2 PENGARUH PEMUPUKAN NITROGEN TERHADAP
PERTUMBUHAN BEBERAPA TANAMAN HIJAUAN
MENJALAR UNTUK SISTEM PASTURA-HUTAN
Pendahuluan
Latar belakang
Berbagai faktor dibutuhkan untuk pertumbuhan tanaman seperti cahaya, CO2, air dan unsur hara mineral (Marschner 1999). Nitrogen merupakan unsur hara utama yang dibutuhkan seluruh tanaman untuk pertumbuhan dan produksi yang optimum. Ketidakcukupan atau kekurangannya akan menyebabkan terhentinya pertumbuhan dan menunjukkan gejala defisiensi unsur hara (Ipinmoroti et al. 2008).
Nitrogen di dalam tanah terdapat dalam berbagai bentuk yang berbeda seperti senyawa nitrogen organik, ion ammonium (NH4+), dan ion nitrat (NO3-) (Eckert 2009; Marx et al. 1999), yang berinteraksi antara satu dengan lainnya dan dengan tanaman, ternak, dan mikroorganisme. Kebanyakan tanaman menggunakan nitrogen dengan cepat. Tanaman menyerap nitrogen dalam bentuk ion nitrat (NO3-) atau amonium (NH4+), yang keduanya merupan ion yang larut dalam air. Ion nitrat merupakan ion yang larut dalam air. Ion nitrat diserap dengan cepat oleh akar tanaman tetapi tercuci dengan mudah dari tanah dengan adanya curah hujan yang tinggi atau irigasi berlebihan (CFF 2009; Marx et al. 1999). Ion ammonium terjerap dalam partikel tanah dan bergerak dengan lambat melalui akar tanaman (CFF 2009). Nitrogen total tidak menunjukkan ketersediaan nitrogen tanaman (Marx et al. 1999). Kandungan nitrogen yang dibutuhkan untuk pertumbuhan yang optimum bervariasi antara 2 dan 5% dari berat kering tanaman, bergantung pada spesies tanaman, fase perkembangan tanaman dan bagian organ tanaman (Marschner 1999).
Kebanyakan tanaman mengambil nitrogen dari tanah secara berkelanjutan dalam daur hidupnya dan kebutuhan nitrogen biasanya meningkat dengan meningkatnya ukuran tanaman. Tanaman yang mempunyai ketersediaan nitrogen yang cukup akan tumbuh dengan cepat dan menghasilkan sejumlah sukulen, bagian tanaman yang hijau (Eckert 2009). Penambahan pupuk N umumnya
meningkatkan rasio akar-tajuk (Barbar 1984). Rasio akar-tajuk merupakan rasio berat akar terhadap tajuk suatu tanaman. Tanaman dengan proporsi akar yang lebih besar dapat berkompetisi lebih efektif untuk mendapatkan unsur hara tanah, sedangkan tanaman dengan proporsi tajuk yang lebih besar dapat mengumpulkan lebih banyak energi (Allaby 2004). Terdapat suatu karakteristik rasio tajuk-akar untuk setiap spesies pada setiap fase pertumbuhan. Rasio tajuk-akar cenderung meningkat dengan meningkatnya ukuran tanaman (menurunkan akar tanaman), mencerminkan meningkatnya asimilasi bagian atas tanaman. Rasio tajuk-akar dipengaruhi oleh kondisi lingkungan seperti cahaya, ketersediaan unsur hara, suhu dan ketersediaan air. Perubahan ini biasanya mencerminkan keuntungan adaptasi dari tanaman dalam menyesuaikan diri pada sumber yang terbatas (Marschner 1999)
Beberapa spesies rumput dan legum mempunyai keuntungan secara morfologi tumbuh di bawah kondisi ternaungi. Stür (1998) menyatakan bahwa spesies hijauan yang berhasil tumbuh dalam tanaman perkebunan harus beradaptasi tidak hanya dengan rendahnya level cahaya tetapi juga dengan iklim (curah hujan, suhu, variasi panjang hari), tanah (pH, kesuburan, tekstur dan drainase) dan manajemen (cekaman defoliasi atau penggembalaan dan input pupuk). Rendahnya produksi potensial seluruh spesies pada kondisi cahaya rendah merupakan hambatan utama terhadap produktivitas sistem tanaman pohon-pakan dengan adanya penutupan kanopi sejalan dengan pertambahan umur tanaman pohon (Wong 1991; Addison 2003). Beberapa penelitian telah menemukan beberapa spesies rumput dan legum yang mampu beradaptasi terhadap naungan, mulai dari naungan rendah sampai sangat tinggi, seperti Brachiaria humidicola, Arachis pintoi, Axonopus compressus and Paspalum sp (Reynolds 1995; Wong, 1991).
Respon produksi beberapa spesies hijauan tropis terhadap naungan menggambarkan kuatnya hubungan antara produktivitas dan cahaya, tetapi lebih sering juga digabungkan dengan ketersediaan nitrogen tanah (Wong 1991). Unsur hara harus ada di dalam tanah dan tersedia melalui sumber yang sesuai dalam jumlah yang cukup dan dalam bentuk yang siap digunakan oleh tanaman dalam mencapai produksi yang tinggi. Nitrogen merupakan unsur hara utama yang dibutuhkan oleh seluruh tanaman untuk pertumbuhan dan produksi. (Ipinmoroti et
al. 2008) dan nitrogen secara normal merupakan unsur hara yang sangat memengaruhi untuk pertumbuhan tanaman yang optimum (Espinoza et al. 2007).
Beberapa spesies hijauan toleran naungan diteliti responnya dalam hal karakteristik morfologi dan produksi berat kering terhadap pemupukan nitrogen. Perbedaan pola pertumbuhan tanaman menyebabkan perbedaan manajemen dan strategi dalam pengembangannya. Informasi ini berguna untuk mendesain komponen sistem yang kompatibel untuk meningkatkan produktivitas sistem integrasi tanaman pohon-pakan.
Bahan dan Metode
Waktu dan tempat
Penelitian ini dilakukan dalam skala rumah kaca mulai dari Agustus 2010 sampai dengan Desember 2010. Penelitian dilakukan di dalam rumah plastik didesain dengan konstruksi bamboo. Atap plastik dilapisi dengan net plastik untuk menciptakan kondisi naungan, dengan dinding yang separuh tertutup (separuh dari bagian bawah ditutup dengan plastik), separuh bagian atas ditutup dengan net plastik. Kondisi ini memungkinkan sirkulasi udara yang akan mengurangi panasnya rumah plastik. Temperatur minimum dan maksimum selama penelitian berkisar 24°C sampai dengan 34°C dengan kelembaban minimum dan maksimum berkisar 67% sampai 89%.
Bahan dan alat
Dalam penelitian ini digunakan beberapa spesies tanaman hijauan menjalar dan nitrogen (bersumber dari urea) sebagai pupuk. Alat yang digunakan dalam penelitian ini pot bulat datar dengan diameter 43 cm dan tinggi 12 cm, gunting, pisau dan thermohygrometer digital.
Penelitian ini didesain dengan menggunakan rancangan acak lengkap dengan pola faktorial 4 spesies, 2 level pemupukan dengan 3 kali ulangan. Spesies tanaman hijauan menjalar (stoloniferous) yang digunakan adalah PN = Paspalum notatum, BH = Brachiaria humidicola, AC = Axonopus compressus dan AP = Arachis pintoi. Level pemupukan nitrogen/urea, yaitu (-N) = tanpa pupuk dan (+N) = 300 Kg urea/ha.
Prosedur penelitian
Tanaman hijauan di tanam di tengah pot bulat datar yang berisikan 5 kg tanah. Bahan tanam berasal dari anakan untuk rumput dan stek untuk legum. Pemupukan nitrogen diaplikasikan 3 hari setelah tanam. Pemupukan dilakukan 2 kali; 150 kg urea/ha diberikan pada 3 hari setelah tanam dan pemupukan kedua diberikan pada bulan awal kedua pertumbuhan tanaman. Selama penelitian tanaman disiram setiap hari. Komposisi unsur hara hasil analisis tanah sebelum penanaman adalah
pH 4.5 – 6.5, 0.10% nitrogen, 84 mg/100g P2O5 dan 10 mg/100g K2O (BPT 2010). Hasil analisis tanah awal ini mengindikasikan bahwa kondisi tanah sebelum penanaman rendah akan nitrogen dan kalium, namun akan fosfor, dengan pH yang sedikit masam ke netral (acuan Tabel 1).
Tablel 1 Kriteria penilaian sifat-sifat kimia tanah
Elemen
Kisaran Sangat
rendah Rendah Sedang Tinggi Kelebihan
N (%) <0.10 0.10 - 0.20 0.21 – 0.50 0.51 – 0.75 > 0.75 P2O5(HCl 25%) (mg/100g) <15.0 15.0 – 20.0 21.0 – 40.0 41.0 – 60.0 > 60.0 K2O (HCl 25%) (mg/100g) <10 10.0 – 20.0 21.0 – 40.0 41.0 - 60.0 > 60.0 Sangat masam Cukup masam Sedikit masam Netral Cukup alkalin pH (H2O) < 4.5 4.5 – 5.5 5.6 -6.5 6.6 – 7.5 7.6 – 8.5 Sumber: BPT (2005)
Peubah yang diamati
Pengamatan pertumbuhan tanaman selama 90 hari meliputi:
a. Panjang tanaman (cm/tanaman); dengan mengukur tanaman mulai dari permukaan tanah sampai ujung daun paling panjang dan diukur pada akhir penelitian.
b. Pertambahan panjang tanaman (cm/tanaman); diukur dengan menghitung perbedaan pengukuran pengamatan minggu satu dengan pengamatan minggu berikutnya.
c. Jumlah daun (helai/tanaman); diukur pada akhir penelitian.
d. Bobot kering tajuk tanaman (g/tanaman); bobot kering akar tanaman dengan mengeringkan sampel dalam oven 80°C selama 48 jam. Kemudian sampel kering digiling untuk analisis N tajuk (metode Kjeldahl).
e. Rasio tajuk-akar.
f. Senescense; bagian tanaman yang menua dan jatuh di permukaan tanah. g. Status N, P dan K tanah.
Analisis data
Data yang diperoleh dianalisis sidik ragam berdasarkan RAL faktorial. Perbedaan antar perlakuan dianalisis dengan Uji Beda Nyata Terkecil (Steel dan Torrie 1989).
Hasil
Pertumbuhan tanaman
Tiga bulan periode penelitian tercatat bahwa pertumbuhan spesies hijauan menjalar tidak nyata dipengaruhi oleh pemupukan, tetapi pertumbuhan hijauan menjalar berbeda secara nyata menurut spesies. Beberapa pot penelitian, khususnya pada P. notatum, diduga menderita ketidakcukupan akan unsur hara tertentu. Hal ini ditunjukkan oleh warna kuning kecoklatan pada bagian ujung daun pada minggu ke-6 periode penelitian. Berdasarkan pengamatan, P. notatum menunjukkan pertumbuhan yang paling rendah dibandingkan dengan spesies lainnya.
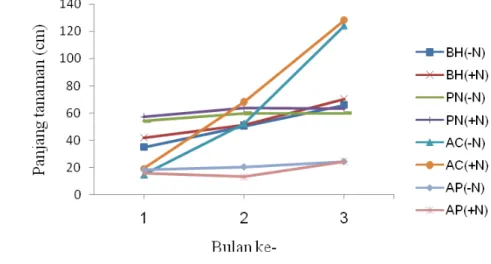
Tinggi bibit hijauan menjalar pada saat awal penelitian adalah 5 cm di atas permukaan tanah. Pada bulan pertama, pertumbuhan seluruh tanaman menunjukkan adanya peningkatan pada panjang tanaman, akan tetapi pada bulan kedua, A. pintoi menunjukkan pertumbuhan yang menurun. A. compressus pada perlakuan (-N) dan (+N) rata-rata mencapai panjang 126.8 cm. Pada bulan ketiga, kebanyakan spesies menunjukkan peningkatan pertambahan panjang, kecuali P. notatum cenderung menurun (Gambar 1).
Gambar 1 Tren pertumbuhan panjang spesies hijauan menjalar selama 3 bulan penelitian (PN = P. notatum, BH = B. humidicola, AC = A.
compressus dan AP = A. pintoi; (-N) = tanpa pupuk dan (+N) = 300 kg
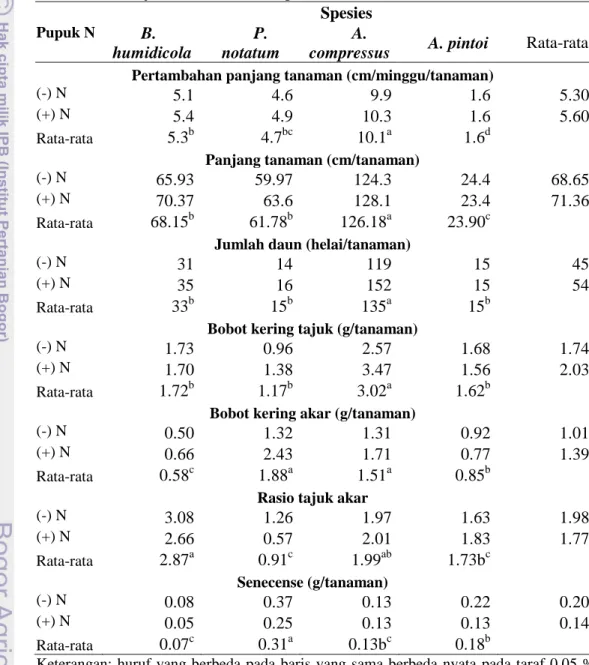
Tabel 2 menunjukkan beberapa peubah pertumbuhan tanaman hijauan menjalar selama 3 bulan periode penelitian. Penelitian ini menemukan bahwa terdapat pengaruh yang sangat nyata dari spesies terhadap pertumbuhan.
Tabel 2 Pertambahan panjang, panjang tanaman, jumlah daun, bobot kering tajuk dan akar, rasio tajuk-akar dan senescense tanaman hijauan menjalar selama 3 bulan penelitian
Pupuk N Spesies B. humidicola P. notatum A.
compressus A. pintoi Rata-rata Pertambahan panjang tanaman (cm/minggu/tanaman)
(-) N 5.1 4.6 9.9 1.6 5.30
(+) N 5.4 4.9 10.3 1.6 5.60
Rata-rata 5.3b 4.7bc 10.1a 1.6d
Panjang tanaman (cm/tanaman)
(-) N 65.93 59.97 124.3 24.4 68.65
(+) N 70.37 63.6 128.1 23.4 71.36
Rata-rata 68.15b 61.78b 126.18a 23.90c
Jumlah daun (helai/tanaman)
(-) N 31 14 119 15 45
(+) N 35 16 152 15 54
Rata-rata 33b 15b 135a 15b
Bobot kering tajuk (g/tanaman)
(-) N 1.73 0.96 2.57 1.68 1.74
(+) N 1.70 1.38 3.47 1.56 2.03
Rata-rata 1.72b 1.17b 3.02a 1.62b
Bobot kering akar (g/tanaman)
(-) N 0.50 1.32 1.31 0.92 1.01
(+) N 0.66 2.43 1.71 0.77 1.39
Rata-rata 0.58c 1.88a 1.51a 0.85b
Rasio tajuk akar
(-) N 3.08 1.26 1.97 1.63 1.98 (+) N 2.66 0.57 2.01 1.83 1.77 Rata-rata 2.87a 0.91c 1.99ab 1.73bc Senecense (g/tanaman) (-) N 0.08 0.37 0.13 0.22 0.20 (+) N 0.05 0.25 0.13 0.13 0.14 Rata-rata 0.07c 0.31a 0.13bc 0.18b
Keterangan: huruf yang berbeda pada baris yang sama berbeda nyata pada taraf 0.05 % dengan uji BNT. (-N) = tanpa pupuk dan (+N) = 300 Kg urea/ha
Dari Tabel 2 terlihat A. compressus secara nyata menunjukkan pertumbuhan yang terbaik dibandingkan spesies lainnya dalam hal pertambahan panjang
tanaman per minggu, panjang tanaman, jumlah daun, bobot kering tajuk dan akar. Semakin panjang tanaman A. compressus, maka semakin banyak buku terbentuk yang menjadikan tanaman ini mempunyai jumlah daun yang banyak. Daun tumbuh pada setiap buku baru. Pertumbuhan bagian atas tanaman yang baik pada A. compressus juga diikuti dengan pertumbuhan akar yang baik. Meskipun perlakuan pemberian pupuk tidak memberikan pengaruh yang nyata, namun tanaman A. compressus yang diberi pupuk terlihat mempunyai akar yang lebih banyak (Gambar 2). Kondisi ini diikuti oleh B. humidicola, P. notatum, and A. pintoi. Stolon dari A. compressus menyebar dengan cepat dan jika masih terdapat area untuk bertumbuh, tanaman ini akan menutupinya.
Gambar 2 Struktur akar tanaman hijauan menjalar (-N = tanpa pupuk dan +N = 300 kg Urea/ha)
P. notatum menghasilkan bobot kering akar dan senescence yang tertinggi yaitu masing-masing 1.88 dan 0.33 g/tanaman, sedangkan yang terendah ditunjukkan oleh B. humidicola yaitu masing-masing sebesar 0.58 dan 0.07 g/tanaman. Daun tua dan mati disepanjang pertumbuhan tanaman merupakan daun yang berasal dari sisa pemotongan bahan tanam awal (Gambar 3).
Gambar 3 Daun menua dan mati sisa dari pemotongan awal dari bahan tanam (dalam lingkaran). (PN = P. notatum, BH = B. humidicola, AC = A. compressus dan AP = A. pintoi)
Status N, P dan K tanah dan serapan N
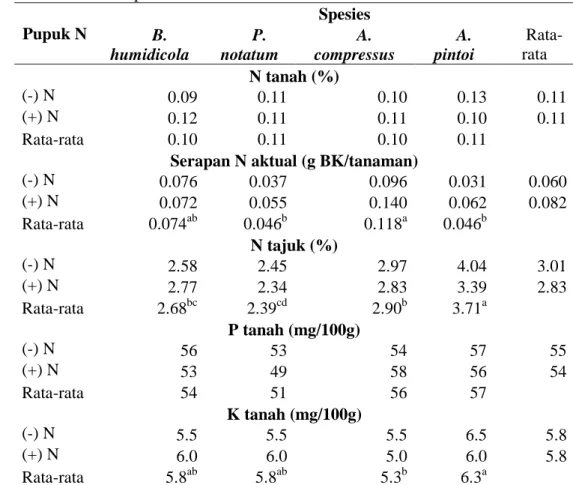
Serapan hara N oleh tanaman hijauan menjalar dan status unsur hara tanah pada pot penelitian setelah penelitian dapat dilihat pada Tabel 3. Dari Tabel 3 di atas terlihat bahwa hasil analisis tanah pada akhir penelitian untuk kandungan nitrogen tanah terlihat sama dengan sebelum aplikasi nitrogen, tetapi tidak untuk konsentrasi P dan K tanah. Penurunan konsentrasi P dan K selama 3 bulan periode penelitian dari nilai yang tinggi menjadi rendah. Hal ini menunjukkan bahwa tanaman menggunakan N, P dan K dari dalam tanah untuk pertumbuhannya. Hijauan pakan menjalar mengunakan P dalam proporsi yang hampir sama, sehingga konsentrasi P tidak nyata berbeda diantara spesies. Konsentrasi kalium pada A. pintoi terlihat yang paling tinggi. Konsentrasi N tajuk yang paling tinggi terlihat pada A. Pintoi dan diikuti oleh A. compressus, yaitu masing-masing 3.71 dan 2.9%. Kandungan N tajuk dan K tanah tertinggi sampai terendah terlihat pada spesies legum A. pintoi, A. compressus, B. humidicola dan P. notatum. Serapan N tertinggi sampai terendah terlihat pada A. compressus, B. humidicola, P. notatum dan A. pintoi.
Tabel 3 N-tanah, serapan N aktual, N-tajuk dan kandungan P and K tanah pada akhir penelitian Pupuk N Spesies B. humidicola P. notatum A. compressus A. pintoi Rata-rata N tanah (%) (-) N 0.09 0.11 0.10 0.13 0.11 (+) N 0.12 0.11 0.11 0.10 0.11 Rata-rata 0.10 0.11 0.10 0.11
Serapan N aktual (g BK/tanaman)
(-) N 0.076 0.037 0.096 0.031 0.060 (+) N 0.072 0.055 0.140 0.062 0.082 Rata-rata 0.074ab 0.046b 0.118a 0.046b N tajuk (%) (-) N 2.58 2.45 2.97 4.04 3.01 (+) N 2.77 2.34 2.83 3.39 2.83 Rata-rata 2.68bc 2.39cd 2.90b 3.71a P tanah (mg/100g) (-) N 56 53 54 57 55 (+) N 53 49 58 56 54 Rata-rata 54 51 56 57 K tanah (mg/100g) (-) N 5.5 5.5 5.5 6.5 5.8 (+) N 6.0 6.0 5.0 6.0 5.8 Rata-rata 5.8ab 5.8ab 5.3b 6.3a
Keterangan: huruf yang berbeda pada baris yang sama berbeda nyata pada taraf 0.05 % dengan uji BNT. (-N) = tanpa pupuk dan (+N) = 300 Kg urea/ha
Pembahasan
Pertumbuhan tanaman
Produksi tajuk merupakan gabungan panjang tanaman dan jumlah daun. Sejalan dengan panjang tanaman dan jumlah daun, bobot kering tajuk dan akar
A.compressus terlihat paling tinggi dibandingkan dengan spesies lainnya seperti B. humidicola, P. notatum dan A. pintoi yang terlihat sama.
Jumlah daun yang paling rendah terdapat pada P. notatum dan A. pintoi dengan rata-rata 15 helai daun masing-masingnya selama 3 bulan penelitian (Tabel 2). Pakiding (1998) menemukan tidak ada pengaruh terhadap jumlah daun per anakan dengan meningkatnya level nitrogen dan tinggi pemotongan. Hirata (2000) juga menemukan level nitrogen mempunyai pengaruh yang kecil terhadap laju pemunculan daun (LAR) dari P. notatum, LAR sedikit meningkat dengan meningkatnya level nitrogen.
Anakan merupakan sumber potensial bagi produksi jumlah daun. Anakan yang banyak tumbuh, mengakibatkan jumlah daun semakin meningkat. Jumlah daun yang rendah pada P. notatum disebabkan oleh rendahnya respon P. notatum terhadap pemupukan nitrogen. Anakan pertama dari P. notatum (+N) muncul sekitar minggu ke-10 dari periode penelitian. Hirata (1993) melaporkan bahwa jumlah anakan rumput Bahia membutuhkan waktu 3 bulan untuk menunjukkan respon yang nyata terhadap tinggi pemotongan. Respon pembentukan anakan dapat dipengaruhi oleh faktor seperti genotip, suhu, intensitas cahaya, ketersediaan air, nutrisi mineral, photoperiod dan pengatur pertumbuhan. Penelitian lebih jauh dari rumput Bahia oleh Hirata and Pakiding (2001) menemukan bahwa level N memberikan pengaruh yang rendah terhadap jumlah anakan dan anakan anak, laju pemunculan daun (LAR), laju daun yang mati (LDR) dan yang jatuh, kecuali untuk anakan pada rumput Bahia pada musim anakan aktif (Mei – Juni). Kepadatan anakan rumput Bahia sangat stabil karena tingginya longevitas anakan (rendah laju kematian anakan) disamping juga rendahnyalaju pemunculan anakan(TAR). Relatif TAR yang rendah pada rumput Bahia disebabkan oleh rendahnya laju pengisian anakan. Islam dan Hirata (2005) bahwa melaporkan pertumbuhan anakan anak pada axil daun adalah nihil atau
sangat jarang terjadi pada rumput centipede (Eremochloa ophiuroides)dan Bahia pada pertumbuhan daun muda pertama sampai ke-tiga. Aktivitas pembentukan anakan pada rumput Bahia juga nihil atau sangat rendah ketika pertumbuhan daun muda ke-empat dan ke-enam, kecuali pada bulan Mei dan Juni, ketika pembentukan anakan terjadi dengan level pemupukan nitrogen yang tinggi. Bogdan (1977) menyatakan rumput Bahia tumbuh mencapai tinggi maksimum antara 15-70 cm (kira-kira 6-28 inchi), terkadang mencapai tinggi 100 cm (sekitar 40 inchi)
Pada spesies B.humidicola (-N) dan A. compressus (+N), pemunculan anakan pertama terjadi pada minggu pertama periode penelitian. A. compressus menyebar dengan cepat melalui stolon dan rhizome di bawah kondisi yang memungkinkan dibandingkan Axonopus affinis (FAO). Manidool (1992) menyatakan bahwa tanaman muda dari A. compressus mulai tumbuh dalam jalur melingkar. Dengan sedikit kompetisi, jalur dapat mencapai sampai diameter 1 meter dalam satu musim. Tanaman ini tumbuh seperti gulma dan membentuk hamparan rumput yang padat. Wong (1991) menambahkan bahwa spesies indigenous tahan naungan seperti Axonopus compressus, Stenotaphrum secundatum, Brachiaria miliiformis dan Paspalum conjugatum menunjukkan lebih persisten dan produktif di bawah kondisi cahaya rendah.
Menurut Abdullah (2009) jumlah anakan B. humidicola (Rendle) Schweick bergantung pada jumlah buku dan panjang stolon, sehingga pola pertumbuhan anakan mengikuti pola dinamik keduanya. Pada penelitian ini panjang stolon dari B. humidicola dan P. notatum nyata lebih rendah dibandingkan dengan A. compressus.
Panjang tanaman dan jumlah daun merupakan sumber potensial bagi fotosintesis tanaman. Lebih banyak daun, lebih luas area untuk fotosintesis. Dengan demikian akan menghasilkan lebih banyak fotosintat untuk akumulasi produksi bagian atas tanaman. Ditambah lagi, pertambahan biomassa bagian atas tanaman yang tinggi, ditunjang oleh pertumbuhan struktur akar yang baik.Gastal dan Durand (1999) menyatakan bahwa sintesis struktur tajuk akar bergantung pada keseimbangan antara permintaan morphogenetik dan ketersediaan substrat karbohidrat. Permintaan morphogenetik ditentukan oleh potensi morphogenetik
tanaman (pertumbuhan potensial daun, batang, akar, anakan dan organ reproduksi), di bawah kontrol genom dan di bawah pengaruh ekspresinya yang diakibatkan faktor lingkungan (suhu, kualitas cahaya). Gastal dan Durand (1999) juga menambahkan fotosintesis kanopi bergantung pada luas area daun yang menghasilkan perluasan daun. Davidson dan Robson (1985) menyatakan laju fotosintesis yang lebih tinggi dari Clover dapat dihasilkan dari tingginya potensial fotosintesis daun atau lebih besarnya intersepsi cahaya.
Pembentukan anakan di bawah suplai nitrogen sedang terlihat tertekan pada rumput pendek, akan tetapi pemanjangan daun tetap terjadi meskipun lajunya mengalami penurunan (Nelson 2000). Hirata (2000) juga mengamati bahwa pemanjangan daun terjadi secara cepat ketika masih daun pertama dan ke-dua tetapi berhenti memanjang ketika daun ke-empat. Hirata et al. (2009) menemukan bahwa panjang anakan yang paling tinggi terjadi pada pemotongan yang tinggi pada periode musim panas dengan nitrogen yang rendah, sedang dan tinggi yaitu masing-masing 399, 396, 365 mm.
Meningkatnya panjang tanaman, jumlah daun dan biomassa berat kering bagian atas tanaman mengakibatkan tingginya rasio tajuk-akar pada A. compressus, meskipun berbeda tidak nyata dengan B.humidicola. Rasio tajuk-akar cenderung meningkat dengan meningkatnya ukuran tanaman (menurun untuk akar), mencerminkan meningkatnya asimilat bagian atas tanaman (Marschner 1999).
Davidson (1994) menemukan bahwa di dalam akar terdapat spektrum yang luas dari respon berbagai komponen morphogenetik terhadap nitrogen. Terdapat kisaran respon yang sangat luas antara spesies. Wong (1991) melaporkan meningkatnya berat kering terhadap komponen daun yang disebabkan oleh pengaruh akar menghasilkan rasio tajuk akar, rasio daun batang, rasio berat daun dan luas daun yang lebih tinggi, khususnya pada rumput. Allaby (2004) menyatakan tanaman dengan proporsi akar yang lebih banyak dapat berkompetisi lebih efektif untuk unsur hara tanah, sedangkan tanaman dengan proporsi tajuk yang lebih tinggi dapat mengumpulkan lebih banyak cahaya energi.
Berat kering tajuk B. humidicola tidak meningkat dengan aplikasi pupuk nitrogen, akan tetapi lebih tinggi pada perlakuan tanpa pemupukan nitrogen.
Variasi dalam bobot biomassa hijauan (berkisar antara 23–560 g BK/m) merupakan akibat dari variasi berat anakan (Hirata dan Pakiding 2002). Ditegaskan oleh Nakamura et al. (2002) bahwa B. humidicola lebih baik beradaptasi pada kondisi terbatas nitrogen dibandingkan dengan dua spesies
rumput lainnya, B. decumbens dan B. Brizantha. Subarao et al. (2009)
menemukan bahwa B. humidicola melepaskan Braquialactona oleh akar yang mengakibatkan menurunnya dan bahkan tertekannya nitrifikasi dalam rhizosfer tanah. Hambatan nitrifikasi merupakan strategi untuk memperpanjang waktu keberadaan N di dalam tanah dan meningkatkan efisiensi penggunaan N (Rodgers 1986).
Dalam penelitian ini, berat kering akar B. humidicola terlihat yang paling rendah dibandingkan dengan spesies lainnya. Dias-Filho (2000) dalam penelitiannya menemukan bahwa spesies B. humidocola dan B. brizantha menghasilkan biomassa bagian bawah yang rendah dengan meningkatnya naungan.
Laju fotosintesis dan ekspor gula (energi) dari daun menurun pada daun yang mulai menua. Penurunan kandungan klorofil (klorosis) merupakan gejala yang telihat pada daun yang menua (Marschner 1999). Daun tua dan mati yang terbanyak terdapat pada P.notatum, sedangkan yang terendah pada B. humidicola. Daun yang menua dan mati merupakan daun yang tertinggal dari pemotongan awal (bibit). Ditambah lagi, lebih banyak daun P. notatum yang menunjukkan warna kuning kecoklatan. Hodgson (1990) menyatakan tanda yang nampak dari proses penuaan daun adalah warna kuning yang sangat progresif dan bahkan kecoklatan dan dehidrasi. Kehilangan karena daun yang menua dan mati dan dekomposisi awalnya meningkat namun lambat, bergantung pada jumlah jaringan tua yang tertinggal setelah pemotongan dan kemudian meningkat dengan progresif dan akan sampai pada laju pertumbuhan yang sama.
Status N, P dan K tanah dan serapan N
Kandungan kalium tanah yang ditanam Cynodon nlenfluensis meningkat ketika ditambahkan serasah A.pintoi, khususnya pada pengamatan terhadap rumput yang ditanam secara monokultur. Legum ini mempunyai laju dekomposisi yang lebih
cepat dan lebih besar melepas unsur hara (Oliviera et al. 2002). Pelepasan unsur hara (N, P, K, dan Ca) dari serasah A.pintoi sangat cepat (Thomas 1994).
Konsentrasi N tajuk yang paling tinggi terlihat pada A. pintoi. Hal ini disebabkan oleh kemampuan A. pintoi untuk memfiksasi nitrogen bebas sebagaimana spesies legum lainnya. Cook (1992) melaporkan bahwa kandungan N dalam A. pintoi berkisar antara 2.5 – 3 %, dan P berkisar antara 0.18 – 0.37. A.pintoi mempunyai toleransi terhadap naungan yang tinggi (sampai dengan 50% cahaya) dan menunjukkan potensi yang tinggi sebagai cover crops. Tanaman ini menunjukkan kemampuan yang baik mengontrol gulma dan dapat memfiksasi sejumlah besar nitrogen (Rika 1995).
Manidool (1992) menemukan bahwa konsentrasi N pada A. compressus berkisar antara 1-2%. Meningkatnya konsentrasi nitrogen pada daun tanaman rumput berhubungan dengan efisiensi dari konversi energi matahari (Sophanodora 1989). Beberapa penelitian menunjukkan bahwa A. compressus dapat memfiksasi nitrogen atmosfer dan menambahkannya ke dalam tanah (Sao et al. 2007). Pekerja CSIRO telah menemukan bahwa rumput ini mempunyai sistem nitrogenase aktif yang mampu memfiksasi 13 kg N/ha/hari selama 12 minggu periode pertumbuhan selama musim panas (FAO). Secara relatif terdapat sejumlah nitrogen yang dapat difiksasi melalui asosiasi non simbiosis dalam rhizosfer A.compressus. Meskipun demikian, rendahnya level nitrogen dapat membantu bertahan pada musim pertumbuhan yang pendek dan mempertahankan rumput pada fase vegetatif. A. compressus tidak terlihat berespon dengan baik terhadap fosfor, karena dapat tumbuh dengan baik pada tanah rendah fosfor (Tropical Forages).
Serapan N tanaman diukur dengan mengalikan berat kering dengan konsentrasi hara yang ada, sehingga serapan N yang tertinggi terlihat pada A.compressus. Serapan N ini berbeda sangat nyata dibandingkan dengan spesies lainnya karena tingginya bobot kering A.compressus dibandingkan dengan spesies lainnya. Touraine et al. (1994) menemukan serapan nitrogen diatur oleh status fisiologis tanaman secara keseluruhan.
Simpulan
Dari penelitian ini dapat disimpulkan bahwa pertumbuhan tanaman hijauan menjalar berbeda disebabkan oleh spesies. Pengaruh pemupukan nitrogen berbeda tidak nyata terhadap keempat spesies tanaman menjalar yang diuji. A. compressus nyata menunjukkan pertumbuhan terbaik diantara spesies yang lain dalam hal panjang tanaman, jumlah daun, bobot tajuk dan akar, N tajuk dan serapan N aktual, kemudian diikuti oleh spesies B. humidicola, P. notatum, dan A. pintoi. Penelitian lebih lanjut diperlukan untuk mengamati kompabiliti setiap spesies dalam sistem tanaman pohon-ternak.