tct cga gct atg tgt cct gct ttc aca-3’,dengan menggunakan DNA genom sebagai templat PCR. Target DNA fragmen gen antivirus PmAV adalah pada posisi sekitar 513 bp.
Pada pengamatan ekspresi gen antivirus PmAV pada uji tantang dengan
WSSV, RNA diekstraksi dari 10 mg hepatopankreas larva dengan menggunakan
kit isogen seperti yang dijelaskan pada isolasi gen antivirus pada Bab III. Secara singkat, isolasi RNA dilanjutkan dengan sintesis cDNA menggunakan kit
Ready-To-Go You-Prime Fisrt Strand Beads (GE Healthcare). Hasil cDNA tersebut
dijadikan templat DNA dalam proses amplifikasi PCR dengan menggunakan primer PmAV-F: 5’-tag tgc atg cat atg ggt cat aca atc cta-3’ dan PmAV-R: 5’-ctg tct cga gct atg tgt cct gct ttc aca-3’, dengan target fragmen sepanjang 513 bp. Ekspresi gen β-aktin udang windu digunakan sebagai kontrol ekspresi gen seperti yang telah dikembangkan oleh Sriphaijit & Senapin (2007). Hasil PCR dielektroforesis untuk mengetahui pola ekspresi gen PmAV berdasarkan ketebalan pita yang terbentuk pada gel agarosa.
Proses amplifikasi fragmen gen antivirus PmAV dijalankan pada mesin PCR GenAmp AB-7200 (Applied Biosystem) dengan program suhu pre-denaturasi 94oC selama 3 menit; 35 siklus untuk (denaturasi 94oC selama 30 detik,
annealing 58oC selama 30 detik, dan ekstensi 72oC selama 45 detik); serta final
ekstensi 72oC selama 3 menit. Untuk melihat keberhasilan amplifikasi fragmen DNA target, hasil PCR dieletroforesis pada gel agarose 1,0% dan didokumentasi
dengan Gel Documentation System (Biometra). Untuk menentukan berat molekul fragmen DNA digunakan marker VC 100bp Plus DNA Ladder (Vivantis).
Udang windu transgenik F0 yang dihasilkan dipelihara dalam bak terkontrol dengan kepadatan 150 ekor/ton. Pakan berupa pelet dengan dosis 30% dari berat badan diberikan pada pagi dan sore hari. Untuk mengetahui respons pertumbuhan udang windu transgenik, pengukuran panjang total dan berat tubuh dilakukan pada 10 ekor udang windu pada umur 47 hari. Udang windu non-transgenik dijadikan sebagai pembanding (kontrol).
Analisis Data
Hasil cracking, uji orientasi konstruksi gen pProAV-PmAV, ekspresi gen
antivirus pada embrio dan larva, dan keberadaan gen antivirus PmAV pada larva udang windu disajikan secara deskriptif. Kelangsungan hidup larva udang windu yang ditantang dengan WSSV dianalisis ragam (ANOVA), serta berat dan pajang total udang windu dianalisis dengan uji-t menggunakan program Statistix Versi 3.0 (NH Analytical Software) dan dilanjutkan dengan uji beda nyata terkecil pada taraf 5%. Ekspresi gen antivirus PmAV di hepatopankreas larva udang windu yang ditantang WSSV setiap pengamatan disajikan secara deskriptif.
HASIL DAN PEMBAHASAN
Konstruksi gen pProAV-PmAV telah berhasil dibuat dengan ukuran total
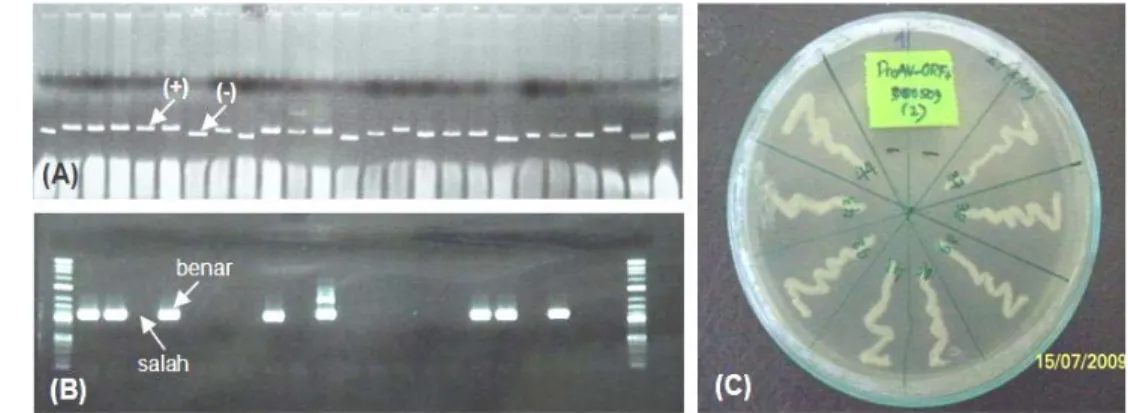
plasmid adalah sekitar 4,4 kb. Uji cracking memperlihatkan adanya insersi promoter ProAV dan gen antivirus PmAV dibandingkan dengan kontrol bakteri koloni biru (Gambar 12A), selanjutnya dengan konfirmasi uji orientasi juga menunjukkan keberhasilan penyisipan gen dengan arah yang benar (Gambar 12B). Arah ligasi yang benar pada konstruksi gen pProAV-PmAV ini didapatkan lebih rendah, yakni 8 klon dari 21 klon bakteri (38,1%) dibandingkan dengan arah ligasi yang benar pada kloning konstruksi gen pProAV-EGFP (86,0%). Rendahnya persentase arah ligasi yang benar dalam konstruksi ini kemungkinan disebabkan karena proses penyambungan (ligasi) gen dilakukan dalam dua tahap, yakni
pertama dengan promoter ProAV dengan menggunakan bantuan situs resriksi
BamHI, kemudian diikuti dengan insersi gen antivirus PmAV dengan bantuan
situs restriksi SalI. Pada pembuatan konstruksi ProAV-EGFP hanya dilakukan satu kali insersi promoter dengan bantuan situs restriksi BamHI (lihat Bab IV). Selain itu, kualitas sel kompeten bakteri yang digunakan mungkin berbeda dan mempengaruhi keberhasilan dalam memperoleh koloni bakteri dengan arah ligasi yang benar. Klon-klon yang positif membawa konstruksi ProAV-PmAV selanjutnya digoreskan pada media agar dalam cawan petri (Gambar 12C) sebagai bahan untuk digunakan pada kegiatan selanjutnya.
Gambar 12 Hasil analisis cracking dan PCR konstruksi gen pProAV-PmAV. A=hasil cracking klon bakteri pembawa promoter dan PmAV pada gel agarosa dimana tanda panah dengan indikator positif menunjukkan klon pembawa gen insersi sedangkan indikator negatif sebagai kontrol bakteri koloni biru, B=hasil uji orientasi ligasi promoter pProAV-PmAV dalam vektor dimana tanda panah menunjukkan perbedaan arah ligasi yang benar dan yang salah, dan C=hasil plating klon bakteri pembawa konstruksi gen pProAV-PmAV.
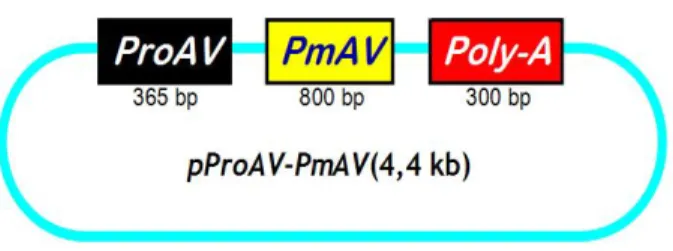
Dengan masuknya promoter ProAV dan gen antivirus PmAV ke dalam vektor pBlueskript-SK, maka didapatkan konstruksi gen dengan komponen secara berurutan adalah pProAV-PmAV. Peta konstruksi gen pProAV-PmAV disajikan pada Gambar 13.
Gambar 13 Peta konstruksi gen pProAV-PmAV yang digunakan dalam transfeksi gen antivirus PmAV ke embrio udang windu P. monodon
Transfeksi konstruksi gen pProAV-PmAV dilakukan dalam dua tahap, dimana konsentrat telur yang digunakan adalah 370 butir/2 mL pada tahap pertama dan 235 butir/2 mL pada tahap kedua. Hasil pengamatan derajat penetasan telur udang windu tidak memperlihatkan perbedaan yang nyata (P>0,05) antara kontrol positif (transfeksi tanpa gen antivirus) dan kontrol negatif (tanpa transfeksi). Rata-rata daya tetas telur yang didapatkan adalah 38,1% pada perlakuan transfeksi gen antivirus, 43,1% pada kontrol positif, dan 49,0% pada kontrol negatif. Hasil tersebut berimplikasi bahwa penggunaan larutan jetPEI dan plasmid DNA dari konstruksi gen tidak memberikan efek yang berbahaya pada daya tetas embrio udang windu. Daya tetas embrio, deteksi DNA, dan cDNA pada udang windu setiap tahap transfeksi disajikan pada Tabel 4. Walaupun metode transfeksi pada udang vaname memperlihatkan metode yang lebih baik dibanding dengan mikroinjeksi dan elektroporasi dilihat dari daya tetas telur yang dihasilkan (Sun et al. 2005), tetapi metode mikroinjeksi pada ikan medaka Oryzias latifes memberikan daya tetas telur yang cukup tinggi yakni 70% dibandingkan dengan daya tetas 78% pada ikan kontrol (Winkler et al. 1991), pada ikan lele Clarias sp. mencapai 55,0-93,3% (Ath-Thar 2007), dan pada ikan sea bream Pagrus major sekitar 53-63% (Kato et al. 2007).
Selain itu, untuk mengetahui masuk atau tidaknya gen eksogenus antivirus PmAV pada larva, dilakukan deteksi melalui ekstraksi DNA genom dan ekspresi gen antivirus PmAV diamati melalui ekstraksi RNA total yang dilanjutkan dengan sintesis cDNA. Secara umum, Tabel 4 memperlihatkan bahwa hasil deteksi DNA dan cDNA yang dilakukan memperlihatkan masuknya gen eksogenous gen antivirus ke dalam embrio atau larva udang windu (positif).
Tabel 4 Daya tetas embrio, deteksi DNA dan cDNA pada embrio dan larva udang windu P. monodon hasil transfeksi konstruksi gen ProAV-PmAV.
Tahap Daya tetas telur (%) Deteksi*) Transfeksi
PmAV
Kontrol
positif Kontrol negatif DNA cDNA 1 2 48,0±14,2a 28,1±6,4a 60,0±3,0a 26,2±6,9a 66,8±12,1a 31,2±14,4a (+) (+) (+) (+)
Keterangan: Angka pada baris yang sama dan diikuti oleh huruf yang sama menunjukkan
tidak berbeda nyata (P>0,05), angka ditulis dalam rataan ± SD, *) = analisis
dilakukan terhadap 50 embrio (pooled sample), (+) = gen PmAV positif terdeteksi pada genom DNA dan cDNA.
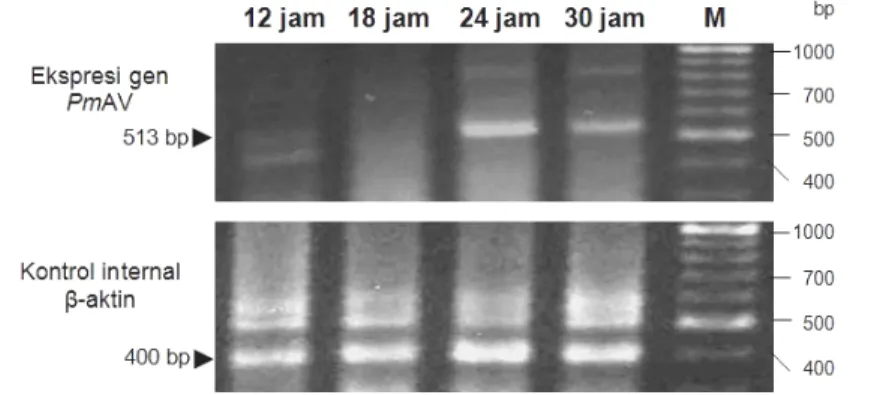
Analisis ekspresi gen antivirus PmAV pada embrio dan larva udang windu
(Gambar 14) memperlihatkan pola yang relatif sama degan pola ekspresi gen
EGFP pada embrio dan larva udang windu (lihat Bab IV). Pada pengamatan 12 jst,
ekspresi gen antivirus PmAV sudah mulai terlihat tetapi masih lemah sampai dengan 18 jst. Ekspresi gen tersebut memuncak pada jam ke-24 atau satu hari setelah transfeksi dan kembali menurun pada 30 jst. Hasil penelitian menunjukkan bahwa ekspresi transgen mulai menurun setelah udang menetas menjadi naupli. Berdasarkan beberapa penelitian, ekspresi gen asing umumnya dimulai setelah fase mid-blastula dan levelnya meningkat selama embriogenesis, dan selanjutnya menurun setelah menetas (Alimuddin et al. 2003).
Gambar 14 Ekspresi gen antivirus PmAV pada embrio dan larva udang windu P.
monodon. Pengamatan ekspresi gen antivirus PmAV pada 12, 18, 24,
dan 30 jst; M= VC 100bp Plus DNA Ladder (Vivantis); tanda kepala panah menunjukkan posisi fragmen gen antivirus PmAV dan gen β-aktin udang windusebagai kontrol internal.
Ekspresi transgen yang dianalisis pada penelitian ini bersifat sementara (transient expression). Ekspresi transgen meningkat diduga terjadi akibat adanya replikasi ekstra kromosomal DNA asing, dan selanjutnya level ekspresinya menurun yang diikuti dengan degradasi dari eksra kromosomal DNA asing. Ekspresi sementara gen hrGFP yang dikontrol oleh promoter β-aktin ikan medaka pada ikan lele Clarias sp. mulai terlihat pada 4 jam setelah mikroinjeksi tetapi masih sangat lemah. Ekspresi tersebut semakin meningkat pada jam 8 dan ke-12 dan setelahnya (jam ke-16, 20, 24) menunjukkan tanda penurunan ekspresi dan akhirnya tidak terdeteksi (Ath-thar 2007). Dengan menggunakan promoter yang sama, ekspresi GFP pada ikan mas Cyprinus carpio memperlihatkan pola yang serupa dengan ikan lele, dimana didapatkan tingkat ekspresi tertinggi pada jam ke-12 sampai jam ke-18 setelah mikroinjeksi dan kemudian setelah menetas ekspresi mulai melemah sampai tidak kelihatan pada larva berumur 1 hari (Purwanti 2007).
Masuknya gen antivirus PmAV ke dalam embrio atau larva udang windu merupakan indikator utama keberhasilan transfer gen. Hasil analisis DNA genom pada penelitian ini menunjukkan bahwa persentase udang windu F0 yang membawa gen eksogenus PmAV adalah 37,5-75,0%. Efisiensi transfer gen asing ke embrio dipengaruhi oleh metode transfer dan spesies yang digunakan. Beberapa peneliti telah melaporkan keberhasilannya dalam mengintroduksi gen asing ke embrio krustase. Laju introduksi gen pada udang kuruma Marsupenaeus
japonicus relatif rendah yakni 1% untuk metode mikroinjeksi dan 0,42% untuk
metode particle-bombardment. Keberhasilan transfer gen asing ke embrio udang putih L. schmitti sekitar 36% dengan metode elektroporasi (Arenal et al. 2008). Efesiensi introduksi gen TSV-CP yang relatif tinggi (72%) telah dilaporkan oleh Sun et al. (2005) pada udang L. vannamei dengan menggunakan metode transfeksi. Selanjutnya, dengan metode elektroporasi, Tseng et al. (2000) telah membuktikan bahwa transfer gen bacterial alkaline phosphatase (BAP) dapat terintegrasi dalam DNA genom udang windu, dimana laju integrasi gen tersebut dapat mencapai 31%. Integrasi gen yang diintroduksi ke ikan mas India Labeo
rohita telah dilaporkan oleh Rajesh & Majumdar (2005) dengan menggunakan
transfer gen asing ke embrio udang tidak lagi merupakan kendala utama dalam pengembangan produksi udang transgenik.
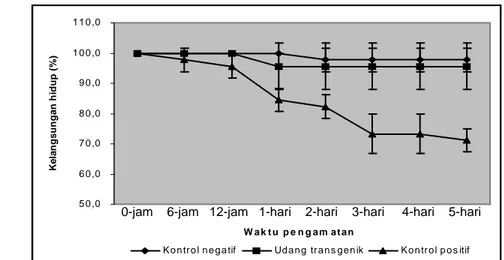
Pada penelitian uji tantang larva udang dengan WSSV diperoleh hasil bahwa respons imunitas udang transgenik F0 lebih tinggi dibandingkan dengan larva udang non-transgenik. Penilaian respons imun udang didasarkan pada kelangsungan hidup larva. Kematian larva udang windu kontrol mulai terlihat pada 12 jam (hari-1) setelah uji tantang sampai dengan hari ke-3 dan setelah itu kematian udang tidak signifikan (Gambar 15).
Gambar 15 Kelangsungan hidup larva udang windu P. monodon yang ditantang dengan WSSV
Kematian udang ditandai dengan perubahan respons pakan yang menurun, aktivitas renang yang tidak stabil, selalu berada di dasar wadah dan munculnya warna tubuh yang kemerahan, serta gejala bintik putih pada karapaks. Gejala perubahan patologis yang serupa juga telah dilaporkan oleh Alifuddin et al. (2003) pada penelitian penularan WSSV pada larva udang windu P. monodon non-transgenik. Selanjutnya dikatakan bahwa karakteristik perubahan seluler akibat infeksi virus WSSV pada udang windu adalah terjadinya pembengkakan inti sel (hipertropi) akibat perkembangan dan penumpukan virion yang berkembang dalam inti sel sehingga bergerak ke pinggir, kemudian terjadi kariolisis yang pada akhirnya sel akan mengalami kerusakan (lisis). Kerusakan sel tersebut yang diduga memacu kematian udang windu.
5 0 ,0 6 0 ,0 7 0 ,0 8 0 ,0 9 0 ,0 1 0 0 ,0 1 1 0 ,0 W a k t u p e n g a m a t a n K e la ngs unga n h idup (% ) K o n tr o l n e g a tif Ud a n g tr a n s g e n ik K o n tr o l p o s itif
Hasil penelitian menunjukkan bahwa kelangsungan hidup larva udang windu yang ditransfeksi dengan gen antivirus lebih tinggi dibandingkan dengan larva udang normal (kontrol) ketika ditantang dengan WSSV (Tabel 5). Analisis ragam menunjukkan bahwa kelangsungan hidup larva transgenik PmAV (95,6%) berbeda nyata (P<0,05) dengan larva udang kontrol (71,1%), tetapi tidak berbeda nyata dengan kontrol negatif (udang normal tanpa uji tantang). Alifuddin et al. (2003) melaporkan bahwa memaparan WSSV pada larva udang windu dapat menyebabkan kematian dengan tingkat kelangsungan hidup rata-rata berkisar 73,3-91,7% dibandingkan dengan larva pada kontrol negatif (100%), dimana semakin tinggi konsentrasi virus yang diberikan semakin rendah kelangsungan hidup larva yang dihasilkan.
Tabel 5 Kelangsungan hidup larva udang windu P. monodon pada perlakuan uji tantang dengan WSSV
Uraian Uji tantang dengan WSSV Tanpa uji tantang WSSV Transgenik Non-Transgenik Ulangan 1 Ulangan 2 Ulangan 3 Rata-rata*) SD 100,0 86,7 100,0 95,6a 7,7 73,3 73,3 66,7 71,1b 3,8 100,0 93,3 100,0 97.8a 3,9
Keterangan: *)Angka yang diikuti oleh huruf yang sama menunjukkan tidak berbeda nyata (P>0,05) dan SD=standar deviasi.
Hasil penelitian menunjukan bahwa kelangsungan hidup larva udang windu hasil transfeksi adalah 24,5% lebih tinggi dari kontrol udang normal yang ditantang dengan WSSV. Hal ini berimplikasi bahwa over-ekspresi gen antivirus
PmAV dapat meningkatkan resistensi udang windu. Seperti halnya dengan udang
windu, peningkatan resistensi udang vaname L. vannamei melalui transfer gen
TSV-CP telah dilaporkan oleh Lu & San (2005). Penelitian yang dilakukan selama
4 hari tersebut mengungkapkan bahwa larva udang vaname transgenik memiliki resistensi yang lebih tinggi dengan kelangsungan hidup mencapai 83%
dibandingkan dengan kontrol udang normal hanya 44% ketika ditantang dengan virus TSV.
Imunitas ikan yang dihasilkan dari teknologi transgenesis pengkode antimikroba memiliki beberapa keuntungan dibandingkan dengan immunisasi konvensional. Ikan dapat diproteksi mulai dari awal perkembangan larva, jauh sebelum sistem imun mulai matang (matured immunity) dan immunitas yang diberikan pada transgenik dapat menghilangkan persiapan khusus untuk melawan setiap jenis patogen (Dunham 2009). Transfer gen pengkode peptida antimikroba seperti cecropin yang diatur oleh promoter CMV dapat meningkatkan resistensi terhadap penyakit bakteri pada ikan channel catfish Ictalurus punctatus sampai dengan 2-4 kali lipat (Dunham et al. 2002). Ikan catfish transgenik yang membawa konstruksi preprocecropin dapat memperlihatkan kelangsungan hidup yang sangat tinggi (100%) ketika dipapar dengan bakteri Plavobacterium
columnare dibandingkan dengan kontrol ikan yang hanya 27,3%. Sementara itu,
ketika ditantang dengan bakteri Edwardsiella ictaluri, ikan catfish transgenik juga memperlihatkan kelangsungan hidup yang tinggi (40,7%) dibandingkan dengan kontrol ikan non trangenik (14,8%). Transfer gen cecropin ke ikan medaka juga telah memperlihatkan pengaruh peningkatan resistensi terhadap patogen. Sarmasik et al. (2002) melaporkan bahwa ikan transgenik medaka generasi F2 memperlihatkan resistensi yang tinggi terhadap bakteri Pseudomonas fluorescens dengan tingkat kematian 0-10% dibandingkan dengan kontrol 40%, dan ketika ditantang dengan Vibrio anguillarum, ikan medaka transgenik masih mampu bertahan dengan kelangsungan hidup 70-90% sedangkan kontrol medaka hanya berkisar 60%.
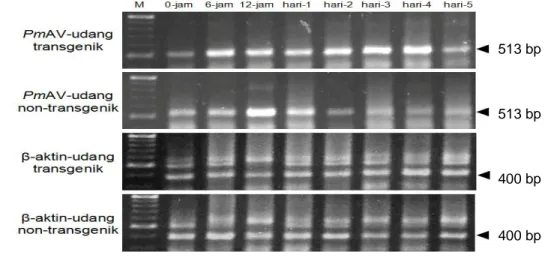
Setelah ditantang dengan WSSV, udang windu transgenik memperlihatkan respons induksi yang meningkat (up-regulated) baik pada larva udang transgenik maupun larva non-transgenik (Gambar 16). Ekspresi gen antivirus PmAV mulai terinduksi sejak 6 jam setelah pemaparan dan selanjunya meningkat dengan tajam sampai dengan hari ke-4 dan sedikit menurun pada hari ke-5 setelah uji tantang. Seperti halnya dengan udang transgenik, udang kontrol non-transgenik juga memperlihatkan induksi yang meningkat dimana terjadi peningkatan sampai mencapai puncak pada hari ke-1 dan selanjutnya menurun pada hari ke-2 dan
seterusnya memperlihatkan ekspresi gen antivirus PmAV yang relatif lebih rendah. Luo et al. (2007) telah melaporkan ekspresi gen antivirus PmAV secara alami pada udang windu non-transgenik melalui uji tantang dengan WSSV. Ekspresi terbesar gen antivirus tersebut didapatkan pada hepatopankreas udang windu, 700 kali lebih tinggi dari ekspresi PmAV pada otot. Pola ekspresi PmAV yang didapatkan pada hepatopankreas relatif sama dengan hasil penelitian ini, dimana pada awal pemaparan pada hari ke-1 ekspresi menurun dan selanjutnya meningkat tajam sampai dengan hari ke-4, tetapi menurun kembali pada hari ke-5. Selanjutnya Luo
et al. (2007) menyatakan bahwa pola ekspresi gen antivirus PmAV sangat relevan
dengan muatan virus WSSV dalam tubuh udang windu. Peningkatan ekspresi gen
PmAV yang tinggi pada hari ke-4 dari penelitian ini mendukung penelitian
sebelumnya pada gen C-type lectin dari udang vaname. Ma et al. (2007) melaporkan bahwa udang vaname yang ditantang dengan WSSV memperlihatkan ekspresi gen C-type lectin yang awalnya menurun pada hari ke-2 dan setelah itu meningkat tajam sampai dengan mencapai puncak pada hari ke-4.
Gambar 16 Ekspresi gen antivirus PmAV pada hepatopankreas larva udang windu
P. monodon yang ditantang dengan WSSV. Tanda kepala panah atas
dan tengah menunjukkan posisi fragmen DNA target gen PmAV dan β-aktin sebagai kontrol internal, M= VC 100bp Plus DNA Ladder (Vivantis), pengamatan ekspresi gen PmAV pada 0 jam (1) 6 jam (2), 12 jam (3), 24 jam (4), 2 hari (5), 3 hari (6), 4 hari (7), dan 5 hari (8) setelah uji tantang.
513 bp
513 bp
400 bp
Hasil pengamatan pola ekspresi gen antivirus PmAV dalam penelitian ini menunjukkan bahwa peningkatan ekspresi gen setelah ditantang dengan patogen mengindikasikan akan keterlibatan gen tersebut dalam proses perlawanan tubuhnya atau dikenal sebagai respons resistensi terhadap patogen. Somboonwiwat et al. (2006) telah melaporkan bahwa peningkatan level ekspresi gen dalam hemosit udang windu P. monodon yang telah dipapar dengan mikroba menunjukkan bahwa gen tersebut terlibat dalam respons mikroba misalnya
glucosa transporter-1, interferon-related developmental regulator-1, lisozim,
profilin, dan serpin-B3. Hal ini berarti bahwa gen-gen tersebut mengalami induksi meningkat (up-regulated gene). Beberapa gen pada krustase yang sudah diketahui mengalami induksi meningkat ketika dipapar dengan patogen. Gen antibakteria penaeidin memperlihatkan ekspresi yang kuat pada udang vaname L. vannamei (Destoumieux et al. 2000b) dan pada udang Fenneropenaues chinensis (Kang et
al. 2007), ketika ditantang dengan patogen. Demikian pula ekspresi gen Rab GTPase pada udang P. japonicus terinduksi ketika udang ditantang dengan virus WSSV (Wu & Zhang 2007), dan gen lisozim pada udang vaname L. vannamei
ketika diinjeksi dengan Vibrio campellii (Burge et al. 2007).
Untuk mengetahui pengaruh introduksi gen antivirus PmAV dalam penampilan morfologi atau pertumbuhan, maka larva udang dipelihara dalam kondisi terkontrol. Pengamatan yang dilakukan pada saat larva berumur 47 hari menunjukkan bahwa larva udang transgenik tidak memiliki perbedaan penampilan secara morfologi (Gambar 17). Larva udang transgenik mencapai ukuran berat 0,21±0,12 g dengan panjang 3,3±0,51 cm, sedangkan larva udang kontrol memiliki berat 0,30±0,16 g dan panjang 3,5±0,63 cm. Berdasarkan uji-t yang dilakukan antara kedua kelompok udang tersebut menunjukkan tidak adanya perbedaan yang nyata (P>0,05) baik berat maupun panjang udang windu. Selanjutnya, berdasarkan pengamatan dalam jangka waktu panjang, udang transgenik terlihat relatif aktif, sehat dengan morfologi yang normal selama berlangsungnya pemeliharaan.
Gambar 17 Penampilan secara morfologi udang windu P. monodon transgenik (A) dan non-transgenik (B) pada umur 47 hari.
Hal yang serupa dilaporkan oleh Lu & Sun (2005) pada udang vaname transgenik. Pengamatan yang dilakukan sampai dengan pemeliharaan 236 hari menunjukkan bahwa udang transgenik dan non-transgenik tidak memperlihatkan perbedaan penampilan secara morfologi. Berat rata-rata udang vaname transgenik pada umur tersebut adalah 7,67 g, tidak berbeda nyata secara statistik dibandingkan dengan udang non-transgenik (kontrol) yakni 9,17 g. Selanjutnya Lu & Sun (2005) menyatakan bahwa penurunan pertambahan berat kemungkinan disebabkan oleh integrasi gen target yang diintroduksi pada daerah spesifik pada genom akan memberikan sedikit pengaruh ke pertumbuhan awal larva udang vaname.
Penelitian ini berimplikasi bahwa analisis ekspresi gen antivirus PmAV pada embrio dan larva merupakan salah satu bukti keberhasilan transfer gen antivirus PmAV pada udang windu. Sementara itu, pengamatan kelangsungan hidup dan ekspresi gen antivirus pada udang windu ketika ditantang dengan WSSV memberikan gambaran umum mengenai keterlibatan gen antivirus PmAV dalam sistem imunitas udang windu sehingga respons gen antivirus tersebut dapat diketahui.
KESIMPULAN
Konstruksi gen antivirus pProAV-PmAV telah berhasil dibuat. Ekspresi
transgen antivirus PmAV mampu meningkatkan resistensi terhadap infeksi WSSV dan tidak mempengaruhi pertumbuhan udang windu.