KOMPATIBILITAS Bacillus spp. DAN AKTINOMISET
SEBAGAI AGENS HAYATI Xanthomonas oryzae pv. oryzae DAN
PEMICU PERTUMBUHAN PADI
M. CANDRA PUTRA
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
ABSTRAK
M. CANDRA PUTRA. Kompatibilitas Bacillus spp. dan Aktinomiset sebagai
Agens Hayati Xanthomonas oryzae pv. oryzae dan Pemicu Pertumbuhan Padi. Dibimbing oleh GIYANTO.
Penyakit hawar daun bakteri adalah salah satu penyakit penting pada tanaman padi yang disebabkan oleh bakteri Xanthomonas oryzae pv. oryzae. Bakteri ini dapat terbawa benih dan bertahan dalam waktu yang lama. Beberapa mikroba diketahui berpotensi sebagai agens hayati patogen ini, seperti Bacillus spp. dan aktinomiset. Bacillus spp. dan aktinomiset telah diketahui menghasilkan antibiotik yang dapat menghambat perkembangan bakteri patogen dan menghasilkan spora yang dapat bertahan pada kondisi ekstrim. Penelitian ini bertujuan untuk mendapatkan isolat aktinomiset sebagai agens hayati patogen X. oryzae pv. oryzae dan kompatibel terhadap Bacillus spp.yang dapat diaplikasikan pada benih padi. Aktinomiset yang diisolasi tanah, diseleksi dengan metode pembiakkan ganda pada media agar dan media tumbuh berupa tanah dan pupuk kandang (1:1). Dari 16 isolat aktinomiset yang berhasil diisolasi, terdapat tiga isolat yang bersifat antagonis terhadap patogen X. oryzae pv. oryzae dan kompatibel terhadap Bacillus spp., yaitu APS7, APS9, dan APS12. Bacillus spp. dan aktinomiset diaplikasikan pada benih padi varietas Ciherang dengan metode pelapisan benih (seed coating) dan bahan pembawa yang digunakan ialah tepung dengan arang sekam sebagai komposisi utamanya. Aplikasi Bacillus spp. dan aktinomiset pada benih padi dapat menurunkan populasi patogen X. oryzae pv. oryzae pada bibit padi serta meningkatkan pertumbuhan bibit padi. Perlakuan aktinomiset dengan kode isolat APS9 dapat menekan populasi X. oryzae pv. oryzae sebesar 88,89%. Perlakuan B12+APS7 merupakan perlakuan dengan hasil persen kemunculan bibit paling tinggi di antara perlakuan lain, yaitu 83,33% pada 7 HST, diikuti dengan perlakuan APS9 dengan 79%. Perlakuan Bacillus spp. dan aktinomiset juga mampu memicu pertumbuhan tajuk tanaman sebesar 13,35-26,53% pada 7 HST.
KOMPATIBILITAS Bacillus spp. DAN AKTINOMISET
SEBAGAI AGENS HAYATI Xanthomonas oryzae pv. oryzae DAN
PEMICU PERTUMBUHAN PADI
M. CANDRA PUTRA
Skripsi
sebagai salah satu syarat untuk memperoleh
gelar Sarjana Pertanian
pada
Departemen Proteksi Tanaman
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
Judul : Kompatibilitas Bacillus spp. dan Aktinomiset sebagai Agens Hayati Xanthomonas oryzae pv. oryzae dan Pemicu Pertumbuhan Padi Nama : M. Candra Putra
NRP : A34063063
Disetujui Dosen Pembimbing
Dr. Ir. Giyanto, M.Si. NIP. 19670709 199303 1 002 Diketahui Ketua Departemen Dr. Ir. Dadang, MSc. NIP. 19640204 199002 1 002 Tanggal Lulus:
RIWAYAT HIDUP
Penulis bernama lengkap M. Candra Putra, dilahirkan di Cirebon pada tanggal 30 November 1988. Penulis merupakan anak bungsu dari empat bersaudara dari pasangan Mumud Sunaryo dan Mumun Munarsih.
Penulis menempuh pendidikan sekolah dasar di Indramayu dan sekolah menengah di Cirebon. Pendidikan sekolah dasar diselesaikan di SDN Kedokan Agung III pada tahun 2000. Pendidikan menengah penulis tempuh di SMP Negeri 1 Cirebon, lulus pada tahun 2003. Pendidikan kemudian dilanjutkan di SMA Negeri 1 Cirebon selama tiga tahun dan lulus pada tahun 2006. Penulis melanjutkan pendidikan di IPB melalui jalur SPMB (Seleksi Penerimaan Mahasiswa Baru) pada tahun 2006, dan pada tahun 2007, penulis diterima sebagai mahasiswa Departemen Proteksi Tanaman, Fakultas Pertanian.
Selama menjalani pendidikan di perguruan tinggi, penulis aktif dalam beberapa organisasi. Pada tahun 2006-2007, penulis aktif di Ikatan Kekeluargaan Cirebon-IPB (IKC-IPB) sebagai anggota divisi Hubungan Masyarakat dan Forum Silaturahmi Alumni ESQ IPB (FOSMA IPB) sebagai anggota. Pada tahun 2008, penulis bergabung dalam Himpunan Mahasiswa Proteksi Tanaman (HIMASITA) sebagai anggota Divisi Administrasi Rumah Tangga. Penulis juga aktif dalam beberapa club mitra HIMASITA, yaitu sebagai editor Majalah Metamorfosa dan sebagai anggota di Organic Farming Club serta Entomology Club. Pada Januari 2009 penulis diamanahkan untuk menjadi ketua umum HIMASITA selama satu periode kepengurusan hingga Desember 2009. Penulis juga aktif dalam mengikuti Program Kreativitas Mahasiswa (PKM) pada tahun 2008-2009. Penulis telah berhasil menghasilkan satu laporan hasil PKM-Pengabdian Masyarakat dan empat laporan PKM-Penelitian.
PRAKATA
Segala pujian dan ucapan syukur penulis panjatkan ke hadirat Allah SWT atas segala nikmat dan ridho-Nya sehingga penulis dapat menyelesaikan skripsi dengan judul “Kompatibilitas Bacillus spp. dan Aktinomiset sebagai Agens Hayati Xanthomonas oryzae pv. oryzae dan Pemicu Pertumbuhan Padi”. Penyusunan ini dilakukan berdasarkan atas penelitian yang telah penulis lakukan pada bulan Febuari hingga September 2010 di Laboratorium Bakteriologi Tumbuhan, Departemen Proteksi Tanaman IPB.
Penulis ucapkan terima kasih kepada Dr. Ir. Giyanto, M.Si atas bimbingan, nasihat, dan motivasi yang telah diberikan kepada penulis selama pelaksanaan penelitian hingga penyusunan skripsi ini, Dr. Ir. I Wayan Winasa M.Si selaku dosen penguji tamu pada sidang skripsi yang berkenan memberikan saran dan kritik yang membangun, Ir. Titiek S. Yuliani, SU. selaku dosen pembimbing akademik yang telah memberikan bimbingan dan dorongan selama penulis menjadi mahasiswa di Departemen Proteksi Tanaman (DPT).
Penulis juga ucapkan terima kasih dan apresiasi yang luar biasa kepada kedua orang tua tercinta atas segala perhatian, dukungan, motivasi, dan doa yang selalu tercurah kepada penulis. Ucapan terimakasih juga penulis ucapkan kepada kakak-kakak tercinta (Sukmawati, Suryanto, Agustina, Boy, dan Iwan) yang selalu memberikan semangat dan nasihat kepada penulis. Terima kasih kepada sahabat-sahabat mahasiswa Departemen Proteksi Tanaman IPB atas semangat dan inspirasi yang diberikan kepada penulis dalam menyelesaikan penyusunan skripsi ini, terutama untuk Redi, Faishol, Adde, Ibnu, Fitrah, Herlie, Lara, Sari, Ita, Dilah, Ana, Yeni serta rekan-rekan DPT lainnya (angkatan 42, 43, 44, dan 45) yang tidak dapat disebutkan satu persatu. Terima kasih kepada rekan-rekan di Laboratorium Bakteriologi Tumbuhan, Mbak Didi, Mbak Nilda, Pak Rustam, Pak Husda, dan Ibu Lia, atas semua bantuan dan nasihat yang diberikan selama penulis melakukan penelitian di Laboratorium Bakteriologi Tumbuhan. Tak lupa pula ucapan terima kasih penulis ucapkan kepada teman-teman kosan Fullhouse dan semua pihak yang telah membantu penulis dalam pelaksanaan penelitian hingga penyusunan skripsi ini.
Penulis menyadari bahwa skripsi ini bukan merupakan suatu karya tulis ilmiah yang sempurna. Oleh sebab itu, penulis mengharapkan masukan berupa kritik dan saran yang bersifat membangun dari pembaca. Semoga skripsi ini dapat bermanfaat baik bagi penulis maupun para pembacanya.
Bogor, Januari 2011
DAFTAR ISI
Halaman
DAFTAR TABEL ... viii
DAFTAR GAMBAR ... ix DAFTAR LAMPIRAN ... x PENDAHULUAN ... 1 Latar Belakang ... 1 Tujuan ... 4 Manfaat ... 4 TINJAUAN PUSTAKA ... 5 Pengandalian Hayati ... 5 Bacillus spp. ... 5 Aktinomiset ... 7
BAHAN DAN METODE ... 9
Waktu dan Tempat ... 9
Peremajaan Isolat Bacillus spp. ... 9
Isolasi dan Seleksi Aktinomiset sebagai Agens Hayati ... 10
Uji Kompatibilitas Bacillus spp. dan Aktinomiset ... 10
Pengujian Aplikasi Bacillus spp. dan Aktinomiset terhadap Penekanan X. oryzae pv. Oryzae dan Pertumbuhan Tanaman Padi ... 11
Penyiapan Spora Bacillus spp. dan Aktinomiset ... 11
Penyiapan Formulasi Spora Bacillus spp. dan Aktinomiset pada Bahan Pembawa Berbentuk Tepung... 12
Aplikasi Formulasi Bacillus sp. dan Aktinomiset pada Benih Padi ... 13
Pengujian Formulasi Bacillus spp. dan Aktinomiset terhadap
Penekanan Populasi X. oryzae pv. oryzae ... 14
Pengujian Formulasi Spora Bacillus spp. dan Aktinomiset terhadap Pertumbuhan Bibit Padi... 11
Analisis Statistik ... 16
HASIL DAN PEMBAHASAN ... 17
Isolat Bacillus spp. ... 17
Isolasi dan Seleksi Aktinomiset sebagai Agens Hayati ... 17
Uji Kompatibilitas Bacillus spp. dan Aktinomiset ... 20
Pengujian Aplikasi Bacillus spp. dan Aktinomiset terhadap Penekanan X. oryzae pv. Oryzae dan Pertumbuhan Tanaman Padi ... 23
Pengujian Formulasi Bacillus spp. dan Aktinomiset terhadap Penekanan Populasi X. oryzae pv. oryzae ... 23
Pengujian Formulasi Spora Bacillus spp. dan Aktinomiset terhadap Pertumbuhan Bibit Padi... 24
PENUTUP ... 28
Kesimpulan ... 28
Saran ... 28
DAFTAR PUSTAKA ... 29
DAFTAR TABEL
Nomor Halaman
1. Perlakuan terhadap benih padi pada pengujian formulasi spora
Bacillus spp. dan aktinomiset ... 15 2. Daftar isolat aktinomiset hasil isolasi berdasarkan media isolasi ... 18 3. Pembentukan zona hambatan oleh aktinomiset terhadap koloni
X. oryzae pv. oryzae pada media TSA ... 19 4. Pembentukan zona hambatan oleh aktinomiset terhadap koloni
Bacillus spp. pada media TSA ... 20 5. Pengaruh perlakuan formulasi pada benih padi terhadap populasi
X. oryzae pv. oryzae pada bibit padi berumur 7 HST ... 23 6. Pengaruh perlakuan formulasi pada benih padi terhadap persen
perkecambahan benih ... 24 7. Pengaruh perlakuan formulasi pada benih padi terhadap tinggi
DAFTAR GAMBAR
Nomor Halaman
1 Koloni Bacillus spp. pada media TSA ... 17 2 Koloni aktinomiset pada media agar padat dan penampakan
mikroskopis ... 18 3 Pengaruh uji kompatibilitas bakteri terhadap populasi
Bacillus spp. dan aktinomiset pada media tanah dan pupuk
kandang steril ... 21 4 Pengaruh bibit padi yang tidak diberi perlakuan (kiri) dan
DAFTAR LAMPIRAN
Nomor Halaman
1 Hasil analisis ragam pengujian formulasi spora Bacillus spp. dan Aktinomiset terhadap penekanan populasi X. oryzae
pv. oryzaea pada 7 HST ... 32 2 Hasil analisis ragam pengaruh perlakuan formulasi spora
Bacillus spp. dan aktinomiset pada benih padi terhadap
persen kemunculan bibit padi pada 4 HST ... 32 3 Hasil analisis ragam pengaruh perlakuan formulasi spora
Bacillus spp. dan aktinomiset pada benih padi terhadap
persen kemunculan bibit padi pada 5 HST ... 32 4 Hasil analisis ragam pengaruh perlakuan formulasi spora
Bacillus spp. dan aktinomiset pada benih padi terhadap
persen kemunculan bibit padi pada 6 HST ... 33 5 Hasil analisis ragam pengaruh perlakuan formulasi spora
Bacillus spp. dan aktinomiset pada benih padi terhadap
persen kemunculan bibit padi pada 7 HST ... 33 6 Hasil analisis ragam pengujian formulasi spora Bacillus spp.
PENDAHULUAN
Latar Belakang
Permasalahan yang cukup dominan dalam dunia pertanian adalah masalah organisme pengganggu tanaman (OPT), termasuk di Indonesia yang memiliki iklim tropis yang cocok bagi perkembangan patogen penyebab penyakit tanaman. Penyebab penyakit adalah organisme hidup patogenik maupun faktor lingkungan fisik (Agrios, 1988). Kerugian yang sering kali ditimbulkan oleh penyakit tanaman ini antara lain adalah penurunan kualitas dan kuantitas hasil produksi pertanian. Penyakit hawar daun bakteri, atau dikenal juga dengan istilah penyakit kresek, merupakan salah satu penyakit penting pada tanaman padi. Penyakit ini dapat merusak tanaman mulai dari fase bibit hingga generatif. Kerugian yang ditimbulkan bekisar antara 20-30%, bergantung pada varietas yang ditanam dan musim tanam (CAB Internasional 2005, dalam Sulistiani 2009). Penyakit ini disebabkan oleh bakteri X. oryzae pv. oryzae yang bersifat fitopatogenik. Bakteri X. oryzae pv. oryzae ini dapat terbawa benih dan bertahan dalam waktu yang cukup lama. Hal ini dikarenakan bakteri berada pada fase dorman ketika berada pada benih (Singh dan Mathur 2004).
Untuk mengamankan produksi akibat serangan OPT, petani seringkali menggunakan pestisida secara berlebihan, sehingga menimbulkan dampak negatif yang tidak diinginkan. Untuk mengurangi penggunaan pestisida, maka diperlukan alternatif pengendalian OPT yang ramah lingkungan. Saat ini, perhatian mulai beralih ke sumber daya biologi dalam meningkatkan kesehatan dan ketahanan tanaman terhadap penyakit, antara lain melalui peran mikroba tanah yang bermanfaat.
Mikroba yang bersifat menguntungkan bagi tanaman, termasuk sebagai agens penginduksi ketahanan, hidup di daerah sekitar perakaran (rhizosphere), di mana terdapat eksudat yang dikeluarkan akar sebagai nutrisi bagi mikroba itu sendiri. Mikroba yang banyak diteliti adalah kelompok rizobakteria pemacu pertumbuhan tanaman atau dikenal dengan PGPR (Plant Growth Promoting Rhizocacteria). PGPR merupakan agens pengendali hayati yang menjanjikan dapat menekan OPT di lapang (Nelson 2004).
Salah satu PGPR yang telah dikembangkan dan dimanfaatkan sebagai agens pengendali biologi adalah Bacillus spp., yang merupakan bakteri Gram positif pendegradasi amilum yang umumnya ditemukan di tanah. Bacillus spp. mampu menghasilkan beberapa jenis antibiotik dan hormon pengatur tumbuh. Di luar negeri Bacillus spp. telah dikomersilkan secara luas sebagai agens antagonis atau sebagai bakteri pemicu pertumbuhan tanaman dalam berbagai merek dagang maupun formulasi. Menurut Fravel et al. (1998) produk-produk tersebut banyak digunakan dalam pengendalian berbagai macam penyakit pada tanaman. Bakteri ini memiliki kemampuan membentuk struktur bertahan berupa endospora, yang memungkinkan organisme ini tahan terhadap kondisi lingkungan yang ekstrim, seperti kadar air yang rendah.
Selain Bacillus spp., aktinomiset juga merupakan mikroba yang berpotensi memiliki kemampuan dalam mengendalikan mikoroorganisme penyebab penyakit pada tanaman. Madigan et al (1996) menyatakan bahwa aktinomiset merupakan kelompok besar dari bakteri berbentuk filamen. Fakta mengindikasikan bahwa secara kuantitatif dan kualitatif aktinomiset memiliki peranan yang penting pada rizosfer, di mana aktinomiset berpengaruh terhadap pertumbuhan tanaman dan melindungi perakaran tanaman dari serangan fungi patogen akar. Hal ini menarik banyak ahli mikrobiologi untuk menggunakan aktinomiset sebagai agens hayati dalam mengendalikan patogen tanah. Beberapa hasil penelitian menunjukkan bahwa aktinomiset merupakan kelompok mikroba yang menjanjikan dalam pengendalian biologis (Crawford et al. 1993). Mikroba ini pun telah banyak diketahui dapat menghasilkan beragam antibiotik sebagai metabolit sekundernya (Sabaratnam dan Traquair 2001).
Aktinomiset termasuk golongan bakteri Gram positif dan menghasilkan struktur bertahan berupa spora yang dapat bertahan dalam kondisi tidak menguntungkan, seperti rendahnya kadar air dan suhu tinggi serta dapat bertahan dalam waktu yang lama. Tidak seperti bakteri penghasil spora lainnya, aktinomiset memproduksi spora sebagai cara utama bakteri ini dalam melakukan pemencaran atau dispersal (Schaad et al. 2000). Aktinomiset dikenal sebagai bakteri yang bersifat saprofit dan sangat umum dijumpai di rhizosfer hingga lapisan tanah dalam. Menurut Schaad et al. (2000), dari sekitar 400 spesies
Streptomyces, sangat sedikit yang diketahui menjadi patogen pada tanaman. Isolasi aktinomiset dari jaringan tanaman dan lahan pertanian sering kali diperoleh aktinomiset yang bersifat saprofitik.
Baik Bacillus spp. maupun aktinomiset, keduanya dapat menghasilkan spora yang dapat bertahan pada kondisi ekstrim, seperti rendahnya kadar air. Berdasarkan persamaan sifat ini, maka kedua mikroba ini berpotensi untuk dapat diaplikasikan pada bahan pembawa dalam bentuk kering berupa tepung. Hal ini dilakukan agar Bacillus spp. maupun aktinomiset dapat digunakan dalam pengendalian penyakit tanaman yang efektif dan aman bagi manusia serta lingkungan sekitarnya. Bahan pembawa dalam bentuk tepung juga dinilai lebih efisien dalam penggunaannya karena lebih mudah diaplikasikan. Selain itu, mikroba dalam bahan pembawa berbentuk tepung memiliki umur simpan yang relatif lebih panjang karena mikroba berada dalam fase dorman, yaitu dalam bentuk spora yang dapat bertahan dalam waktu yang cenderung lebih lama daripada bentuk sel vegetatifnya.
Kondisi tersebut tentu didukung dengan terjaganya kadar air dalam tepung tersebut agar tetap rendah. Penggunaan bahan pembawa yang tepat dan murah pun akan menjadikan aplikasi mikroba dalam bentuk tepung menjadi dinilai lebih efektif dan efisien bagi penggunanya. Muis (2006) melaporkan bahwa penggunaan beberapa bahan, seperti talc, tepung beras, tepung singkong, abu pegunungan, dan arang, dengan konsentrasi tertentu dapat menjadi bahan pembawa bagi mikroba yang baik pada perlakuan benih dengan metode seed coating. Hasil penelitian Putra et al. (2009) aplikasi Bacillus spp. pada benih padi dengan bahan pembawa 70% tepung singkong, 1% tepung cangkang udang, 1% ragi, 10% dedak halus, dan 18% talc mampu menghambat pertumbuhan patogen X. oryzae pv. oryzae sebesar 96,6% serta meningkatkan tinggi tajuk tanaman padi sebesar 6,83% pada usia 3 minggu setelah tanam.
Namun, untuk membuat suatu formulasi yang mengandung lebih dari satu jenis mikroba, terlebih dahulu diperlukan adanya kajian mengenai kompatibilitas mikroba tersebut. Hal ini diperlukan agar mikroba yang digunakan tidak saling meniadakan karena memiliki sifat antagonis antar keduanya. Oleh sebab itu, penelitian ini akan mengkaji kompatibilitas Bacillus spp. dan aktinomiset,
kemampuan keduanya dalam menekan patogen yang menyebabkan penyakit pada tanaman, serta peranannya dalam mendukung pertumbuhan tanaman.
Tujuan
Penelitian ini bertujuan untuk mendapatkan isolat aktinomiset sebagai agens hayati patogen X. oryzae pv. oryzae penyebab penyakit hawar daun bakteri dan mengetahui kompatibilitasnya terhadap Bacillus spp.yang dapat diaplikasikan pada benih padi.
Manfaat
Manfaat dari penelitian ini berupa informasi dan data isolat aktinomiset sebagai agens hayati patogen X. oryzae pv. oryzae penyebab penyakit hawar daun bakteri dan kompatibilitasnya terhadap Bacillus spp.yang dapat diaplikasikan pada benih padi dalam rangka penyediaan teknik pengendalian alternatif patogen X. oryzae pv. oryzae.
TINJAUAN PUSTAKA
Pengendalian Hayati
Pengendalian hayati adalah proses pengurangan kepadatan inokulum atau aktivitas patogen dalam menimbulkan penyakit yang berada dalam keadaan aktif maupun dorman oleh satu atau lebih oragisme baik secara aktif maupun manipulasi lingkungan dan inang, dengan menggunakan agens antagonis, atau dengan mengintroduksi secara massal satu atau lebih organisme antagonis (Baker dan Cook 1974).
Pada dasarnya manusia telah melakukan pengendalian hayati sejak manusia mengenal budidaya pertanian dan munculnya OPT yang merugikan. Suwahyono (2010) mengemukakan bahwa pengendalian hayati yang dilakukan saat itu hanya bersifat percobaan berdasarkan pengalaman, dengan pendekatan ilmiah yang minim.
Saat muncul Revolusi Hijau, kegiatan pengendalian hayati sempat jarang dilakukan karena penggunaan bahan kimia sintetik dinilai lebih efektif dan efisien dalam mengendalikan OPT. Namun beberapa dekade belakangan ini, seiring dengan perkembangan dunia pertanian dan merebaknya isu lingkungan, penggunaan bahan kimia sintetik mulai dikurangi secara bertahap. Pengendalian hayati pun mulai gencar dilakukan kembali karena dinilai lebih ramah lingkungan. Penggunaan organisme hidup sebagai agens antagonis dalam mengendalikan OPT mulai dilakukan dengan pendekatan ilmiah yang lebih jauh dan perkembangannya tampak semakin pesat. Agens antagonis dapat berupa mikroorganisme yang dapat mempengaruhhi kemampuan bertahan atau berpengaruh negatif terhadap aktivitas patogen dalam menimbulkan penyakit (Agrios 1997).
Bacillus spp.
Bacillus spp. ialah kelompok bakteri yang umum ditemukan di berbagai lingkungan ekologi, baik di tanah, air, maupun udara. Bakteri ini merupakan bakteri Gram positif yang dapat membentuk endospora yang berbentuk oval di bagian sentral sel. Spora berfungsi untuk bertahan hidup antara lain pada suhu dan kondisi lingkungan yang ekstrim. Sel Bacillus spp. berbentuk batang, berukuran
0,3-2,2 x 1,2-7,0 µm dan mempunyai flagel peritrikus. Bakteri ini dapat tumbuh pada suhu 45° C, pH 5-7, NaCl 7%, menghidrolisis pati, serta membentuk asam sitrat dari karbohidrat glukosa, arabinosa, manitol, dan silosa (Sonenshein et al. 2002).
Pada umumnya Bacillus spp. dapat digunakan sebagai agens biokontrol terhadap patogen tanaman walaupun diketahui terdapat strain yang dapat membusukkan biji kedelai. Biji kedelai yang diinokulasikan B. subtilis strain virulen (isolat VS) pada suhu 30-35° C dan kelembaban udara relatif 98% akan menunjukkan busuk berlendir 5 hari setelah inokulasi. (Sinclair dan Backman 1989, dalam Desmawati 2006).
Bakteri Bacillus spp. yang bersifat antagonis mampu menekan pertumbuhan mikroorganisme lain karena memproduksi antibiotik berupa lipopeptida yang disebut basitrasin dengan mekanisme merusak membran sel bakteri (Leary dan Chan 1988, dalam Desmawati 2006). Jenis metabolit sekunder lain yang diproduksi Bacillus spp. adalah bio-surfaktan yang disebut surfaktin atau subtilisin. Surfaktin merupakan lipopeptida siklik yang berfungsi menurunkan tegangan permukaan air dan juga bersifat antibiotik (Hommel dan Ratledge 1933, Desai dan Desai 1933, dalam Dirmawati 2004).
Sebagian besar anggota Bacillus spp. tidak dianggap sebagai bakteri patogen terhadap manusia, walaupun dapat mengkontaminasi makanan, namun jarang menimbulkan keracunan (Sonenshein,et al. 2002). Schaad et al. (2000) menyatakan bahwa hanya terdapat tiga kelompok Bacillus yang diketahui sebagai patogen tanaman, yaitu B. circulans, B. megaterium pv. cerealis, dan B. polymyxa. Bacillus spp. memiliki aktivitas antifungal yang tinggi (Jing dan Qian 2007) dan berperan dalam menekan beberapa fungi yang bersifat patogen, seperti Rhizoctonia, Fusarium (Zhang et al. 2009) dan Aspergilus (Muis 2006). Selain memiliki kemampuan dalam menekan perkembangan fitopatogen, Bacillus spp. pun diketahui dalam mendukung pertumbuhan tanaman. McQuilken et al. (1998) mengemukakan bahwa aplikasi Bacillus spp. pada benih kedelai mampu mengurangi kerusakan bibit karena kerusakan saat imbibisi. Selain itu, perlakuan benih dengan Bacillus spp. untuk merangsang pertumbuhan tanaman dan
membantu mengurangi patogen terbawa benih telah menjadi bahan penelitian yang menarik selama lebih dari 20 tahun terakhir.
Aktinomiset
Aktinomiset adalah kelompok besar dari bakteri berfilamen, umumnya bersifat Gram positif, dan membentuk filament yang bercabang. Pertumbuhan aktinomiset yang sukses mampu mengahasilkan jaringan berfilamen yang memiliki cabang-cabang yang rumit, disebut juga dengan miselium. Ada juga yang menyebutnya miselia aerial karena miselia dapat tumbuh pada lapisan udara. Ukuran miselium umumnya memiliki diameter 0,5-1,0 µm, dengan panjang yang tidak tentu, dan tidak memiliki sekat pada fase vegetatif (Madigan et al. 1993). Sebagian besar aktinomiset mampu menghasilkan spora dari ujung-ujung miselium yang terbentuk. Spora aktinomiset dikenal dengan eksospora, karena terbentuk tidak dari dalam sel serta memiliki dinding yang tidak terlalu tebal (Janse 2005).
Aktinomiset dapat ditemukan di berbagai lingkungan ekologi, seperti air laut (Patil et al. 2001, air sungai (Rifaat 2003), dan tanah gua (Nakaew et al. 2009). Bahkan, Khan et al. (2008) berhasil mengisolasi aktinomiset dari floppy dan compact disc. Beberapa aktinomiset juga diketahui merupakan organisme endofit dan dapat diisolasi dari tanaman (Kunoh 2002).
Salah satu anggota Aktinomiset yang memiliki kemampuan sebagai agens hayati adalah Streptomyces sp. Bakteri ini merupakan mikroorganisme yang banyak menghasilkan substansi antibiotik, salah satunya aktif menghambat pertumbuhan cendawan patogen pada tumbuhan. Hasil penelitian menunjukkan bahwa Streptomyces sp. berpengaruh terhadap penghambatan pertumbuhan cendawan, salah satunya adalah Fusarium sp. (Murdiyah 2008).
Pada tahun 1944 Selman Waksman menemukan streptomisin yang merupakan salah satu antibiotik yang dihasilkan oleh Streptomyces sp. Perkembangan ini merangsang penelitian lebih lanjut terhadap genus Streptomyces dalam usaha mencari mikroorganisme penghasil antibiotik. Sejak saat itu penelitian aktinomiset, terutama Streptomyces sp., menjadi gudang utama untuk memperoleh antibiotik baru. Di berbagai lembaga penelitian dilakukan
pencarian antibiotik dari berbagai tipe mikroorganisme terutama dari kelas aktinomiset dan telah berhasil menemukan antibiotik baru seperti khlortetrasiklin, neomisin, oksitetrasiklin, dan eritromisin (Suwandi 1993).
Dalam bidang fitopatologi, khususnya dalam hal pengendalian patogen pada tanaman, aktinomiset diketahui dapat mengendalikan beberapa patogen penting, seperti Rhizoctonia solani (Sabaratnam dan Traquair, 2001) dan Phytium ultimum (Crawford et al., 1993). Bahkan dari hasil penelitian Nakaew et al. (2009) melaporkan bahwa salah satu aktinomiset yang diisolasi dari gua di Thailand, Spirillospora albida, diketahui dapat menujukkan aktivitas antimoikrobial terhadap Bacillus cereus, Staphylococcus aureus, dan Paenibacillus larvae. Hasil penelitian Charoensopharat et al. (2007) menyatakan bahwa Streptomyces sp. menunjukkan aktivitas antibakteri yang signifikan terhadap perkembangan bakteri Gram negatif, seperti Xanthomonas sp..
Aktinomiset dapat menjadi objek penelitian yang sangat potensial untuk masa yang akan datang (Sabaratnam dan Traquair, 2001). Streptomyces sp. telah menunjukkan adanya produksi senyawa toksik antifungal, rhizostreptin, dalam kapasitas yang tinggi, baik pada media biakan maupun pada perakaran tomat. Bakteri ini pula menunjukkan produksi eksoenzim pendegradasi dinding sel cendawan seperti kitinase. Kajian mengenai kemampuan produksi aktinomiset akan senyawa metabolit lainnya, kemampuan metabolit tersebut menekan pertumbuhan patogen, hingga efikasi dan persistensinya di perakaran akan sangat penting untuk menghasilkan senyawa yang dapat diaplikasikan sebagai antimikroba untuk berbagai agroekosistem.
BAHAN DAN METODE
Waktu dan Tempat
Penelitian dilakukan pada bulan Maret hingga September 2010 di Laboratorium Bakteriologi Tumbuhan Departemen Proteksi Tanaman IPB, Fakultas Pertanian, Institut Pertanian Bogor.
Peremajaan Isolat Bacillus spp.
Isolat Bacillus spp. yang digunakan dalam penelitian ini diperoleh dari kultur stok koleksi Laboratorium Bakteriologi Tumbuhan Departemen Proteksi Tanaman IPB. Isolat tersebut berupa suspensi bakteri pada larutan Gliserol 20% dan disimpan dalam keadaan beku pada suhu -20°C. Sebanyak satu lup suspensi beku tersebut diambil dan digores pada media Tryptone Soy Agar (TSA) secara aseptik dengan menggunakan jarum oose. Selanjutnya, cawan diinkubasi pada suhu ruang selama dua hari. Koloni bakteri akan tumbuh pada media setelah masa inkubasi. Peremajaan isolat Bacillus spp. ini dilakukan secara aseptik di laminar air flow cabinet.
Isolasi dan Seleksi Aktinomiset sebagai Agens Hayati
Aktinomiset merupakan salah satu mikroba yang umum ditemukan di tanah. Penelitian ini menggunakan isolat aktinomiset asal tanah. Tanah sampel, yang diambil dari daerah Bogor, berasal dari tanah bagian top soil. Kemudian tanah tersebut dikeringanginkan pada suhu ruang selama 10 hari. Sebanyak 10 gr tanah sampel yang sudah kering disuspensikan pada 100 ml air steril kemudian diinkubasi pada inkubator bergoyang selama 15 menit. Pencawanan dilakukan terhadap suspensi ini dengan didahului teknik pengenceran berseri dengan konsentrasi 10-1 hingga 10-8. Pencawanan dilakukan pada media semi selektif Water-yeast extract Agar (WYE) (0,25 g yeast extract, 0,5 g K2HPO4, 18 g agar,
dan 1 L aquades) dan Casamino Acids-yeast extract-glucose-agar (YCED) (0,3 g Casamino Acids, 0,3 g Yeast extract, 0,3 g D-Glucose, 2 g K2HPO4, 18 g agar,
dan 1 L aquades). Koloni Aktinomiset akan tumbuh dalam waktu 7-10 hari, kemudian dilakukan pemurnian terhadap koloni tunggal yang terbentuk dan ini
dijadikan sebagai isolat murni. Setelah diberi label kode, isolat murni ini yang akan dijadikan sebagai bahan pada pengujian selanjutnya.
Seleksi Aktinomiset dilakukan dengan menggunakan metode biakkan ganda (dual culture). Sebanyak satu lup biakan X. oryzae pv. oryzae berusia 24 jam diambil dari media Yeast Dextrose Calcium carbonate Agar (YDCA) (15 gr Yeast Extract, 20 gr Dextrose, 20 gr CaCO3, 15 gr Agar, dan 1 L aquades) dan
diinokulasikan ke erlenmeyer berisi media 10 ml media LB serta diinkubasi pada inkubator bergoyang dengan kecepatan 100 rpm pada suhu ruang selama 18 jam. Pada waktu yang sama, sebanyak satu lup spora aktinomiset, yang diperoleh dari biakan berusia tujuh hari, diinokulasikan ke erlenmeyer berisi 10 ml media cair Luria Broth (LB) (1 gram Triptone, 0,5 gram NaCl, 0,5 gram Yeast Extract), dan diinkubasi pada inkubator bergoyang dengan kecepatan 100 rpm pada suhu ruang selama 18 jam. Setelah masa inkubasi, sebanyak 100 µl suspensi bakteri X. oryzae pv. oryzae disebar pada media TSA, kemudian dikeringanginkan selama 15 menit pada laminar air flow cabinet. Sebanyak empat buah kertas saring berdiameter 0,5 cm diletakkan pada media TSA yang telah disebar suspensi bakteri X. oryzae pv. oryzae. Pada masing-masing kertas saring diteteskan suspensi aktinomiset sebanyak 10 µl. Setelah itu, media diinkubasi pada suhu ruang selama beberapa hari. Pengamatan terhadap aktivitas antagonisme dilakukan setiap hari. Aktivitas antagonisme ditunjukkan dengan pembentukkan zona bening di sekitar kertas saring. Isolat aktinomiset yang menunjukkan sifat antagonis terhadap bakteri patogen X. oryzae pv. oryzae kemudian dijadikan isolat stok yang akan digunakan pada pengujian selanjutnya.
Uji Kompatibilitas Bacillus spp. dan Aktinomiset
Uji kompatibilitas pada media padat dilakukan dengan membiakkan Bacillus spp. dan aktinomiset secara dual culture. Sebanyak 10 μl suspensi biakan aktinomiset diteteskan pada kertas saring yang terdapat pada media agar yang telah disebar dengan 100 μl susupensi Bacillus spp. Media agar yang digunakan ialah media umum untuk membiakkan bakteri, yaitu TSA. Aktinomiset yang bersifat antagonis terhadap Bacillus spp. akan membentuk zona bening dan tidak akan digunakan sebagai bahan uji.
Isolat aktinomiset yang menunjukkan hasil positif terhadap uji kompatibilitas melalui dual culture kemudian dilakukan pengujian lebih lanjut. Bacillus spp. dan aktinomiset dibiakkan pada media tumbuh berupa tanah dan pupuk kandang yang sudah disterilisasi. Perbandingan antara tanah dan pupuk kandang ialah 1:1. Bacillus spp. dan aktinomiset yang diinokulasikan pada media tumbuh masing-masing sebanyak 105 cfu/gr media tumbuh. Setelah diinkubasi selama dua minggu pada suhu ruang, kemudian dilakukan penghintungan kepadatan populasi Bacillus spp. dan aktinomiset pada masing-masing media tumbuh dengan metode pengenceran berseri. Isolat aktinomiset yang diketahui tidak memiliki interaksi yang negatif terhadap Bacillus spp. akan dijadikan sebagai bahan pengujian selanjutnya.
Pengujian Aplikasi Bacillus spp. dan Aktinomiset terhadap Penekanan X.
oryzae pv. oryzae dan Pertumbuhan Tanaman Padi
Penyiapan Spora Bacillus spp. dan Aktinomiset
Bakteri Bacillus sp. adalah bakteri Gram positif yang dapat membentuk spora. Pembentukan spora oleh Bacillus sp. merupakan salah satu respon bakteri ini dalam bertahan pada kondisi tertentu (Muis 2006). Pada penelitian ini, produksi spora diawali dengan peremajaan biakan Bacillus spp. secara aseptik. Biakan Bacillus spp. hasil peremajaana berumur satu hari tersebut diinokulasi sebanyak satu lup ke dalam tabung reaksi berisi 5 ml media cair LB dan diinkubasi pada inkubator bergoyang dengan kecepatan 100 rpm selama 13 jam. Setelah itu sebanyak 5 ml suspensi Bacillus spp. diinokulasi ke dalam erlenmeyer 1000 ml berisi 300 ml media cair Dickinson (6 g Pancreatic Digest of Gelatin, 4 g Pancreatic Digest of Casein, 3 g Yeast Extract, 1 g Beef Extract, 0,3 g Manganolis Sulfat) dan diinkubasi pada inkubator bergoyang dengan kecepatan 100 rpm selama enam hari. Penghitungan spora Bacillus spp. dilakukan dengan memanaskan suspensi pada suhu 80 °C selama 15 menit dengan tujuan untuk mematikan sel vegetatif Bacillus spp., tetapi tidak mematikan sporanya. Setelah itu, dilakukan pengenceran berseri dan dilanjutkan dengan pencawanan pada media TSA. Pengamatan dilakukan dengan menghitung jumlah koloni Bacillus
spp. yang tumbuh setelah diinkubasikan selama 48 jam. Jumlah koloni yang tumbuh selanjutnya dikonversikan ke dalam satuan cfu/ml dengan rumus:
Populasi bakteri = Keterangan:
x = jumlah koloni yang tumbuh pada cawandengan faktor pengenceran ke- (cfu) p = faktor pengenceran ke-
v = volume suspensi yang disebar pada cawan (ml)
Isolat aktinomiset yang telah terseleksi kemudian dilakukan perbanyakan untuk keperluan pengujian selanjutnya. Perbanyakan aktinomiset dilakukan dengan menggoreskan kultur murni ke media WYE atau YCED, tergantung pada tiap isolat. Untuk persediaan, setiap isolat dengan sama yang berbeda diperbanyak hingga 10 cawan Kemudian cawan diinkubasi hingga 8-10 hari. Spora akan tumbuh pada permukaan media yang ditumbuhi koloni aktinomiset dengan penampakan kasar seperti beludru dan mengandung pigmen tertentu sehingga akan tampak memiliki beragam (Madigan et al. 1996). Selain itu, dilakukan juga perbanyakan isolat aktinomiset dengan penggunaan agar miring. Hal ini dilakukan untuk menyediakan kultur stok yang murni dan dapat diperbanyak sewaktu-waktu.
Penyiapan Formulasi Spora Bacillus spp. dan Aktinomiset pada Bahan Pembawa Berbentuk Tepung
Spora bakteri dipanen dengan sentrifugasi eppendorf pada 7.500 rpm pada suhu ruang selama 6 menit dari media kultur yang telah berumur 7 hari. Pelet dari spora bakteri kemudian dicuci 2 kali dengan larutan 0,05M Phosphate Buffer Saline (PBS) (8 gr NaCl, 0,2 gr KH2PO4, 1,15 gr Na2HPO4, 0,2 gr KCl, 1 L
aquades) pH 7,0. Spora bakteri yang telah dicuci dengan PBS selanjutnya diresuspensikan kembali pada larutan PBS dan kemudian kepadatan spora bakteri pada suspensi tersebut dihitung dengan teknik pengenceran berseri.
Spora aktinomiset dipanen dari kultur aktinomiset yang telah berumur 10 hari pada cawan petri berisi media padat. Sebanyak 10 ml aquades steril dituang pada cawan tersebut. Kemudian dengan menggunakan spatula steril, koloni
aktinomiset digores dengan lembut hingga sporanya lepas dari permukaan agar dan tersuspensi pada aquades steril. Setelah itu, suspensi spora tersebut diteteskan pada cawan kultur lain dengan menggunakan pipet. Selanjutnya dilakukan metode yang sama hingga mencapai 5 cawan. Penghitungan terhadap kepadatan spora dilakukan dengan alat Haemacytometer.
Setelah dipanen, kemudian spora diinokulasikan pada bahan pembawa yang akan digunakan. Setiap bahan pembawa yang digunakan, sebelumnya telah disaring dengan saringan 50 mess. Bahan pembawa yang akan diuji dalam penelitian ini adalah campuran tepung arang sekam, dedak halus, tepung jagung, dan tepung cangkang udang dengan perbandingan 86,5 : 3 : 10 : 0,5. Bahan pembawa yang sudah dicanpur kemudian dimasukkan ke dalam plastik tahan panas untuk dilakukan sterilisasi secara panas uap dengan menggunakan autoklaf selama 15 menit.
Formulasi dibuat dengan cara menyebarkan susupensi spora secara merata pada bahan pembawa dengan perbandingan 5 ml untuk setiap 10 g bahan pembawa. Suspensi tersebut diteteskan ke dalam plastik berisi bahan pembawa dan diaduk secara merata. Adapun kepadatan spora Bacillus spp. dan aktinomiset yang diinokulasi ialah 108 cfu/gr bahan pembawa. Formulasi bahan pembawa spora Bacillus spp. dan aktinomiset selanjutnya diletakan pada aluminium foil steril dan dilakukan pengeringan dengan oven pada suhu 60°C selama 2 jam. Selanjutnya formulasi ini digunakan untuk pengujian pada bahan tanaman.
Aplikasi Formulasi Bacillus sp. dan Aktinomiset pada Benih Padi
Benih padi yang digunakan dalam pengujian ini ialah benih padi varietas Ciherang. Benih yang akan digunakan dalam pengujian ini, sebelum diberi perlakuan dan ditanam, dilakukan analisis terhadap populasi X. oryzae pv. oryzae dengan teknik pencawanan dengan pengenceran berseri. Sebanyak 20 benih dipilih dan disterilisasi permukaan lalu digerus serta disuspensikan pada 10 ml larutan PBS. Suspensi tersebut kemudian dilakukan pengenceran berseri hingga 10-8 kali dan disebar pada cawan berisi media YDCA. Selanjutnya media diinkubasi selama 2 hari dan dilakukan peghitungan koloni X. oryzae pv. oryzae yang tumbuh. Schaad et al. (2000) menyatakan bahwa pada media YDCA koloni
X. oryzae pv. oryzae akan tampak mukoid dan berwarna kuning dengan permukaan licin.
Penanaman benih padi dilakukan pada nampan plastik yang berisi cocopeat steril sebagai media semainya. Sebelum diberi perlakuan dan ditumbuhkan pada media semai, benih padi direndam dalam tabung erlenmeyer steril berisi air steril selama semalam. Sebanyak 100 benih padi disebar pada formulasi spora bakteri yang diletakan di atas aluminium foil steril dan dicampurkan secara merata hingga seluruh benih tertutupi formulasi bakteri. Tahap ini dinamakan seed dressing atau seed coating. Kemudian benih yang sudah diberi perlakuan ditanam pada media yang telah disiapkan.
Pengujian Formulasi Bacillus spp. dan Aktinomiset terhadap Penekanan Populasi X. oryzae pv. oryzae
Analisis populasi X. oryzae pv. oryzae dilakukan pada bibit tanaman padi yang berumur 7 HST. Sebanyak 20 dari 100 sampel bibit pada setiap ulangan dari setiap perlakuan digerus pada plastik steril kemudian disuspensikan pada 50 ml air steril. Kemudian X. oryzae pv. oryzae diisolasi dari suspensi tersebut dengan metode pengenceran berseri dan pencawanan pada media YDCA. Pengamatan dilakukan dengan menghitung jumlah koloni X. oryzae pv. oryzae yang tumbuh setelah diinkubasikan selama 48 jam. Jumlah koloni yang tumbuh selanjutnya dikonversikan ke dalam satuan cfu/ml dengan rumus:
Populasi bakteri = Keterangan:
x = jumlah koloni yang tumbuh pada cawandengan faktor pengenceran ke- (cfu) p = faktor pengenceran ke-
v = volume suspensi yang disebar pada cawan (ml)
Pengujian formulasi spora Bacillus spp. dan aktinomiset terhadap penekanan populasi X. oryzae pv. oryzae dilakukan dengan menggunakan rancangan acak lengkap. Perlakuan yang diuji dapat dilihat pada Tabel 1.
Tabel 1 Perlakuan terhadap benih padi pada pengujian formulasi spora Bacillus spp. dan aktinomiset
Perlakuan Keterangan
KONTROL Benih padi tanpa aplikasi Bacillus spp. dan aktinomiset
B12 Aplikasi benih padi dengan formulasi Bacillus spp. APS7 Aplikasi benih padi dengan formulasi aktinomiset
isolat APS7
APS9 Aplikasi benih padi dengan formulasi aktinomiset isolat APS9
APS12 Aplikasi benih padi dengan formulasi aktinomiset isolat APS12
B12+APS7 Aplikasi benih padi dengan formulasi Bacillus spp. dan aktinomiset isolat APS7
B12+APS9 Aplikasi benih padi dengan formulasi Bacillus spp. dan aktinomiset isolat APS9
B12+APS12 Aplikasi benih padi dengan formulasi Bacillus spp. dan aktinomiset isolat APS12
Parameter pengamatan yang diambil ialah jumlah koloni X. oryzae pv. oryzae yang tumbuh berdasarkan hasil pengenceran berseri dan pencawanan pada media YDCA. Pengujian dilakukan dengan tiga ulangan. Pada setiap ulangan diambil 20 unit sampel yang dipilih secara acak yang akan dijadikan sebagai sumber parameter pengamatan.
Pengujian Formulasi Spora Bacillus spp. dan Aktinomiset terhadap Pertumbuhan Bibit Padi
Pengujian formulasi spora Bacillus spp. dan aktinomiset terhadap pertumbuhan tanaman dilakukan dengan menggunakan rancangan acak lengkap. Perlakuan yang diuji dapat dilihat pada Tabel 1.
Parameter pengamatan yang diambil ialah persen kemunculan bibit padi dan tinggi tajuk dari bibit yang tumbuh. Pengujian dilakukan dengan tiga ulangan. Pada setiap ulangan diambil 20 unit sampel yang dipilih secara acak yang akan dijadikan sebagai sumber parameter pengamatan.
Analisis Statistik
Data pengujian formulasi spora Bacillus spp. dan aktinomiset terhadap populasi X. oryzae pv. oryzae dan pertumbuhan tajuk pada bibit padi diolah menggunakan Analisis Sidik Ragam (ANOVA) dengan program The Statistical Analysis System (SAS) for Windows 9.0. Pengaruh yang berbeda nyata akan dilakukan uji lanjut dengan uji selang berganda Duncan dengan taraf nyata (α) = 5%.
HASIL DAN PEMBAHASAN
Isolat Bacillus spp.
Pada media agar padat, Bacillus spp. berbentuk tidak beraturan dengan pinggiran bergerigi dan berwarna putih pucat kecoklatan (Gambar 1). Muis (2006) mengungkapkan bahwa Bacillus spp. berpotensi dalam mengendalikan penyakit yang disebabkan oleh X. oryzae.Waites et al. (1970) mengemukakan bahwa Bacillus spp. tergolong ke dalam bakteri Gram positif yang dapat membentuk struktur berupa endospora yang dapat bertahan pada kondisi ekstrim dan dalam waktu yang lama hingga hitungan tahun.
Gambar 1 Koloni Bacillus spp. pada media TSA
Isolasi dan Seleksi Aktinomiset sebagai Agens Hayati
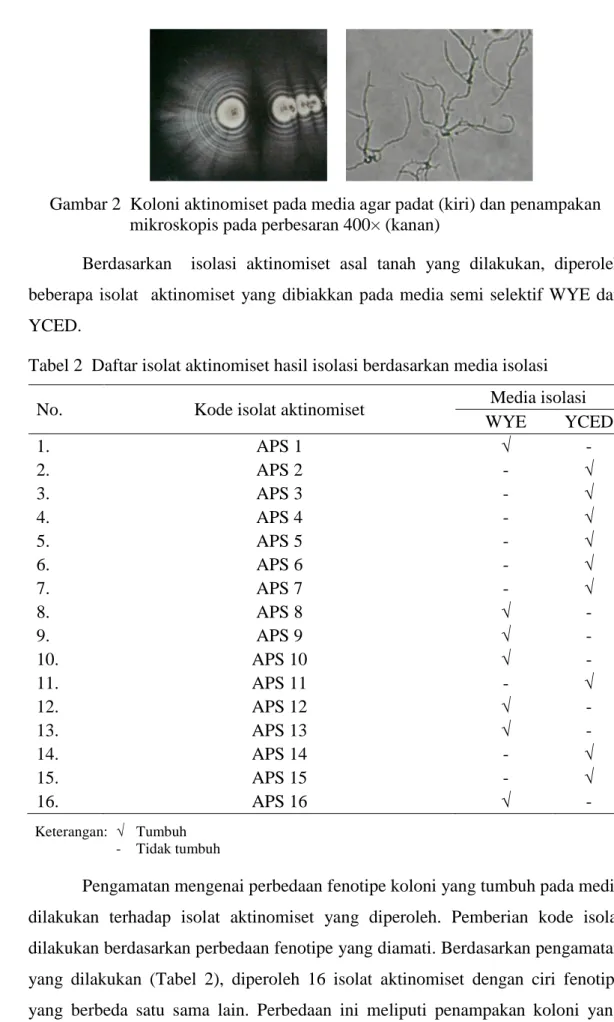
Menurut Madigan et al. (1996), koloni aktinomset pada cawan agar mudah dibedakan dengan koloni bakteri lain, bahkan cendawan sekalipun. Koloni aktinomiset memiliki ciri khas berupa penampakan yang terlihat berdebu atau bertekstur seperti beludru (Gambar 2). Penampakan tersebut merupakan spora yang dihasilakan oleh hifa aerial yang hanya dimiliki oleh aktinomiset. Hal ini berbeda dengan bakteri yang memiliki koloni yang mukoid ataupun permukaan licin yang khas. Bakteri ini sudah dikenal selama bertahun-tahun sebagai bakteri yang dapat memproduksi antibiotik sebagai metabolit sekundernya (Stevenson 1956). Aktinomiset merupakan suatu kelompok bakteri berfilamen yang umum ditemukan di tanah. Aktinomiset termasuk ke dalam golongan bakteri Gram positif dan dapat menghasilkan struktur bertahan berupa spora.
Gambar 2 Koloni aktinomiset pada media agar padat (kiri) dan penampakan mikroskopis pada perbesaran 400× (kanan)
Berdasarkan isolasi aktinomiset asal tanah yang dilakukan, diperoleh beberapa isolat aktinomiset yang dibiakkan pada media semi selektif WYE dan YCED.
Tabel 2 Daftar isolat aktinomiset hasil isolasi berdasarkan media isolasi
No. Kode isolat aktinomiset Media isolasi
WYE YCED 1. APS 1 √ - 2. APS 2 - √ 3. APS 3 - √ 4. APS 4 - √ 5. APS 5 - √ 6. APS 6 - √ 7. APS 7 - √ 8. APS 8 √ - 9. APS 9 √ - 10. APS 10 √ - 11. APS 11 - √ 12. APS 12 √ - 13. APS 13 √ - 14. APS 14 - √ 15. APS 15 - √ 16. APS 16 √ - Keterangan: √ Tumbuh - Tidak tumbuh
Pengamatan mengenai perbedaan fenotipe koloni yang tumbuh pada media dilakukan terhadap isolat aktinomiset yang diperoleh. Pemberian kode isolat dilakukan berdasarkan perbedaan fenotipe yang diamati. Berdasarkan pengamatan yang dilakukan (Tabel 2), diperoleh 16 isolat aktinomiset dengan ciri fenotipe yang berbeda satu sama lain. Perbedaan ini meliputi penampakan koloni yang
terbentuk pada media agar, warna koloni, serta pertumbuhan isolat tersebut dalam menghasilkan spora. Keenambelas isolat ini tumbuh pada dua media isolasi yang berbeda, yaitu WYE dan YCED. Penggunaan media WYE dan YCED pernah dilakukan Crawford et al. (1993) dalam mengisolasi aktinomiset. Media dengan kandungan nutrisi yang rendah baik digunakan dalam melakukan isolasi aktinomiset karena media jenis ini dapat mengurangi kontaminasi yang dapat diakibatkan oleh fungi dan bakteri. Adapun isolat yang tumbuh pada media WYE adalah APS 1, APS 8, APS 9, APS 10, APS12, APS 13, dan APS 16. Sedangkan isolat yang tumbuh pada media YCED adalah APS 2, APS 3, APS4, APS 5, APS 6, APS 7, APS 11, APS 14, dan APS 16.
Tabel 3 Pembentukan zona hambatan oleh aktinomiset terhadap koloni X. oryzae pv. oryzae pada media TSA
No. Kode isolat aktinomiset Pembentukan zona bening
1. APS 1 - 2. APS 2 - 3. APS 3 - 4. APS 4 + 5. APS 5 - 6. APS 6 - 7. APS 7 + 8. APS 8 - 9. APS 9 + 10. APS 10 - 11. APS 11 - 12. APS 12 + 13. APS 13 - 14. APS 14 - 15. APS 15 - 16. APS 16 -
Keterangan: + Terbentuk zona bening - Tidak terbentuk zona bening
Isolat aktinomiset yang diperoleh kemudian dilakukan pengujian terhadap daya antagonisme yang muncul terhadap bakteri X. oryzae pv. oryzae. Hasil pengujian menunjukkan bahwa tidak semua aktinomiset memiliki kemampuan dalam menghambat pertumbuhan X. oryzae pv. oryzae. Dari 16 isolat aktinomiset yang berhasil diisolasi, hanya empat isolat yang memiliki daya hambat terhadap
X. oryzae pv. oryzae, yaitu aktinomiset dengan kode isolat APS 4, APS 7, APS 9, dan APS 12 (Tabel 3). Ada tidaknya kemampuan daya hambat yang dimiliki aktinomiset dapat diketahui dengan terbentuknya zona hambatan pada media biakan cawan. Zona bening yang terbentuk ini merupakan suatu akibat dari adanya aktivitas antimikroba yang dimiliki oleh aktinomiset. Aktivitas antimikroba ini dapat dikarenakan oleh suatu senyawa antibiotik yang dihasilkan aktinomiset, seperti Amphotericin, Cyclohexamide, Nystatin, dan Streptomycin yang dikenal memiliki sifat antimikroba (Ainsworth 1971).
Uji Kompatibilitas Bacillus spp. dan Aktinomiset
Uji kompatibilitas dilakukan dalam dua tahap, yaitu dengan menggunakan metode dual culture pada media agar dan dengan menumbuhkan kedua bakteri ini di media tanah dan pupuk kandang steril. Pengujian dengan metode dual culture, yang dilakukan pada media TSA, menunjukkan hasil bahwa hampir seluruh isolat aktinomiset tidak menunjukkan aktivitas antagonisme terhadap Bacillus spp. Satu-satunya isolat aktinomiset yang menunjukkan aktivitas antagonisme terhaap Bacillus spp. ialah aktinomiset dengan kode isolat APS 4. Hal ini diperoleh dari pembentukan zona bening pada media TSA yang dilakukan pengujian.
Tabel 4 Pembentukan zona hambatan oleh aktinomiset terhadap koloni Bacillus spp. pada media TSA
No. Kode Isolat Pembentukan zona
bening
Kompatibilitasnya terhadap Bacillus spp.
1. APS 4 + Tidak kompatibel
2. APS 7 - Kompatibel
3. APS 9 - Kompatibel
4. APS 12 - Kompatibel
Keterangan: + Terbentuk zona bening - Tidak terbentuk zona bening
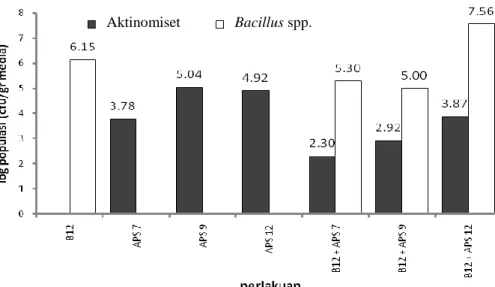
Gambar 3 Pengaruh uji kompatibilitas bakteri terhadap populasi Bacillus spp. dan aktinomiset pada media tanah dan pupuk kandang steril
Berdasarkan hasil pengujian (Tabel 4), sebanyak 3 isolat aktinomiset yang kompatibel terhadap Bacillus spp., yaitu isolat APS 7, APS 9, dan APS 12. Ketiga isolat ini kemudian dilakukan pengujian lebih lanjut dengan menumbuhkan Bacillus spp. dan ketiga isolat aktinomiset pada suatu media tumbuh, baik secara terpisah maupun dikombinasikan antara Bacillus spp. dan aktinomiset media tumbuh tersebut berupa tanah dan pupuk kandang (1:1) yang telah disterilisasi.
Inkubasi dilakukan terhadap media yang telah diinokulasi bakteri selama tiga minggu. Setelah masa inkubasi selama tiga minggu, tanah yang telah diinokulasi bakteri dengan berbagai perlakuan kemudian dilakukan analisis penghitungan bakteri yang tumbuh dengan metode pengenceran berseri yang disertai dengan teknik pencawanan. Bedasarkan penghitungan yang telah dilakukan (Gambar 3), dapat diketahui bahwa terdapat perbedaan jumlah populasi antara bakteri yang ditumbuhkan secara tunggal dan ganda. Secara umum, baik Bacillus spp. maupun aktinomiset, keduanya memiliki nilai populasi yang lebih kecil saat keduanya ditumbuhkan secara bersamaan. Bacillus spp. yang ditumbuhkan secara tunggal memiliki nilai log populasi yang lebih tinggi, yaitu sebesar 6,15 (1,4×106 cfu/gr media). Namun ketika ditumbuhkan secara bersamaan dengan APS 7 dan APS 9 hanya memiliki nilai log populasi sebesar 5,30 (2×105 cfu/gr media) dan 5,00 (1×105 cfu/gr media).
Hal yang sama terjadi pada aktinomiset, ketika ditumbuhkan secara tunggal, nilai log populasinya mencapai 3,78 (6×103 cfu/gr media) untuk APS 7,
Bacillus spp. Aktinomiset
5,04 (1,1×105 cfu/gr media) untuk APS 9 dan 4,92 (8,3×104 cfu/gr media) APS 12. Namun pada kondisi dimana keduanya ditumbuhkan bersama dengan Bacillus spp., nilai populasinya menurun hingga 2,30 cfu/gr media untuk APS 7, 2,92 cfu/gr media untuk APS 9, dan 3,87 cfu/gr media untuk APS 12. Penyebab dari penurunan populasi yang terjadi, baik pada Bacillus spp. mapun aktinomiset, terjadi karena beberapa hal. Pada saat bakteri ini ditumbuhkan secara tunggal, tidak terjadi kompetisi dalam hal nutrisi maupun ruang tumbuh, sehingga bakteri dapat tumbuh dengan maksimal. Namun ketika ditumbuhkan secara bersamaan, baik Bacillus spp. bersama dengan APS 7 maupun APS 9, terjadi kompetisi dalam memperoleh nutrisi serta ruang hidup, sehingga pertumbuhan dan perkembangan bakteri ini lebih terbatas. Tetapi kompetisi ini tidak bersifat antagonis karena bakteri masih dapat tumbuh walaupun dalam kondisi yang terbatas sehingga hubungan antara Bacillus spp. dan aktinomiset ini dapat dikatakan kompatibel.
Populasi Bacillus spp. ketika ditumbuhkan dengan APS 12 mengalami kenaikan yang sigifikan, yaitu mencapai 7,56 (3,67×107 cfu/gr media). Hal ini menunjukkan bahwa perbedaan isolat aktinomiset dapat mempengaruhi pertumbuhan Bacillus spp. Perbedaan kode isolat mengindikasikan perbedaan spesies aktinomiset. Kemungkinan terdapat suatu interaksi tertentu yang bersifat mendukung yang terjadi antara APS 12 dengan Bacillus spp. sehingga populasi Bacillus spp. yang ditumbuhkan bersama dengan APS 12 lebih tinggi dibandingkan Bacillus spp. yang ditumbuhkan secara tunggal.
Crawford et al. (1993) menyatakan bahwa aktinomiset merupakan bakteri yang aktif dalam mendegradasi bahan-bahan organik, seperti lignoselulosa, kitin, dan pati dalam tanah. Schaad et al. (2000) pernah melaporkan bahwa Bacillus spp. juga memiliki hasil yang positif terhadap uji hidrolisis pati dan menghasilkan enzim amylase. Kemampuan keduanya dalam mendegradasi senyawa kompleks menjadi senyawa sederhana ini diduga mempengaruhi kompatibilitas keduanya dalam pengujian pada media tanah dan pupuk kandang.
Pengujian Aplikasi Bacillus spp. dan Aktinomiset terhadap Penekanan
X. oryzae pv. oryzae dan Pertumbuhan Tanaman Padi
Pengujian Formulasi Bacillus spp. dan Aktinomiset terhadap Penekanan Populasi X. oryzae pv. oryzae
Perlakuan mikroba pada benih padi juga berpengaruh pada populasi bakteri patogen X. oryzae pv. oryzae pada bibit padi (Lampiran 1). Sebelum digunakan, sebanyak 20 benih padi diambil secara acak dari masing-masing sampel yang akan digunakan. Benih tersebut dilakukan analasis populasi X. oryzae pv. oryzae yang terbawa benih dengan cara teknik pencawanan dengan pengenceran berseri pada media YDCA. Hasil analisis menunjukkan bahwa benih yang digunakan sebagai bahan uji dalam penelitian ini rata-rata mengandung X. oryzae pv. oryzae sebanyak 2,7 × 104 cfu/gr benih padi.
Tabel 5 Pengaruh perlakuan formulasi pada benih padi terhadap populasi X. oryzae pv. oryzae pada bibit padi berumur 7 HST
Perlakuan Populasi X, oryzae pv, oryzae
(× 105 cfu/gr bibit)* KONTROL 108,00a B12 45,33c APS 7 91,00b APS 9 12,00d APS 12 23,33d B12 + APS 7 53,00c B12 + APS 9 16,00d B12 + APS 12 48,00c
* Angka yang diikuti huruf yang berbeda menunjukkan hasil yang berbeda nyata (uji selang berganda Duncan α = 0,05)
Setelah benih padi ditanam pada media perkecambahan berupa cocopeat dengan perlakuan yang berbeda, kemudian dilakukan analisis terhadap populasi bakteri X. oryzae pv. oryzae yang terkandung dalam bibit padi. Pada Tabel 4 disajikan data populasi X oryzae pv. oryzae pada bibit padi yang berumur 7 HST. Hasil menunjukkan bahwa seluruh perlakuan mikroba dapat menekan populasi X. oryzae pv. oryzae dibandingkan dengan benih yang tidak diberi perlakuan (KONTROL). Perlakuan aktinomiset dengan kode isolat APS 9 dapat menekan populasi X. oryzae pv. oryzae sebesar 88,89%. Hal ini menunjukkan bahwa
adanya sifat antagonisme yang dimiliki oleh aktinomiset terhadap bakteri X. oryzae pv. oryzae yang terdapat pada bibit padi berumur 7 HST tersebut. Jika dibandingkan dengan populasi awal X. oryzae pv. oryzae pada benih padi sebelum ditanam, yaitu 2,7 × 104 cfu, data menunjukkan bahwa bakteri X. oryzae pv. oryzae mengalami perbanyakan. Namun dengan diberinya perlakuan, perkembangan X. oryzae pv. oryzae dapat ditekan hingga 12,00 × 105 cfu, dengan bibit yang tidak diberi perlakuan mencapai 108,00 × 105 cfu.
Bakteri patogen X. oryzae pv. oryzae merupakan bakteri Gram negatif yang tidak dapat membentuk spora dalam siklus hidupnya. Madigan et al (1996) menyatakan bahwa beberapa spesies aktinomiset dapat menghasilkan antibiotik yang aktif dalam menekan perkembangan bakteri Gram negatif seperti X. oryzae pv. oryzae ini, seperti Streptomycin yang dihasilkan oleh Streptomyces griseus dan Tetracyline yang dihasilkan oleh S. aureofaciens.
Pengujian Formulasi Bacillus spp. dan Aktinomiset terhadap Pertumbuhan Bibit Padi
Adanya aktivitas mikroba pada benih padi menimbulkan keragaman dalam pertumbuhan bibit padi pada usia semai. Hasil analisis statistik dengan sidik ragam pengaruh perlakuan aktinomiset, Bacillus spp., maupun kombinasi keduanya dalam suatu bahan pembawa menunjukkan bahwa adanya pengaruh yang nyata terhadap persen perkecambahan benih padi selama masa pengamatan hingga 7 HST (Lampiran 2, 3, 4, dan 5).
Tabel 6 Pengaruh perlakuan formulasi pada benih padi terhadap persen kemunculan bibit padi
Perlakuan Persen kemunculan bibit padi*
4 HST 5 HST 6 HST 7 HST
KONTROL 6,67b 23,67bc 40,00bc 56,67c
B12 6,33b 16,67c 30,67c 56,33c
APS 7 20,67a 35,33ab 44,33abc 73,33abc
APS 9 12,67ab 29,33abc 58,33ab 79,00ab
APS 12 7,67b 15,67c 35,67bc 63,00bc
B12 + APS 7 20,67a 42,33a 64,67a 83,33a
B12 + APS 9 7,67b 17,33c 30,33c 57,00c
B12 + APS 12 16,00ab 27,00bc 46,33abc 77,00bc
* Angka yang diikuti huruf yang berbeda pada kolom yang sama menunjukkan hasil yang berbeda nyata (uji selang berganda Duncan α = 0,05)
Pada tabel 6 disajikan data kemunculan bibit padi dengan berbagai perlakuan bakteri yang diberikan terhadap benih padi. Aplikasi bakteri pada benih padi menunjukkan hasil yang beragam terhadap kemunculan bibit padi. Perlakuan kombinasi bakteri Bacillus spp, dan aktinomiset dengan kode isolat APS 7 (B12+APS7) merupakan perlakuan dengan hasil paling tinggi di antara perlakuan lain. Pada 4 HST sebanyak 20,67% bibit yang muncul. Kemudian pada 5, 6, dan 7 MST masing-masing ialah 42,33%, 64,67%, dan 83,33%. Dapat dikatakan bahwa perlakuan Bacillus spp, dan aktinomiset dapat meningkatkan persen kemunculan bibit padi sebesar 47,04%. Hal ini menunjukkan adanya akivitas mikroba yang dapat memacu proses kemunculan bibit padi hingga menyebabkan jumlah benih dengan perlakuan bakteri memiliki tingkat kemunculan bibit yang lebih banyak dibandingkan dengan benih yang tidak diberi perlakuan.
Bakteri X. oryzae pv. oryzae terdapat pada bagian bawah glume dan terkadang berada pada endosperma dari benih padi. Keberadaan bakteri pada bagian ini yang dapat mengganggu proses metabolisme dari benih tersebut (Singh dan Mathur 2004). Hal ini mengakibatkan benih yang tidak diberi perlakuan (KONTROL) memiliki tingkat kemunculan bibit yang paling rendah, karena bakteri X. oryzae pv. oryzae dapat berkembang dan mengganggu proses metabolisme benih padi. Adanya aktivitas antagonisme yang dimiliki Bacillus spp. maupun aktinomiset mengakibatkan terhambatnya perkembangan X. oryzae pv. oryzae. Hal ini yang menyebabkan benih dengan perlakuan memiliki tingkat kemunculan bibit yang lebih tinggi dibandingkan benih yang tidak diberi perlakuan (KONTROL).
Bakteri Bacillus spp., telah diketahui mampu memacu pertumbuhan bagi tanaman karena diketahui dapat membantu menghasilkan hormon pertumbuhan seperti asam indoleasetat (IAA), asam giberelat, sitokinin, dan etilen pada tanaman (Sulistiani 2009). Dalam jumlah yang sesuai, hormon tersebut dapat memacu pertumbuhan tanaman menjadi lebih baik. Aktinomiset juga merupakan bakteri yang diketahui banyak menghasilkan antibiotik yang dapat dimanfaatkan dalam menekan populasi mikroba yang dapat mengganggu proses pertumbuhan tanaman.
Tabel 7 Pengaruh perlakuan formulasi pada benih padi terhadap tinggi tajuk rata-rata bibit padi berumur 7 HST
Perlakuan Tinggi tajuk rata-rata (cm)*
KONTROL 7,64e B12 9,09cd APS 7 9,89abc APS 9 10,40a APS 12 8,66d B12 + APS 7 8,80d B12 + APS 9 9,21bcd B12 + APS 12 10,07ab
* Angka yang diikuti huruf yang berbeda menunjukkan hasil yang berbeda nyata (uji selang berganda Duncan α = 0,05)
Perlakuan aplikasi mikroba pada benih juga mengakibatkan perbedaan yang nyata pada tinggi tajuk bibit padi berumur 7 HST (Lampiran 6). Seluruh perlakuan mikroba yang diberikan pada benih mengakibatkan tinggi tajuk rata-rata bibit padi menjadi lebih tinggi dibandingkan dengan unit uji tanpa perlakuan (kontrol). Benih padi dengan perlakuan APS12 dapat meningkatkan tinggitajuk padi sebesar 13,35% dan benih yang diberi perlakuan aktinomiset dengan kode isolat APS 9 dapat meningkatkan tinggi tajuk bibit padi sebesar 26,53%. Hal ini ditunjukkan dengan data tinggi tajuk rata-rata bibit yang dimiliki, yaitu 8,66 cm dengan perlakuan APS12 dan 10,40 cm untuk perlakuan APS9, jika dibandingkan dengan benih yang tidak diberi perlakuan (kontrol) yang hanya 7,64 cm (Tabel 7).
Gambar 4 Bibit padi yang tidak diberi perlakuan (kiri) dan yang diberi perlakuan B12+APS12 (kanan)
Adanya perbedaan dalam pertumbuhan kecambah padi, baik persen kemunculan bibit maupun tinggi tajuk rata-rata, tidak hanya diakibatkan oleh adanya aktivitas produksi hormon yang dihasilkan Bacillus spp., namun kemungkinan selain menghasilkan antibiotik, aktinomiset juga dapat menghasilkan hormon pertumbuhan yang dapat memacu pertumbuhan tanaman. Igarashi et al. (2005) pernah melaporkan bahwa Streptomyces hygroscopicus S-17, salah satu anggota aktinomiset, mampu memacu pertumbuhan tomat dua kali lebih tinggi dan delapan kali lebih berat dibandingkan dengan perlakuan kontrol. Aktinomiset diketahui memproduksi toyocamycin, hormon mirip cytokinin, yang dapat memacu pertumbuhan kalus dan asam pteridic, hormon mirip auksin, yang dapat memacu perkembangan akar.
KESIMPULAN DAN SARAN
Kesimpulan
Bacillus spp. dan aktinomiset merupakan dua jenis mikroba yang dapat mengendalikan X. oryzae pv. oryzae. Dari 16 isolat aktinomiset yang diperoleh, terdapat tiga isolat yang memiliki sifat antagonis terhadap patogen X. oryzae pv. oryzae dan kompatibel terhadap Bacillus spp., yaitu APS 7, APS 9, dan APS 12. Aplikasi Bacillus spp. dan aktinomiset pada benih padi dapat menurunkan populasi patogen X. oryzae pv. oryzae pada bibit padi serta meningkatkan pertumbuhan bibit padi. Perlakuan aktinomiset dengan kode isolat APS 9 dapat menekan populasi X. oryzae pv. oryzae pada bibit padi sebesar 88,89%. Perlakuan B12+APS7 merupakan perlakuan dengan hasil persen kemunculan bibit paling tinggi di antara perlakuan lain, yaitu 83,33% pada 7 HST, diikuti dengan perlakuan APS 9 dengan 79%. Perlakuan Bacillus spp. dan aktinomiset juga mampu memicu pertumbuhan tajuk tanaman sebesar 13,35 untuk perlakuan APS12 hingga 26,53% untuk perlakuan APS9 pada 7 HST.
Saran
Penelitian ini dapat dikembangkan, sehingga diperlukan adanya kajian lebih lanjut mengenai hal-hal yang mendukung suatu formulasi mikroba dalam bentuk kering, seperti eksplorasi aktinomiset dari berbagai habitat asal yang berbeda, kandungan dan konsentrasi berbagai bahan pembawa yang tepat, daya tahan mikroba dalam masa penyimpanan, hingga pengujian in vivo formulasi tersebut dengan waktu pengamatan lebih panjang.
DAFTAR PUSTAKA
Agrios GN. 1988. Plant Pathology. Gainesville : Academic Press, Inc.
Ainsworth GC. 1971. Ainsworth & Bisby’s Dictionary of the Fungi. Surrey: Commonwealth Mycological Institute.
Baker KF dan Cook RJ. 1974. Biological Control of Plant Pathogens. San Francisco: Freeman & Co.
Charoensopharat K, Thummabenjapone P, dan Thammasirirak S. 2007. Study of Antimicrobial Substance Produced by Streptomyces ssp., 87 [abstrak]. Di Dalam The 5th International Symposium on Biocontrol and Biotechnology; Nong Khai, Thailand, 1-3 November 2007. Nong Khai, Thailand: King Mongkut’s Institute of Technology Ladkrabang and Khon Kaen University. Hlm 37. O-28.
Crawford DL, James ML, John MW, dan Margaret AO. 1993. Isolation and Characterization of Actinomycete Antagonis of a Fungal Root Pathogen. Applied and Environmental Microbiology 11: 3899-3905.
Desmawati. 2006. Pemanfaatan Plant Growth Promoting Rhizobacteria (PGPR) Prospek yang Menjanjikan dalam Berusahatani Tanaman Holtikultura [tesis]. Bogor : Sekolah Pascasarjana IPB.
Dirmawati SR. 2004. Kajian Keefektifan Beberapa Komponen Pengendalian Ramah Lingkungan terhadap Penyakit pustul Bakteri Kedelai [disertasi]. Bogor: Sekolah Pascasarjana IPB.
Fravel DR, Connick WJ, dan Lewis JA. 1998. Formulation of Microorganisms to Control Plant Diseases. Di Dalam: Burges HD, editor. Formulation of Microbial Biopesticides. London: Kluwer Academic Publisher. hlm 187-202.
Igarashi Y, S Miura, M Azumi, T Furumai, dan R Yoshida. 2005. Studies on Plant-associated Actinomycetes and Their Secondary Metabolites. Journal of Bioscience, Biotechnology, and Biochemistry. 21: 1427-1441.
Janse JD. 2005. Phytobacteriology: Principle ang Practice. Cambridge: CAB International Publishing.
Jing L dan Qian Y. 2007. Purification and Properties of Antifungal Protein Produced by Bacillus Subtilis B29 [abstrak]. Di Dalam The 5th International Symposium on Biocontrol and Biotechnology; Nong Khai, Thailand, 1-3 November 2007. Nong Khai, Thailand: King Mongkut’s Institute of Technology Ladkrabang and Khon Kaen University. Hlm 37. O-27.
Khan MR, Saha ML, dan Zuha SB. 2008. Bacteria and Actinomycetes Growing on Floppy and Compact Disc Under Ambient Conditions. Bangladesh J. Bot. 37(1): 7-14.
Kunoh H. 2002. Endophytic Actinomycetes: Attractive Biocontrol Agents. J. Gen. Plant Pathol. 68: 249-252.
Madigan MT, John MM, dan Jack P. 1996. Brock Biology of Microorganisms 8th edition. New Jersey: Prentice-Hall, Inc.
McQuilken MP, Halmer P, dan Rhodes DJ. 1998. Application of Microorganisms to Seeds. Di Dalam: Burges HD, editor. Formulation of Microbial Biopesticides. London: Kluwer Academic Publisher. hlm 255-285.
Muis, Amran. 2006. Biomass Production and Formulation of Bacillus subtilis for Biological Control. Indonesian Journal of Agriculture Science. 7(2): 51-56. Murdiyah S. 2008. Daya hambat Streptomyces sp terhadap pertumbuhan jamur
patogen tumbuhan Fusarium sp dan Rhizoctonia sp. digilib.unej.ac.id [23 September 2008].
Nakaew N, Wasu P, dan Saisamorn L. 2009. First Record of the Isolation, Identification and Biological Activity of a New Strain of Spirillospora albida from Thai Cave Soil. Actinomycetologica 23:1–7
Nelson LM. 2004. Plant Growth Promoting Rhizobacteria (PGPR): Prospect for New Inoculant. Plant Management Network. www.plantmanagement network.org/pub/cm/review/2004/rhizobacteria/ [14 Desember 2010]. Patil R, Jeyasekaran G, Shanmugam SA, dan Shakila RJ. 2001. Control of
Bacterial Pathogens, Associated with Fich Diseases, by Antagonistic Marine Actinomycetes Isolated from Marine Sediment. Journal of Marine Science. 30(4): 264-267.
Putra MC, Solichah YR, Murgiyanto F, Aminudi, dan Nurhasanah Y. 2009. Formulasi Bacillus subtilis pada Tepung Singkong sebagai Probiotik Tanaman. [Laporan Penelitian]. Bogor: Institut Pertanian Bogor.
Rifaat HM. 2003. The Biodiversity of Actinomycetes in the River Nile Exhibiting Antifungal Activity. Journal of Mediterranean Ecology. 4:5-7.
Sabaratnam S dan James A. Traquair. 2001. Formulation of a Streptomyces Biocontrol Agent for the Supression of Rhizoctonia Damping-off in Tomato Transplant. Journal of Biological control 23: 245-253.
Schaad NW, Jones JB, dan Chun W. 2000. Laboratory Guide for Identification of Plant Phatogenic Bacteria. Minnesota: APS Press.
Singh D dan Mathur SB. 2004. Histopathology of Seed Borne Infection. Florida: CRC Press.
Sonenshein AL. et al. 2002. Bacillus subtilis and Its Closest Relatives from Genes to Cell. Washington DC :ASM Press.
Stevenson IL. 1956. Antibiotic Activity of Actinomycetes in Soil and their Controlling Effects on Root-rot of Wheat. Journal of Genetic Microbiology. 14, 440498
Sulistiani. 2009. Formulasi Spora Bacillus subtilis sebagai Agens Hayati dan PGPR (Plant Growth Promoting Rhizobacteria) pada Berbagai Bahan Pembawa [skripsi]. Bogor: Departemen Proteksi Tanaman Fakultas Pertanian IPB.
Suwahyono U. 2010. Biopestisida: Cara Membuat dan Petunjuk Penggunaan. Jakarta: Penebar Swadaya.
Suwandi U. 1993. Perkembangan Antibiotik. http://www.kalbe.co.id/files/cdk/ files/19PerkembanganAntibiotik083.pdf/19PerkembanganAntibiotik083.ht ml. [23 September 2008].
Waites WM et al. 1970. Sporulation in Bacillus subtilis: Correlation of Biochemical E vents with Morphological Changes in Asporogenous Mutants. Journal of Biochemistry. 118: 667-676.
Zhang JX dan Tambong JT. 2009. Evaluation of Seed and Soil Treatments with Novel Bacillus subtilis Strains for Control of Soybean Root Rot Caused by Fusarium oxysporum and F. graminearum. Plant Dis. 93:1317-1323.