EKSPRESI GEN PENYANDI ALGINAT
Sargassum polycystum
C.
Agardh ASAL PANTAI MANGANTI KEBUMEN
Oleh
Dra. Dwi Sunu Widyartini, MSi
Fakultas Biologi, Universitas Jenderal Soedirman. Jl. Dr. Soeparno 63, Purwokerto,
Banyumas 53122
Tel./Fax. Tel.+62-281638794,Fax: +62-281-631700, E-mail:
[email protected]
ABSTRAK
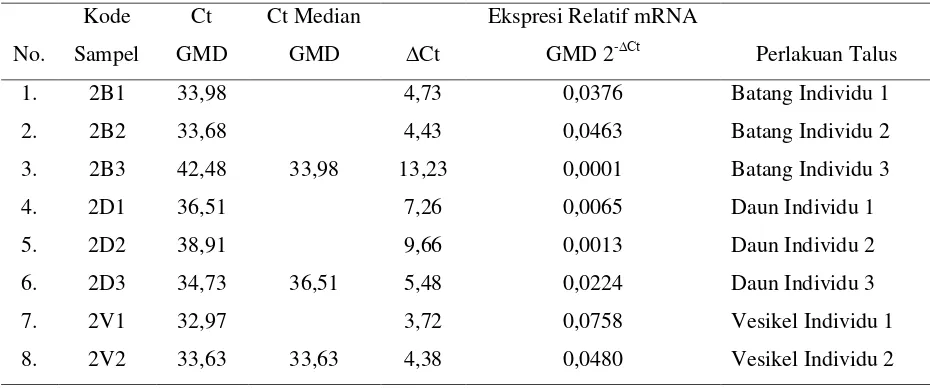
Sargassum polycystum C. Agardh merupakan rumput laut penghasil alginat. Kemampuan talus rumput lautini dalam menghasilkan alginat, dipengaruhi oleh aktivitas enzim yang disandi oleh gendan ekspresinya dipengaruhi oleh faktor lingkungan. Penelitian ini bertujuan untuk mengetahui ekspresi gen penyandi alginat S.polycystum yang diisolasi dari bentuk talus berbeda, sehingga dapat diiinterpretasikan bagian talus yang menunjukkan ekspresi gen paling tinggi. Penelitian ini menggunakan metode survei, pengambilan sampel secara purposive random sampling di perairan Pantai Manganti Kebumen. Untuk mengidentifikasi kandidat gen GDP-Man dehidrogenase, dengan urutan genom Ectocarpus siliculosus dandiamplifikasi oleh PCR. Pengamatan ekspresi gen berdasarkan kurva 2-ΔCt sehingga dapat ditentukan ekspresi relatif gen tertinggi dalam penyandi alginat. Hasil penelitian menunjukkan bahwa ekspresi relatif mRNA GDP mannose dehydrogenase pada S. polycystum berkisar 0,000125 - 0,076415. Dengan nilai median bentuk talus batang 0,02825433 (min. 0,000125 – maks. 0,046714), pada bentuk talus daun 0,01012533 (min. 0,001245
– maks. 0,022561) dan pada bentuk talus vesikel 0, 04733100 (min. 0,017217 – maks. 0,076415). Terdapat perbedaan ekspresi relatif mRNA GDP mannose dehydrogenase yang signifikan antar bentuk talus S. polycystum. Ekspresi tertinggi pada talus vesikel.
Kata kunci: bagian talus; ekspresi gen; perairan Manganti; S. polycystum
ABSTRACT
Sargassum polycystum C. Agardh is an alginate-producing seaweed. The ability of this seaweed talus to produce alginate, influenced by enzyme activity encoded by the gene and its expression influenced by the environmental factor. This study aims to determine the expression of gene encoding alginate S. polycystum isolated from different talus forms, so that can be interpreted the section of talus which shows the highest gene expression. This research used survey method, purposive sampling random sampling in the waters of Manganti Beach. To identify GDP-Man dehydrogenase gene candidates, in the order of the Ectocarpus siliculosus genome and amplified by PCR. Observation of gene expression based on the 2-ΔCt curve so that the highest relative gene expression in alginate encoding can be determined. The results showed that the relative expression of mannose dehydrogenase GD mRNA in S. polycystum ranged from 0.000125 - 0.076415. With the median value of the stalk thallus 0,02825433 (min 0.000125 - max 0.046714), in the form of leaves 0,01012533 (min 0.001245 - max 0.022561) and on the form of vesicles 0,04733100 (min 0.017217 - max 0.076415). There are significant differences in relative mRNA expression of mannose dehydrogenase GDP between the S. polycystum thallus form. The highest expression on the esicle.
PENDAHULUAN
Sargassum memiliki potensi yang cukup tinggi karena pertumbuhannya yang cepat,
kemampuan menyesuaikan diri terhadap perubahan musim dan memiliki prospek ekonomi yang
baik (Husni et al., 2012; Widyartini et al., 2012). Alga Sargassum yang dikenal karena potensi
ekonominya adalah Sargassum polycystum (Masduqi et al., 2014). S. polycstum memiliki bentuk
tubuh berupa talus (Anggadiredja, 2008). Bentuk talus ada yang menyerupai daun, akar, batang,
bahkan ada yang menyerupai buah (Purwanti, 2013). Menurut Widyartini et al. (2012), bentuk talus
dapat mempengaruhi kandungan alginat (Widyartini et al., 2012).
Alginat banyak digunakan sebagai bahan baku industri, antara lain industri makanan 30%,
farmasi 5%, industri tekstil sekitar 50%, welding rods 5%, industri kertas 6%, dan industri lainnya
4% (Mc. Hugh, 2008). Alginat berfungsi sebagai pembentuk gel (gelling agent), penstabil
(stabilizer), pengemulsi (emulsifier), pensuspensi (suspending agent), dan pendispersi suatu produk
(Kadi, 2005). Biosintesa alginat melibatkan beberapa enzim diantaranya adalah heksokinase,
phosphomannose isomerase, dan D-mannose-1-phosphate guanylyl transferase yang mengkatalis
gula nukleotida GDP-D-mannose. Enzim guanosine diphosphate mannose dehidrogenase (GDP
mannose dehydrogenase) memiliki aktivitas dalam mengoksidasi asam GDP-D-mannuronic yang
pada akhirnya mengikat asam mannuronat dalam pembentukan suatu alginat (Rasyid, 2003).
Prekusor pembentukan alginat adalah asam GDP-D-mannuronic yang diyakini berasal dari derivat
oksidasi empat elektron gula GDP-D-mannose. Enzim GDP mannose dehydrogenase (GMD)
mengkatalisasi reaksi perubahan dari GDP-D-mannose + 2NAD+ menjadi Asam
GDP-D-mannuronic + 2 NADH (Tenhaken et al., 2011).
Penelitian mengenai ekspresi mRNA GDP mannose dehydrogenase yang merupakan enzim
kunci pembentukan alginat (Tenhaken et al., 2011) bermanfaat dalam memberikan informasi
ilmiah mengenai ekspresi gen pembentuk alginat sehingga dapat mendukung pemanfaatan
sumberdaya alginat secara optimal. Oleh karena itu, perlu adanya penelitian untuk mengetahui
profil ekspresi mRNA GDP mannose dehydrogenase pada S. polycystum. Tahapan sintesa protein
fungsional atau enzim mengikuti dogma sentral dimana informasi genetik dari DNA ditranskripsi
menjadi RNA dan melalui tahapan translasi RNA diubah menjadi protein (Rochmah et al., 2009).
Ekspresi suatu gen secara molekuler dapat dideteksi pada tahap transkripsi (mRNA) (Pardal,
2010). Deteksi ekspresi gen pada tingkat mRNA memerlukan tahapan isolasi mRNA pada fase atau
bagian yang mengekspresikan gen tersebut dan membutuhkan alat yang sensitif. RT-qPCR
(Reverse Transcriptase Quantitative Polymerase Chain Reaction) merupakan teknik Real Time
PCR yang paling sensitif untuk mendeteksi dan mengukur kuantitas mRNA (Pardal, 2010). Tujuan
1. Mengetahui ekspresi relatif mRNA GDP mannose dehydrogenase pada masing-masing bentuk
talus S. polycystum asal pesisir pantai Manganti Kebumen.
2. Menentukan bagian talus S. polycystum yang mempu mengekspresi relatif mRNA GDP
mannose dehydrogenase yang paling tinggi.
METODE PENELITIAN
A.Materi Penelitian
Bahan yang digunakan dalam penelitian ini adalah Sargassum polycystum, zeolith, alkohol
70%, β-merkaptoetanol, etanol 96%, kit ekstraksi RNA (plant RNA lysis solution, , wash buffer 1,
wash buffer 2, dan air bebas nuklease), kit sintesis cDNA (oligo (dT)18 primer, 5x reaction buffer,
inhibitor ribolock RNase, mix dNTP (dATP, dCTP, dGTP, dTTP), dan RevertAID M-MULV RT),
kit qPCR SYBR green (2x master mixSYBRGreen qPCR (Tbr DNA polymerase, SYBR® Green I,
PCR buffer, MgCl2, dan dNTP mix termasuk dUTP)), ddH2O, dH2O, primer forward untuk gen
target (GMD-F), primer reverse untuk gen target (GMD-R), primer forward untuk HKG
(GSPAc-F), dan primer reverse untuk HKG (SPAct-R).
Alat yang digunakan dalam penelitian ini adalah tip putih, tip kuning, tip biru, tube qPCR,
alumunium foil, timbangan digital, autoklaf, masker, sarung tangan, mortar dan pestle, spatula,
microtubes 1,5 ml RNase free dan DNase free, mikropipet, waterbath, collection tube,
RNase-freecollection tube 1,5 ml, purification column, timer, kulkas -20°C, kulkas -80°C, vortek, kamera,
sentrifugator, nano drop spektrofotometer, mesin Real Time PCR, dan komputer.
B. Metode Penelitian
Penelitian ini menggunakan pendekatan deskriptif dengan metode survei. Pengambilan
sampel S. polycystum dilakukan di pesisir Pantai Manganti, Kabupaten Kebumen, Jawa Tengah.
Variabel yang diamati adalah variabel terikat, yaitu nilai ekspresi relatif mRNA GDP mannose
dehydrogenase pada S. polycystum. Variabel bebas, yaitu bentuk talus S. polycystum asal pesisir
pantai Manganti Kebumen. Adapun tahapan kerjanya sebagai berikut:
1. Persiapansampel
Pengambilan sampel pada penelitian ini menggunakan teknik purposive random sampling
yaitu teknik pengambilan sampel dengan memperhatikan pertimbangan-pertimbangan yang dibuat
oleh peneliti (Hadi, 2004). Sampel ditimbang dengan berat masing-masing 100 mg dengan
timbangan digital. Sampel lalu disimpan di kulkas -20°C. Penyortiran, pembersihan dan
penyimpanan sampel dilakukan di hari yang sama untuk menghindari degradasi RNA (Sim et al.,
2. Isolasi RNA Sargassum polycystum
a. Plant RNA lysis solution diambil sebanyak 500 µl lalu ditambahkan dengan β-merkaptoetanol
sebanyak 10 µl kedalam microtubes 1,5 ml RNase free dan DNase free.
b. Bentuk talus batang, bentuk talus daun dan bentuk talus vesikel dengan berat 100 mg dihaluskan
bersama pasir zeolith dengan teknik maserasi. Kemudian dipindahkan hasil maserasi kedalam
microtubes 1,5 ml RNase free dan DNase free yang didalamya sudah terdapat Plant RNA lysis
solution.
c. Lysate (b) diinkubasi selama 5 menit pada suhu 56°C. Lalu, disentrifugasi 7 menit dengan
kecepatan 12.000 rpm.
d. Supernatan diambil sebanyak 550 µl lalu dipindahkan kedalam microtubes yang baru.
e. Lysate (d) ditambahkan etanol 96% dicampur dengan pipet.
f. Lysate (e) dipindahkan ke purification tube yang sudah dimasukkan kedalam collection tube.
Disentrifugasi selama 1 menit dengan kecepatan 12.000 x g (11.000 rpm).
g. Cairan supernatan yang terdapat didalam tube dibuang. Lalu dipasangkan kembali purification
column kedalam collection tube yang baru.
h. Wash buffer 1 yang sebelumnya sudah ditambahkan etanol kemudian ditambahkan sebanyak
700 µl kedalam purification column. Lalu, disentrifugasi selama 1 menit dengan kecepatan
12.000 x g (11.000 rpm). Cairan yang terdapat didalam tube dibuang dan purification column
dipindahkan kedalam collection tube 2 ml yang baru.
i. Wash buffer 2 yang sebelumnya sudah ditambahkan etanol kemudian ditambahkan sebanyak
500 µl kedalam purification coloumn. Lalu, disentrifugasi selama 1 menit dengan kecepatan
12.000 x g (11.000 rpm). Cairan yang terdapat didalam tube dibuang dan purification column
dipindahkan kedalam collection tube yang baru.
j. Perlakuan (i) diulang kembali sebanyak 1 kali. Kemudian, disentrifugasi kembali selama 1
menit dengan kecepatan 12.000 rpm. Cairan yang terdapat didalam tube dibuang dan
purification column dipindahkan kedalam RNase free 1,5 ml collection tube.
k. Air bebas nuklease (water free nuclease) ditambahkan sebanyak 50 µl tepat ditengah-tengah
membran purification column untuk mengelusi RNA. Kemudian, disentrifugasi selama 1 menit
dengan kecepatan 12.000 x g (11.000 rpm).
l. Purification column dibuang. Kemudian, RNA yang terdapat didalam RNase free 1,5 ml
collection tube disimpan dikulkas pada suhu -80°C untuk penyimpanan dalam jangka waktu
lebih dari satu bulan.
3. Sintesis cDNA
a. Template RNA berupa RNA total diambil sebanyak 5 µl dan dimasukkan kedalam microtubes
b. Oligo (dT18) primer ditambahkan sebanyak 1 µl.
c. Air bebas nuklease ditambahkan hingga volume mencapai 12 µl.
d. Lysate (c) disentrifugasi 1 menit dengan kecepatan 1.500 rpm. Lalu, diinkubasi pada suhu 65 °C
selama 5 menit. Dinginkan diatas gel ice lalu divortex selama 1 menit.
e. 5x reaction buffer ditambahkan sebanyak 4 µl.
f. Ribolock Rnase inhibitor (20 U/ µl) ditambahkan sebanyak 1 µl.
g. 10 mM dNTP mix ditambahkan sebanyak 2 µl.
h. RevertAid M-MuLV RT (200 U/ µl) ditambahkan sebanyak 1 µl hingga didapatkan volume total
20 µl.
i. Lysate (h) disentrifugasi selama 2 menit dengan kecepatan 2500 rpm. Kemudian diinkubasi
pada suhu 42 °C selama 60 menit.
j. Suhu dinaikkan menjadi 70 °C untuk memaksimalkan reaksi terminasi akhir selama 5 menit.
k. cDNA disimpan didalam kulkas dengan suhu -80 °C untuk penyimpanan dalam jangka waktu
yang lebih lama.
4. Pengukuran Konsentrasi cDNA dengan Nano Drop Spektrofotometri
a. Mesin nano drop spektrofotometri diatur.
b. Larutan ddH2O digunakan sebagai blanko.
c. cDNA dari masing-masing sampel diambil sebanyak 2 µl dan dimasukan kedalam mesin nano
drop.
d. Hasil pengukuran konsentrasi cDNA akan muncul pada layar mesin kemudian dicatat.
5. Analisis ekspresi mRNA dengan metode RT-qPCR
a. Metode RT-qPCR digunakan untuk mendeteksi ekspresi mRNA GDP mannose dehydrogenase
(GMD) pada sampel S. polycystum asal pesisir Pantai Karimunjawa. Primer sisi kiri
(left/forward primer) (GMD-F) 5’-ATGGATCCATGCCAGGAAAGGAGAACG-3’ dan primer
sisi kanan (right/reverse primer) (GMD-R) 5’-TGAAGCTTCCAGCGAGCTCAACGTGT-3’
didesain dari gen GMD Ectocarpus siliculosus (Tenhaken et al., 2011). House Keeping Gene
(HKG) atau gen pembaku digunakan untuk normalisasi nilai Ct (Cycle treshold). HKG
menggunakan gen β-aktin. Primer sisi kiri (GSPAc-F) 5’-AGGCTCTGGTGGTGGACA-3’ dan primer sisi kanan (SPAct-R) 5’-TCGGTAAGAAGGACGGGGTGT-3’ di desain dari daerah
koding atau ekson pada gen aktin (Ac) yang akan menghasilkan amplikon cDNA berukuran 304
bp (Sim et al., 2012). Campuran RT-qPCR (RT-qPCR mix) dibuat untuk dua jenis gen, yaitu
GMD sebagai gen target dan β-aktin sebagai HKG. Komposisi RT-qPCR mix gen target GMD dan HKG terdiri dari 5 µl 2x master mixSYBRGreen (Tbr DNA polymerase, SYBR® Green I,
kanan, 3 µl dH2O (water PCR grade) dan 1,0 µl cetakan cDNA 10 ng/ µl. Total volume
RT-qPCR mix sebanyak 10 µl
b. Proses amplifikasi gen target dan HKG dilakukan dalam satu waktu PCR yang sama.
Amplifikasi dilakukan pada mesin Real Time PCR Bioer sebanyak 40 siklus dengan kondisi
PCR sebagai berikut : pre denaturation 95 °C selama 7 menit, denaturation 95 °C selama 30
detik, annealing dan extention 60 °C selama 30 detik, dan post extention 30 °C selama 5 menit,
dan storage 4 °C.
yang telah dinormalisasi (Livak & Schmittgen, 2001). Nilai Ct GDP mannose dehydrogenase
dinormalisasi dengan nilai Ct β-aktin (House Keeping Gene). Β-aktin dipilih sebagai gen pembaku karena ekspresinya relatif konstan dan tidak dipengaruhi oleh lingkungan (Heid et al., 1996; Bustin,
2000).
HASIL DAN PEMBAHASAN
Hasil penelitian menunjukkan bahwa nilai ekspresi relatif mRNA GDP mannose
dehydrogenase (GMD) pada talus S. polycystum asal pesisir pantai Manganti berkisar antara 0,0001
– 0,758 dan nilai Ct median GMD talus bentuk batang yaitu 33,98, talus bentuk daun 36,51 dan talus bentuk vesikel 33,63 (Tabel 1).
9. 2V3 35,12 5,87 0,0170 Vesikel Individu 3
Nilai Ct menunjukkan siklus amplifikasi dimana sinyal fluorescent melewati batas ambang
(threshold) yang ditandai dengan jumlah amplikonyang bertambah(Dewi et al., 2015;
Rahmaningtyas et al., 2013). Amplifikasi PCR untuk deteksi ekspresi mRNA GDP mannose
dehydrogenase (GMD) pada S. polycystum menggunakan metode RT-qPCR. Hasil reaksi RT-qPCR
berupa nilai cycle threshold (Ct) pada semua perlakuan talus S. polycystum (Budiman et al., 2015;
Shahib et al., 2015; Livak & Schmittgen, 2001).
Nilai Ct digunakan untuk mengkategorikan tingkatan ekspresi (Dewi et al., 2015; Shahib et
al., 2015; Budiman et al., 2015). Nilai Ct dikategorikan bervariasi dari 15 sampai 40. Nilai Ct > 30
– 35 kategori ekspresi rendah; dan nilai Ct > 35 – 40 kategori ekspresi sangat rendah (Dewi et al., 2015; Shahib et al., 2015; Budiman et al., 2015). Bentuk talus daun menunjukkan ekspresi sangat
rendah (nilai Ct > 35 – 40), bentuk talus batang dan vesikel menunjukkan ekspresi rendah (nilai Ct
> 30 – 35).
Nilai Ct menggambarkan jumlah amplikon(semakin kecil nilai Ct maka semakin banyak
jumlahamplikon) (Schmittgen &Livak, 2008). Nilai Ct sangat berkorelasi dengan kuantitas urutan
DNA target, apabila kuantitas urutan DNA target tinggi di awal reaksi maka nilai Ct akan lebih
cepat diketahui (Giglio et al., 2003). Nilai Ct yang tinggi kemungkinan karena disebabkan
sedikitnya mRNA templatepada reaksi reverse transcriptase, sehingga jumlah cDNA yang
terbentuk sedikit. Jumlah cDNA menentukan banyaknya amplikonyang dihasilkan dalam reaksi
RT-qPCR.Menurut Agrawal (2008) kemurnian cDNA target juga mampu mempengaruhi hasil
PCR, sehingga untuk mendapatkan hasil RT-qPCR yang optimal diperlukan cDNA target yang
murni.
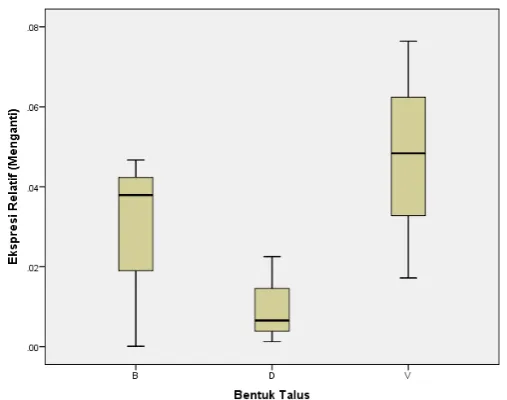
Hasil analisis box-plot menunjukkan nilai median ekspresi relatif mRNA GDP mannose
dehydrogenase pada bentuk talus S. polycystum asal pesisir pantai Manganti. Nilai median ekspresi
relatif mRNA GDP mannose dehydrogenase pada talus bentuk batang 0,0376 (min. 0,0001 – maks.
0,0463), talus bentuk daun 0,0065 (min. 0,0013 – maks. 0,0224) dan talus bentuk vesikel 0,0483
(min. 0,0170 – maks. 0,0758) (Gambar 1). Distribusi data pada ketiga kelompok bentuk talus tidak
simetris, menurut Dahlan (2014) data dikatakan tidak simetris apabila panjang whisker bagian atas
Gambar 1. Hasil Analisis Box-Plot Ekspresi Relatif mRNA GDP Mannose Dehydrogenase
Pada Bentuk Talus Sargassum polycystum.
Hasil uji anova ekspresi relatif mRNA GDP mannose dehydrogenase pada tiap bentuk talus
S. polycystum didapatkan rerata rangking bentuk talus batang, bentuk talus daun dan bentuk talus
vesikel secara berurutan yaitu 4, 5, dan 6 dengan jumlah total sampel sebanyak 9. Nilai
signifikansi p = 0,225 melebihi batas kritis 0,05 sehingga hasil tidak menunjukkan adanya
perbedaan yang signifikan (p>0,05).
Tabel 2. Hasil Uji AnovaEkspresi Relatif mRNA GDP Mannose Dehydrogenase pada Bentuk
Talus Sargassum polycystum
Ekspresi Relatif mRNA
GDP mannose dehydrogenase
Jumlah
(n)
df Nilai p
Bentuk Talus Batang 3 2 0,225
Bentuk Talus Daun 3 6 0,225
Bentuk Talus Vesikel 3 8 0,225
Talus merupakan tempat menyerap unsur hara dan terjadinya proses fotosintesis. Bentuk
talus ini pula yang kemungkinan sangat berpengaruh terhadap kadar alginat yang dihasilkan
(Rasyid, 2009; Widyartini et al., 2012). Variabilitas karakter fenotipik bentuk talus mencerminkan
variabilitas genotipe dan dipengaruhi oleh lingkungan (Tave, 1999). Faktor lingkungan dapat
mempengaruhi bentuk dan ukuran talus sehingga mempengaruhi cadangan makanan alginat yang
dihasilkan. Menurut Soelistyowati et al. (2014) fenotipe pada karakter bentuk talus menunjukkan
adanya hubungan positif antara ekspresi genotipe dengan salinitas sebagai faktor lingkungan.
Salinitas dan parameter kualitas air lainnya dapat mempengaruhi fenotipe morfometrik
melalui berbagai proses fisiologis yang dilakukan oleh S. polycystum melalui proses fotosintesis
dan penyesuaian tekanan osmotik. Penyesuaian terhadap tekanan osmotik dilakukan dalam rangka
mencapai keseimbangan dengan lingkungan yaitu dengan mengatur konsentrasi ion inorganik (K+,
Na+, Cl-) dan beragam molekul organik (floridoside, isofloridoside, digeneside) (Darley, 1982).
Menurut Soelistyowati et al. (2014), salinitas juga berkorelasi positif dengan konduktivitas.
Konduktivitas merupakan gambaran numerik dari kemampuan air untuk meneruskan aliran listrik
sehingga sering disamakan dengan daya hantar listrik yang mempengaruhi peningkatan laju
penyerapan nutrien dari lingkungan ke dalam sel alga, didukung dengan nilai turbiditas
(kekeruhan) yang rendah sehingga intensitas cahaya matahari dapat masuk ke perairan dengan baik
KESIMPULAN
1. Hasil penelitian menunjukkan bahwa ekspresi relatif mRNA GDP mannose dehydrogenase pada
S. polycystum berkisar 0,000125 - 0,076415. Nilai median bentuk talus batang 0,02825433
(min. 0,000125 – maks. 0,046714), pada bentuk talus daun 0,01012533 (min. 0,001245 – maks.
0,022561) dan pada bentuk talus vesikel 0, 04733100 (min. 0,017217 – maks. 0,076415).
2. Talus vesikel rumput laut S. polycystum menunjukkan ekspresi relatif mRNA GDP mannose
dehydrogenase tertinggi dalam menghasilkan alginat.
UCAPAN TERIMA KASIH
Ucapan terima kasih untuk Kementerian Riset Teknologi dan Pendidikan Tinggi sebagai
penyandang dana Penelitian Disertasi Doktor anggaran tahun 2017. Ucapan terima kasih juga
untuk mahasiswa-mahasiswa bimbingan atas kerjasamanya pada penelitian di Pantai Manganti
serta pada pengamatan di laboratorium.
DAFTAR PUSTAKA
Agrawal, S., 2008. Techniques in Molecular Biology. Lucknow : International Book Distributing Co.
Budhiyanti, S.A., Raharjo, S., Marseno, D.W., & Lelana, I.Y., 2012. Antioxidant Activity of Brown Algae Sargassum Species Extracts from The Coastline of Jawa Island. American Journal of Agricultural and Biological Sciences, 7(3), pp. 337-346.
Budiman, F., Zoraya, A., & Nurhalim, M., 2015. The Existence of mRNAs and miRNAs Expression for Mantaining Cell Survival Networks Associated With The Human Transparent and Cataractous Lens. Journal Ocular Biology, 3(1), pp. 8-14.
Caffal, K.H., & Mohnen, D., 2009. The Structure, Function, and Biosynthesis of Plant Cell Wall Pectic Polysaccharides. Carbohydrate Research, 344, pp. 1879-1900.
Czerpak, R., Dobrzyn, P., Krotke, A., & Kicinska, E., 2002. The Effect of Auxin and Salicylic Acid on Chlorophyl and Carotenoid Contents in Wolfia arrhiza (L) Wimm. (Lemnaceae) Growing on Media of Various Trophicities. Polish Journal of Environtmental Studies, 11 (1): 231-235.
Darley, W.M., 1982. Alga Biology: A Physiological Approach. London, UK: Blackwell Scientific Publications.
Dewi, Y.A., Ahwil, C., & Nurhalim, M., 2015. The Role of Myeolid Derived Suppressor Cells and CXCR4 Genes Expression for Nasopharyngeal Carcinoma Progression. Journal of Scientific Research and Studies, 2(8), pp. 195-201.
Giglio S, Monis PT, Saint CP., 2003. Demonstration of Preferential Binding of SYBR Green I to Specific DNA Fragments in Real-Time Multiplex PCR. Nucleic Acids Research, 31:136.
Godfrey, B.J., Mayfield, M.B., Brown, J.A., Gold, M.H., 1990. Chacterization of a Gene Encoding a Manganese Peroxidase from Phanerochaete chrysosporium. Gene 9(3), pp. 119-124. Heid, C., Stevens, K., Livak & Williams, J.K., 1996. Real Time Quantitative PCR. Journal
ofGenome Research, (6), pp. 986-994.
Hildayanti, P., 2017. Studi Komparasi Keanekaragaman Sargassum di Pantai Karimunjawa dan Manganti Jawa Tengah. Fakultas Biologi Universitas Jenderal Soedirman, Purwokerto.
Husni, A., Subaryono., Pranoto, Y., Tazwir., & Ustadi., 2012. Pengembangan Metode Ekstaksi Alginat dari Rumput Laut Sargassum sp. Sebagai Bahan Pengental. Journal Agritech, 32(1), pp. 1-8.
Husni, A., Subaryono., Pranoto, Y., Tazwir., & Ustadi., 2012. Pengembangan Metode Ekstaksi Alginat dari Rumput Laut Sargassum sp. Sebagai Bahan Pengental. Journal Agritech, 32(1), pp. 1-8.
Ho, C.L., Phang, S.M., Sinnapah, N.D., & Pang, T., 1996. Molecular Approaches in The Taxonomy of The Red and Brown Seaweeds. In : Chaudary BR and Agrawal SB (Eds.)
Cytolology, Genetics and Molecular Biology of Algae. Amsterdam: SPB Academic Publishing, pp 351-362.
Jun, S.Y., Kang, S.H., & Lee, K.H., 2008. Continous-Exchange Cell-Free Protein Synthesis Using PCR-Generated DNA and an Rnase E-deficient Extract. Journal Bioctechniques, 44, pp. 387-391.
Kadi, A., 2006. Struktur Komunitas Makro Algae di Pulau Pengelap, Dedap, abang Besar dan Abang Kecil dan Kepulauan Riau. Jurnal Ilmu Kelautan, 11(4), pp. 234-240.
Livak, K.J., & Schmittgen, T.D., 2001. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and The 2-∆∆CT Method. Elsevier Science, (25), pp. 402-408.
Masduqi, A.F., Izzati, M., & prihastanti, E., 2014. Efek Metode Pengeringan Terhadap Kandungan Bahan Kimia dalam Rumput Laut Sargassum polycystum. Buletin anatomi dan Fisiologi, 22(1), pp. 1-9.
Matanjun, P., Mohamed, S., Mustapha, N.M., Muhammad, K., & Ming, C.H., 2008. Antioxidant Activities and Phenolics Content of Eight Species of Seaweeds From North Borneo. Journal of Applied Phycology, 20, pp. 367-373.
Nababan,M.G, Munasik, I.Y., Kartawijaya, T., Prasetia, R., Ardiwijaya, R.L., Pardede, S.T., Sulisyati, R., Mulyadi, & Syaifudin, R., 2010. Status Ekosistem di Taman Nasional Karimunjawa. Wildlife Conservation Indonesia Programme, 10(1), pp. 78.
Pardal, S.J., 2010. Menguji Ekspresi Gen Menggunakan Real-Time PCR. Warta Penelitian dan Pengembangan Pertanian. 32(6). Pp. 13-14.
Purwanti, A., 2013. Optimasi Kondisi Proses Pengambilan Asam Alginat dari Alga Coklat. Jurnal Teknologi Technoscientia, 5(2), pp. 125-133.
Rahmaningtyas, V., Kurniawan, N., & Oktavianie, D.A., 2013. Isolasi danKuantifikasi Ekspresi mRNA Feline Tetherin/BST-2 dari Peripheral Blood Mononuclear Cells (PBMCs) yang Diinduksi Interferon-Alpha (IFN-α). Program Studi Pendidikan Dokter Hewan, Universitas Brawijaya. Malang.
Rasyid, A., 2003. Algae Coklat (Phaepohyta) sebagai Sumber Alginat. Jurnal Oseana, 28(1), pp. 33-38.
Riani, Y., 2016. Korelasi Kadar Besi dan Magnesium Terhadap Kandungan Klorofil dan Alginat Rumput Laut Sargassum polycystum dari Dua Pantai. Fakultas Biologi Universitas Jenderal Soedirman, Purwokerto.
Roeder, V., Collén, J., Rousval, S., Corre, E., Leblanc, C., & Boyen, C., 2005. Identification of Stress Gene Transcripts in Laminaria digitata (Phaeophyta) Protoplast Cultures by Expressed Sequence Tag Analysis. Journal Phycology, 41, pp. 1227-1235.
Shahib, M.N., Budiman, F., Feranty, Z.A., 2015. Studies on Gene Expressions at The RNA Level Associated with The Senile Lens Changes in Human Lens Cataract. Donnish Journal of Medicine and Medical Sciences, 2(3), pp. 011-018.
Sim, M.C., Ho, C.L., &Phang, S.M., 2013. A Simple and Effective Method for RNA Isolation and cDNA Library Construction from The Brown Seaweed Sargassum polycystum (Fucales, Phaeophyceae). J. App Phycol, 10, pp. 1-9.
Soelistyowati, D.T., Murni, I.A., & Wiyoto., 2014. Variasi Morfologi Rumput Laut Gracillaria
spp. Yang Dibudidayakan pada Salinitas Berbeda di Tambak Desa Pantai Sederhana, Muara Gembong. Jurnal Akuakultur Indonesia, 13(1), pp. 94-104.
Tenhaken, R., Voglas, E., Cock, J.M., &Huber, C.G., 2011. Characterization of GDP-Mannose Dehydrogenase from the Brown Alga Ectocarpus siliculosus Providing the Precursor for the Alginate Polymer. Journal of Biological Chemistry, 286(1), pp. 16707-16715.
Wang, X., Tian, W., & Li, Y., 2008. Development of an Efficient Protocol of RNA Isolation from Recalcitrans Tree Tissues. Molecular Biotechnology 38, pp. 57-64.
Widyartini, D.S., Insan, A.I., &Sulistyani. 2015. Kandungan Alginat Sargassum polycystum pada Metode Budidaya dan Umur Tanam Berbeda. Biosfera, 32(2), pp. 119-125.