Role of In Vitro System as Continuous Betalain Sources
Retno Mastuti
Biology Department, Faculty of Mathematics and Natural Sciences, University of Brawijaya, Malang, Indonesia ([email protected])
Abstract
Plants are sources of many important secondary metabolites including pigments. Betalain, one of plant major pigments is of growing interest for colorant of food and pharmaceutical industries. This natural colorant has been concerned as an alternative for synthetic colorants due to its positive effects on health. Recent reports show that betalains have high anti-oxidative, free radical scavenging activities and potential for anti malaria. In higher plants betalain occurrence is restricted to 13 families of the order Caryophyllales. Their occurence is mutually exclusive to that of the anthocyanins. Although the major pigment group shows wide occurence in different plant tissues, the production is very affected by environmental conditions. Therefore, an in vitro system under controlled conditions is one promising alternative pigments source for all seasons. Plant cells and tissue cultures can be established routinely under sterile conditions from explants, such as plant leaves, stems, and roots for both multiplication and extraction of secondary metabolites including pigments. Some in vitro strategies have been applied to improve betalain production. Strain improvement, methods for the selection of high-producing cell lines, and medium optimizations could enhance secondary metabolite production. In addition, this paper will discuss hairy root cultures for pigment production, precursor addition for betalain improvement, elicitation of in vitro products, and bioreactors scaling up of production.
Keywords: betalains, plant in vitro systems, optimization strategies
I. INTRODUCTION
The plant major pigments are chlorophyll, carotenoids, anthocyanins and betalains. Except chlorophyll, the three families of pigments play important ecological functions, for example in the attraction of pollinators and seed dispersal animals. All of them are secondary metabolite which economically important such as in food and medicinal industry. Interest in betalains has grown since their antiradical activity was characterized (1), and they are widely used as additives in the food industry because of their natural colorant properties and absence of toxicity (2). Consequently, the sustain availability of betalain pigments become important. However, the distribution of secondary metabolites in plants is far more restricted than that of primary metabolites. Also, the secondary metabolites often accumulate in the plant in small quantities, sometimes in specialized cells which caused the difficulty in their extraction. Although extraction of secondary metabolites from plants is still commercially important a large number of these metabolites are difficult or impossible to synthesize at economic values (3). Therefore, development of alternatives to the intact plant for the production of plant secondary metabolites has been intensively approached. In the past three decades, research has been concentrated on the use of plant cell and tissue cultures. Biotechnological approaches, specifically plant tissue culture plays a vital role in search for alternatives to production of desirable compounds from plants (4). This method can provide a continuous, reliable source of natural products. In
vitro system offers several advantages over field cultivation: it is independent of geographical and seasonal variations, environmental factors, and political interference; in addition, it allows optimal and stable growth conditions, voluntary modulation of growth parameters, and constant quality control (5-6). It also eliminates negative biological influences (microorganisms and insects) that affect secondary metabolites production in nature; and possibility to select cultivars with higher production of secondary metabolites (7). Moreover, colourants produced in this way are classified as 'natural' rather than 'nature-identical', which increases their desirability to customers. In some cases, the yield per gram fresh weight of secondary metabolites in plant cell culture may exceed that which is found in nature. Therefore, a priority in current and future efforts towards sustainable conservation and rational utilization of biodiversity is searching for new plant-derived chemicals (8). Plant-produced secondary compounds have been incorporated into a wide range of commercial and industrial applications. Different strategies using cell cultures systems have been extensively studies with the objective of improving the production of bioactive secondary metabolites.
amount of secondary metabolites in plant cell cultures was accompanied by emphasizing the biochemical and molecular research on the secondary metabolism of plants (11-16). These lead to the successful manipulation of secondary metabolism and significantly increase the amounts of the compound(s) production. Now, it seems that any substances of plant origin can be produced by plant cell culture techniques. Identification of cell lines that can produce amounts of compounds equal or even higher than those in the plant from which they derived has been possible.
II. BETALAIN – BIOCHEMISTRY, DISTRIBUTION AND CHARACTERS
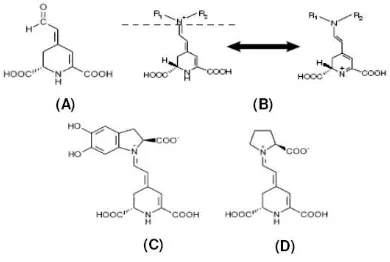
Betalains, known for a long time as safe colorants for food or other industrial purposes. Betalains are water-soluble nitrogen-containing pigments which also known as immonium derivatives of betalamic acid (Fig. 1A) (17).
Fig. 1. Chemical structures of betalamic acid (A), betalain and its resonance (B), betanidin (C), and indicaxanthin (D) (17,18)
They are synthesized from tyrosine by the condensation of betalamic acid (19) The structure below the dashed line is present in all betalain molecules (Fig. 1B). Betalains are commonly classified based on their structural characteristics and divided in two groups red-violet betacyanin and yellow betaxanthin depending on the identity of the R1 and R2 residues. They are immonium conjugates of betalamic acid with cyclo-dopa and amino acids or amines, respectively, respectively. The first betalains identified by chemical means are: betalamic acid (Fig. 1A), the chromophore of all betalains, betanidin (Fig. 1C), the main aglycone of all betacyanins, and indicaxanthin (Fig. 1D) (20), a proline-containing betaxanthin. To date, in nature comprise approximately 50 red betacyanins and 20 yellow betaxanthins (21), all with the same basic structure.
Betalains are restricted to higher plants to most families of the order Caryophyllales (old name Centrospermae); 13 betalain producing families have
been identified: Achatocarpaceae, Aizoacea, Amaranthaceae, Basellaceae, Cactaceae, Chenopodiaceae, Didieraceae, Holophytaceae, Hectorellaceae, Nyctaginaceae, Phytolaccaceae, Portulacaceae and Stegnospermataceae (22) (Table 1). The other two families, the Caryophyllaceae and Molluginaceae accumulate anthocyanins. The betalain and anthocyanin pathways in flowering plants has been known as evolutionary mechanisms leading to the mutual exclusion (23-24). The Caryophyllales-specific occurrence of betalains is a prominent example of the chemotaxonomic relevance of plant secondary products.
Table 1. Classification of Caryophyllales (22)
Suborder Family Examples of genus Chenopodiinae
(Betalain-producing anthocyanin-free taxa)
Achatocarpaceae Achataocarpus
Aizoaceae Dorotheanthus, Mesembryanthemum
Amaranthaceae Amaranthus, Celosia, Gomphrena
Basellaceae Basella
Cactaceae Mammilaria, Opuntia, Schlumbergera
Chenopodiaceae Beta, Chenopodium, Salicornia, Spinacia
Didiereaceae Decaryia, Didierea
Halophytaceae Halophytum
Hectorellaceae Hectorella
Nygtaginaceae Bougainvillea, Mirabilis
Phytolaccaceae Gisekia, Phytolacca
Portulacaceae Claytonia, Portulaca
Stegnospermataceae Stegnosperma
Chenopodiinae (Betalain-free anthocyanin-producing taxa)
Caryophyllaceae Dianthus, Melandrium, Silene
Molluginaceae Mollugo, Pharnaceum
Betalains are found in different plant organs namely flowers, fruits and occasionally in vegetative tissues of plants and they are accumulated in cell vacuoles, mainly in epidermal and subepidermal tissues (Table 2) (25). However, they are sometimes accumulated in plant stalks such as in the underground parts of red beet. Many plants accumulate betalains but only two (Beta vulgaris
and the prickly pear Opuntia ficus-indica) are approved to be used in food (26-27). Betalains are also present in some higher fungi Amanita, Hygrocybe, and Hygrosporus (28). The functions of betalains in plant flower and fruit colouration are obvious. Meanwhile, their role in fungi is unknown. Betalain derived from red beetroot is approved to be used in food industries as food additive in United State (21 CFR 73.40) and in European Union (E162).
Table 2. Betalain distribution in plant structure (25)
Plant Structures Color Produced Examples Flowers Red, yellow, pink
and orange
Aizoaceae and Potrulacaceae plants Fruits Yellow, red and
purple
Prickly pear
Roots Red-purple Red-beet root Bracts Wide ranges of
colors
Bougainvillea sp. Seeds Yellow and red,
among others
III. ACCUMULATION OF BETALAIN AND CELL GROWTH
Many secondary metabolites accumulate in specific tissues and cell of higher plants, or at specific stages during the growth of cultured cells. The accumulation of most secondary metabolites in cultured cells is maximum during the stationary phase of growth. On the contrary, peaks of accumulation of secondary products during the logarithmic phase of growth have been observed in only a few cases. Accumulation of betacyanin was suppressed when growth of cells in suspension cultures of Phytolacca americana was inhibited by aphidicolin (an inhibitor of DNA synthesis) (29). Betacyanin content in cell suspension culture of B. vulgaris also increased in parallel with the cell growth during the log phase (30). Peaks of accumulation of betalains during the logarithmic phase were also observed in suspension cultures of
Chenopodium rubrum (31). Close correlation
between growth and accumulation of betacyanin was also reported in suspension cultures of P. americana
(32). Subsequently, it was suggested a positive correlation between the accumulation of betacyanin and cell proliferation (29).
A similar results has been reported for cell suspension from the violet callus of B. vulgaris (35). Lack of phosphate or addition of aphidicolin to the medium suppressed both the division of cells and the accumulation of betacyanin. Conversely, re-addition of phosphate and removal of aphidicolin initiated both process. Tracer experiments using labeled tyrosine revealed that the incorporation of radioactivity from tyrosine to betacyanin was inhibited while that from DOPA was not. Therefore, it is concluded that the step from DOPA to betacyanin is not coupled with cell division, but that from tyrosine to DOPA is coupled (34). Furthermore such a relationship between the betacyanin accumulation and cell growth has been observed in cell suspension culture of B. vulgaris (30) and
Portulacca (35).
IV. MAXIMIZATION OF SYNTHESIS AND PRODUCTION
Many efforts have focused on the stimulation of biosynthetic activities of cultured cells using various methods. Since the tissue culture cells typically accumulate large amounts of secondary compounds only under specific conditions the maximization of accumulation secondary metabolites requires: manipulating the parameters of the environment and medium; selecting high yielding cell clones precursor feeding and elicitation.
A. Optimization of medium composition
Since the economically viable is prerequisite, it is important to develop methods that would allow for consistent generation of high yields of products (36). Several products were found to be accumulating in cultured cells at a higher level than those in native plants through optimization of cultural conditions (37). There is no similar trend of availability of phosphate ion in affecting betalain content. In B. vulgaris cv Detroit Dark Red hairy root cultures there is phosphate ions reduction from initial levels of 120 to 10-15 mg L-1 after the start of the exponential phase of cultivation (38). However the same culture in phosphate-free medium resulted in total betalain contents rising to 19 g L-1 : 4.8 times higher than the amounts accumulated in a standard MS medium (39).
The concentration of nitrate ions exhibited different color phenotypes of B. vulgaris cv. Bikores Monogerm cell lines (33). While combination of nitrogen concentrations with some concentrations in microelement increased betacyanin production in red beet cell suspension (40), there was no significant effect on betalain accumulation in various nitrogen concentrations (39). Ammonium ions are utilized more rapidly than nitrate ions but no significant amount of either ammonium or nitrate ions are consumed in the first 5 days of B. vulgaris hairy root cultures (41). In suspension cultures of P. americana
L., betacyanin accumulation per cell increased with increasing total nitrogen concentration (initial NH+4:NO−3 ratio 1:2) in the range 0–40 mM and then remained almost constant in the range 40–80 mM. (42).
Microelements in medium also influenced betalain production. Increasing Fe2+ concentration up to 20-fold higher than that in the standard LS medium markedly increased betacyanin content without any affect on cell growth of B. vulgaris cell suspension (30). Apparently, concentration of microelement was significantly affected by basal medium. Removing of boron, iodine, manganese and molybdenum was found to decrease both the cell growth and betacyanin content whereas absence of copper and cobalt did not show any negative effect on red beet cell suspension using LS basal medium (43). On the other hand in Gamborg B5 medium except manganese the addition high concentration of copper, iron, molybdenum, zinc and cobalt showed positive effect on betalain production.
adding DOPA (44) and specific amino acids (45) does not increase the betacyanin content.
The type of inoculums strongly influences betacyanin production. The cell culture of Portulaca grandiflora derived from the basipetal cut of a hypocotyls segment is superior to the cell culture of acropetal origin with regard to betacyanin concentration. These differences indicate the polarization of physiological conditions in the hypocotyls segments (46).
B. Selection of highly productively cell lines
Plant cell cultures represent a heterogenous population in which physiological characteristics of individual plant cells are different (37) Proper selection of productive cells and cultural conditions results in accumulation of products in high level. Not all cells express the pigmented phenotype, giving rise to variegated patches (47). The pattern of patch formation is different from one callus to another, although all calluses stem from the same original clone. Therefore, the selection of high-producing strains is necessary. Secondary callogenesis of B. vulgaris cv. Bikores Monogerm, on media with various ratios of auxin (2,4-D) and cytokinin (BAP), showing white, green, yellow, orange, red and violet pigmentations, a pattern of coloration similar to that exhibited by the flowers of betalain production species (Fig. 1). It represents the overall pathways of betalain metabolism (48). It shows that genetic pathways which repressed at the whole plant level under given conditions may be activated in tissue culture.
Fig 1. Five basis inherited stable phenotypes of red beet cell cultures. Left to right: green, yellow, orange, red, violet (48)
C. Hairy root cultures
Hairy root system based on inoculation with
Agrobacterium rhizogenes has become popular as a method of producing secondary metabolites synthesized in plant roots (49). They often grow as fast as or faster than plant cell cultures (50) do not require hormones in the medium. In contrast to callus cultures, they are usually considered as genetically stable. The secondary metabolites produced by hairy roots are the same as those usually synthesized in intact parent roots, with similar or higher yields (51) The red beet hairy root culture obtained from cv. Detroit Dark Red is a prospective producer of betalain pigments because it
has stable morphological, growing and biosynthetic characteristics and biosynthesizes 13.2 mg/g DW pigments (4.4 mg/g DW are betacyanins and 8.8 mg/g DW betaxanthins, respectively) (52).
D. Elicitation
The accumulation of secondary metabolites in plants is a common response of plants to biotic and abiotic stresses. Their accumulation can be stimulated by biotic and abiotic elicitors. Use of elicitors of plant defense mechanisms, i.e. elicitation, has been one of the most effective strategies for improving secondary metabolites production in plant tissue culture. The effect of elicitors depends on many factors, such as the concentration of the elicitor, the growth stage of the culture at the time of elicitation and the contact time of elicitation. Betacyanin levels in suspension cultures of Portulaca sp. Cv. Jewel increased 2.6- and 1.8-fold due to the addition of biotic elicitors methyl jasmonate and B-glucan, respectively (53). While in a wide range of biotic and abiotic elicitors the Ca2+ was reported as the strongest abiotic elicitor which induced a 2.92-fold increase in betalain content in B. vulgaris L. hairy root cultures (54).
E. Bioreactors scaling up
The most important source of betalain pigments is B. vulgaris root. Until now it has been the only one commercially exploited for betalain production. However, alternative sources are found in plants from the Amaranthaceae and Cactaceae families. Another alternative source is plant cell culture in bioreactors, although optimization of pigment production seems necessary (55). Many different types of bioreactor systems for cultivating plant cultures in vitro are available, and choosing the most appropriate for specific applications is of great importance (56). In hairy root culture bioreactors can be roughly divided into three types: liquid-phase, gas-liquid-phase, or hybrid reactors that are a combination of both. Therefore, the ideal type of in vitro system for producing betalains from any kind of plant cells or tissues needs to be carefully investigated on a case-by-case basis. Cell suspension cultures and hairy roots of B. vulgaris have been grown in shake flasks (40,57) and in several types of reactors. Until recently, the shake flask method of cultivation offered higher betalain yield than cultivation of cells in bioreactors (58). Cultivation of
This technology can also produce novel metabolites since new molecules which have not been found previously in plants have been produced by cell cultures. Plant tissue culture techniques offer the rare opportunity to alter the chemical profile of a phytochemical product, by manipulation of the chemical or physical microenvironment, to produce a compound of potentially more value for human use.
REFERENCES
[1] J. Escribano, M. A. Pedreno, F. Garcıa-Carmona and R. Mun˜oz, “Characterization of the antiradical activity of betalains from Beta vulgaris L. roots”, Phytochem. Anal., no. 9, pp. 124–127, 1998.
[2] S. J. Schwartz, J. H. von Elbe, M. W. Pariza, T. higher-plant cell cultures: biotechnological and economic aspects”. Options Méditerranéennes – Sérié Séminaires, no. 14, pp. 59-74, 1991.
[4] R. S. Rao and G. A. Ravishankar, “Plant tissue
cultures; chemical factories of secondary
metabolites”, Biotechnol. Adv., no. 20, pp. 101-153, 2002.
[5] D.A. Moreno, C. García-Viguera, J.I. Gil, A. Gil-Izquierdo, “Betalains in the era of global agri-food science, technology and nutritional health”,
Phytochem. Rev., no. 7, pp. 261–280, 2008.
[6] S. Ramachandra Rao, G.A. Ravishankar, “Plant cell cultures:Chemical factories of secondary metabolites”, Biotechnol. Adv., no. 20, pp.101–153, 2002.
[7] V. Mulabagal and H. Tsay, “Plant Cell Culture – An Alternative and efficient source for the Production of Biologically Important Secondary Metabolites”, Int. J. Appl. Sci. Engineer., vol. 2, no. 1, pp. 29-48, 2004. [8] Philipson JD (1990). Plants as source of valuable
products. In: B.V.Chalwood and M.J. Rhodes (Eds.), Secondary products from plant tissue culture, Oxford, Clarendon Press. pp.1-21.
[9] B. Kaul, and E. J. Staba, “Ammi visnaga L. Lam. tissue cultures: multiliter suspension growth and examination for furanochromones”, Planta Medica, Journal of Medicinal Plant Research, pp.145-56, 1967.
[10]M. R. Heble, S. Narayanaswamy, and M.S. Chadha, “Diosgenin and sitosterol isolation from Solanum xanthocarpum tissue cultures”, Science, no. 161, pp. 1145, 1968.
[11]M. Fowler, “Plant cell biotechnology to produce desirable substances”, Chemistry and Industry, no. 7, pp. 229-33, 1981.
[12]M. W. Fowler, “Time for plant cell culture?”, Nature, vo1.307, no. 5951, pp. 504, 1984.
[13]M. W. Fowler, “Plant-cell culture: natural products and industrial application”, Biotechnol. Genetic Engineer. Rev., no. 2, pp. 41-67, 1984.
[14]M. Misawa, Production of useful metabolites. Adv. Biochem. Eng./Biotech., no. 31, pp. 59-88, 1985. [15]J. Berlin, S. Sieg, D. Strack, M. Bokern and H.
Harms, “Production ofbetalains by suspension cultures of Chenopodium rubrum L.”, Plant Cell Tiss. Org. Cult., no. 5, pp. 163–174, 1986.
[16]A. Stafford, P. Morris, M. W. Fowler, “Plant cell biotechnology: a perspective”, Enzyme Microb. Technol., no. 8, pp. 578-87, 1986.
[17]F. Delgado-Vargas and O. Paredes-López, Natural colorants for food and nutraceutical uses. CRC Press. Boca Raton, Florida, pp. 192 – 211, 2003. [18]D. Strack, T. Vogt and W. Schliemann, “Recent
advances in betalain research”, hyrocem. No. 62, pp.247-269, 2003
[19]E. Grotewold, “The genetics and biochemistry of floral pigments”, Annual Review of Plant Biology, no. 57, pp. 761-780, 2006.
[20]M. Piattelli, L. Minale, ”Pigments of centrospermae. 1. Betacyanins from Phyllocactus hybridus Hort. and
Opuntia ficus indica Mill.”, Phytochem., no. 3, pp. 307–311, 1964.
[21]F. J. Francis, Anthocyanins and betalains. In F.J Francis (Ed.), Colorants (pp. 55–66). St Paul, MN: Eagan Press. (1999).
[22]J. S. Clement and T. J. Mabry, “Pigment evolution in the Caryophyllales: A systematic overview”, Bot Acta, no. 109, pp. 360–367, 1996.
[23]Kimler L, Mears J, Mabry TJ, Roesler H (1970) On thequestion of the mutual exclusiveness of betalains andanthocyanis. Taxon 19:875–878
[24]H. A. Stafford, “Anthocyanin and betalain: evolution of the mutually exclusive pathways”, Plant Science, no. 101, pp. 91-98, 1994
[25]F. Delgado-Vargas, A. R. Jiménez, and O. Paredes-López, “Natural pigments:carotenoids, anthocyanins, and betalains — characteristics, biosynthesis, processing and stability”, Critical Reviews in Food Science and Nutrition, no.40, pp. 173–289, 2000. [26]R. L. Jackman, and J. L. Smith, Anthocyanins and
betalains, in Natural Food Colorants. G.A.F. Hendry and J.D. Houghton, Eds. Chapman & Hall, New York, pp. 244–310, 1996.
[27]T. J. Mabry, A. Taylor and B. I. Turner, “The betacyanins and their distribution” Phytochemistry, no. 2, pp. 61–64, 1963.
Komamine, “Accumulation of Betacyanin in
Phytolacca americana Cells and of Anthocyanin in Vitis sp. Cells in Relation to Cell Division in Suspension Cultures”, Plant Cell Physiol., vol. 31, no. 2, pp. 267-271, 1990.
[30]T. Akita, Y. Hina, T. Nishi, “Production of betacyanins by a cellsuspension culture of table beet (Beta vulgaris L.)”, BiosciBiotech Biochem no. 64, pp.1807–1812, 2000.
[31]J. Berlin, S. Sieg, D. Strack, M. Bokern and H. Harms, “Production of betalains by suspension cultures of Chenopodium rubrum L.”, Plant Cell Tiss. Org. Cult., no. 5, pp. 163-174, 1986.
[32]M. Sakuta, T. Takagi and A. Komamine, “Growth related accumulation of betacyanin in suspension cultures of Phytolacca Americana L.,” Journal of Plant Physiol, no. 125, pp. 337-343, 1986.
[33]R. Leathers, C. Davin, and J. Zrÿd, “Betalain
betacyanin and anthocyanin in relation to cell division activity in suspension cultures”, Plant Cell Tis. Org. Cult., no. 38, pp. 167-169, 1994.
[35]M. Bhuiyan, K. Murakami and T. Adachi, “Variation in betalain content and factors affecting the biosynthesis in Portulaca sp. “Jewel’ ceel cultures”, Plant Biotechnology, vol. 19, no. 5, pp. 369-376, 2002.
[36]J. Berlin and F. Sasse, “Selection and screening techniques for plant cell cultures”, Advanced Biochemistry and Engineering, no. 31, pp:99-132, 1985.
[37]V. Mulabagal and H. Tsay, “Plant Cell Culture – An Alternative and efficient source for the Production of Biologically Important Secondary Metabolites, International Journal of Applied Science and Engineering, vol. 2, no. 1, pp. 29-48, 2004
[38]A. Pavlov, V. Georgiev and M. Ilieva, “Betalain biosynthesis by red beet (Beta vulgaris L.) hairy root culture. Process Biochem., no. 40, pp. 1531-1533, 2005.
[39]M. Taya, K. Yakura, M. Kino-Oka and S. Tone, Influence of medium constituent on enhancement of pigment production by batch culture of red beet hairy roots. J. Fermen Bioeng, no. 77, pp. 215-217, 1994. [40]T. Akita, Y. Hina and T. Nishi, “New medium
composition for highbetacyanin production by a cell suspension culture of table beet (Beta vulgaris L.)”, Biosci Biotech Biochem., no. 66, pp. 902–905, 2002. [41]K. S. Shin, D. Chakrabarty, Y. J. Ko, S. S. Han and
K. Y. Paek, “Sucrose utilization and mineral nutrient uptake during hairy root growth of red beet (Beta vulgaris L.) in liquid culture”, Plant Growth Reg. no. 39, pp. 187–193, 2003.
[42]M. Sakuta, T. Takagi and A. Komamine, “ Effects of nitrogen source on betacyanin accumulation and growth in suspension cultures of Phytolaca americana”, Physiologia Plantarum, no. 71, pp. 459-463., 1987
[43]T. Akita, Y. Hina and T. Nishi, “Effect of zinc deficiency onbetacyanin production in a cell suspension culture of table beet (Beta vulgaris L.)”, Biosci Biotech Biochem., no. 65, pp. 962–965, 2001. [44]J. Berlin, S. Sieg, D. Strack, M. Bokern and H.
Harms, “Production of betalains by suspension cultures of Chenopodium rubrum L.”, Plant Cell Tiss. Org. Cult., no. 5, pp. 163-174, 1986.
[45]J. Hempel and H. Bo¨hm, “Betaxanthin pattern of hairy roots from Beta vulgaris var. lutea and its alteration by feeding of amino acids”, Phytochem., no. 44, pp. 847–852, 1997.
[46]W. Schroeder and H. Bohm, “Betacyanin
concentrations in young cell cultures from Portulacca grandiflora an analysis of variation. Plant Cell Reports, no. 3, pp. 14-17, 1984
[47]P. A. Girod and J. P. Zryd, “Cloal variability and light induction of betalain synthesisi in red beet cell cultures. Plant Cell Rep., no. 6, pp. 27-30, 1987. [48]P. A. Girod and J. Zrÿd, “Secondary metabolism in
cultured red beet (Beta vulgaris) cells: differential
regulation of betaxanthin and betacyanin
biosynthesis”, Plant Cell Tiss. Org. Cult., no. 25, pp. 1–12, 1991.
[49]J. Palazon, M. T. Pinol, R. M. Cusido, C. Morales and M. Bonfill, “Application of transformed root
technology to the production of bioactive
metabolites. Recent Res. Dev. Plant Phys, no. 1, pp: 125-143, 1997.
[50]H. E, Flores, J. M. Vivanco, V. M. Loyola-Vargas, “Radicle biochemistry: the biology of root-specific metabolism”, Trends Plant Sci., no. 4, pp. 220–226, 1999.
[51]N. Sevon, K. M. Oksman-Caldentey, “Agrobacterium rhizogenesmediates transformation: Root cultures as a source of alkaloids”, Planta Med., no. 68, pp. 859-868, 2002.
[52]A. Pavlov, P. Kovatcheva, V. Georgiev, I. Koleva and M. Ilieva, “Biosynthesis and Radical Scavenging Activity of Betalains during the Cultivation of Red Beet (Beta vulgaris) Hairy root Cultures”, Z. Naturforsch, no. 57c, pp. 640-644, 2022.
[53]N. H. Bhuiyan and T Adachi, “Stimulation of betacyanin synthesis through exogenous methyl jasmonate and other elicitors in suspension-cultured cells of Portulaca”, J Plant Physiol., no. 160, pp. 1117–1124, 2003.
[54]Savitha BC, Thimmaraju R, Bhagyalakshmi N, G. A. Ravishankar, “Different biotic and abiotic elicitors influence betalain production in hairy root cultures of Beta vulgaris in shake-flaskand bioreactor”, Process Biochem., no. 41, pp. 50–60, 2006.
[55]D. Pavokovik and M. Krsnik-Rasol, “Complex Biochemistry and Biotechnological Production of Betalains”, Food Technol. Biotechnol., vo.49, no. 2, pp. 145–155, 2011.
[56]Eibl R, Eibl D (2002) Bioreactors for plant cell and tissue cultures. In:Oksman-Caldentey KM, Barz WH (eds) Plant Biotechnology and Transgenic Plants. Marcel & Dekker, New York, pp 163–200.
[57]G. Trejo-Tapia, A. Jimenez-Aparicio, M. Rodriguez, G. Sepulveda, G. Salcedo, B. Martinez,Influence of medium constituents on growth and betalain production in cell suspension cultures of Beta vulgaris, Asia-Pacific J. Mol. Biol. Biotechnol., no. 7, pp. 167–172, 1999.
[58]B. Neelwarne, R. Thimmaraju, “Bioreactor for cultivation of red beet hairy roots and in situ recovery of primary and secondary metabolites”, Eng. Life Sci. no. 9, pp.227–238, 2009.