253 (2000) 193–209
www.elsevier.nl / locate / jembe
Effects of temperature and salinity on nitrogenous excretion
by Litopenaeus vannamei juveniles
a b a
*
Dong-Huo Jiang , Addison L. Lawrence , William H. Neill , Hui Gong
a
Shrimp Mariculture Project, Texas Agricultural Experiment Station, Texas A&M University System, 1300 Port Street, Port Aransas, TX 78373, USA
b
Department of Wildlife and Fisheries Sciences, Texas A&M University, College Station, TX 77843, USA
Received 15 February 2000; received in revised form 27 June 2000; accepted 11 July 2000
Abstract
Excretion rates of ammonia-N, nitrite-N, nitrate-N, and dissolved organic nitrogen (DON) for juvenile Litopenaeus vannamei (3.8560.83 g) were quantified in response to nine different combinations of temperature (24, 28, and 328C) and salinity (10, 25, and 40 ppt) under laboratory conditions. Results indicated that L. vannamei is ammonotelic, with ammonia-N accounting for 61.9–84.3% of total nitrogen (TN) excretion. There were significant effects of temperature and salinity, but no significant interaction between them, on ammonia-N excretion rate (RAN). RAN increased with increasing temperature, over the interval 24–328C. RAN was lower at 25 ppt than at 10 and 40 ppt, at all temperatures. DON excretion rate (RDON) was not significantly influenced by
21 21
either temperature or salinity; the overall mean RDON was about 5.24mg-N g h . However, the percentages of DON in TN (PDON) varied from 15.4 to 36.4% under the various temperature– salinity combinations. PDON at 28 and 328C was significantly lower than at 248C, and PDONat 10 ppt was significantly lower than at 25 and 40 ppt. Only very small amounts of nitrogen were excreted by L. vannamei as nitrite-N and nitrate-N. 2000 Elsevier Science B.V. All rights reserved.
Keywords: Litopenaeus vannamei; Nitrogenous excretion; Salinity; Temperature
1. Introduction
The white shrimp, Litopenaeus vannamei Boone, is a tropical species with its natural
*Corresponding author. Present address: Arizona Mariculture Associates, 50621 Agua Caliente Road, Dateland, AZ 85333, USA. Tel.:11-520-454-2364; fax:11-512-454-2364.
E-mail address: [email protected] (D.-H. Jiang).
distribution in the Eastern Pacific Ocean, off Northern Mexico to Northern Peru, between the 208C isotherms (Holthuis, 1980). L. vannamei is the dominant penaeid species currently cultivated in the Western Hemisphere. Temperature and salinity are two very important environmental factors in the culture of this and other shrimp species. The optimal temperature for the growth of L. vannamei has been reported to be size-specific, around 28–308C for postlarvae (Ponce-Palafox et al., 1997), greater than 308C for small juveniles (,5 g) and about 278C for subadults (Wyban et al., 1995). It is known that L.
vannamei can tolerate a wide salinity range from brackish water of 1–2 ppt to
hypersaline water of 50 ppt (Pante, 1990; Stern et al., 1990). Boyd (1989) considered salinity of 15–25 ppt to be ideal for L. vannamei culture. But, in view of inconsistencies in published information regarding salinity effects on shrimp survival and growth, the optimum salinity for L. vannamei is still not conclusive. Significant effects of temperature and salinity have been reported on survival (Ogle et al., 1992), molting frequency (Pante, 1990), oxygen consumption (Villarreal et al., 1994; Martinez-Pakacios et al., 1996), and growth of L. vannamei (Huang, 1983; Wyban et al., 1995).
In decapod crustaceans, nitrogen is mainly excreted as ammonia (60–70%), with relatively small amounts of amino acids, urea, and uric acid (Regnault, 1987). Because ammonotelism is so dominant in aquatic gill-breathers, ammonia excretion rate typically has been used to evaluate the effects of various factors on total nitrogen excretion by crustaceans. But the importance of non-ammonia nitrogen excretion (amino acids, urea, etc.) has rarely been scrutinized. Adequate knowledge about nitrogen excretion by shrimp is required for successful design and operation of intensive production systems. Underestimation of total nitrogen excretion is potentially a serious problem, especially for closed recirculating water systems (Wickins, 1985). Quantification of the proportions of excretory products is also of importance in understanding effects of environmental factors on nitrogen metabolism of shrimp.
Nitrogen excretion rates of crustaceans vary with environmental conditions (Regnault, 1987). Significant effects of temperature and salinity on ammonia excretion have been documented for several commonly cultured species – Macrobrachium rosenbergii (Nelson et al., 1977; Stern et al., 1984), Fenneropenaeus indicus (Gerhardt, 1980), F.
chinensis (Chen and Lin, 1992, 1995), Marsupenaeus japonicus (Chen and Lai, 1993),
and Penaeus monodon (Chen et al., 1994). However, no comparable data have been reported for any species of the genus Litopenaeus, which includes the open-thelycum white shrimp Litopenaeus vannamei.
The purpose of this study was to determine nitrogenous excretory products and rates for L. vannamei juveniles exposed to various combinations of temperature and salinity under controlled experimental conditions. Temperature and salinity were within the normal ranges encountered in the culture of L. vannamei.
2. Materials and methods
2.1. Shrimp and acclimation
Shrimp Farms, Ltd. (Los Fresnos, TX, USA) and reared in the Nutrition Laboratory of the Shrimp Mariculture Project, Texas A&M University System, at 2561 ppt and 30618C. Shrimp used for the experiment were acclimated to nine combinations of three temperatures (24, 28, and 328C) and three salinities (10, 25, and 40 ppt). Water temperature was adjusted by decreasing or increasing values by 2–48C per day, and salinity was adjusted by decreasing or increasing values by 2–3 ppt per day until the desired levels were reached. Before the initiation of the excretion measurements, shrimp were kept at a constant temperature and salinity for 2–3 weeks to ensure complete acclimation. During the acclimation period, shrimp were fed three times daily. The feed used was prepared in the laboratory with 34% protein and 7.6% total lipid (Table 1). Photoperiod of 12-h light–12-h dark was maintained.
Table 1
Ingredients and proximate analysis of the experimental diet
Component Diet (%)
ICN Pharmaceuticals, Inc., Costa Mesa, CA, USA.
b
United States Biochenicals, Cleveland, OH, USA.
c
Inual, Santiago, Chile.
d
Zapata Haynei Corp., Reedville, VA, USA.
e
Sigma Chemical Company, Cleveland, OH, USA.
f
Riceland Foods Inc., Stuttgart, AR, USA.
g
Zapata Protein, Inc., Hammond, LA, USA.
h
Dawes Laboratories, Arlington Heights, IL, USA.
i
Roche Vitamins and Fine Chemicals, Pendergrass, GA, USA.
j
ICN Pharmaceuticals, Inc., Costa Mesa, CA, USA.
k
2.2. Experimental protocol
Filtered (5mm) and UV-sterilized seawater was used in the experiment. Salinity was adjusted by adding PHG evaporated salt (Gargill, Inc., MN, USA) or diluting with de-ionized water. Water temperatures were maintained by using water-bath tables
2
equipped with thermostat-controlled heaters. Polyethylene tanks (0.09 m bottom area), which had been thoroughly cleaned and contained 8 l of test seawater, were positioned in the water baths. Each individual tank was covered with a plastic lid to prevent shrimp escape and to reduce water evaporation. Water in each tank was gently aerated via a
21
single air-stone to keep DO above 6 mg l . In a preliminary experiment, an average 21
biomass about 1–1.5 g l was established as sufficient to produce an appropriate amount of nitrogen for accurate analyses, but not so great as to compromise shrimp performance during a 24-h interval.
Prior to each experimental trial, shrimp were starved for 12 h to ensure maximum gut evacuation. Shrimp of similar size (in the range of 3–5 g) were selected from holding tanks. To avoid metabolic fluctuation due to molt cycle (Stern and Cohen, 1982), only shrimp in the intermolt stage were selected for use. The molt stage was determined according to the methods of Smith and Dall (1985). Sexes were not separated since no previous study had ever indicated a significant difference in nitrogen excretion between male and female shrimp within the size range used in this study. A group of four or five shrimp was transferred into each test tank. Water samples were taken initially and after 24 h. Nitrogen analyses were conducted immediately after water samples were collected. After the trial, shrimp were gently dried on paper towels and weighed to the nearest 0.01 g by using a digital balance. Each shrimp was used only once in the experiment. The average weight and standard deviation for all tested shrimp was 3.8560.83 g, with no significant difference between treatments. There were two or three replicates of each treatment each time, and the excretion trial was repeated three times under the same conditions. In addition, to limit any other potential effects, such as bacterial activity, one tank without shrimp served as a control for each temperature–salinity combination. Water was sampled from the control chamber in the same manner as that from the experimental chambers; these data then were used as a basis for correction. To determine any ammonia loss into the atmosphere via aerial diffusion during the 24-h interval,
21
chambers initially containing water with 0.5mg-N l ammonia sulfate were processed with the same procedures as previously described. No significant loss was detected.
2.3. Measurements
2 2
Total ammonia-N (TAN), nitrite-N (NO ), and nitrate-N (NO ) were measured by2 3
methods adapted from Solorzano (1969) and Spotte (1979), from Strickland and Parsons (1972) and Spotte (1979), and from Mullin and Riley (1955) and Spotte (1979), respectively. Total dissolved nitrogen (TDN) was measured by the wet-oxidation method (Solorzano and Sharp, 1980; Parsons et al., 1984). Dissolved organic nitrogen (DON) was calculated based on the difference between TDN and ammonia-N1
nitrite-21 21
and DON were calculated by measuring the change in concentrations over the experimental period using the following formula:
R5[(Cf2C )i 3V ] /(W3t)
21
where C is the corrected final concentration of sample (f mg-N l ), C is the initiali
21
concentration of sample (mg-N l ), V is the water volume of the tank (l), W is the live weight of shrimp (g), and t is the time interval between initial and final water sampling (h).
The responses of metabolic rate of shrimp to temperature can be expressed as the thermal quotient (Q ). Q10 10 values for nitrogen excretion were calculated for each temperature interval at each salinity using the following formula:
10 / (T22T )1 Q105(R /R )2 1
where, R and R are metabolic rates of shrimp (in terms of ammonia excretion rate,2 1 RTAN) at temperatures T and T , respectively.2 1
2.4. Statistical analyses
Repeated measurements of nitrogen excretion resulted in both inter-treatment vari-ability and time-to-time variation. All data, after log transformation, were statistically analyzed by three-way analysis of variance (ANOVA), blocking by trial, to determine the effects of temperature and salinity on nitrogenous excretion rates. If significant differences were indicated at a 50.05, then Duncan’s multiple range test was used to resolve significant differences among treatments (Duncan, 1955). Further, regression analysis was used to model the relationships among nitrogen excretion rate, temperature, and salinity (SAS Institute, Inc., 1989–1996).
3. Results
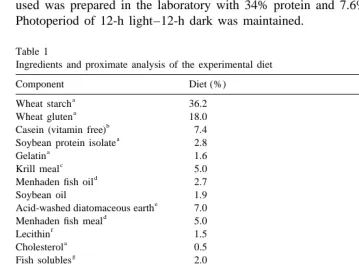
ANOVA indicated that there were significant effects of temperature and salinity on ammonia excretion (P,0.05), but the interaction between temperature and salinity was not significant (P.0.05). Ammonia excretion rate (RTAN) significantly increased with temperature. RTAN at 10 ppt was significantly higher than that at 25 or 40 ppt, with the difference between rates at 25 and 40 ppt not statistically significant (Fig. 1). The relationship between RTAN and temperature (T ) and salinity (S ) could be described as
0.062T 2 2
follows: RTAN54.54e 21.13S10.0205S (R 50.91, N59). The salinity for minimum RTANwas estimated to be 27.5 ppt. Relatively large variation was observed in the excretion rate of dissolved organic nitrogen (RDON), both among treatments and within the same treatment. Statistically significant effects of temperature and salinity on
RDON could not be detected (Fig. 1). The overall mean (6standard deviation) of RDON
21 21
was 5.24 (62.33) mg-N g h .
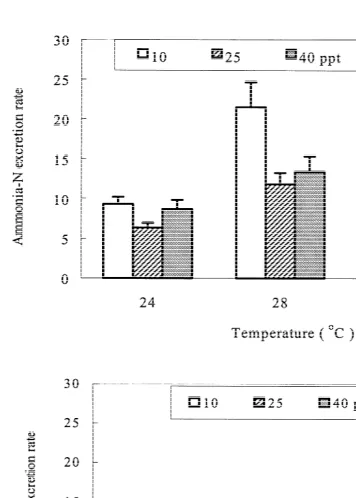
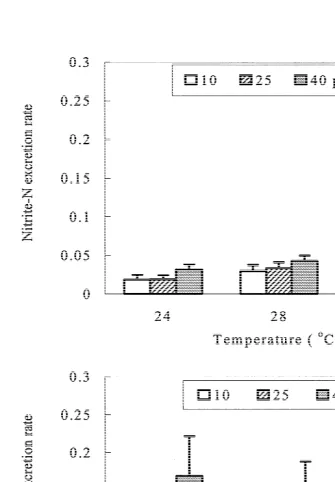
There was no significant interaction between temperature and salinity on either nitrite excretion rate (RNO2) or nitrate excretion rate (RNO2) of shrimp (Fig. 2). RNO2 ranged
21 21
21 21
21 21
from 0.019 to 0.043 mg-N g h ; it increased significantly with salinity and was significantly higher at 288C and 328C than at 248C. RNO2 ranged from 0.056 to 0.226
3
21 21
mg-N g h ; it was not influenced significantly by temperature, but increased significantly with salinity.
The excretion rate of total nitrogen (RTN) and the percentages of RTN comprised by ammonia-N, nitrite-N, nitrate-N, and DON are presented in Table 2. RTN was lowest
21 21 21 21
(10.03 mg-N g h ) at 248C and 25 ppt, and highest (30.24 mg-N g h ) at 328C and 40 ppt. A multiple regression analysis showed that RTN was significantly affected both by temperature and salinity, but not by temperature–salinity interaction. RTN
increased with temperature, with RTN at 28 and 328C significantly higher than at 248C.
RTNat 10 ppt was significantly higher than at 25 ppt, but the difference between 25 and 40 ppt was not significant. The relationship between RTN and temperature (T ) and
0.051T 2
salinity (S ) could be described by the equation: RTN58.10e 21.21S10.0228S
2
(R 50.91, N59). The salinity for minimum RTN was thus estimated to be 26.6 ppt. The results indicated that L. vannamei is strongly ammonotelic, with ammonia-N accounting for 61.9–84.3% of the total nitrogen excreted under the experimental conditions. Percentage ammonia-N in total nitrogen excreta (PTAN) was significantly influenced both by temperature and salinity. PTANincreased with temperature, with PTAN
at 28 and 328C significantly higher than at 248C. PTAN at 10 ppt was significantly higher
Table 2
21 21
Excretion rate for total nitrogen (RTNinmg-N g h ) and percentages (%) of RTNcomprised by ammonia-N,
a
nitrite-N, nitrate-N and DON for L. vannamei juveniles at various temperatures (T ) and salinities (S) T S RTN Percentage of total nitrogen excretion (%)
21 21
(8C) (ppt) (mg-N g h ) Ammonia-N Nitrite-N Nitrate-N DON
24 10 14.96 66.82 0.06 0.47 32.65
(2.32) (4.27) (0.013) (0.15) (4.40)
25 10.03 63.90 0.12 1.44 34.53
(1.06) (1.83) (0.016) (0.56) (1.69)
40 14.30 61.93 0.16 1.57 36.35
(2.30) (1.88) (0.021) (0.47) (1.63)
28 10 26.20 84.34 0.04 0.24 15.38
(2.17) (3.03) (0.011) (0.09) (2.97)
25 18.17 64.17 0.11 0.67 35.06
(3.01) (4.10) (0.010) (0.27) (4.38)
40 18.29 73.30 0.17 1.23 25.31
(3.66) (2.79) (0.026) (0.37) (2.94)
32 10 28.39 83.74 0.11 0.21 15.94
(4.78) (3.16) (0.032) (0.04) (3.21)
25 22.59 77.78 0.10 0.68 21.44
(3.34) (4.60) (0.018) (0.25) (4.66)
40 30.24 75.86 0.13 0.80 23.21
(2.66) (1.28) (0.016) (0.20) (1.36)
a
Table 3
Q10values for ammonia-N excretion rate (RTAN) of L. vannamei juveniles over two temperature ranges at each salinity
Salinity Q10calculated by RTAN
(ppt) 24–288C 28–328C
10 8.0 1.2
25 4.7 2.6
40 3.0 3.8
than that at 25 ppt, but the difference between 25 and 40 ppt was not significant (Table 2). The relationship between PTAN, temperature (T ), and salinity (S ) could be described
2 2
by the equation: PTAN538.9111.86T21.53S10.0254S (R 50.83, N59). Unlike RDON, the percentage DON in total nitrogen excretion (PDON), which ranged from 15.4 to 36.4%, was significantly affected by temperature and salinity. PDON
decreased with increased temperature; PDON at 28 and 328C were significantly lower than at 248C (P,0.05). PDONat 10 ppt was significantly lower than at 25 and 40 ppt, but the difference between 25 and 40 ppt was not significant. The relationship between
PDON, temperature (T ), and salinity (S ) could be described by the equation: PDON5
2 2
59.2621.79T11.46S20.0246S (R 50.81, N59).
There was only a small amount of total nitrogen excreted by L. vannamei as nitrite-N and nitrate-N. Percentages of nitrite (PNO2) and nitrate (PNO2) in total nitrogen excreta
2 3
ranged from 0.04 to 0.16% and from 0.24 to 1.56%, respectively.
The Q10 values for L. vannamei were determined at each salinity for two temperature ranges 24–288C and 28–328C (Table 3). In general, larger Q10 values were observed in the temperature range of 24–288C than in range of 28–328C, with an exception at salinity 40 ppt.
4. Discussion
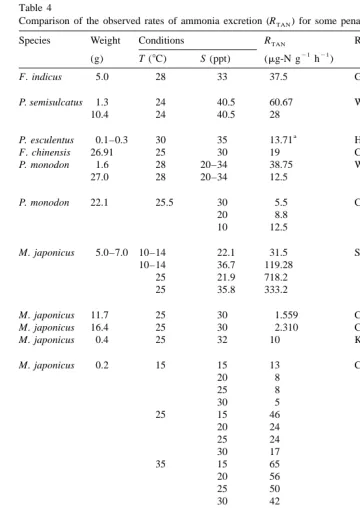
measuring the effect of various factors on nitrogen excretion (Regnault, 1987). There exists great variability in the published data for RTAN, even within the same species (Table 4). Part of variance results from variability in experimental methods and environmental conditions. In the present study, shrimp were fully adapted to the test media prior to testing. The data represent the physiological status of the shrimp at steady
21 21
state with ambient environment. RTANwas highest (23.26mg-N g h ) at 328C and 10
21 21
ppt and lowest (6.38 mg-N g h ) at 248C and 25 ppt. Since endogenous nitrogen excretion rate varies with shrimp species, size, molt stage, and environmental conditions (i.e., temperature and salinity), a direct comparison of absolute values between the results in the present study and those in the literature is difficult. However, the trend and relative magnitude of RTAN can be compared. Results obtained seem in general agreement with those reported by Gerhardt (1980); Wickins (1985); Chen et al. (1993); Koshio et al. (1993); Chen and Chen (1997).
The RTAN of crustaceans is affected both by intrinsic factors such as species, size, molt stage, and starvation and by external factors such as temperature, salinity, pH, DO, and light intensity. Effects of temperature and salinity on nitrogen excretion by crustaceans have been reviewed by Regnault (1987). Generally, the RTAN of shrimp increases with temperature due to a higher metabolic rate at elevated temperatures (Spaargaren et al., 1982; Chen and Lai, 1993; the present study).
The effect of salinity upon nitrogen excretion of crustaceans appears species-specific (Regnault, 1987). For penaeids, an increase in ammonia excretion frequently has been observed as salinity decreases. For M. japonicus, RTAN has been reported to increase with the decline of salinity in the range of 21–37 ppt (Spaargaren et al., 1982) and in the range of 15–30 ppt (Chen and Lai, 1993); for P. monodon, in the range of 15–35 ppt (Lei et al., 1989) and in the range of 10–30 ppt (Chen et al., 1994); and for F. chinensis, in the range of 10–30 ppt (Chen and Lin, 1995). In this study, RTAN increased for L.
vannamei as salinity decreased from 25 to 10 ppt. The reasons for that could be: (1) a
decrease in external osmotic concentration results in an increased metabolic rate of shrimp (Subramanian and Krishnamurthy, 1986; Chen and Lai, 1993); (2) at low salinity, shrimp are prone to use protein, not lipid, as their primary energy source (Chen, 1998); (3) at low salinity, the osmotic water inflow to shrimp increases, which is compensated by increased urine production to maintain water balance. To prevent losses
1 1 1
of the alkali ions (Na and K ), increased NH4 partially replaces alkali ions in the formation of urine (Spaargaren et al., 1982); and (4) a decreased concentration of free amino acids in the tissue (Lange, 1972; Dalla Via, 1986), and an increased catabolism of amino acids result in evaluated nitrogen excretion which is mainly ammonia (Chen and Kou, 1996). Also, it is noteworthy that RTAN of L. vannamei was lower at 25 ppt than at 40 ppt for all temperature levels, even though the difference was not statistically significant. Under hyper-osmotic conditions, more energy is required for osmoregulation when the ion gradient between body fluids and external medium is increased (Spaar-garen, 1975, 1976). Total nitrogen excretion of L. vannamei was lowest at 26.6 ppt
21
Table 4
Comparison of the observed rates of ammonia excretion (RTAN) for some penaeid species
Species Weight Conditions RTAN Ref.
21 21
(g) T (8C) S (ppt) (mg-N g h )
F. indicus 5.0 28 33 37.5 Gerhardt, 1980
P. semisulcatus 1.3 24 40.5 60.67 Wajsbrot et al., 1989
10.4 24 40.5 28
a
P. esculentus 0.1–0.3 30 35 13.71 Hewitt and Irving, 1990 F. chinensis 26.91 25 30 19 Chen et al., 1993 P. monodon 1.6 28 20–34 38.75 Wickins, 1985
27.0 28 20–34 12.5
P. monodon 22.1 25.5 30 5.5 Chen et al., 1994
20 8.8
10 12.5
M. japonicus 5.0–7.0 10–14 22.1 31.5 Spaargaren et al., 1982
10–14 36.7 119.28
25 21.9 718.2
25 35.8 333.2
M. japonicus 11.7 25 30 1.559 Chen and Cheng, 1993a M. japonicus 16.4 25 30 2.310 Chen and Cheng, 1993b M. japonicus 0.4 25 32 10 Koshio et al., 1993
M. japonicus 0.2 15 15 13 Chen and Lai, 1993
20 8
M. japonicus 0.3 27 18 33.25 Chen and Chen, 1997
26 24.17
34 19.75
L. vannamei 3.0–5.0 24 10 9.35 Present study
1976) and Subramanian and Krishnamurthy (1986), all of whom suggested that metabolic rates increased as salinity deviated from the isosmotic point because of increasing energy-cost due to osmoregulation.
Haberfield et al. (1975) observed that ammonia excretion of Carcinus maenas was affected by a salinity decrease while amino acid excretion rate did not change. Spaargaren et al. (1982) documented relative constancy in non-ammonia nitrogen excretion of M. japonicus at various temperature–salinity treatments. Similarly, Quarmby (1985) found that the organic nitrogen excretion of Pandalus platyceros was not affected by either temperature or salinity. In the present study, large variation was found in RDON, but a significant effect of temperature and salinity on RDON was not shown. However, percentage of DON in total nitrogen excretion was significantly affected by temperature and salinity. DON is mainly free amino acids and urea (Regnault, 1987). The release of amino acids which are metabolically useful substances is probably inadvertent and should be considered more as leakage than excretion (Dagg, 1976). In the present study, lower PDON was found at 28 and 328C than at 248C, and at 10 ppt than at 25 and 40 ppt. This suggests that the protein metabolism of L. vannamei is more efficient at high temperatures and low salinity.
Spaargaren (1985) reported that nitrate-N excretion of Carcinus maenas was higher at intermediate salinity (22.7–24.7 ppt) than at high salinity (38.5–42.5 ppt) or low salinity (4.7–15.1 ppt). Also, he suggested that nitrate formation may serve in the detoxification of ammonia and maintenance of hemolymph electroneutrality. Chen et al. (1994) reported both nitrite-N and nitrate-N excretion of P. monodon was higher at 20 ppt than at 10 or 30 ppt. Nitrite-N excretion increased when shrimp were exposed to ambient ammonia (Chen and Cheng, 1993b; Chen et al., 1994), apparently because the ammonia entering the hemolymph may be converted to nitrite under ammonia stress (Chen and Cheng, 1993b). The results of this study showed that, even though the excretion of both nitrite-N and nitrate-N was affected by temperature and salinity, only very small amounts of nitrogen were excreted by L. vannamei as nitrite and nitrate. For the conditions of the present study, nitrite-N and nitrate-N make up 0.04–0.16% and 0.24–1.56% of nitrogen excretion of L. vannamei, respectively. In contrast, nitrite-N and nitrate-N accounted for about 9.59–14.49% and 6.41–10.31%, respectively, of nitrogen excretion by P. monodon (Chen et al., 1994).
An experimental period of 24 h has often been used to determine excretion rates of aquatic animals. Such long periods can minimize the impact of high initial activity due to transferring shrimp into new tanks and the effect of light cycle on nitrogen excretion. However, bacterial activity during 24 h might introduce an experimental artifact in excretion studies (Armstrong et al., 1981). But, assuming that autotrophic bacteria take up ammonia-N while heterotrophic bacteria assimilate organic nitrogen, the apparent excretion rate of total nitrogen may not be affected much. Moreover, sterilized seawater and controls (without shrimp) were used in this study; thus, the effect of bacteria on results was not a large source of error.
Dietary factors also influence nitrogen excretion of shrimp. Regnault (1983) found that high-protein diets did not influence growth or body nitrogen storage of shrimp
excretion rate. Koshio et al. (1993) observed that post-feeding ammonia excretion of M.
japonicus increased with dietary protein content. They also observed that ammonia
excretion after 24-h starvation was constant, indicating that a 24-h starvation period eliminated differences in ammonia excretion rates due to variation in dietary protein content. In this study, the shrimp were fully acclimated to temperature and salinity, and had been starved over 12 h. The test tanks allowed shrimp to move freely but not actively swim, resulting in an uncontrolled but minimum motor activity. According to Brett and Groves (1979), who defined routine metabolism as the metabolic rate of fish during normal ‘‘spontaneous activity’’, the mean values of nitrogen excretion rate within a 24-h interval should represent the routine level of nitrogen metabolism of shrimp. In addition, it is known that in crustaceans protein and lipids appear to be the major source of energy (Barclay et al., 1983), while carbohydrates are not used as energy substrates during starvation (Regnault, 1981). Using P. esculentus, Hewitt and Irving (1990) found that carbohydrates and lipids were important energy substrates during the first 8–14 h post-feeding and that protein catabolism became increasingly significant in the pro-duction of energy thereafter. It is clear that nitrogen excreted by starving shrimp originates from the endogenous amino acids. Therefore, the rate of nitrogen loss from the endogenous pool in fasting L. vannamei due to excretion was estimated to be
21
0.24–0.73 mg-N g shrimp per day.
With intensification of shrimp farming, there is a significant increase in stocking density and harvest biomass. Accumulation of ammonia is a primary factor constraining shrimp biomass and stocking density in intensive systems because of its toxicity to shrimp. Using ammonia excretion rate together with a 96-h LC50 value, Wajsbrot et al. (1989) tried to estimate the appropriate biomass density of P. semisulcatus in shrimp ponds. But, effects of temperature and salinity should be taken into account because both ammonia toxicity and excretion rate of shrimp are influenced by temperature and salinity. The ‘‘safe level’’ of ammonia for L. vannamei has been reported to be 2.6 mg-N
21
l (Jiang et al., 1999). No attempt is made to calculate the maximum density here because it was not an objective of this research. However, the quantitative data obtained could be used for designing adequate biofiltration capacities in recirculating culture systems. Also, by measuring total nitrogen excretion instead of ammonia-N only, nitrogen loading from shrimp could be better estimated for more accurate nitrogen budgets and dynamic models.
Juvenile shrimp are adapted to estuarine salinities and coastal temperature regimes (Lester and Pante, 1992). Changes in ammonia excretion rate under the experimental conditions could reflect the responses of the shrimp metabolic rate to temperature and salinity. Thus, the thermal quotients (Q ) were calculated based on R10 TAN. However, it is recognized that this calculation may be inappropriate because the response of RTANto the temperature is unlikely monotonic. Larger Q10 values in the temperature range of 24–288C suggests that L. vannamei juveniles were more sensitive to temperature than at 28–328C, especially at low salinity (10 ppt). Coincidentally, in laboratory salinity-preference experiments at 26–328C, Mair (1980) demonstrated L. vannamei has a relatively low salinity preference (1–8 ppt). In nature, the abundance of postlarval L.
5. Conclusions
1. Ammonia is the major end-product of protein catabolism for L. vannamei, accounting for 61.9–84.3% of total nitrogen excretion.
2. Ammonia-N excretion rate (RTAN) was significantly affected by temperature and salinity. RTANincreased with temperature and was significantly higher at 10 ppt than at 25 and 40 ppt.
3. DON is the second major end-product, accounting for 15.4–36.4% of total nitrogen excretion.
4. The excretion rate of DON (RDON) was not significantly influenced by either
21 21
temperature or salinity, with an overall mean of about 5.24 mg-N g h .
5. Only very small amounts of nitrogen were excreted by L. vannamei as nitrite-N and nitrate-N.
6. The excretion rate of total nitrogen instead of ammonia is recommended for use in estimating the nitrogen load from shrimp in nitrogen budgets of production systems.
Acknowledgements
This research was funded in part by Project H-8158 of the Texas Agricultural Experiment Station and United States Department of Commerce Marine Shrimp Farming Program CSREES Grants No. 95-38808-1424 and No. 92-38808-6920. The publication is also a contribution of the Living Marine Resources Panel under the United States, Peoples’ Republic of China Marine and Fisheries Science and Technology Agreement Program. [SS]
References
Armstrong, D.A., Strange, K., Crowe, J., Knight, A., Simmons, M., 1981. High salinity acclimation by the prawn Macrobrachium rosenbergii: uptake of exogenous ammonia and changes in endogenous nitrogen compounds. Biol. Bull. 160, 349–365.
Barclay, M.C., Dall, W., Smith, D.M., 1983. Changes in lipid and protein during starvation and the molting cycle in the tiger prawn, Penaeus esculentus Haswell. J. Exp. Mar. Biol. Ecol. 68, 229–244.
Boyd, C.E., 1989. Water quality management and aeration in shrimp farming. In: Fisheries and Allied Aquacultures Departmental Series, Vol. No. 2. Alabama Agricultural Experiment Station, Auburn Universi-ty, Auburn, AL, USA, p. 83.
Brett, J.R., Groves, T.D.D., 1979. Physiological energetics. In: Hoar, W.S., Randall, D.J., Brett, J.R. (Eds.). Fish Physiology, Vol. III. Academic Press, Inc, New York, USA, pp. 280–352.
Castille, Jr. F.L., Lawrence, A.L., 1981. The effect of salinity on the osmotic, sodium and chloride concentrations in the hemolymph of euryhaline shrimp of the genus Penaeus. Comp. Biochem. Physiol. 68A, 75–80.
Chen, H.Y., 1998. Nutritional requirements of the black tiger shrimp: Penaeus monodon. Rev. Fish. Sci. 6, 79–95.
Chen, J.C., Chen, K.W., 1997. Oxygen uptake and ammonia-N excretion of juvenile Penaeus japonicus during depuration following 1-day exposure to different concentrations of saponin at different salinity levels. Aquaculture 156, 77–83.
Chen, J.C., Cheng, S.Y., 1993a. Haemolymph osmolality, acid–base balance and shift of ammonotelic to uerotellic excretory pattern of Penaeus japonicus exposed to ambient ammonia. Comp. Biochem. Physiol. 106C, 733–737.
Chen, J.C., Cheng, S.Y., 1993b. Urea excretion by Penaeus japonicus Bate exposed to different concentrations of ambient ammonia. J. Exp. Mar. Biol. Ecol. 173, 1–9.
Chen, J.C., Kou, C.T., 1996. Nitrogenous excretion in Macrobrachium rosenbergii at different pH levels. Aquaculture 144, 155–164.
Chen, J.C., Lai, S.H., 1993. Effects of temperature and salinity on oxygen consumption and ammonia-N excretion of juvenile Penaeus japonicus Bate. J. Exp. Mar. Biol. Ecol. 165, 161–170.
Chen, J.C., Lin, C.Y., 1992. Oxygen consumption and ammonia-N excretion of Penaeus chinensis juveniles exposed to ambient ammonia at different salinity levels. Comp. Biochem. Physiol. 102C, 287–291. Chen, J.C., Lin, C.Y., 1995. Response of oxygen consumption, ammonia-N excretion and urea-N excretion of
Penaeus japonicus Bate. J. Exp. Mar. Biol. Ecol. 165, 161–170.
Chen, J.C., Nan, F.H., Cheng, S.Y., Sheen, S.S., 1993. Effects of ambient ammonia on ammonia-N and protein concentrations in hemolymph and ammonia-N excretion of Penaeus chinensis. Mar. Ecol. Prog. Ser. 98, 203–208.
Dagg, M.J., 1976. Complete carbon and nitrogen budgets for the carnivorous amphipod Calliopus laeviusculus (Kroyer). Hydrobiol. 61, 297–357.
Dalla Via, G.J., 1986. Salinity responses of the juvenile penaeid shrimp Penaeus japonicus. II. Free amino acids. Aquaculture 55, 307–316.
Duncan, D.B., 1955. Multiple-range and multiple F test. Biometrics 11, 1–42.
Gerhardt, H.V., 1980. Nitrogen excretion by the juvenile prawn Penaeus indicus Milne Edwards at various temperatures. S. Afr. J. Sci. 76, 39–40.
Haberfield, E.C., Haas, L., Hamman, C.S., 1975. Early ammonia release by a polychaete Nereis virens and a crab Carcinus maenas in diluted seawater. Comp. Biochem. Physiol. 52A, 501–503.
Hewitt, D.R., Irving, M.G., 1990. Oxygen consumption and ammonia excretion of the brown tiger prawn Penaeus esculentus fed diets of varying protein content. Comp. Biochem. Physiol. 96A, 373–378. Holthuis, L.B., 1980. Shrimps and Prawns of the World. FAO Species Catalogue, Vol. 1. Food and Agriculture
Organization of the United Nations, Rome, Italy.
Huang, H.J., 1983. Factors Affecting the Successful Culture of Penaeus stylirostris and Penaeus vannamei at an Estuarine Power Plant Site: Temperature, Salinity, Inherent Growth Variability, Damselfly Nymph Predation, Population Density and Distribution, and Polyculture. Texas A&M University, College Station, TX, USA, Ph.D. Dissertation.
Jiang, D.H., Lawrence, A.L., Neill, W.E., Grant, W.H., Gong, H., 1999. Lethal effect of ammonia to postlarval Penaeus vannamei at two temperatures, 25 and 308C. In: American Aquaculture Society Annual Conference, 27–30 January, Tampa, Florida, USA, p. 77.
Koshio, S., Teshima, S., Kanazawa, A., Watase, T., 1993. The effect of dietary protein content on growth digestion efficiency and nitrogen excretion of juvenile kuruma prawns, Penaeus japonicus. Aquaculture 113, 101–114.
Kutty, M.N., Murugapoopathy, G., Krishnan, T.S., 1971. Influence of salinity and temperature on the oxygen consumption in young juveniles of the indian prawn Penaeus indicus. Mar. Biol. 11, 125–131. Lange, R., 1972. Some recent work on osmotic, ionic and volume regulation in marine animals. Rev.
Oceanograph. Mar. Biol. 10, 97–136.
Lei, C.H., Hsieh, L.Y., Chen, C.K., 1989. Effects of salinity on the oxygen consumption and ammonia-N excretion of young juveniles of the grass shrimp, Penaeus monodon Fabricus. Bull. Inst. Zool. Academia Sinica 28, 245–256.
Lester, L.J., Pante, M.J.R., 1992. Penaeid temperature and salinity responses. In: Fast, A.W., Lester, L.J. (Eds.), Marine Shrimp Culture: Principles and Practices. Elsevier, Amsterdam, pp. 515–534.
Martinez-Pakacios, C.A., Ross, L.G., Valenzuela, L.J., 1996. The effect of temperature and body weight on the oxygen consumption of Penaeus vannamei Boone, 1931. J. Aqua. Trop. 11, 59–65.
Mullin, J.D., Riley, J.P., 1955. The spectrophotometric determination of nitrate in natural waters, with particular reference to seawater. Anal. Chem. Acta. 12, 464–480.
Nelson, S.G., Armstrong, D.A., Knight, A.W., Li, H.W., 1977. The effects of temperature and salinity on the metabolic rate of juvenile Macrobrachium rosenbergii (Crustacea: Palaemonidae). Comp. Biochem. Physiol. 56A, 533–537.
Ogle, J.T., Beaugez, K., Lotz, J.M., 1992. Effects of salinity on survival and growth of postlarval Penaeus vannamei. Gulf Res. Rep. 8, 415–421.
Pante, M.J.R., 1990. Influence of Environmental Stress on the Heritability of Molting Frequency and Growth Rate of the Penaeid Shrimp, Penaeus vannamei. University of Houston-Clear lake, Houston, TX, USA, M.Sc. Thesis.
Parsons, T.R., Maita, Y., Lalli, C.M., 1984. A Manual of Chemical and Biological Methods for Seawater Analysis. Pergamon Press Inc, New York, USA.
Ponce-Palafox, J., Martinez-Palacios, C.A., Ross, L.G., 1997. The effects of salinity and temperature on the growth and survival rate of juvenile white shrimp, Penaeus vannamei, Boone, 1931. Aquaculture 157, 107–115.
Quarmby, L.M., 1985. The influence of temperature and salinity on the nitrogenous excretion of the spot prawn, Pandalus platyceros Brandt. J. Exp. Mar. Biol. Ecol. 87, 229–239.
Regnault, M., 1981. Respiration and ammonia excretion of the shrimp Crangon crangon L.: metabolic response to prolonged starvation. J. Comp. Physiol. 141, 549–555.
Regnault, M., 1983. Influence a long terme du taux proteique du regime sur l’excretion d’azote et le metabolism de la crevette Crangon cragon L. Oceanis. 9, 241–255.
Regnault, M., 1987. Nitrogen excretion in marine and fresh-water crustacea. Biol. Rev. 62, 1–24.
SAS Institute Inc, Inc., 1989–1996. SAS System for Windows. Release 6.12. SAS Institute Inc, Cary, NC, USA.
Smith, D.M., Dall, W., 1985. Moult-staging the tiger prawn, Penaeus esculentus Haswell. In: Rothlisberg, P.C., Hill, B.J., Staples, D.J. (Eds.), Second Australian National Prawn Seminar, NPS2, Cleveland, Qld., Australia, pp. 85–93.
Solorzano, L., 1969. Determination of ammonia in natural waters by the phenolhypochlorite method. Limnol. Oceanogr. 14, 799–801.
Solorzano, L., Sharp, J.H., 1980. Determination of total dissolved nitrogen in natural waters. Limnol. Oceanogr. 25, 751–754.
Spaargaren, D.H., 1975. Energy relations in the ion regulation in three crustacean species. Comp. Biochem. Physiol. 51A, 543–548.
Spaargaren, D.H., 1976. A comparative study on the regulation of osmotic, ionic and organic solute concentrations in the blood of aquatic organism. Comp. Biochem. Physiol. 53A, 31–40.
Spaargaren, D.H., 1985. The significance of nitrate in the nitrogenous excretion of Carcius maenas. Neth. J. Sea Res. 19, 119–124.
Spaargaren, D.H., Richard, P., Ceccaldi, H.J., 1982. Excretion of nitrogenous products by Penaeus japonicus Bate in relation to environmental osmotic conditions. Comp. Biochem. Physiol. 72A, 673–678. Spotte, S., 1979. Fish and Invertebrate Culture: Water Management in Closed Systems, 2nd Edition. John
Wiley and Sons, New York, USA.
Stern, S., Brout, A., Cohen, D., 1984. The effect of salinity and ion composition on oxygen consumption and nitrogen excretion of Macrobrachium rosenbergii (De Man). Comp. Biochem. Physiol. 79A, 271–274. Stern, S., Cohen, D., 1982. Oxygen consumption and ammonia excretion during the molt cycle of the
freshwater prawn Macrobrachium rosenbergii (De Man). Comp. Biochem. Physiol. 73A, 417–419. Stern, S., Daniels, H., Letellier, E., 1990. Tolerance of post larvae and juvenile Penaeus vannamei to low
salinity. In: World Aquaculture 90, Halifax, Nova Scotia, Canada, T30. 12. National Research Council Canada, Ottawa, Canada, Abstract.
Strickland, J.D.H., Parsons, T.R., 1972. In: A Practical Handbook of Seawater Analysis, 2nd Edition. Fish. Res. Board Canada, Ottawa, Canada, p. 310, Bull. 167.
Villarreal, H., Hinojosa, P., Naranjo, J., 1994. Effect of temperature and salinity on the oxygen consumption of laboratory produced Penaeus vannamei postlarvae. Comp. Biochem. Physiol. 108A, 331–336.
Wajsbrot, N., Krom, M.D., Gasith, A., Samocha, T., 1989. Ammonia excretion of green tiger prawn Penaeus semisulcatus as a possible limit on the biomass density in shrimp pond. Bamidgeh 41, 159–164. Wickins, J.F., 1985. Ammonia production and oxidation during the culture of marine prawns and lobsters in
laboratory recirculation systems. Aquacult. Eng. 4, 155–174.