ANALISIS EKSPRESI GEN PENYANDI METALLOTHIONEIN TIPE II PADA Melastoma affine L. YANG MENDAPAT CEKAMAN pH RENDAH
DAN ALUMINIUM
Niken Trisnaningrum P 055050081
PROGRAM STUDI BIOTEKNOLOGI SEKOLAH PASCA SARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2009
Dengan ini saya menyatakan bahwa tesis yang berjudul Analisis Ekspresi Gen Penyandi Metallothionein Tipe II pada Melastoma affine L. yang Mendapat Cekaman pH Rendah dan Aluminium adalah karya saya bersama dengan komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Juni 2009
Niken Trisnaningrum NRP P055050081
ABSTRACT
NIKEN TRISNANINGRUM. Expression of the gene encoding metallothionein type II in Melastoma affine L. under low pH and aluminum stress. Supervised by SUHARSONO and MUHAMAD YUNUS
Aluminum is toxic to plants, affects plant development and reduces crop production. Indonesia has 47.500.000 ha of acid soil with very low pH and high content of aluminum. The application of tolerant crop varieties is the best approach to overcome this problem and the availability of genes controlling tolerance to this abiotic stress is essential to develop new tolerant varieties. Melastoma affine is the indigenous plant in tropical rain forest, very tolerant to acid condition and aluminum. Therefore this plant could be used for source of genes for acid and aluminum tolerance. Metallothionein is one of genes responsible for aluminum tolerance. MaMt2 gene encoding metallothionein from M. affine was isolated. The expression of this gene in M. affine under low pH and aluminum stresses was investigated using Real Time PCR in this research. MaMt2 primers were designed based on the sequence of MaMt2. β-actin was used as internal control, and the primers for β-actin were designed based on cDNA of the region between exon 1 and exon 2 of this gene. Total RNA was successfully isolated from roots and young leaves separately. Total cDNA was synthesized from total RNA by reverse transcription. This cDNA was used as template for Real Time PCR to analyse the expression of MaMt2. The expressions of MaMt2 in leaves and roots were not affected by low pH. At high concentration (3.2 mM) aluminum induced the expression of MaMt2 in the roots but not in leaves.
.
Tipe II pada Melastoma affine L. yang Mendapat Cekaman pH Rendah dan Aluminium. Dibimbing oleh SUHARSONO dan MUHAMAD YUNUS
Indonesia yang mempunyai tanah ultisol sebanyak 47,5 juta ha yang memiliki karakteristik pH rendah dan kandungan Al yang tinggi. Salah satu cara untuk mengatasi kandungan Al yang sangat tinggi tersebut adalah dengan melakukan perakitan tanaman yang memiliki ketahanan terhadap keracunan Al. Melastoma affine merupakan tanaman yang toleran terhadap pH rendah dan cekaman Al yang sangat tinggi, sehingga tanaman ini dapat digunakan sebagai sumber gen toleransi terhadap pH rendah dan Al. Salah satu gen yang diduga berperan dalam ketahanan M. affine terhadap pH rendah dan Al adalah gen penyandi metallothionein (MaMt2).
Penelitian ini bertujuan untuk menganalisis ekspresi gen penyandi MT2 pada Melastoma affine L. yang mendapat cekaman pH rendah dan Al. Untuk itu, M. affine diperbanyak dengan stek pucuk dan diberi cekaman pH rendah dan cekaman Al secara terpisah pada kultur cair selama satu minggu. Penelitian ini dilakukan dengan dua ulangan. Cekaman pH rendah diberikan pada dua tingkat pH yaitu 6 dan 4. Cekaman Al dilakukan pada pH 4 dengan 3 tingkat konsentrasi Al yaitu 0, 0,8, dan 3,2 mM. Satu minggu setelah perlakuan, RNA total diisolasi dari daun dan akar secara terpisah, lalu digunakan untuk sintesis cDNA total. Analisis ekspresi MaMt2 dilakukan dengan menggunakan cDNA total dengan Real Time PCR dan pengamatan semikuantitatif pada gel agarose. Analisis ekspresi MaMt2 dilakukan dengan membandingkan ekspresi gen MaMt2 yang telah dinormalisasi dengan gen β aktin antara kontrol (pH 6 dan 0 mM Al) dengan perlakuan yang dilakukan dengan metode comparative Ct (∆∆Ct). Analisis dengan menggunakan Real Time PCR dilakukan sebanyak dua ulangan, selanjutnya analisis statistik dilakukan dengan program SPSS 13 untuk membandingan ekspresi perlakuan. Hasil Real Time PCR di cek dengan menggunakan gel agarose untuk mendapatkan hasil ekspresi MaMt2 semikuantitatif.
RNA total yang berkualitas baik telah berhasil diisolasi dari daun dan akar M. affine. Selanjutnya, cDNA yang bebas dari kontaminasi DNA genom, disintesis dari RNA total. Berdasarkan hasil uji statistik terhadap ekspresi MaMt2, cekaman pH rendah tidak mempengaruhi ekspresi MaMt2 pada daun dan akar M. affine, begitu juga dengan cekaman Al terhadap ekspresi MaMt2 di daun. Sedangkan cekaman Al pada akar meningkatkan ekspresi MaMt2 sebanyak 2,1 kali pada konsentrasi Al 3,2 mM. Hal ini terjadi karena metallothionein merupakan protein yang terutama dibutuhkan untuk homeostasis logam dan detoksifikasi logam berat, sehingga ketiadaan logam berlebih pada media dengan pH rendah tidak menginduksi MaMt2. Akar merupakan bagian tanaman yang berinteraksi langsung dengan cekaman Al sehingga kemungkinan respon gen tersebut terjadi di akar dari pada di daun. Selain itu M. affine merupakan tanaman yang sangat toleran terhadap Al, sehingga hanya pada konsentrasi Al yang tinggi, Al menginduksi ekspresi MaMt2.
Judul Penelitian : Analisis Ekspresi Gen Penyandi Metallothionein Tipe II pada Melastoma affine L. yang Mendapat Cekaman pH Rendah dan Aluminium
Nama Mahasiswa : Niken Trisnaningrum
NRP : P055050081
Disetujui
Komisi Pembimbing
Dr. Ir. Suharsono, DEA Ketua
Ir. Muhamad Yunus, MSi, PhD Anggota
Diketahui
Ketua Program Studi Bioteknologi
Dr. Ir. Muhammad Jusuf
Dekan Sekolah Pascasarjana
Prof. Dr. Ir. Khairil A. Notodiputro, MS
Puji syukur penulis panjatkan kepada Allah SWT yang telah melimpahkan karunia-Nya, sehingga penulis dapat menyelesaikan tesis yang berjudul : Analisis Ekspresi Gen Penyandi Metallothionein Tipe II pada Melastoma affine L. yang Mendapat Cekaman pH Rendah dan Aluminium. Penelitian ini dilaksanakan di Laboratorium BIORIN, PAU, IPB dan Laboratorium Terpadu, BB Biogen.
Ucapan terimakasih penulis sampaikan kepada :
1. Dr. Suharsono dan Dr. Muhamad Yunus selaku Komisi Pembimbing, atas bimbingan, saran dan masukan selama penelitian dan penulisan tesis ini. 2. Proyek Penelitian Kerjasama Kemitraan Penelitian Pertanian dengan
Perguruan Tinggi (KKP3T) yang telah membiayai penelitian ini melalui SPK No. 748/LB.620/I.1/3/2008 atas nama Dr. Suharsono.
3. Bapak Adi selaku teknisi rumah kaca, Bapak Mulya selaku staf Laboratorium Biologi Molekuler dan Bioteknologi IPB, dan Ibu Pepy selaku staf Laboratorium BIORIN atas bantuan yang diberikan selama pelaksanaan penelitian.
4. Ayah dan Ibu atas dukungan finansial dan spiritual.
5. Teman-teman Laboratorium Biorin atas masukan dan diskusi yang sering diberikan.
Penulis menyadari bahwa tesis ini masih ada kekurangan. Namun demikian penulis berharap semoga penelitian ini dapat berguna bagi pihak-pihak yang memerlukannya.
Bogor, Juni 2009
RIWAYAT HIDUP
Penulis dilahirkan di Bondowoso pada tanggal 31 Agustus 1981 dari pasangan Bapak Sutrisno dan Ibu Sri Mastutik. Penulis merupakan anak kedua dari tiga bersaudara.
Tahun 1999 penulis lulus dari SMU Negeri 2 Bondowoso dan lulus seleksi masuk Universitas Brawijaya melalui jalur UMPTN di Program Studi Pemuliaan Tanaman, Fakultas Pertanian dan selesai pada tahun 2005. Pada tahun yang sama penulis diterima di Program Studi Bioteknologi Sekolah Pascasarjana IPB.
DAFTAR TABEL ... viii DAFTAR GAMBAR ... ix DAFTAR LAMPIRAN... x I. PENDAHULUAN... 1 1.1Latar Belakang ... 1 1.2Tujuan Penelitian... 3 1.3Hipotesis Penelitian ... 3
II. TINJAUAN PUSTAKA ... 4
2.1Mekanisme Toleransi Terhadap Aluminium (Al) ... 4
2.2Metallothionein... 6
2.3Melastoma affine L ... 7
2.4Real Time Polimerase Chain Reaction (Real Time PCR) ... 8
III. BAHAN DAN METODE ... 16
3.1Waktu dan Tempat... 16
3.2Bahan Penelitian ... 16
3.3Metode ... 16
3.3.1Perlakuan cekaman pada kultur air ... 17
3.3.2Isolasi RNA total dari Melastoma affine L ... 17
3.3.3Pembentukan cDNA total... 18
3.3.4Analisis ekspresi gen dengan menggunakan Real Time PCR ... 19
IV. HASIL DAN PEMBAHASAN... 21
4.1Isolasi RNA Total ... 21
4.2Sintesis cDNA Total ... 21
4.3Ekspresi Gen Metallothionein ... 22
4.3.1Pengaruh pH rendah terhadap ekspresi gen metallothionein ... 22
4.3.2Pengaruh aluminium terhadap ekspresi gen metallothionein ... 24
V. SIMPULAN DAN SARAN ... 26
5.1Simpulan ... 26
5.2Saran ... 26
DAFTAR PUSTAKA LAMPIRAN ... 32
DAFTAR TABEL
Halaman 1. Nilai relative quantification (RQ) dari transkripsi MaMT2
pada daun dan akar M. affine L.yang mendapat cekaman pH rendah.. 24
2. Nilai relative quantification (RQ) dari transkripsi MaMT2
pada daun dan akar M. affine L. yang mendapat perlakuan Al pada medium dengan pH 4... 25
Halaman
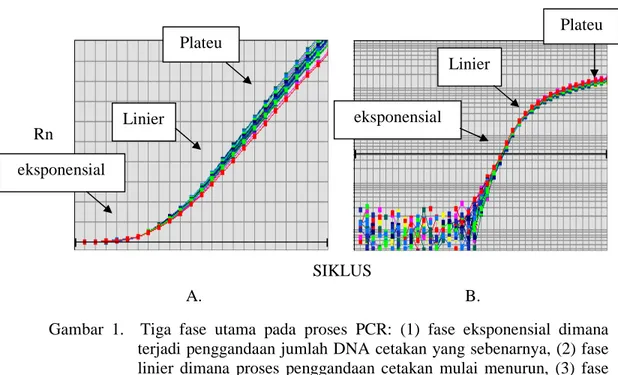
1. Tiga fase utama pada proses PCR: (1) fase eksponensial dimana terjadi penggandaan jumlah DNA cetakan yang sebenarnya, (2) fase linier dimana proses penggandaan cetakan mulai menurun, (3) fase plateu dimana tidak terjadi lagi
proses penggandaan cetakan dan DNA mulai terdegradasi. Deteksi untuk Real Time PCR terjadi pada fase eksponensial, sedangkan deteksi pada PCR biasa terjadi pada fase plateu yang digambarkan dalam bentuk kurva linier (A) dan kurva log (B)
(Applied Biosystem 2007)... 10
2. Proses deteksi pada Real Time PCR dengan menggunakan SYBR Green. SYBRGreen akan terdeteksi bila terikat pada utas ganda DNA saat fase perpanjangan DNA (extention)
(Dorak 2006)... 11
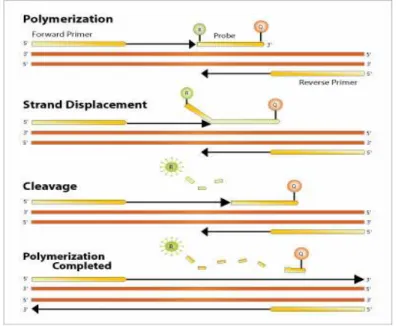
3. Proses deteksi pada Real Time PCR dengan menggunakan
TaqMan. Deteksi akan terjadi bila TaqMan probe yang menempel diantara primer forward dan primer reverse rusak selama fase
perpanjangan DNA (extention) (Dorak 2006) ... 11
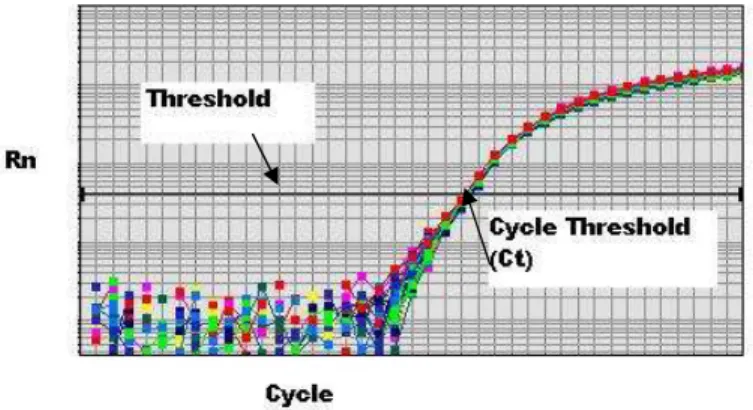
4. Kurva amplifikasi pada Real Time PCR. Threshold adalah daerah deteksi yang digunakan dalam analisis. Cycle Threshold (Ct) adalah siklus pada saat fluorescence reporter berpotongan dengan threshold. Rn adalah jumlah fluorescence reporter yang terdeteksi selama proses PCR. Cycle adalah banyaknya siklus yang digunakan dalam proses PCR (Applied Biosystem 2007)... 12
5. RNA total M. affine L. hasil isolasi dari akar (1) dan daun (2) ... 21
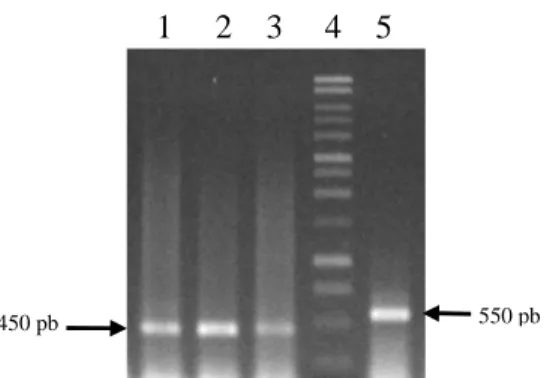
6. Hasil amplifikasi β-actin. Lajur 1,2, dan 3 = hasil amplifikasi dengan menggunakan cDNA sebagai cetakan. Lajur 4 = 1 kb ladder. Lajur 5 = hasil amplifikasi dengan menggunakan DNA genom
sebagai cetakan... 22
7. Ekspresi MaMt2 semikuantitatif pada daun dan akar dari M. affine L. yang mendapat cekaman pH rendah... 23
8. Ekspresi MaMt2 semikuantitatif pada daun dan akar dari M. affine L. yang mendapat cekaman Al ... 24
DAFTAR LAMPIRAN
Halaman 1. Tabel ANOVA dari pengaruh pH terhadap ekspresi
MaMt2 di daun dan akar M. affine L. ... 32
2. Nilai ekspresi gen MaMt2 terhadap β-actin dari M. affine L. yang mendapat cekaman pH rendah pada daun dan akar ... 33
3. Tabel uji Tukey dari pengaruh konsentrasi aluminium terhadap
I. PENDAHULUAN
1.1 Latar Belakang
Salah satu usaha untuk meningkatkan produksi pertanian di Indonesia adalah dengan cara ekstensifikasi pertanian, diantaranya dengan pemanfaatan tanah ultisol yang merupakan salah satu jenis tanah di Indonesia yang mempunyai sebaran luas yaitu mencapai 47,5 juta ha (Subagyo et al. 2004), terutama yang berasal dari bahan dasar sedimen seperti pada tipe hapludults (Pusat Penelitian Tanah dan Agroklimat 2000). Tipe tersebut memiliki pH tanah yang sangat rendah berkisar pada pH 4 dan memiliki kandungan aluminium tinggi (Prasetyo & Suriadikarta 2006). Bentuk Al3+ banyak ditemui pada tanah dengan pH berkisar 4 dimana konsentrasi Al dalam larutan air tanah dapat melebihi 100 µM (Okada & Wissuwa 2004). Keberadaan Al dalam tanah, terutama dalam bentuk Al3+ akan sangat berbahaya dan merupakan racun bagi tanaman, karena akan mengganggu perpanjangan akar dan menghalangi daya tukar kation lain misalnya Ca2+, Mg2+, dan K+.
Salah satu cara untuk mengatasi kandungan Al yang sangat tinggi tersebut adalah dengan melakukan perakitan tanaman yang memiliki ketahanan terhadap keracunan Al. Untuk itu, pencarian sumber daya genetik untuk gen-gen yang menyebabkan tanaman toleran terhadap Al perlu dilakukan. Tanaman yang dapat menjadi sumber gen toleransi terhadap keracunan Al antara lain berasal dari famili Melastomataceae yang dapat ditemui terutama di daerah tropis. Menurut Jansen et al. (2002), Melastomataceae merupakan tanaman akumulator unsur logam termasuk Al. Sebagai contoh Melastoma malabraticum yang mampu mengakumulasi lebih dari 10000 mg Al/kg daun tua dan lebih dari 7000 mg Al/kg daun muda tanpa mengalami keracunan (Watanabe et al. 2003). Analisis akumulasi Al pada Melastoma affine L. yang mendapat cekaman 3,2 mM Al pada pH 4 dalam media cair menunjukkan bahwa M. affine L. mampu mengakumulasi 8,81 mg Al/g daun tua setelah 2 bulan (Mutiasari 2008). Hal ini menunjukkan bahwa tanaman ini memiliki mekanisme toleransi terhadap Al sehingga dapat menjadi salah satu sumber gen ketahanan terhadap cekaman pH rendah dan Al tinggi.
2
Menurut Hall (2002), terdapat beberapa mekanisme seluler untuk toleransi terhadap logam yang berbahaya, dan salah satunya dengan memproduksi metallothionein. Metallothionein (MT) merupakan protein yang banyak mengandung sistein dan berfungsi sebagai peptida pengikat logam (metal binding peptides). Cobbett dan Goldbrough (2002) membagi MT menjadi empat macam berdasarkan pada susunan residu sistein (cys) yang dimiliki, yaitu MT tipe 1, MT tipe 2, MT tipe 3 dan MT tipe 4. MT tipe 1 (MT1) tersusun atas enam motif Cys-Xaa-Cys (Xaa menunjukkan asam amino selain cystein) yang terdistribusi pada dua wilayah secara seimbang. MT tipe 2 memiliki dua wilayah yang kaya Cys, dengan motif susunan cystein berupa Cys-Cys pada asam amino ke tiga dan keempat dari protein ini. Selain itu MT tipe 2 (MT2) memiliki motif Cys-Gly-Gly-Cys pada akhir N-terminal dari wilayah cystein-rich dan pada wilayah C-terminal terdapat 3 motif Cys-Xaa-Cys. MT tipe 3 (MT3) hanya terdiri dari empat residu Cys yang ada pada wilayah N-terminal. Tiga residu pertama membentuk motif Cys-Gly-Asn-Cys-Asp-Cys. Sedangkan residu yang keempat membentuk motif tersendiri yaitu Gln-Cys-Xaa-Lys-Lys-Gly. MT tipe 4 (MT4) berbeda dari kelompok MT tanaman yang lainnya karena memiliki tiga wilayah cystein-rich yang masing-masing memiliki 5 sampai 6 residu cystein yang biasanya membentuk motif Cys-Xaa-Cys.
Menurut Duncan et al. (2006), motif dan susunan residu cys yang dimiliki oleh MT diduga berkaitan dengan kemampuan MT mengikat logam dan kestabilan protein. Pada hewan, logam akan terikat pada residu cys dalam formasi logam-cys-(thiolate) dengan cara pertukaran ion logam misalnya ion Zn tergantikan dengan ion Cu, Cd, atau Hg (Duncan et al. 2006) dan bisa juga terjadi pengikatan logam yang berbeda pada saat bersamaan seperti yang terjadi pada alga coklat (Fucus vesiculosus) yang mampu mengikat Cu dan Cd sekaligus (Zimeri et al. 2005). Hasil penelitian Zimeri et al (2005), menunjukkan bahwa pada arabidopsis, pemberian Cd akan meningkatkan stabilitas MT1. Namun mekanisme pertukaran ion logam dan pengikatan ion logam tersebut masih belum diketahui secara pasti.
Selama ini, penelitian terhadap jenis logam yang mampu menginduksi ekspresi MT masih sangat terbatas, yaitu hanya pada tembaga (Cu), seng (Zn),
kadmium (Cd), dan aluminium (Al) (Robinson et al. 1993; Hall 2001; Snowden & Gardner, 1993; Snowden et al, 1995). Pada tanaman Silene vulgaris (Hoof et al. 2001) dan arabidopsis (Murphy & Taiz, 1995), ekspresi MT2 terinduksi oleh Cu dan keberadaan gen penyandi MT2 membuat mutan kapang yang sensitif terhadap Cu mampu tumbuh dengan penambahan Cu pada medianya (Hoof et al, 2001). Selain terinduksi oleh logam, ekspresi gen MT terinduksi oleh cekaman oksidatif pada jaringan gabus (Mir et al, 2004), terekspresi saat fase senesen pada daun (Buchanan-Wallaston, 1994), dan saat pematangan buah (Davies & Robinson, 2000).
Beberapa gen yang ekspresinya diinduksi oleh Al telah berhasil diisolasi dan dikarakterisasi dari gandum, diantaranya adalah gen penyandi MT (Snowden & Gardner, 1993; Snowden et al, 1995). Suharsono et al (2009) telah mengisolasi cDNA gen penyandi MT2 dari Melastoma affine L (MaMt2). MaMt2 diduga berperan dalam toleransi M. affine terhadap cekaman Al. Untuk mengetahui peranan MaMt2 terhadap Al, maka analisis ekspresi MaMt2 di M. affine L. yang mendapat cekaman Al sangat penting dilakukan.
1.2 Tujuan Penelitian
Tujuan dari penelitian ini adalah untuk menganalisis ekspresi gen penyandi MT2 dari Melastoma affine L. yang mendapat cekaman pH rendah dan Al.
1.3 Hipotesis Penelitian
- Ekspresi gen penyandi MT2 pada daun dan akar Melastoma affine L. akan meningkat bila mendapat cekaman pH rendah.
- Ekspresi gen penyandi MT2 pada daun dan akar Melastoma affine L. akan meningkat bila mendapat cekaman Al.
4
II. TINJAUAN PUSTAKA
2.1 Mekanisme Toleransi Terhadap Aluminium (Al)
Pada prinsipnya ada dua mekanisme toleransi tanaman terhadap cekaman Al. Menurut Taylor (1991), mekanisme pertama adalah mekanisme eksklusi dengan cara mencegah Al masuk kedalam simplas dan mencapai daerah metabolik yang peka. Mekanisme kedua adalah mekanisme internal dengan imobilisasi, kompartementasi dan detoksifikasi saat Al masuk kedalam simplas.
Mekanisme eksklusi dapat berupa: (1) immobilisasi Al pada dinding sel: dinding sel tanaman memiliki sifat yang berbeda dalam menghalangi pergerakan Al. Sebagian besar Al yang diserap akar masuk melalui apoplas dengan menggantikan kedudukan Ca2+ dalam sel tanaman. Immobilisasi Al pada dinding sel akan mengurangi pergerakan Al ke dalam simplas; (2) Permeabilitas selektif membran plasma terhadap Al: membran plasma bertindak sebagai penghalang terhadap masuknya Al ke sitosol; (3) Meningkatkan pH di sekitar perakaran: tanaman yang toleran terhadap Al cenderung menaikkan pH lebih cepat dibandingkan tanaman yang peka sehingga dapat membentuk Al menjadi bentuk yang tidak beracun; (4) Eksudasi senyawa-senyawa pengkelat: Al dapat dikeluarkan dari simplas jika ligan pengkelat dibebaskan ke rizosfir dan ligan tersebut membentuk kompleks yang stabil dengan Al, sehingga mengurangi aktifitas ion Al bebas pada daerah akar-tanah serta mengurangi absorpsi melalui membran plasma. Contoh ligan pengkelat adalah asam organik seperti asam oksalat, asam sitrat, asam suksinat, dan asam malat; dan (5) Eksudasi fosfat: fosfat yang secara aktif dikeluarkan dari akar tanaman dapat membentuk kompleks tak larut dengan Al, sehingga mengurangi pengaruh Al pada permukaan menbran dan/atau penyerapan melalui membran plasma.
Mekanisme internal mencakup: (1) kelatisasi Al di sitosol: meningkatnya produksi asam organik pada tanaman yang toleran Al dapat berperan dalam mengurangi toksisitas Al dalam sitosol; (2) kompartemensasi Al di vakuola yakni Al diasingkan pada tempat yang tidak peka terhadap Al; (3) Protein pengikat Al: banyak jenis tanaman memiliki protein pengikat logam yang pembentukannya dapat terinduksi oleh logam-logam terutama Cd, Zn, Cu, dan Pb yang dikenal
dengan nama fitokelatin; dan (4) Induksi sintesis protein tertentu: pada tanaman gandum kultivar PT741 (toleran Al) Basu et al. (1994) menemukan protein sebesar 51 kD yang diisolasi dari membran mikrosomal pada akar. Protein ini disintesis pada ujung akar karena terinduksi cekaman Al 100 µM sedangkan pada kultivar Neepawa (peka terhadap Al) tidak terbentuk protein tersebut.
Beberapa spesies tanaman memiliki kemampuan untuk mengakumulasi Al di dalam tunas daun dan akar. Tanaman yang memiliki kemampuan ini antara lain teh (Carr et al. 2003; Ruan & Wong 2004), melastoma (Watanabe & Osaki 2002), dan buckwheat (Ma et al. 1998 ; Zheng et al. 1998). Tanaman disebut sebagai akumulator logam bila mampu mengakumulasi logam sedikitnya 0,1 % pada jaringan (Barcel & Poschnenrider 2003).
Mekanisme tanaman dalam mengakumulasi Al masih belum diketahui secara pasti. Namun beberapa penelitian menunjukkan bahwa Al yang masuk ke dalam tanaman terjerap oleh asam organik yang dihasilkan oleh tanaman. Pada teh diduga bahwa Al terikat pada senyawa chatecin (Vitorello et al. 2005), sedangkan pada melastoma Al terikat pada oksalat dan malat dalam daun (Watanabe & Osaki 2002).
Ekspresi beberapa gen pada gandum terinduksi oleh Al, dan salah satu diantaranya adalah gen penyandi metallothionein (Snowden et al. 1995). Metallothionein merupakan protein yang diproduksi oleh tanaman untuk mengikat logam (Cobbett & Goldsbrough 2002). Gen penyandi metallothionein juga berhasil diisolasi dan dikarakterisasi dari Melastoma affine L. yang merupakan salah satu tanaman akumulator Al (Suharsono et al. 2009).
Snowden dan Gardner (1993) menyatakan bahwa Al meningkatkan ekspresi gen MT pada gandum. Al merupakan kation yang sangat reaktif dan termasuk ke dalam golongan hard cation bedasarkan klasifikasi dari Niebour dan Richardson (1980). Oleh karena itu, Al merupakan pengikat yang kuat untuk kelompok hard negative donor, misalnya kelompok karboksil (-COOH), hidroksil (-OH), karbonil (-CO), dan fosfat (-PO3). Sedangkan gugus sulfidril tidak dapat mengikat Al dengan kuat, walaupun berada pada pengkelat seperti misalnya pada metallothionein dan pada phytochelatin (Vitorello et al 2005). Oleh karena itu, induksi gen MT2 oleh cekaman Al diduga berkaitan dengan interferensi Al
6
terhadap ion logam lain yang berpengaruh terhadap metabolisme tanaman secara normal, sehingga MT2 dibutuhkan untuk menjaga homeostasis ion logam tersebut. Ion logam esensial yang dibutuhkan oleh tanaman dan berikatan dengan MT2 adalah Cu dan Zn (Snowden & Gardner 1993).
2.2 Metallothionein
Tanaman memiliki mekanisme untuk mengendalikan dan merespon masuknya ion logam yang berbahaya ke dalam selnya. Salah satu mekanisme internal yang dimiliki oleh tanaman yaitu dengan mengikat ion logam tersebut pada protein atau senyawa organik tertentu dan mengakumulasikannya pada bagian tanaman tertentu misalnya daun. Tanaman tingkat tinggi memiliki dua kelompok besar peptida pengikat logam, yaitu metallothionein (MT) dan phytochelatin (PC) (Cobbett & Goldsbrough 2002).
MT merupakan polipeptida yang memiliki banyak ikatan sistein (cys) yang disandikan oleh gen, memiliki berat molekul yang rendah, dan merupakan protein pengikat logam. Sebagai konsekuensi dari banyaknya kandungan asam amino Cys maka protein ini mengandung kelompok ’thiol’ (sulfidril, -SH) dalam jumlah yang besar. Kelompok ’thiol’ mengikat logam-logam berat dengan sangat kuat dan efisien, termasuk zink, merkuri, tembaga, dan kadmium. Residu sulfidril dari Cys mampu mengikat logam dimana satu ion logam diikat oleh tiga residu –SH atau satu ion logam dengan 2 residu –SH. Koordinasi pengikatan dari setiap ion logam melalui residu –SH yang ada pada Cys, membentuk struktur tetrahedral tetrathiolate (Schultze et al. 1988; Robbins et al. 1991). Pada hewan, logam akan terikat pada residu cys dalam formasi logam-cys-(thiolate) dengan cara pertukaran ion logam misalnya ion Zn tergantikan dengan ion Cu, Cd, atau Hg (Duncan et al. 2006). Menurut Zangger et al. (2001), MT mengikat logam dengan sangat kuat namun pertukaran ikatan logam dapat berlangsung dengan mudah karena ikatan MT terhadap logam memiliki stabilitas termodinamik yang tinggi dan stabilitas kinetik yang rendah.
Selain terinduksi oleh logam, gen MT terinduksi oleh cekaman oksidatif pada jaringan gabus (Mir et al. 2004), terekspresi pada saat fase senescense pada daun (Buchanan-Wallaston 1994), dan saat pematangan buah (Davies & Robinson
2000). Seringkali beberapa tipe MT ditemui pada tanaman yang sama dan memiliki kesamaan kinerja seperti yang ditemui pada Arabidopsis, dimana Hernandez et al. (1998) menemukan bahwa terdapat overlapping antara MT1 dan MT2 dalam pola ekspresinya. Pada arabidopsis, MT ditemukan terekspresi walaupun tidak ada cekaman Cu. Saat tidak ada cekaman, MT1 terekspresi pada jaringan trikoma dan vaskular daun, akar, bunga, dan benih. Sedangkan saat dikenai cekaman Cu, MT1 diekspresikan pada jaringan mesofil daun dan tunas. MT2 ditemukan pada trikoma Arabidopsis, baik yang dikenai cekaman maupun yang tidak dikenai cekaman Cu.
Kemampuan MT untuk melakukan kompartemensasi Cd pada vakuola ditunjukkan oleh hasil penelitian Vogeli-Lange (1990). Pada vakuola daun tembakau (Nicotiana rustica), Cd terikat pada protein yang memiliki gugus (γ-Glu-Cys)3Gly dan (γ-Glu-Cys)4Gly, yang kemudian dikenali sebagai MT tipe 3.
Selain berfungsi untuk detoksifikasi logam berat, berbagai fungsi seluler MT adalah untuk pengaturan homeostasis logam esensial, perlindungan terhadap radiasi dan kerusakan oksidatif, dan kontrol seluler dalam poliferasi dan apoptosis (Klaassen 1998).
2.3 Melastoma affine L.
M. affine L. merupakan salah satu spesies dari famili Melastomataceae yang mampu mengakumulasi Al (Jansen et al. 2002), setelah Rubiaceae (Jansen et al. 2003). Menurut Li et al (2000), M. affine L. dapat hidup pada tanah dengan pH dibawah 4 dan mampu mengakumulasi Al sebanyak 9099 – 12810 mg/kg daun dan 3499 – 3633 mg/kg akar. Tanaman diklasifikasikan sebagai akumulator bila mengakumulasi Al sedikitnya 1000 mg/kg daun (Jansen et al. 2002).
Penelitian Watanabe et al (2001) menunjukkan bahwa selain tahan terhadap cekaman Al, pertumbuhan melastoma juga dipacu oleh Al pada konsentrasi yang rendah. Penelitian oleh Muhaemin (2008) menunjukkan bahwa perlakuan 3,2 mM Al pada pH 4 pada M. affine L. mempunyai pengaruh positif yang ditandai dengan pertambahan pertumbuhan akar sebesar 70,41 % setelah 6 minggu perlakuan dan tidak ditemui kerusakan jaringan pada ujung akar. Cekaman Al pada tanaman toleran akan menginduksi sejumlah gen untuk
8
menghindari pengaruh ion Al. Pada melastoma, gen-gen ini diduga tidak hanya berperan dalam mendetoksifikasi Al, tetapi juga menginduksi hormon pertumbuhan.
Watanabe dan Osaki (2003) menyatakan bahwa Al mampu menembus jaringan endodermis dan masuk ke dalam pembuluh xilem dan ditimbun di dalam daun. Penelitian Mutiasari (2008) menunjukkan adanya akumulasi Al sebesar 8,81 mg/gr daun tua tanaman M. affine L. yang diberi cekaman 3,2 mM Al pada pH 4 selama 2 bulan. Akumulasi Al tersebut dapat ditemui pada jaringan mesofil daun (Mutiasari 2008). Penelitian lain yang dilakukan oleh Watanabe dan Osaki (2002) menyatakan bahwa akumulasi Al terjadi terutama pada bagian vakuola daun. Selain itu juga terjadi akumulasi pada jaringan periderm sampai xilem pada akar dan pada jaringan korteks, xilem sekunder dan empulur batang (Mutiasari 2008). Didalam jaringan melastoma, Al ditemukan dalam bentuk Al monomerik, Al-oksalat, Al-(oksalat)2, dan Al-(oksalat)3 (Watanabe et al. 1998). Karena kemampuan Melastoma untuk mengakumulasi Al dalam jumlah yang tinggi, maka melastoma dapat dijadikan sebagai tanaman model bagi ketahanan Al terutama bagi tanaman dikotil.
2.4 Real Time Polimerase Chain Reaction (Real Time PCR)
Kemampuan sel untuk tumbuh, melakukan aktifitas, dan pertahanan terhadap cekaman merupakan refleksi dari ekspresi gen. Oleh karena itu, tingkat transkripsi dari gen spesifik dapat digunakan untuk mengetahui fungsi dari gen. Terdapat empat metode yang biasanya digunakan untuk mengetahui tingkat transkripsi yaitu northern blotting, in situ hybridization, Rnase protection assay, dan quantitatif reverse transcriptase. Dengan menggunakan analisis Northern, bisa diketahui ukuran dari mRNA dan integritas RNA sampel. Rnase protection assay memungkinkan untuk melakukan pemetaan daerah inisiasi pada transkripsi, daerah terminasi, batas ekson atau intron, dan membedakan mRNA yang memiliki ukuran yang sama, yang akan ditampilkan sebagai satu pita pada analisis Northern. In situ hybridization merupakan satu-satunya metode yang dapat digunakan untuk menunjukkan transkripsi lokal pada sel spesifik suatu jaringan. Kekurangan dari ketiga metode tersebut adalah kurang akurat (Bustin 2000).
Untuk mengetahui kuantitas ekspresi dari suatu gen, biasanya digunakan northern blotting atau PCR biasa yang menggunakan produk akhir sebagai dasar perhitungan. Penggunaan northern blotting untuk mengetahui ekspresi gen memerlukan jumlah RNA yang cukup banyak (Hunt 2006). Selain itu, pengukuran kuantitas ekspresi suatu gen dengan menggunakan hasil akhir dari proses PCR mempunyai kelemahan yaitu presisi yang kurang, tidak sensitif, resolusi rendah, tidak otomatis, diskriminasi hanya dari ukuran saja, hasil tidak diekspresikan secara kuantitatif (Applied Biosystem 2007).
Pada proses PCR, terdapat tiga fase utama yaitu eksponensial, linier, dan plateu (datar) (Gambar 1). Pada fase eksponensial, terjadi penggandaan dari jumlah sesungguhnya DNA atau RNA yang digunakan sebagai cetakan, sehingga perhitungan kuantitas DNA pada fase ini memiliki akurasi 100 %. Pada fase linier sudah terjadi penggunaan bahan reaksi, reaksi penggandaan terjadi pelan dan produk mulai terdegradasi. Dan saat plateu, tidak terjadi lagi proses penggandaan DNA dan pada saat ini juga terjadi degradasi DNA produk, oleh sebab itu deteksi ekspresi gen berdasarkan hasil akhir PCR memiliki akuransi yang rendah (Applied Biosystem 2007).
10
Gambar 1. Tiga fase utama pada proses PCR: (1) fase eksponensial dimana terjadi penggandaan jumlah DNA cetakan yang sebenarnya, (2) fase linier dimana proses penggandaan cetakan mulai menurun, (3) fase plateu dimana tidak terjadi lagi proses penggandaan cetakan dan DNA mulai terdegradasi. Deteksi untuk Real Time PCR terjadi pada fase eksponensial, sedangkan deteksi pada PCR biasa terjadi pada fase plateu yang digambarkan dalam bentuk kurva linier (A) dan kurva log (B) (Applied Biosystem 2007)
Real Time PCR adalah metode kuantifikasi PCR yang memungkinkan untuk mengamati pertambahan amplikon selama proses amplifikasi. Pada Real Time PCR, perhitungan amplikon dilakukan pada saat fase eksponensial (Gambar 1). Sistem Real Time PCR menggunakan dasar pada deteksi dan kuantifikasi fluorescence reporter. Signal akan meningkat sesuai dengan jumlah produk PCR pada setiap siklus (Dorak 2006). Ada dua macam fluorescence reporter yang biasanya digunakan dalam Real Time PCR, yaitu SYBR Green (Gambar 2) dan TaqMan (Gambar 3). SYBR Green merupakan senyawa kimia yang akan mendeteksi semua utas ganda DNA, biasa digunakan untuk perhitungan kuantitas DNA dengan tujuan yang tidak spesifik. Sedangkan TaqMan lebih spesifik daripada SYBR Green karena menggunakan TaqMan probe, yaitu probe yang komplemen dengan utas DNA spesifik yang berada diantara primer forward dan primer reverse dan didesain sedemikian rupa sehingga bila probe tersebut rusak akibat proses perpanjangan DNA pada proses PCR maka akan melepaskan energi yang dapat ditangkap oleh mesin Real Time PCR dengan dasar teknologi FRET (Fluorescent Resonance Energy Transfer). Teknologi FRET didasari oleh transfer Rn SIKLUS eksponensial Linier Plateu eksponensial Linier Plateu A. B.
energi dari pelacak berenergi tinggi (reporter dye) yang diletakkan didekat pelacak berenergi rendah (quencer dye), transfer energi tersebut akan terjadi bila pelacak berenergi tinggi rusak (Applied Biosystem 2007).
Gambar 2. Proses deteksi pada Real Time PCR dengan menggunakan SYBR Green. SYBR Green akan terdeteksi bila terikat pada utas ganda DNA saat fase perpanjangan DNA (extention) (Dorak 2006)
Gambar 3. Proses deteksi pada Real Time PCR dengan menggunakan TaqMan. Deteksi akan terjadi bila TaqMan probe yang menempel diantara primer forward dan primer reverse rusak selama fase perpanjangan DNA (extention) (Dorak 2006)
12
Fluorescence reporter akan meningkat sampai pada jumlah yang bisa terdeteksi oleh Real Time PCR dan digambarkan dalam bentuk kurva amplifikasi (Gambar 4). Kurva amplifikasi mengandung beberapa informasi penting: (1) threshold yaitu daerah deteksi yang terjadi pada saat jumlah fluorescence reporter bisa dideteksi oleh mesin Real Time PCR, agar hasil perhitungan lebih akurat threshold berada pada fase eksponensial; (2) Cycle Threshold (Ct) yaitu siklus pada proses PCR saat jumlah fluorescence reporter berpotongan dengan garis threshold yang ditetapkan untuk perhitungan pada Real Time PCR; (3) Rn yaitu jumlah fluorescence reporter yang terdeteksi; dan (4) Cycle yaitu jumlah siklus yang digunakan dalam proses PCR (Applied Biosystem 2007).
Gambar 4. Kurva amplifikasi pada Real Time PCR. Threshold adalah daerah deteksi yang digunakan dalam analisis. Cycle Threshold (Ct) adalah siklus pada saat fluorescence reporter berpotongan dengan threshold. Rn adalah jumlah fluorescence reporter yang terdeteksi selama proses PCR. Cycle adalah banyaknya siklus yang digunakan dalam proses PCR (Applied Biosystem 2007)
Terdapat dua macam perhitungan yang bisa dilakukan dengan Real Time PCR, yaitu (1) absolute quantitation yang digunakan untuk mengetahui jumlah molekul DNA yang tidak diketahui dengan menggunakan kurva standar yang sudah diketahui jumlah molekul DNA-nya. (2) Relative quantitation yang digunakan untuk mengetahui jumlah molekul DNA yang tidak diketahui dengan membandingkannya dengan sampel DNA yang lain yang digunakan sebagai pembanding (Applied Biosystem 2004).
Relative quantitation banyak digunakan untuk mengetahui tingkat ekspresi suatu gen pada suatu perlakuan dibandingkan dengan ekspresi gen yang sama pada perlakuan yang lain. Untuk mengetahui ekspresi suatu gen maka dibutuhkan gen referensi sebagai pembanding internal (endogenous control) jumlah DNA agar tidak terjadi kesalahan interpretasi akibat jumlah sel yang berbeda. Gen referensi adalah gen yang tidak dipengaruhi oleh lingkungan. Gen referensi yang paling banyak digunakan adalah β-aktin dan Glyceraldehyde-3-phosphat-dehydrogenase (GAPDH) (Bustin 2000).
Ada dua metode yang bisa digunakan untuk mengetahui tingkat ekspresi gen dengan menggunakan relative quantitation, yaitu metode kurva standar relatif (relative standard curve method) dan metode comparative Ct (comparative Ct method/∆∆Ct). Metode kurva standar relatif menggunakan kurva standar relatif yang akan digunakan untuk menghitung kuantitas sampel. Kurva standar yang digunakan dapat dibuat dari DNA apapun yang sudah diketahui kuantitasnya dengan pengenceran berseri. Oleh karena itu metode ini membutuhkan tingkat akurasi yang tinggi dalam pemipetan sampel DNA yang akan digunakan untuk kurva standar dan tidak membutuhkan validasi PCR sebelumnya. Namun metode ini membutuhkan lebih banyak bahan reaksi kimia dan membutuhkan banyak tempat reaksi PCR. Selanjutnya kuantitas gen target dan gen referensi dapat diketahui dengan memasukkan nilai Ct gen target dan gen referensi ke dalam kurva standar (log ng DNA dibanding nilai Ct). Ekspresi gen target pada suatu perlakuan dilakukan dengan menormalisasi kuantitas gen target dengan gen referensi dan membandingkan hasil normalisasi gen target pada perlakuan dengan hasil normalisasi gen target pada perlakuan kontrol (kalibrator) (Applied Biosystem 2004).
Gen target yang dinormalisasi (perlakuan/kalibrator)
referensi gen ng et t gen ng arg =
Ekspresi gen target pada perlakuan relatif terhadap kalibrator (Relative Quantification/RQ) kalibrator pada asi dinormalis yang et t gen perlakuan pada asi dinormalis yang et t gen arg arg =
14
Metode ∆∆Ct banyak digunakan untuk menghitung ekspresi gen dari sampel yang banyak. Karena tidak perlu menggunakan kurva standar, metode ini lebih menghemat bahan reaksi PCR dan tidak membutuhkan akuransi dalam pemipetan. Dalam metode ∆∆Ct, dibutuhkan validitas efisiensi reaksi PCR yang biasa dilakukan dengan merancang primer yang menghasilkan amplikon berukuran lebih kecil daripada 150 pb, dimana pada ukuran amplikon tersebut maka efisiensi PCR yang terjadi adalah 100%.
Metode ∆∆Ct menggunakan rumus aritmatika 2-∆∆CT untuk mengetahui ekspresi gen target yang telah dinormalisasi gen referensi, dan relatif terhadap kalibrator (Relative Quantification/RQ). Rumus aritmatika tersebut diperoleh dari rumus eksponensial amplifikasi proses PCR yaitu:
Xn = X0 x (1 + EX) Ct
= K dimana K= konstanta (1)
Dimana : Xn = jumlah DNA pada siklus ke n X0 = jumlah DNA mula-mula EX = efisiensi PCR
Ct = adalah siklus yang diinginkan atau Cycle threshold
Bila gen target disimbolkan dengan X dan gen referensi disimbolkan dengan R, maka rumus tersebut diatas dapat digunakan untuk mencari KX maupun KR. Sehingga ekspresi gen X dinormalisasikan dengan gen referensi R, adalah
R X CTR R CTX X T T K K E x R E x X R X = + + = ) 1 ( 0 ) 1 ( 0 (2)
Nilai efisiensi gen X (EX) dan gen R (ER) sama, sehingga
K E x R X + CTX−CTR = ) 1 ( 0 0 (3)
Nilai ekspresi suatu gen yang sesungguhnya adalah jumlah DNA awal yang digunakan sebagai cetakan, sehingga rumus (4) dapat diturunkan menjadi
XN = K x (1+E)-∆CT (4) Dimana : XN = 0 0 R X ∆CT = CTX-CTR
Dan untuk mengetahui ekspresi gen target X pada perlakuan relatif terhadap kalibrator, maka ekspresi gen X pada perlakuan (XNP) dibagi dengan ekspresi gen X pada kalibrator (XNK)
CT CTK CTP NK NP E E x K E x K X X −∆∆ ∆ − ∆ − + = + + = (1 ) ) 1 ( ) 1 ( (5) Dimana : -∆∆CT = -(∆CTP - ∆CTK)
Bila amplikon yang dihasilkan berukuran kurang dari 150 pb, maka nilai effisiensi PCR adalah 100% yang nilainya sama dengan 1, sehingga jumlah gen X pada suatu perlakuan yang telah dinormalisasi dengan gen referensi, dibandingkan dengan kalibrator adalah
2-∆∆CT (6)
(Livak & Schmittgen 2001) Real Time PCR sudah digunakan secara luas, yaitu untuk deteksi patogen, kuantifikasi virus, kuantifikasi ekspresi gen, efisiensi terapi pada obat-obatan baru, dan memastikan kerusakan DNA (Applied Biosystem 2007). Untuk ekspresi gen metallothionein (MT), beberapa peneliti juga menggunakan Real Time PCR antara lain van Hoof et al (2001) yang menggunakan metode ini untuk mengetahui perbedaan ekspresi MT pada tanaman yang sensitif terhadap tembaga (Cu) dan tanaman yang toleran terhadap Cu yang ditumbuhkan pada konsentrasi CuSO4 0,1 µM dan Moyle et al (2005) yang menggunakan kuantitatif Real Time PCR untuk mengetahui ekspresi gen MT pada saat pematangan buah nanas.
16
III. BAHAN DAN METODE
3.1 Waktu dan Tempat
Penelitian ini dilaksanakan pada bulan Juni 2007 – Desember 2008 di Laboratorium Biologi Molekuler dan Seluler Tanaman, Laboratorium BIORIN Pusat Penelitian Sumberdaya Hayati dan Bioteknologi Institut Pertanian Bogor, dan Laboratorium Terpadu, Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian, Badan Penelitian dan Pengembangan Pertanian. Perlakuan cekaman pH dan Al dilakukan di rumah kaca Institut Pertanian Bogor.
3.2 Bahan Penelitian
M. affine L. digunakan sebagai bahan tanaman. Primer forward (MF) 5’TCGAGAAAAATGTCTTGCTGTG3’ dan primer reverse (MR1) 5’GTCTCGCCGGAGAATCCCAAG3’ yang didesain dari MaMt2 (Suharsono et al. 2009) menghasilkan produk sebesar 113 pb digunakan untuk analisis ekspresi MT2. Primer forward (ActF) 5’ATGGCAGATGCCGAGGATAT3’ dan primer reverse (ActR2) 5’TCGAGGTCGGCCAACAATAC3’ yang didesain dari gen β actin kedelai (Shah et al. 1982) dan menghasilkan produk sebesar 123 pb, digunakan sebagai pembaku dalam analisis ekspresi. Primer forward (ActF) 5’ATGGCAGATGCCGAGGATAT3’ dan primer reverse (ActR) 5’CAGTTGTGCGACCACTTGCA3’ yang didesain dari gen β actin kedelai (Shah et al. 1982), digunakan untuk mengetahui kontaminasi DNA genom pada cDNA.
3.3 Metode
Penelitian ini dilakukan dengan melalui beberapa tahapan yaitu perlakuan cekaman pH dan Al rendah pada kultur air, isolasi RNA total dari M. affine L, sintesis cDNA total, dan analisis ekspresi gen dengan menggunakan Real Time PCR.
3.3.1 Perlakuan cekaman pada kultur air
Bahan tanam yang digunakan berasal dari stek pucuk tanaman M. affine L. yang diambil dari Jasinga, Kabupaten Bogor, Jawa Barat. Pengadaan stek pucuk dilakukan dengan memotong 4 - 6 ruas dari pucuk tanaman. Pucuk ditanam pada tanah Jasinga dan ditutup dengan plastik selama seminggu untuk mengurangi penguapan. Setelah itu, tutup plastik dibuka dan dibiarkan selama sebulan sampai tumbuh akar. Selanjutnya pucuk yang mempunyai akar kurang lebih 1 cm ditanam pada tempat yang berisi larutan nutrisi standar pH 6 dengan komposisi NH4NO3 2,14 mM; NaH2PO4 3,2 µM; K2SO4 : KCl (1:1) 0,77 mM; CaCl 1,2 mM; MgSO4 0,82 mM; FeSO4 35,8 µM; MnSO4 9,1 µM; H3BO3 46,3 µM; ZnSO4 3,1 µM; CuSO4 0,16 µM; (NH4)6Mo7O24 0,05 µM (Watanabe et al. 1998) dan diberi aerasi, selama seminggu. Setelah satu minggu, tanaman siap untuk diberi perlakuan cekaman pH dan Al.
Percobaan dilakukan dengan dua macam perlakuan dan dua ulangan. Perlakuan yang diberikan adalah pH dan konsentrasi Al. Perlakuan pH menggunakan 2 tingkat, yaitu pH 6 (kontrol) dan 4. Sedangkan perlakuan konsentrasi Al yang diberikan adalah 0 mM (kontrol), 0,8 mM dan 3,2 mM yang dilakukan pada pH 4. Sampel berupa daun dan akar diambil 1 minggu setelah perlakuan untuk isolasi RNA total agar didapatkan ekspresi gen yang sesungguhnya.
3.3.2 Isolasi RNA total dari Melastoma affine L.
RNA total diisolasi dari akar dan daun secara terpisah, dari dua ulangan dan dua macam perlakuan, menggunakan metode CTAB (Chang et al. 1993) yang dimodifikasi. Untuk itu, 1 g bahan tanaman digerus didalam 10 ml buffer ekstraksi (CTAB 2%; Tris pH 9,5 0,1 M; EDTA 20 mM; NaCl 1,4 M; PVP 25000 2%; dan β-mercaptoethanol 1%) hangat hingga membentuk suspensi sel. Suspensi sel dimasukkan ke tabung 20 ml dan diinkubasi pada suhu 65oC selama 10 menit, kemudian didinginkan, dan ditambah 10 ml kloroform: isoamil alkohol (24 : 1). Setelah dicampur dengan menggunakan vortex, campuran disentrifugasi dengan kecepatan 42000xg (rotor SW11, Sorval Ultra Pro 80) pada suhu 4oC selama 10 menit. Cairan bagian atas dipindahkan ke tabung 20 ml, ditambahkan ¼
18
volume LiCl 10 M dan diinkubasi pada suhu -32oC selama 2,5 jam. Campuran disentrifugasi pada kecepatan 42000xg pada suhu 4oC selama 10 menit. Cairan dibuang dan endapan RNA total disuspensikan dalam 500 µl TE 1X (Tris HCl pH 7,4 10 mM dan EDTA 1 mM) kemudian dipindahkan ke tabung 1,5 ml. Suspensi RNA total diekstraksi dengan penambahan 1X volume fenol pH 9, divortex dan disentrifugasi pada kecepatan 8000xg (Jouan BR4i) pada suhu 20oC selama 10 menit. Cairan bagian atas yang mengandung RNA total diambil, dimasukkan dalam tabung 1,5 ml, dan diekstraksi kembali dengan 1X volume fenol : kloroform : isoamil alkohol (25:24:1), divortex dan disentrifugasi pada kecepatan 8000xg (Jouan BR4i) pada suhu 20oC selama 10 menit. Cairan bagian atas diambil, dimasukkan dalam tabung 1,5 ml, ditambah ¼X volume LiCl 10 M dan kemudian diinkubasi pada suhu -32oC selama 2,5 jam. Cairan disentrifugasi pada kecepatan 8000xg pada suhu 4oC selama 15 menit. Cairan dibuang dan endapan RNA total dibilas dengan penambahan 500 µl ethanol 70 % dan disentrifugasi pada kecepatan 5000xg pada suhu 4oC selama 10 menit. Endapan RNA total dikeringkan dengan pengering vakum dan disuspensikan dengan 20 µl DEPC H2O. Keutuhan RNA total diuji dengan melakukan elektroforesis RNA total di gel agarose 1 % dalam buffer MOPS 1X (MOPS 4,2 g/l; Na asetat 0,41 g/l; Na2EDTA 0,37 g/l). Visualisasi RNA total dilakukan di atas UV transluminator GelDoc (Labquip) setelah diwarnai dengan EtBr (0,5 µg/ml) selama 30 menit dan dibilas dengan air.
3.3.3 Pembentukan cDNA total
Untuk mendapatkan RNA yang bebas dari DNA genom, suspensi RNA total diberi Dnase sebelum sintesis cDNA dengan mencampur 5 µg RNA total, buffer Dnase I 1X, Dnase I bebas RNase (Fermentas) 0,2 U, dan DEPC H2O sampai volume total mencapai 10µl. Larutan diinkubasi pada suhu ruang selama 5 menit. Setelah itu ditambahkan EDTA 2,5 mM dan diinkubasi kembali pada suhu 65oC selama 10 menit.
Sintesis cDNA total dilakukan dengan mencampur 5 µg RNA total, First-Strand Reverse Transcriptase Buffer 1X, oligo (dT) 20 pmol, dNTP 4 mM, DTT 10 mM, enzim SuperScript II Reverse Transcriptase (Invitrogen) 40 U, dan
DEPC-H2O dengan volume reaksi 20 µl. Proses sintesis cDNA dilakukan dengan alat PCR (MJ Research TM 100) dengan menginkubasi campuran pada suhu 30oC selama 10 menit, kemudian inkubasi pada suhu 42oC selama 50 menit, inkubasi pada suhu 95oC selama 5 menit dan 20oC selama 5 menit.
Keberhasilan dan kemurnian sintesis cDNA total dan kemurnian cDNA total dari DNA genom diverifikasi dengan menggunakan PCR menggunakan primer forward ActF dan primer reverse ActR yang spesifik untuk ekson1 – ekson2 dari aktin. Reaksi PCR yang digunakan adalah 1 µl cDNA total, Taq buffer 1X, dNTP mix 4 mM, ActF 20 pmol dan ActR 20 pmol, enzim Taq DNA polimerase (RBC) 0,5 U, DMSO 4% dan dH2O sampai volume akhir 10 µl. PCR dilakukan dengan kondisi pra-PCR pada 94oC, 5 menit; denaturasi pada 94oC, 30 detik; penempelan primer pada 55oC, 30 detik; pemanjangan pada 72oC, 1,5 menit; pasca PCR pada 72oC, 7 menit; dan pendinginan 20 oC, 5 menit. PCR dilakukan sebanyak 30 siklus. Amplifikasi gen β actin ekson1-ekson2 dengan menggunakan DNA genom melastoma sebagai cetakan digunakan sebagai kontrol terjadinya kontaminasi terhadap sintesis cDNA. Keberhasilan sintesis cDNA diketahui dari terbentuknya pita berukuran 450 pb. Bila terjadi kontaminasi DNA genom pada cDNA, maka akan didapatkan 2 pita yang masing-masing berukuran 450 pb (dari cDNA) dan 550 pb (dari DNA genom).
3.3.4 Analisis ekspresi gen dengan menggunakan Real Time PCR
Real Time PCR dilakukan dengan komposisi 5 µl cDNA, Power SYBR Green PCR master mix 1X, primer MF atau ActF 50 nM, primer MR1 atau ActR2 50 nM dalam total reaksi 50 µL. Reaksi dilakukan untuk dua jenis gen, yaitu MaMt2 sebagai gen target dan aktin sebagai gen pembaku. Deteksi ekspresi gen dilakukan dengan menggunakan mesin Real Time PCR ABI Prism 7000 Sequence Detection System (Applied Biosystem). Amplifikasi gen target dengan menggunakan sistem touchdown PCR, yaitu dengan kondisi aktivasi enzim taq pada suhu 95oC selama 10 menit, kemudian diikuti dengan touchdown PCR yaitu denaturasi pada suhu 95oC selama 15 detik, annealing pada suhu 65oC selama 30 detik, dan sintesis DNA pada suhu 72oC selama 30 detik, dimana suhu annealing diturunkan 2oC setiap 3 siklus dari suhu awal 65oC sampai suhu annealing
20
mancapai 55oC. Saat suhu annealing mencapai 55oC, proses PCR dilakukan sebanyak 30 siklus.
Hasil dari Real Time PCR berupa kurva amplifikasi dari masing-masing reaksi dianalisa dengan menggunakan program Relative Quantification Study dengan metode ∆∆CT berdasarkan metode Livak (2001). Dari kurva amplifikasi dapat diperoleh nilai CT dari masing-masing reaksi. Nilai ∆CT dari masing-masing perlakuan diperoleh dari normalisasi gen target dengan gen pembaku yaitu dengan cara pengurangan nilai CT gen MT2 sebagai gen target dengan nilai CT gen aktin sebagai gen referensi, sehingga rumus untuk mendapatkan ∆CT adalah : ∆CT = CT MT2 – CT actin. Sedangkan nilai ∆∆CT diperoleh dengan cara pengurangan nilai ∆CT dari perlakuan dan nilai ∆CT dari kalibrator (kontrol). Dalam penelitian ini terdapat dua kontrol yaitu pH 6 untuk mengetahui pengaruh perlakuan pH pada ekspresi gen MT2 dan pH 4 dengan 0 mM Al untuk mengetahui pengaruh cekaman aluminium pada ekspresi gen MT2. Selanjutnya nilai ∆∆CT tersebut akan digunakan untuk mengetahui ekspresi (RQ) gen MT2 pada perlakuan relatif terhadap kontrol dengan memasukkannya dalam rumus aritmatika 2 –∆∆CT. Analisis ekspresi gen dengan menggunakan Real Time PCR ini diulang 2 kali dan nilai RQ antar perlakuan dianalisis dengan menggunakan program Statistical Product and Service Solution (SPSS) 13.0 for windows. Amplikon hasil dari Real Time PCR, dicek pada gel agarose untuk mendapatkan ekspresi secara semikuantitatif dan kualitas pita DNA.
IV. HASIL DAN PEMBAHASAN
4.1 Isolasi RNA Total
RNA total telah berhasil diisolasi dari daun dan akar M. affine L. Keutuhan RNA total ditunjukkan dengan adanya 2 pita, yaitu rRNA 28S dan 18S (Gambar 5) karena rRNA merupakan jenis RNA yang paling banyak ditemui di dalam sel dan rRNA 28S dan 18S merupakan dua jenis RNA yang memiliki ukuran paling besar, dan pada saat dimigrasikan kedua jenis rRNA ini tampak paling jelas sehingga bisa digunakan untuk menentukan integritas RNA total yang didapatkan. Bila terjadi degradasi pada RNA total, maka intensitas kedua pita rRNA tersebut tidak sama (Bowtell & Sambrook 2003). Sehingga, keberadaan kedua pita rRNA ini menunjukkan bahwa RNA total yang diisolasi mempunyai keutuhan yang tinggi sehingga sangat baik digunakan sebagai cetakan untuk sintesis cDNA total. Selain kedua pita tersebut, RNA total dari daun mengandung beberapa pita RNA yang kemungkinan adalah rRNA yang terdapat pada mitokondria dan kloroplas yang berukuran 23S, 16S, dan 5S.
Gambar 5. RNA total M. affine L. hasil isolasi dari akar (1) dan daun (2)
RNA total mengandung mRNA sehingga RNA total yang utuh juga mengandung mRNA yang utuh. Oleh sebab itu, keutuhan RNA total sangat penting untuk sintesis cDNA total.
4.2 Sintesis cDNA Total
Dari RNA total yang diisolasi, telah berhasil disintesis cDNA total melalui transkripsi balik dengan menggunakan RNA total sebagai cetakan. Dengan primer oligo-dT, hanya mRNA yang dapat disintesis menjadi cDNA karena mempunyai poliA pada ujung 3’ sedangkan rRNA dan tRNA tidak mempunyai poliA. PCR
1. 2.
28S 18S
22
dengan menggunakan primer ActF dan ActR yang dirancang dari ekson 1 dan ekson 2 gen aktin menghasilkan DNA yang berukuran sekitar 450 pb (Gambar 6) yang menunjukkan bahwa daerah yang diamplifikasi adalah cDNA bukan DNA genom dari gen actin. Gen aktin memiliki ukuran sekitar 550 pb karena mengandung intron yang akan dibuang pada saat pembentukan mRNA. Teramplifikasinya cDNA aktin dengan ukuran 450 pb menunjukkan bahwa sintesis cDNA total melalui proses transkripsi balik telah berlangsung dengan baik. Hal ini juga menunjukkan bahwa RNA total yang telah diisolasi mempunyai kualitas yang sangat bagus, karena selain dapat digunakan untuk mensintesis cDNA juga terbebas dari kontaminasi DNA. Oleh sebab itu, cDNA total ini dapat digunakan sebagai bahan untuk analisis ekspresi gen.
Gambar 6. Hasil amplifikasi β-actin. Lajur 1,2, dan 3 = hasil amplifikasi dengan menggunakan cDNA sebagai cetakan. Lajur 4 = 1 kb ladder. Lajur 5 = hasil amplifikasi dengan menggunakan DNA genom sebagai cetakan.
4.3 Ekspresi Gen Metallothionein
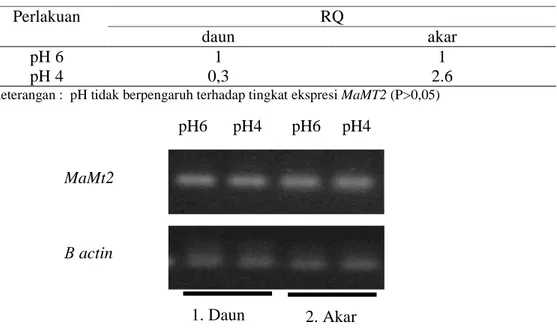
4.3.1 Pengaruh pH rendah terhadap ekspresi gen metallothionein
Untuk mengetahui tingkat ekspresi MaMt2 pada pH rendah, perlakuan pH 6 digunakan sebagai pembanding dan pH 4 sebagai perlakuan. Analisis ekspresi gen dilakukan pada daun dan akar M. affine L. Hasil ANOVA (Lampiran 1) terhadap ekspresi gen MaMt2 dari M. affine L. menunjukkan bahwa ekspresi MaMt2 pada akar dan pada daun antara M. affine L. yang mendapat cekaman pH 6 dan pH 4 tidak berbeda nyata (Tabel 1), walaupun terdapat perbedaan ekspresi yang relatif besar antara perlakuan pH 6 dan pH 4. Hal ini mungkin disebabkan oleh perbedaan ekspresi yang tinggi dalam ulangan pada perlakuan yang sama (Lampiran 2). Pengamatan pada gel agarose (Gambar 7) tidak menunjukkan
450 pb
1 2 3 4 5
adanya perbedaan ekspresi gen MaMt2 yang terlihat dari perpendaran pita MaMt2 yang sama antara perlakuan pH 6 dan pH 4 baik pada akar ataupun pada daun.
Pada perlakuan pH rendah, tidak ada logam yang diberikan secara berlebihan pada media tumbuh. Metallothionein merupakan protein yang berfungsi untuk mendetoksifikasi logam dan untuk menjaga homeostasis logam didalam sel (Cobbett and Goldbrough, 2002). Bila logam berupa ion bebas yang disebabkan pengaruh pH rendah masih berada dibawah konsentrasi yang dapat menyebabkan keracunan, maka ion ini tidak mempengaruhi ekspresi MaMt2. Ion logam (Cu, Mg, Mn, Fe, Zn) yang terdapat pada medium Watanabe et al. (1998) merupakan logam esensial yang dibutuhkan tanaman untuk tumbuh dengan baik, untuk membentuk struktur protein dan katalisis enzim yang terlibat dalam metabolisme dan pertumbuhan tanaman (Haydon et al. 2007).
Tabel 1. Nilai relative quantification (RQ) dari transkripsi MaMt2 pada daun dan akar M. affine L. yang mendapat cekaman pH rendah
RQ Perlakuan
daun akar
pH 6 1 1
pH 4 0,3 2.6
Keterangan : pH tidak berpengaruh terhadap tingkat ekspresi MaMT2 (P>0,05)
Gambar 7. Ekspresi MaMt2 semikuantitatif pada daun dan akar dari M. affine L. yang mendapat cekaman pH rendah
MaMt2
Β actin
pH6 pH4 pH6 pH4
24
4.3.2 Pengaruh aluminium terhadap ekspresi gen metallothionein
Pengaruh aluminium pada ekspresi gen metallothionein dapat diketahui dengan cara melihat tingkat ekspresi MaMt2 pada perlakuan pemberian Al 0,8 mM dan 3,2 mM setelah dibandingkan dengan perlakuan Al 0 mM sebagai kontrol. Analisis ekspresi dilakukan pada daun dan akar M. affine L.
Uji Tukey (Lampiran 3) menunjukkan bahwa tidak ada perbedaan nyata antar perlakuan Al 0 mM, Al 0,8 mM, dan Al 3,2 mM terhadap ekspresi gen MaMt2 di daun (Tabel 2). Sedangkan pada akar, perlakuan Al 0,8 mM tidak berbeda nyata terhadap ekspresi MaMt2 bila dibandingkan dengan Al 0 mM. Namun, perlakuan Al 3,2 mM menginduksi ekspresi gen MaMt2 di M. affine L. Ekspresi MaMt2 di akar dari tanaman M. affine L. yang mendapatkan cekaman Al 3,2 mM meningkat 2 kali lipat dibandingkan dengan perlakuan Al 0 mM dan 0,8 mM. Pengecekan pada gel agarose menunjukkan perpendaran pita MaMt2 yang sama pada perlakuan Al 0 mM dan Al 0,8 mM baik pada daun maupun pada akar, sedangkan perpendaran pita MaMt2 pada perlakuan Al 3,2 mM pada akar meningkat intensitasnya (Gambar 8).
Tabel 2. Nilai relative quantification (RQ) dari transkripsi MaMt2 pada daun dan akar M. affine L. yang mendapat perlakuan Al pada medium dengan pH 4 RQ Perlakuan Al daun akar 0 mM 1 1 0.8 mM 1.1 1 3.2 mM 0.5 2.1*
Keterangan : * mengindikasikan beda nyata secara statistik (P<0,05)
Gambar 8. Ekspresi MaMt2 semikuantitatif pada daun dan akar dari M. affine L. yang mendapat cekaman Al
MaMT2
β actin
0 mM 3,2 mM 0 mM 0,8 mM 3,2 mM
1. Daun 2. Akar
Cekaman Al tidak meningkatkan ekspresi MaMt2 di daun, tetapi meningkatkan ekspresi MaMt2 di akar. Hal ini dapat terjadi karena akar merupakan organ pada tumbuhan yang berinteraksi langsung dengan Al pada media tumbuh. Menurut Hernandes (1998) ekspresi MT dipengaruhi oleh spesies tanaman, jenis jaringan dan tipe MT, sehingga ekspresi jenis MT yang sama pada spesies yang berbeda akan berbeda, begitu juga bila berada pada jaringan yang berbeda.
Pada akar, ekspresi MaMt2 meningkat pada kondisi cekaman Al yang tinggi (3,2 mM) selama 7 hari. Sedangkan pada cekaman Al 0,8 mM, tidak terjadi perubahan tingkat ekspresi dibandingkan dengan tanpa cekaman Al. Hal ini dapat terjadi karena M. affine L. merupakan tanaman yang sangat toleran terhadap Al, sehingga pada konsentrasi Al 0,8 mM, diduga belum menginduksi ekspresi MaMt2, bila dibandingkan dengan tanpa cekaman Al.
Pada gandum varietas Warigal yang peka terhadap Al, cekaman Al 10 µM selama 48-96 jam meningkatkan ekspresi MT2 secara nyata, sedangkan pada varietas Waalt yang toleran Al cekaman 10 µM Al tidak menginduksi ekspresi MT2. Pada varietas yang toleran ini, ekspresi MT2 diinduksi pada cekaman Al dengan konsentrasi yang lebih tinggi yaitu 100 µM (Snowden et al. 1993). Hal ini menunjukkan bahwa semakin toleran suatu tanaman, maka semakin tinggi konsentrasi Al yang dibutuhkan untuk menginduksi ekspresi MT2. Hal ini terjadi pada M. affine L. yang merupakan tumbuhan yang sangat toleran Al, bahwa konsentrasi 0,8 mM Al belum dapat meningkatkan ekspresi MT2 dan peningkatan ekspresi MT2 diinduksi oleh konsentrasi Al yang sangat tinggi yaitu 3,2 mM.
26
V. SIMPULAN DAN SARAN
5.1 Simpulan
Cekaman pH 4 tidak menyebabkan kenaikan ekspresi MaMt2 baik pada akar maupun pada daun. Sedangkan cekaman aluminium pada konsentrasi 3,2 mM menyebabkan kenaikan ekspresi MaMt2 pada akar tetapi tidak pada daun dari M. affine L.
5.2 Saran
Untuk melakukan konfirmasi peranan MaMt2 dalam toleransi tanaman terhadap aluminium, MaMt2 harus diekspresikan secara berlebih (over-expression) pada tanaman sensitif Al seperti tembakau dan ditiadakan ekspresinya pada tumbuhan yang sangat toleran terhadap Al seperti M. affine L.
DAFTAR PUSTAKA
Applied Biosystem. 2004. Guide to Performing Relative Quantitation of Gene Expression Using Real-Time Quantitative PCR. Applied Biosystem
Applied Biosystem. 2007. Real Time PCR VS Traditional PCR. Applied Biosystem
Barcelo J, Poschenrieder C. 2003. Phytoremediation : principles and perspectives. Science 2 (3) : 332-344
Bowtell D, Sambrook J. 2003. DNA Microarrays: A Molecular Cloning Manual. Cold Spring Harbor Laboratory Press. New York
Buchanan-Wollaston V. 1994. Isolation of cDNA clones for genes that are expressed during leaf senescence in Brassica napus. Identification of a gene encoding a senescence-specific metallothionein-like protein. Plant Physiol 105: 839-846
Bustin SA. 2000. Absolute quantification of mRNA using real time reverse transcription polymerase chain reaction assays. Review. J Mole Endocrin 25: 169-193
Bustin SA, Benes V, Nolan T, Pfaffl MW. 2005. Quantitative real time RT-PCR – a perspective. Review. J Mole Endocrin 34: 597–601
Carr HP, Lombi E, Kupper H, Mcgrath SP, Wong MH. 2003. Accumulation and distribution of aluminum and other elements in tea (Camellia sinensis) leaves. Agronomie 23: 705-710
Cobbett C, Goldbrough P. 2002. Phytochelatin and metallothionein: roles in heavy metal detoxification and homeostatis. Annu Rev Biol 53: 159-182
Davies C, Robinson SP. 2000. Differential screening indicates a dramatic change in mRNA profiles during grape berry ripening. Cloning and characterization of cDNAs encoding putative cell wall and stress response proteins. Plant Physiol 122: 803-812
Dorak MT. 2006. Real Time PCR. Advance method series. www.dorak.info/genetics/realtime.html [29Apr 2007]
Duncan KER et al. 2006. Peptide folding, metal binding mechanisms, and binding site structure in metallothioneins. Society for experimental biology and medicine. Pp:1488-1499
28
Frank BC. 2007. Two step real time RT-PCR protocol. http://pga.tigr.org/sop/RT-PCR.pdf [3 Mei 2007]
Hall Jl. 2001. Cellular mechanisms for heavy metal detoxification and tolerance. J Exp Bot 53 (366): 1-11
Haydon MJ, Cobbett CS. 2007. Transporters of ligands for essential metal ions in plants. Research review. NewPhytologist 174:499 – 506
Hernandez AG, Murphy A, Taiz L. 1998. Metallothioneins 1 and 2 have distinct but overlapping expression patterns in Arabidopsis. Plant Physiol 118: 387–397
Hunt M. 2006. Real Time PCR. The Board of Trustees of the University of South Carolina
Jansen S, Watanabe T, Smets E. 2002. Aluminum accumulation in leaves of 127 species in Melastomataceae with comment on the order Myrtales. Annals Bot 90: 53-64
Jansen S, Watanabe T, Dessein S, Smets E, Robbrecht E. 2003. A comparative study of metal level in leaves of some Al-accumulating rubiaceae. Annals Bot 91 : 657-663
Klaassen CD. 1998. Metallothionein IV. Birkhauser Verlag. Basel. Switzerland
Livak KJ, Schmittgen TD. 2001. Analysis of relative gene expression data using real time quantitative PCR and the 2 –∆∆CT method. Methods 25 : 402-408
Li J, Xie Z, Ye Z, Wong MH. 2000. Aluminum uptake and accumulation by aluminum excluder and accumulators as influenced by pH in plants and pH change in rhizophere soil. www.chinawater.net[29 Apr 2007]
Ma JF, Hiradate S, Matsumoto H. 1998. High aluminium resistant in buckwheat. II. Oxalic acid detoxifies aluminium internally. Plant Physiol 117: 753-759
Mir G et al. 2004. A plant type 2 metallothionein (MT) from cork tissue responds to oksidative stress. J Exp Bot 55 (408): 2483–2493
Moyle R et al. 2005. Developing pineapple fruit has a small transcriptome dominated by metallothionein. J Exp Bot 55 (409): 101-112
Muhaemin. 2008. Analisis pertumbuhan melastoma (Melastoma malabathricum auct. non L. dan M. affine D. Don) yang mendapat cekaman pH rendah dan aluminium [Thesis]. Bogor: Program Pascasarjana, Institut Pertanian Bogor
Mutiasari A. 2008. Akumulasi aluminium pada Melastoma affine dan Melastoma malabathricum [Thesis]. Bogor: Program Pascasarjana, Institut Pertanian Bogor
Murphy A, Taiz L. 1995. Comparison of metallothionein gene expression and non protein thiol in ten arabidopsis ecotypes. Corelation with cooper tolerance. Plant Physiol 109: 945-954
Okada K, Wisuwa M. 2004. Soil acidity and related problems in upland rice in the tropics. Di dalam: Toriyama K, Heong KL, Hardy B, editor. Rice is life: scientific perspectives for the 21st century. Proceeding of the World Rice Research Conference; Tsukuba, 5-7 Nov 2004. Japan: IRRI Publications. Pp: 454-456
Prasetyo BH, Suriadikarta DA. 2006. Karakteristik, potensi, dan teknologi pengelolaan tanah ultisol untuk pengembangan pertanian lahan kering di Indonesia. J Litbang Pertanian 25 (2): 39-46
PE Biosystem. 2007. Sequence Detection System Quantitative Assay Design and Optimizing. PE Biosystem
Pusat Penelitian Tanah dan Agroklimat. 2000. Atlas Sumberdaya Tanah Eksplorasi Indonesia Skala 1: 1.000.000. Pusat Penelitian Tanah dan Agroklimat. Bogor
Reid KE, Olsson N, Schlosser J, Peng F, Lund ST. 2006. An optimized grapevine RNA isolation procedure and statistical determination of reference genes for real time RT PCR during berry development. Methodology article. BMC Plant Biol 6 (27): 1-11
Ruan J, Wong MH. 2004. Aluminum absorption by intact roots of the Al accumulating plant Camellia sinensis L. Agronomie 24 : 137-142
Robbins AH et al. 1991. Refined crystal structure of Cd, Zn metallothinein at 20A resolution. J Mol Biol 221 (4): 1269-1293
Robinson NJ, Tommey AM, Kuske C, Jackson PJ. 1993. Plant metallothioneins. Biochem J 295: 1-10
Schultze P et al. 1988. Conformation of [Cd7]-metallothionein-2 from rat liver in aqueous solution determined by nuclear magnetic resonance spectroscopy. J Mol Biol 203 (1): 251-268
Shah DM, Hightower RC, Meagher RB. 1982. Complete nucleotide sequence of a soybean actin gene. Proc Natl Acad Sci USA 79: 1022-1026
Snowden KC, Gardner RC. 1993. Five genes induced by aluminum in wheat (Triticum aestivum L.) roots. Plant Physiol 103: 855-861
30
Snowden KC, Richard KD, Gardner RC. 1995. Aluminum induced genes induction by toxic metal, low cadmium, wounding and pattern of expression in root tips. Plant Physiol 107: 341-348
Subagyo H, Suharta N, Siswanto AB. 2004. Tanah-tanah pertanian Indonesia. Di dalam Sumberdaya Lahan Indonesia dan Pengelolaannya. 21-66
Suharsono, Trisnaningrum N, Sulistyaningsih LD, Widyastuti U. 2009. Isolation and cloning of cDNA of gene encoding for metallothionein type 2 from Melastoma affine. Biotropia (in press).
Taylor GJ. 1991. Current views of the aluminum stress response: the physiological basis of tolerance. Curr. Top. Plant Biochem and Physiol. 10 : 57-93
van Hoof NALM et al. 2001. Enhanced copper tolerance in Silene vulgaris (Moench) garcke populations from copper mines is associated with increased transcript levels of a 2b-type metallothionein gene. Plant Physiol 126: 1519–1526
Vitorello VA, Capaldi FR, Stefanuto VA. 2005. Recent advance in aluminum toxicity and resistance in higher plant. Braz J Plant Physiol 17 (1): 129-143
Vogeli-Lange R, Wagner GJ. 1990. Subcellular localization of cadmium and cadmium-binding peptides in tobacco leaves implication of a transport fungtion for cadmium-binding peptides. Plant Physiol 92: 1086-1093
Watanabe T, Osaki M, Yoshihara T, Tadano T 1998. Distribution and chemical speciation of aluminum in the Al accumulator plant, Melastoma malabathricum L. Plant Soil 201: 165-173
Watanabe T, Osaki M. 2001. Influence of aluminum and phosphorus on growth and xylem sap composition in Melastoma malabathricum L. Plant Soil 237 : 63-70
Watanabe T, Osaki M. 2002. Role of organic acid in aluminum accumulation and plant growth in Melastoma malabathricum. Tree Physiol 22: 785-792
Watanabe T, Jansen S, Osaki M. 2003. A physiological study of Melastoma malabathricum, an aluminum accumulating woody plant. http:www.keele.ac.uk/depths/ch/groups/aluminium/watanabe.po.pdf [3 Mei 2007]
Zangger K et al. 2001. Oxidative dimerization in metallothionein is a result of intermolecular disulphide bonds between cysteins in the α-domain. J Biochem 359: 353-360
Zheng SJ, Ma JF, Matsumoto H. 1998. High aluminum resistance in buckwheat I. Al-induced specific secretion of oxalic acid from root tips. Plant Physiol 117: 745–751
Zimeri AM, Dhankher OK, McCaig B, Meagher RB. 2005. The plant MT1 metallothioneins are stabilized by binding cadmium and are required for cadmium tolerance and accumulation. Plant Mol Biol 58: 839-855
32
Lampiran 1. Tabel ANOVA dari pengaruh pH terhadap ekspresi MaMt2 di daun dan akar M. affine L.
ANOVA RQ pada daun
Sumber keragaman Jumlah kuadrat Derajat bebas Kuadrat tengah F Sig. perlakuan 0,437 1 0,437 10,490 0,084 ulangan 0,083 2 0,042 Total 0,520 3
ANOVA RQ pada akar
Sumber keragaman Jumlah kuadrat Derajat bebas Kuadrat tengah F Sig. perlakuan 2,665 1 2,665 2,631 0,246 ulangan 2,026 2 1,013 Total 4,691 3