www.elsevier.nlrlocateraqua-online
Feeding rates and scope for growth of green
ž /
mussels, Perna
Õ
iridis L. and their relationship
with food availability in Kat O, Hong Kong
W.H. Wong, S.G. Cheung
)Department of Biology and Chemistry, City UniÕersity of Hong Kong, 83 Tat Chee AÕenue, Kowloon,
Hong Kong SAR, People’s Republic of China
Received 28 March 2000; received in revised form 10 July 2000; accepted 10 July 2000
Abstract
Ž . Ž .
The feeding and absorption rates ARs of the green mussel PernaÕiridis L. were studied
from December 1997 to November 1998. During each visit, characteristics of seston including
Ž . Ž .
concentrations of total particulate matter TPM , particulate organic matter POM , particulate
Ž . Ž .
inorganic matter PIM and organic fraction of seston f were also determined. Higher values of
Ž . Ž .
clearance rates CRs , ARs and absorption efficiencies AEs were obtained from November to January and lower values in March and April. CR was a negative power function of TPM and a positive linear function of f. ARs and AEs were significantly correlated with TPM and f
ŽP-0.001 , showing that food availability both seston quantity and quality was the key factor. Ž .
influencing feeding behaviour of P.Õiridis. The activity of amylase and cellulase in the digestive
Ž .
gland and crystalline style, and the scope for growth SFG were determined in February, May, July and October to represent the situation in winter, spring, summer and autumn, respectively. There was no correlation between digestive enzyme activities and seasons except for amylase activity in the digestive gland. SFG was lowest in February and highest in October, and was
correlated with f.q2001 Elsevier Science B.V. All rights reserved.
Keywords: Mussels; PernaÕiridis; Feeding; Scope for growth; Food availability
1. Introduction
The green mussel Perna Õiridis is widely distributed in tropical and sub-tropical
areas of the Indo-Pacific region. Its dramatic growth and natural abundance have
)Corresponding author. Tel.:q86-852-27887749; fax:q86-852-27887406.
Ž .
E-mail address: [email protected] S.G. Cheung .
0044-8486r01r$ - see front matterq2001 Elsevier Science B.V. All rights reserved. Ž .
W.H. Wong, S.G. CheungrAquaculture 193 2001 123–137
124
encouraged efforts to culture this species as a cheap protein source in this region which
Ž . Ž .
includes China Menzel, 1988 , Singapore Cheong and Lee, 1981 , the Philippines ŽWalter, 1982 , Thailand Chalermwat and Lutz, 1989 and India Rajagopal et al.,. Ž . Ž
. Ž
1998 . In Singapore, P. Õiridis accounted for 70.4% of aquaculture production Chou
.
and Lee, 1997 . After China, Thailand is the second largest producer of mussels in Asia with nearly 35,000 ha of coastal area that are suitable for mollusc culture, and P.Õiridis
Ž
yields the highest net profit of any bivalves cultured in the country Chalermwat and .
Lutz, 1989 . In Hong Kong, P. Õiridis is distributed widely from oceanic waters to
estuarine waters. It is a dominant intertidal and subtidal species and highest densities
Ž 2. Ž
have been recorded from Victoria Harbour 246 per m and Tolo Harbour )1000 per
2. Ž .
m Huang et al., 1985 . Although this species is not intentionally cultured in Hong Kong, their abundance on fish cages is so great that they are collected by fish farmers as
Ž .
a byproduct. Its high density and fast growth rate in Hong Kong Cheung, 1991 demonstrates the potential of P.Õiridis as an aquaculture species. Although this species
plays an important economic role in southeast Asia, information on the relationship
Ž .
between feeding physiology and food availability is limited Hawkins et al., 1998a . A recent approach to developing a sustainable aquaculture industry is to use ecosystem
Ž
modelling to determine the carrying capacity of the environments Grant et al., 1993; .
Heral, 1993; Dame, 1996; Bacher et al., 1998 . The first step in this approach is to
´
understand the inter-relationship between food availability and feeding response of the Ž
aquaculture species Stenton-Dozey and Brown, 1994; Bacon et al., 1998; MacDonald et .
al., 1998 . In this experiment, the physiological parameters of feeding in P. Õiridis
Ž . Ž .
which included clearance rate CR , absorption rate AR and absorption efficiency ŽAE , and their relationships with food availability in the experimental water column. were studied at Kat O, the site with the fastest growth recorded for P. Õiridis in Hong
Kong. The results may help in the calculation of the carrying capacity for this potential Ž culture site and in the selection of culture sites in different waters of Hong Kong Lee,
. 1986; Cheung, 1991 .
2. Materials and methods
Ž .
Green mussels, P.Õiridis, were collected from a fish farm in Kat O Fig. 1 at the
northeast of Hong Kong and cultured in nylon bags at the same site from December 1997 to November 1998. The site was visited once a month. During each visit, feeding rates including CRs, filtration rates, ingestion and organic ingestion rates, and ARs were determined. Hydrographic parameters including water temperature, salinity, dissolved oxygen level and seston characteristics were also measured monthly for 12 months.
Ž y1.
Seston characteristics which included total particulate matter TPM: mg l ,
particu-Ž y1. Ž y1.
late organic matter POM: mg l , particulate inorganic matter PIM: mg l and organic fraction of seston in the seawater were determined. This was done by collecting a water sample of 300 ml at the time interval of about 30 min from the outflow of the
Ž . Ž .
empty container control see the description of physiological measurements . Each
Ž .
Fig. 1. The map of Hong Kong showing Kat O, the experimental site.
Ž .
rinsed with distilled water, dried in an oven 1108C for 24 h , weighed, and then ashed
Ž .
in a muffle furnace 4508C for 6 h before final weighing. Thus, concentrations of the
Ž y1. Ž y1. Ž y1.
TPM mg l and PIM mg l were measured. The POM concentration mg l was estimated by subtracting PIM from TPM. The organic content of suspended matter was computed as fsPOMrTPM.
To determine feeding rates, each time 13–42 individuals of P. Õiridis of sizes
Ž .
ranging from 25 to 85 cm were placed into separate containers 250 ml into which
Ž .
natural seawater 10 cm below the sea surface was pumped from the sea via a peristaltic pump. One empty container without an animal was used as a control to determine the seston characteristics of the natural seawater. The pump provided a water flow through each chamber of about 100 ml miny1 which, as determined in a
preliminary study with an electronic coulter counter, allowed the mussels by their filtering activity to deplete the particle concentration by less than 40%. The animals were maintained in the circulated natural seawater for about 1 h prior to experimentation so as to evacuate the gut. After that, all the pseudofaeces and faeces produced in the first hour were removed. The animals were then maintained in the containers for about 2–2.5 h, and faeces and pseudofaeces produced were collected. The faeces and pseudofaeces
Ž .
collected were filtered onto preashed GFrC filters Whatman . The total, inorganic and organic weight of faeces and pseudofaeces were determined by the same methods as
Ž
W.H. Wong, S.G. CheungrAquaculture 193 2001 123–137
126
y1. Ž y1.
mg h and rejection rate pseudofaeces produced per h, RR: mg h were calculated.
Ž . Ž .
ER included organic egestion rate OER and inorganic egestion rate IER whereas RR
Ž . Ž .
included organic rejection rate ORR and inorganic rejection rate IRR . AE was
Ž . Ž . Ž Ž
calculated by the Conover ratio method Conover, 1966 in which AEs fyer f= 1 ..
ye , where f was the organic content of suspended matter and e was the percentage
Ž y1. Ž .
of organic matter in faeces. CRs l h were estimated as CRs IRRqIERrPIM ŽIglesias et al., 1996 . ARs. Žmg hy1. were obtained by AE=CR=POM. When
pseudofaeces was produced, AE and AR were obtained by biodeposition method ŽIglesias et al., 1998 . These calculations assume no significant delay in the deposition. of filtered particles, estimating particle availability as the integrated average over
Ž .
corresponding periods of faecal collection Hawkins et al., 1998b . The determination of seston characteristics was discontinuous, however, the feeding rates were continuous. Therefore, the data of food availability obtained in this experiment were actually a pool of estimated mean values. The CR of P. Õiridis showed significant relationship with
Ž .
tissue dry weight Table 1 . CR was then calculated for a standardized animal of 1 g to allow comparisons among different sized groups. CRs in the following text were standardized data without special notice. The AR and AE were found to be independent of body size in the present study.
Ž y1.
Scope for growth SFG: J h is defined as the amount of energy available for production and it is the energy absorbed minus energy expended on respiration. SFG was determined in February, May, July and October 1998 to represent the situation in
Ž y1.
winter, spring, summer and autumn, respectively. Respiration rates mg O2 h of 40–42 individuals with shell lengths of between 25 and 85 cm were determined using a
Ž .
YSI 51 oxygen meter. Each mussel was put into a plastic chamber 220 ml filled with seawater and the chambers were then sealed off for 30–60 min. The time was used such that the oxygen tension in the chambers fell neither fast to make the individuals cease aerobic pumping nor slow to influence the rate of aerobic respiration. Initial and final oxygen content of the seawater in each chamber was measured. Three chambers without
Table 1
Ž y1. Ž .
animals were served as controls. The respiration rate of each individual was calculated
y1 Ž .
as mg O2 h . Energy absorbed was calculated from absorbed rate AR using a
y1 Ž .
conversion factor of 20.78 J mg POM Crisp, 1971 and energy expended on
y1 Ž .
respiration was expressed using a conversion factor of 13.98 J mg O2 Ivlev, 1934 . SFG of each individual was obtained by subtracting energy expended on respiration from the correspondent AR.
Enzymatic activities in digestive glands and crystalline styles of the same group of experimental animals used in the respiration study were also determined. The amylase
Ž
and cellulase activity was determined by the Nelson–Somogyi Method Nelson, 1944; .
Somogyi, 1952 . Digestive glands and crystalline styles were collected in situ at the experimental site, put into liquid nitrogen, transported back to the laboratory and stored at y308C for in vitro enzymatic assays. Wet weights of the digestive glands and
crystalline styles were recorded just before the enzymatic assays. One digestive gland or Ž
two crystalline styles were homogenised in cold 20 mM phosphate buffer pH 6.9 and .
pH 6.5 for the digestive gland and crystalline style, respectively containing 20 mM NaCl, then centrifuged for 15 min at 4000=g. The clear supernatants were used to
determine a-amylase and cellulase activities. Standard calibration curves were set up
with glucose. The substrates for the determination of a-amylase and cellulase were
Ž . Ž .
starch 1% and carboxymethylcellulose 1% , respectively, and were made up with the corresponding phosphate buffer for digestive glands and crystalline styles, respectively. Mass of gland and crystalline style were estimated as protein using the method of Lowry
Ž .
et al. 1951 and the standard calibration curve was set up using bovine serum albumin ŽBSA . The enzyme activity was expressed in mg glucose h. y1 mgy1 protein.
Ž .
Statistical methods used include one-way analysis of variance ANOVA , multiple comparison of Student–Newman–Keuls Test, and multiple stepwise regression analyses
Ž .
of simple linear and non-linear procedures Zar, 1984 . Any regression analysis per-formed in this paper depended on the most appropriate function to be fitted in each case, following standard least square procedures.
3. Results
3.1. Hydrological conditions and seasonal seston characteristics in Kat O
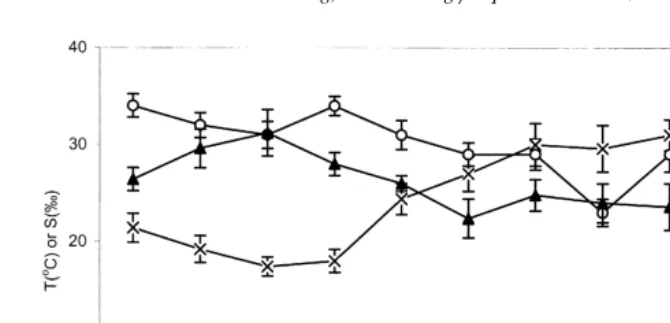
Fig. 2 shows the annual hydrological data in Kat O from December 1997 to November 1998. Temperature varied from about 188C in February to about 308C in
August. Dissolved oxygen level was higher in winter when temperature was lower ŽNovember to April and lower in summer when temperature was higher May to. Ž
.
October 1998 . Salinity remained relatively constant at about 30‰ except in July Ž25‰ . Fig. 3 shows monthly variations of seston at the study site. All seston.
Ž .
parameters, i.e., TPM, POM, PIM and f underwent temporal variations P-0.001 . Two peaks were identified for TPM, POM and PIM, the first and the major one
Ž . Ž .
W.H. Wong, S.G. CheungrAquaculture 193 2001 123–137
128
Ž .
Fig. 2. Monthly variations in temperature, salinity and dissolved oxygen mean"SE, ns9 in Kat O.
whereas f varied from 0.30 in April to 0.62 in January. Relationships between PIM, POM, TPM and f are shown in Fig. 4. There were positive linear relationships between POM, PIM and TPM that is best described by the following equations
POMs0.282=TPMq0.716
Ž
ns338, R2s0.624, P-0.001.
PIMs0.718=TPMy0.716
Ž
ns338, R2s0.915, P-0.001 ..
The contributions of POM and PIM for TPM were 28.2% and 71.8%, respectively. The relationship between f and TPM was best described by a negative power function
fs0.56=TPMy0 .21 n
s338, R2
s0.13, P-0.001 .
Ž
.
3.2. Annual feeding rates, SFG and amylase actiÕity of mussels
Monthly variations in CR, AR and AE are shown in Table 2. Higher values of CR
Ž . Ž .
were obtained from summer September to early winter January and lower values
Ž . Ž .
from winter February to early spring April . Higher values of AE were also obtained in winter, however, lower values were obtained in summer. For AR, higher values were recorded from October to January. In the present study, no pseudofaeces was produced except in January when seven individuals out of 42 produced pseudofaeces. RR of these individuals varied from 0.1 to 0.4 mg hy1.
Both respiration rate and SFG varied significantly with time, with highest values being obtained in October and lowest values in February. There was no significant difference in the amylase and cellulase activity in the crystalline style among the four
Ž .
Fig. 3. Temporal variations in the concentration of TPM, POM, PIM, and organic content of suspended solid
Ž . Žf mean"SE in Kat O..
digestive gland, however, were obtained from May to October. There was no significant
Ž .
difference in the activity of amylase P)0.05 between digestive gland and crystalline style, cellulase activity in digestive gland, however, was significantly higher than that in
Ž .
crystalline style P-0.001 .
3.3. Relationships between feeding rates, SFG, amylase actiÕity and food aÕailability
Ž . Ž .
Food availability, both in terms of quantity TPM and quality f , was shown to be a
Ž .
significant factor determining feeding and ARs Table 4 . CR was a negative power
Ž .
W.H. Wong, S.G. CheungrAquaculture 193 2001 123–137
130 Table 2
Ž .
Temporal variations mean"SE in CR, AE, AR, respiration rate and SFG in P.Õiridis
y1 y1 y1 y1
Note: within columns, treatment means followed by different letters are significantly different P-0.05 . Table 3
Ž
Amylase activity in digestive gland and crystalline style Mean"SE, ns36 for digestive glands and ns18
.
for crystalline styles
Items Enzyme activity in digestive gland Enzyme activity in crystalline style
y1 y1 y1 y1
Žmg glucose h mg protein. Žmg glucose h mg protein.
Amylase activity Cellulase activity Amylase activity Cellulase activity
a a
Note: within columns, treatment means followed by different letters are significantly different P-0.05 . Table 4
Relationships between food availability and enzymatic activity
2
Ž .
Items n Food availability TPMrf R P
y0 .724
CR 468 s2.402=TPM 0.364 -0.001
Ž .
AE 468 sy0.168=Ln TPMq0.591 0.163 -0.001
2
AR 468 sy0.006=TPM q0.21=TPMq0.738 0.061 -0.001
SFG 158 sy0.301=TPMq3.736 0.006 )0.05
a
Amylase in digestive gland 4 s0.646=TPMy0.269 0.951 -0.01
a
Cellulase in digestive gland 4 s0.245=TPMq0.896 0.238 )0.05
1.006
SFG 158 s38.724=fy15.201 0.191 -0.001
a
Amylase in digestive gland 4 s10.731=fy0.816 0.158 )0.05
a
Cellulase in digestive gland 4 sy10.14=fq7.008 0.249 )0.05
a
function of TPM and a positive linear function of f. AEs and ARs showed logarithmic relationships with TPM and varied as a polynomial function of f. SFG increased as a linear function of f but varied independently with TPM. In contrast to SFG, the amylase activity in the digestive gland showed significant relationship with TPM, but not f. Cellulase activity in the digestive gland was not significantly correlated with either TPM or f.
4. Discussions
Kat O maintains high salinity year round as a result of the strong influence of ocean Ž
currents which include the Hainan, Kuroshio and Taiwan Currents Morton and Morton, .
1983 . Occasional heavy rainfalls may contribute to lower salinities obtained in summer. Seawater temperatures demonstrated a seasonal pattern typical in subtropical areas with high values obtained in summer and lower values in winter. Two reasons were responsible for the major peak of TPM recorded in March. Firstly this was due to the growth of microalgae in spring which resulted in an increase in POM. Secondly, the
Ž
resuspension of inorganic sediment resulted in an increase in PIM Morton and Wu, .
1975 . The f value was a negative power function of TPM because the increase in TPM was largely a result of resuspension of sediments. Similar relationships were also found
Ž .
in the Merbok mangrove system, Malaysia Hawkins et al., 1998a , in Bowling Green
Ž .
Bay, Australia Yukihira et al., 1999 and in the lagoon of Takapoto Atoll, French
Ž .
Polynesia Pouvreau et al., 2000 .
Higher CR and AR values were obtained from May 1998 to January 1999. The SFG and amylase activities in the digestive gland were also higher from May to October. These agreed with the results obtained for growth and reproduction of this species that growth occurred mainly from June to November, and gonad dry weight reached a
Ž .
maximum in November Cheung, 1991 . Correlations among CR, AE, AR, respiration rates, SFG and enzyme activity in the digestive gland also showed that faster growth rates were achieved by having higher CR, AR, amylase activity and respiration rate ŽTable 5 . Feeding rate and SFG were lowered from late winter to spring February to. Ž
. Ž .
April when temperatures were low. This agreed with the results reported by Lee 1986 who showed a positive correlation between growth and temperature in day-degrees for
P.Õiridis in Hong Kong with the lowest threshold for growth obtained at 178C. Bayne
Ž .
and Newell 1983 suggested that to attain high growth and reproductive rate, animals may show strong temperature-dependent rates of oxygen consumption and feeding because of the adoption of an AexploitativeB strategy of high energy turnover where ingestion is maximized despite the increased faecal and metabolic losses incurred.
Food availability has been regarded as one of the most important factors influencing Ž
the growth of bivalves Danovaro and Fabiano, 1997; Madon et al., 1998; Sara et al.,
`
.
1998 . Both the total seston available and the proportion of food in the seston were
Ž .
expected to affect seasonal growth patterns in Mytilus edulis Bayne and Worrall, 1980 . Similar to the above studies, the present study showed that feeding and absorption in P.
Ž .
()
W.H.
Wong,
S.G.
Cheung
r
Aquaculture
193
2001
123
–
137
Table 5
Correlation coefficients between CR, AE, AR, respiration rate, SFG and digestive gland enzyme activity
Items CR AE AR Respiration SFG Amylase Cellulase
rate activity in activity in
digestive digestive
gland gland
CR 1
AE y0.719 1
AR 0.825 y0.282 1
) )
Respiration rate 0.702 y0.124 0.981 1
) )
SFG 0.962 y0.525 0.946 0.865 1
) )
Amylase activity in 0.726 y0.417 0.898 0.899 0.821 1
digestive gland
)
Cellulase activity in 0.378 y0.897 y0.017 y0.124 0.174 0.284 1
digestive gland
)
P-0.05.
feeding behaviour under different food conditions to meet physiological requirements is Ž
common in many bivalve species. These include P. Õiridis Hawkins et al., 1998a;
. Ž .
Wong and Cheung, 1999 , M. edulis Bayne et al., 1993; Hawkins et al., 1997 ,
Ž .
Cerastoderma edule Urrutia et al., 1996; Navarro and Widdows, 1997 , Crassostrea
Ž . Ž
gigas Barille et al., 1993 , and Placopecten magellanicus MacDonald and Ward,
´
.
1994; Cranford et al., 1998 . Table 6 shows some functional relationships between bivalves and their food availability. The experiments mentioned in the table were carried
Ž
out with different diets from uniquely phytoplankton diets to mixtures of phytoplankton .
and silt, to natural diets , it is not unreasonable, therefore, that feeding responses were different at intra-specific and inter-specific levels. For example, during natural tidal variations in a mangrove system, Malaysia, TPM varied between 6 and 40 mg ly1and f
varied between 0.06 and 0.22, filtration rates of P.Õiridis were correlated as a power
Ž .
function of TPM Hawkins et al., 1998a . In contrast, the filtration rate was a polynomial function of TPM and an exponential relationship of f in a laboratory study on the same species in which TPM of the seston varied from 4.46 to 24.19 mg ly1
and f
Ž .
varied from 0.12 to 0.64 Wong and Cheung, 1999 .
Pseudofaeces production is a kind of selective behaviour in bivalves. The particle selection is an obvious advantage to filter-feeding organisms, not only in sorting organic from inorganic particles, but also in AchoosingB particles of easier digestibility. Al-though it is generally believed that pseudofaeces are produced in bivalves at high TPM
Ž .
concentrations Bayne et al., 1976 , this selective behaviour was also observed at low
Ž .
TPM Hawkins et al., 1998a; Pouvreau et al., 2000 , showing that other factors are also
Ž .
important Kiørboe et al., 1980; Urrutia et al., 1996 . By using endoscopy, Ward et al.
Table 6
Relationship between feeding rate and food availability in various species of bivalves
Dependent variable Equation Species References
y0 .84 Ž .
Clearance rate s4.2=POM M. edulis Hawkins et al. 1996
y1
Žl h .
y0 .90 Ž .
s12.2=POM P.Õiridis Hawkins et al. 1998b
y0 .304TPM=f Ž .
s1.061=e P.Õiridis Wong and Cheung 1999
y0 .904 Ž .
s103.1=TPM P. margaritifera Yukihira et al. 1999
y0 .0187TPM Ž .
s38.5=10 P. maxima Yukihira et al. 1999
y0 .42 Ž .
s26.96=PIM P. margaritifera Pouvreau et al. 2000 0.96
=POM =Dry Tissue 0.61
Weight
. Ž .
Absorption rate s22.0"0.9 M. edulis Bayne et al. 1989
y1 Žy2.55"0.3POM.
Žmg h . Ž1ye .
Ž .
s0.14q0.202=POM M. edulis Bayne et al. 1993 for 2-day exposures
s0.40q0.202=POM for 12-day exposures
2 Ž .
sy0.0125POM P.Õiridis Wong and Cheung 1999
q0.135POMq0.020
y0 .340 Ž .
Absorption efficiency s0.652TPM P. margaritifera Yukihira et al. 1999
y0 .0147TPM Ž .
W.H. Wong, S.G. CheungrAquaculture 193 2001 123–137
134
Ž1993 found that bivalves could produce pseudofaeces regardless of TPM in the water.. In the present study, almost no pseudofaeces were produced at Kat O during the experimental period except in January. In contrast to the present study, there was a linear relationship between RR and filtration rates for the same species in a field study
Ž .
in Malaysia by Hawkins et al. 1998a and in a laboratory experiment by Wong and
Ž . Ž . Ž y1.
Cheung 1999 . This may be due to higher f 0.25–0.60 and lower TPM 2–12 mg l values obtained in Kat O in this study as compared with the field study by Hawkins et
Ž . Ž .
al. 1998a and the laboratory study by Wong and Cheung 1999 . The significance of f and TPM in pseudofaeces production is confirmed by the study undertaken at an aquaculture farm at Ma Wan, Hong Kong. This site is characterized by lower f and higher TPM values as compared with Kat O, most of the individuals of P.Õiridis at this
site produced pseudofaeces as expected.
High concentrations of PIM in seston lead to dilution of organic matter in suspension. The compensation was achieved in P. Õiridis by having elevated filtration rates and
Ž .
pseudofaeces production and preferential organic ingestion Wong and Cheung, 1999 . In C. edule, RRs were expressed as a simultaneous multiple regression model based on
Ž .
FR, f, TPM and the interaction term of filtration rate=f Iglesias et al., 1996 . Although mechanisms involved in preferential ingestion are not fully understood, gill structure, nervous innervation, muscular control of the gill filaments, particle size structure, chemosensory capabilities, gut residence time, digestive enzyme suites, and
Ž .
mucous secretions may all be important Shumway et al., 1990 . This kind of plastic Ž
behaviour is a physiological adaptation at both feeding and digestive levels Iglesias et .
al., 1996 . The flexibility of physiological responses in bivalves demonstrates a rich diversity of feeding behaviour, particularly when it is measured under natural field
Ž .
conditions Bayne, 1998 .
In Hong Kong, P. Õiridis is distributed widely from the east which is oceanic in
nature to the west which is estuarine. The highest monthly growth rate of 8–10 mm was
Ž .
recorded in Kat O at the northeast of Hong Kong Lee, 1986 . This indicated that Kat O has great potential for large-scale mussel culture. To make mussel culture sustainable, however, carrying capacity of the environment should be determined by means of ecosystem modelling. The first step towards this goal is to understand the inter-relation-ship between food availability and feeding responses of the aquaculture species. The present study demonstrated the significance of the annual seston dynamics on the feeding physiology and SFG of P.Õiridis. Future works will attempt to determine the
optimal culturing density of mussels to ensure maximum yield by using modelling tools ŽGrant et al., 1993; Heral, 1993; Dame, 1996; Bacher et al., 1998 .
´
.Acknowledgements
References
Bacher, C., Duarte, P., Ferreira, J.G., Heral, M., Raillard, O., 1998. Assessment and comparison of the´
Ž . Ž .
Marennes-Oleron Bay France and Carlingford Lough Ireland carrying capacity with ecosystem models.´ Aquat. Ecol. 31, 379–394.
Ž .
Bacon, G.S., MacDonald, B.A., Ward, J.E., 1998. Physiological responses of infaunal Mya arenaria and
Ž .
epifaunal Placopecten magellanicus bivalves to variations in the concentration and quality of suspended particles I. Feeding activity and selection. J. Exp. Mar. Biol. Ecol. 219, 105–125.
Barille, L., Prou, J., Heral, M., Bourgrier, S., 1993. No influence of food quality, but ration-dependent´ ´ retention efficiencies in the Japanese oyster Crassostrea gigas. J. Exp. Mar. Biol. Ecol. 171, 91–106. Bayne, B.L., 1998. The physiology of suspension feeding by bivalve molluscs: an introduction to the
PlymouthATROPHEEBworkshop. J. Exp. Mar. Biol. Ecol. 219, 1–19.
Bayne, B.L., Newell, R.C., 1983. Physiological energetics of marine molluscs. In: Saleuddin, A.S.M., Wilbur,
Ž .
K.M. Eds. , The Mollusca 4. Academic Press, New York, pp. 407–515.
Bayne, B.L., Worrall, C.M., 1980. Growth and production of mussels Mytilus edulis from two populations. Mar. Ecol.: Prog. Ser. 3, 317–328.
Ž .
Bayne, B.L., Thompson, R.J., Widdows, J., 1976. Physiology: I. In: Bayne, B.L. Ed. , Marine Mussels: Their Ecology and Physiology. Cambridge Univ. Press, London, pp. 121–206.
Bayne, B.L., Hawkins, A.J.S., Navarro, E., Iglesias, J.I.P., 1989. Effects of seston concentration on feeding, digestion and growth in the mussel Mytilus edulis. Mar. Ecol.: Prog. Ser. 55, 47–54.
Bayne, B.L., Iglesias, J.I.P., Hawkins, A.J.S., Navarro, E., Heral, M., Deslous-Paoli, J.M.D., 1993. Feeding´ behaviour of the mussel, Mytilus edulis: responses to variations in quantity and organic content of the seston. J. Mar. Biol. Assoc. U.K. 73, 813–829.
Chalermwat, K., Lutz, R.A., 1989. Farming the green mussel in Thailand. World Aquacult. 20, 41–46. Cheong, L., Lee, H.B., 1981. Improvements to rope design for the raft culture of green mussels, PernaÕiridis
Ž .L. , in Singapore. Singapore J. Primary Ind. 9, 38–53.
Cheung, S.G., 1991. Energetics of transplanted populations of the green-lipped mussel Perna Õiridis
ŽLinnaeus. ŽBivalve: Mytilacea in Hong Kong: I. Growth condition and reproduction. Asian Mar. Biol. 8,.
117–131.
Chou, R., Lee, H.B., 1997. Commercial marine fish farming in Singapore. Aquacult. Res. 28, 767–776. Conover, R., 1966. Assimilation of organic matter by zooplankton. Limnol. Oceanogr. 11, 338–354. Cranford, P.J., Emerson, C.W., Hargrave, B.T., Milligan, T.G., 1998. In situ feeding and absorption responses
Ž .
of sea scallops Placopecten magellanicus Gmelin to storm-induced changes in the quality and composi-tion of the seston. J. Exp. Mar. Biol. Ecol. 219, 45–70.
Crisp, D.J., 1971. Energy flow measurements. Methods for the Study of Marine Benthos. In: Holme, N.A.,
Ž .
Mcintyre, A.D. Eds. , International Biological Programme 16. Blackwell Scientific Publication, Oxford, pp. 197–279.
Dame, R.F., 1996. Ecology of marine bivalves: an ecosystem approach. CRC Press, Florida, 254 pp. Danovaro, R., Fabiano, M., 1997. Seasonal changes in quality and quantity of food available for benthic
Ž .
suspension-feeders in the Golf Marconi North-western Mediterranean . Estuarine Coastal Shelf Sci. 44, 723–736.
Grant, J., Dowd, M., Thompson, K., Emerson, C., Hatcher, A., 1993. Perspectives on field studies and related
Ž .
biological models of bivalve growth and carrying capacity. In: Dame, R.F. Ed. , Bivalve Filter Feeders in Estuarine and Coastal Ecosystem Processes, NATO ASI series, Vol. G 33. Springer-Verlag, Berlin, pp. 371–421.
Hawkins, A.J.S., Smith, R.F.M., Bayne, B.L., Heral, M., 1996. Novel observations underlying fast growth of´ suspension-feeding shellfish in turbid environments: Mytilus edulis. Mar. Ecol.: Prog. Ser. 131, 179–190. Hawkins, A.J.S., Smith, R.F.M., Bougrier, S., Bayne, B.L., Heral, M., 1997. Manipulation of dietary´ conditions for maximal growth in mussels, Mytilus edulis, from the Marennes-Oleron Bay, France. Aquat.´ Living Resour. 10, 13–22.
Hawkins, A.J.S., Simth, R.F.M., Tan, S.H., Yasin, Z.B., 1998a. Suspension-feeding behaviour in tropical bivalve molluscs: PernaÕiridis, Crasssostrea belcheri, Crassostrea iradelei, Saccostrea cucculata and
Pinctada margarifera. Mar. Ecol.: Prog. Ser. 166, 173–185.
W.H. Wong, S.G. CheungrAquaculture 193 2001 123–137
136
M.B., 1998b. Some general relationships in comparing the feeding physiology of suspension-feeding bivalve molluscs. J. Exp. Mar. Biol. Ecol. 219, 87–103.
Heral, M., 1993. Why carrying capacity models are useful tools for management of bivalve molluscs culture.´
Ž .
In: Dame, R.F. Ed. , Bivalve Filter Feeders in Estuarine and Coastal Ecosystem Processes, NATO ASI series Vol. G 33. Springer-Verlag, Berlin, pp. 421–454.
Huang, Z.G., Lee, S.Y., Mak, P.M.S., 1985. The distribution and population structure of Perna Õiridis
ŽBivalvia: Mytilacea in Hong Kong waters. In: Morton, B.S., Dudgeon, D. Eds. , Proceedings of the. Ž .
Second International Workshop on the Malacofauna of Hong Kong and Southern China. Hong Kong Univ. Press, Hong Kong, pp. 465–471.
Iglesias, J.I.P., Urrutia, M.B., Navarro, E., Alvarez-Jorna, P., Larretxea, X., Bougrier, S., Heral, M., 1996.´
Ž .
Variability of feeding processes in the cockle Cerastoderma edule L. in response to changes in seston concentration and composition. J. Exp. Mar. Biol. Ecol. 197, 121–143.
Iglesias, J.I.P., Urrutia, M.B., Navarro, E., Ibarrola, I., 1998. Measuring feeding and absorption in suspension-feeding bivalves: an appraisal of the biodeposition method. J. Exp. Mar. Biol. Ecol. 219, 71–86. Ivlev, V.S., 1934. Eine Micromethode zur Bestimmung des Kaloriengehalts von Nahrstoffen. Biochem. Z. 6,¨
49–55.
Kiørboe, T., Møhlenberg, F., Nøhr, O., 1980. Feeding, particle selection and carbon absorption in Mytilus
edulis in different mixtures of algae and resuspended bottom material. Ophelia 19, 193–205.
Ž . Ž .
Lee, S.Y., 1986. Growth and reproduction of the green mussel PernaÕiridis L. Bivalvia: Mytilacea in contrasting environments in Hong Kong. Asian Mar. Biol. 3, 111–127.
Lowry, O.H., Rosebrough, N.J., Farr, A.L., Randall, R.J., 1951. Protein measurements with the Folin phenol reagent. J. Biol. Chem. 193, 265–275.
MacDonald, B.A., Ward, J.E., 1994. Variation in food quality and particle selectivity in the sea scallop
Ž .
Placopecten magellanicus Mollusca: Bivalvia . Mar. Ecol.: Prog. Ser. 108, 251–264.
Ž .
MacDonald, B.A., Bacon, G.S., Ward, J.E., 1998. Physiological responses of infaunal Mya arenaria and
Ž .
epifaunal Placopecten magellanicus bivalves to variations in the concentration and quality of suspended particles. II: Absorption efficiency and scope for growth. J. Exp. Mar. Biol. Ecol. 219, 127–141. Madon, S.P., Schneider, D.W., Stoeckel, J.A., Sparks, R.E., 1998. Effects of inorganic sediment and food
concentrations on energetic processes of the zebra mussel, Dreissena polymorpha: implications for growth in turbid rivers. Can. J. Fish. Aquat. Sci. 55, 401–413.
Menzel, W., 1988. Mussel culture in China. J. Shellfish Res. 7, 569–570.
Morton, B., Morton, J.E., 1983. The sea shore Ecology of Hong Kong. Hong Kong Univ. Press, Hong Kong, 350 pp.
Morton, B.S., Wu, S.S., 1975. The hydrology of the coastal waters of Hong Kong. Environ. Res. 10, 319–347. Navarro, E., Widdows, J., 1997. Feeding physiology of Cerastoderma edule in response to a wide range of
seston concentrations. Mar. Ecol.: Prog. Ser. 152, 175–186.
Nelson, N., 1944. A photometric adaptation of the Somogyi method for the determination of glucose. J. Biol. Chem 153, 375–380.
Pouvreau, S., Bodoy, A., Buestel, D., 2000. In situ suspension feeding behaviour of the pearl oyster, Pinctada
margaritifera: combined effects of body size and weather-related seston composition. Aquaculture 181,
91–113.
Rajagopal, S., Venugopalan, V.P., Nair, K.V.K., van der Velde, G., Jenner, H.A., den Hartog, C., 1998.
Ž .
Reproduction, growth rate and culture potential of the green mussel, Perna Õiridis L. in Edaiyur backwaters, east coast of India. Aquaculture 162, 187–202.
Sara, G., Manganaro, A., Cortese, G., Pusceddu, A., Mazzola, A., 1998. The relationship between food`
Ž .
availability and growth in Mytilus galloproÕincialis in the open sea southern Mediterranean . Aquaculture 167, 1–15.
Shumway, S.E., Newell, R.C., Crisp, D.J., Cucci, T.L., 1990. Particle selection in filter-feeding bivalve
Ž .
molluscs: a new technique on an old theme. In: Morton, B.S. Ed. , The Bivalve–Proceedings of a Memorial Symposium in Honour of Sir Charles Maurice Yonge, Edinburgh, 1986. Hong Kong Univ. Press, Hong Kong, pp. 151–165.
Somogyi, M., 1952. Notes on sugar determination. J. Biol. Chem. 195, 19–23.
Stenton-Dozey, J.M.E., Brown, A.C., 1994. Short-term changes in the energy balance of Venerupis corrugatus
Urrutia, M.B., Iglesias, J.I.P., Navarro, E., Prou, J., 1996. Feeding and absorption in Cerastoderma edule
Ž .
under environmental conditions in the bay of Marennesoleron Western France . J. Mar. Biol. Assoc. U.K. 76, 431–450.
Ž .
Walter, C., 1982. Reproduction and growth in the tropical mussel Perna Õiridis Bivalvia: Mytilidae . Kalikasan 11, 83–97.
Ward, J.E., MacDonald, B.A., Thompson, R.J., Beninger, P.G., 1993. Mechanisms of suspension feeding in bivalves: resolution of current controversies by means of endoscopy. Limnol. Oceanogr. 38, 265–272.
Ž .
Wong, W.H., Cheung, S.G., 1999. Feeding behaviour of the green mussel, PernaÕiridis L. : Responses to variation in seston quantity and quality. J. Exp. Mar. Biol. Ecol. 236, 191–207.
Yukihira, H., Klumpp, D.W., Lucas, J.S., 1999. Feeding adaptations of the pearl oysters Pinctada
margari-tifera and P. maxima to variations in natural particulates. Mar. Ecol.: Prog. Ser. 182, 161–173.