www.elsevier.com / locate / bres
Research report
Chronic fluoxetine administration to juvenile rats prevents
age-associated dendritic spine proliferation in hippocampus
*
Seth D. Norrholm, Charles C. Ouimet
Program in Neuroscience, Department of Psychology, Florida State University, 211 Biomedical Research Facility, Tallahassee, FL 32306-4340, USA Accepted 22 August 2000
Abstract
The density of dendritic spines, the postsynaptic sites of most excitatory synapses, increases during the first 2 postnatal months in rat hippocampus. Significant alterations in hippocampal levels of serotonin and norepinephrine impact synaptic development during this time period. In the present study, dendritic spine density was studied in the hippocampus (CA1) and dentate gyrus of juvenile rats acutely and chronically exposed to antidepressant drugs that act on serotonin and norepinephrine. One group of 21-day-old rats was given a single injection of a serotonin specific re-uptake inhibitor (fluoxetine or fluvoxamine), a norepinephrine-specific re-uptake inhibitor (de-sipramine), or saline and killed after 24 h. A second group of rats was injected daily, beginning on postnatal day (PN) 21, for 3 weeks. This group was further subdivided into rats that were killed 1 day or 21 days after the last injection. Golgi analysis showed that a single injection of fluvoxamine produced a significant increase in dendritic spine density in stratum radiatum of CA1 and in the dentate gyrus. Further, acute treatment with all three antidepressants increased the total length of secondary dendrites in CA1, with fluoxetine and desipramine increasing the number of secondary dendrites as well. In fluoxetine-treated animals killed on days 42 or 62 (1 or 21 days post-treatment, respectively), dendritic spine density remained at levels present in CA1 at 21 days. These results show that acute antidepressant treatment can impact dendritic length and spine density, and raise the possibility that chronic fluoxetine treatment arrests spine development into young adulthood. 2000 Elsevier Science B.V. All rights reserved.
Theme: Neurotransmitters, modulators, transporters, and receptors
Topic: Serotonin
Keywords: Antidepressant; Fluoxetine; Prozac; Hippocampus; Dendritic spine; Plasticity
1. Introduction Serotonin (5-HT) influences dendritic spines and
pat-terns of connectivity in both immature and adult brain. In Dendritic spines, small variably shaped protrusions from the developing brain, serotonin autoregulates the migration dendritic shafts, receive the majority of synaptic contacts and differentiation of serotonin-releasing neurons while [63]. Spines compartmentalize second messenger cascades facilitating the growth of target neuronal populations and contain high concentrations of several classes of innervated by these neurons [23,55,89]. In rats, removal of receptors and ion channels [47]. Functionally, spines serotonergic innervation during the first three postnatal provide part of the anatomical substrate underlying synap- weeks can delay neuronal differentiation and decrease the tic transmission, LTP, LTD, and learning and memory number of synapses in cortical and hippocampal subreg-[1,13,16,27,29,32,59,74,76,77]. They are responsive to ions [53–55,87]. In the adult brain, pharmacological, pharmacological [33,46,49,54,66,70,71,87], anatomical anatomical, and environmental perturbation of serotonin [24,73], physiological [25,70], and environmental factors levels markedly alters receptor dynamics [4,34,40,41], [1,5,13,15,26,29,30,36,38,49,59,61,64,65]. intracellular cascades [21,22,61], growth factor expression [61] dendritic morphology [51,56] and synaptic density [12,53,54]. Synaptic density is decreased by blockade of
*Corresponding author. Tel.: 11-850-644-2271; fax:
11-850-644-serotonergic transmission with parachlorophenylalanine
0989.
E-mail address: [email protected] (C.C. Ouimet). (PCPA) [53,54,62]. Conversely, chronic stress-induced
elevations in synaptic serotonin increase the density of animals were weaned from the dam and randomly assigned dendritic spines on serotonergic neurons [75]. Although to one of 15 experimental conditions. One group was given dendritic spines are responsive to alterations in serotoner- a single injection to differentiate between acute and gic transmission, it is not known whether dendritic spine chronic drug effects. This group received a single i.p. density is one of the targets of serotonin-specific re-uptake injection of fluoxetine HCl (Prozac; 5 mg / kg), fluvox-inhibitor (SSRI) action. amine maleate (Luvox; 5 mg / kg), desipramine (Nor-Serotonin-specific re-uptake inhibitors are prescribed for pramin, DMI; 10 mg / kg), or 0.9% saline on PN 21 and the treatment of clinical depression. Acutely, SSRIs in- were killed 24 h later. A second group (chronic treatment) crease the synaptic availability of serotonin, but the long- was subjected to the same dose regimen but for 3 weeks, term synaptic effects as well as the mechanism underlying beginning on PN 21. Injections were given at the same their therapeutic efficacy remain largely unknown [48,52]. time each day and in the home cage of each animal. In Chronic treatment with one SSRI, fluoxetine (Prozac), addition, a non-handled group was included in each alters a number of physiological processes in the adult condition for control purposes. This group was handled hippocampus including sustained fluctuations in synaptic only during weaning and was left undisturbed until sac-serotonin levels [8,10,34,42,48,52,67,68,78], increased rifice. Half of the animals receiving chronic treatment synaptic norepinephrine release [21,22], down-regulation (n520) were killed 24 h after the last injection (on PN 42) of b-adrenergic receptors [20,39], up-regulation of 5-HT2 and the remaining animals were killed following a 21-day receptors [21,22,41,79], supersensitivity of 5-HT2 recep- washout period (on PN 62) during which no injections tors [9], elevated intracellular cAMP levels [21,22,61], and were given. In addition, animals were handled only as increased expression of BDNF and trkB proteins required for cage maintenance during the washout period. [21,22,61].
The anatomy and plasticity of dendritic spines has been 2.2. Tissue preparation most heavily studied in the hippocampus [32], a brain
region implicated in both the pathophysiology of depres- Animals were perfused transcardially with 4% formalde-sion and therapeutic benefits of antidepressant drugs such hyde (in 0.1 M sodium phosphate buffer, pH 7.4) under as fluoxetine [21,22]. Pyramidal cells in hippocampal deep anesthesia with sodium pentobarbital (60 mg / kg) at subfield CA1 of adult rats exposed to fluoxetine for 3 PN 22, 42 or 62. Following fixation, brains were removed weeks displayed longer hyperpolarizations and time con- and placed in 4% formaldehyde (freshly depolymerized stants, greater potency of 5-HT for 5-HT1A receptors, and from paraformaldehyde) overnight. Brains were then sec-decreased potency of baclofen for GABAB receptors [3]. tioned at 100mm on a vibratome and prepared for Golgi In addition, the number of 5-HT uptake sites in CA1 impregnation using a protocol adapted from Izzo et al. increases by 111% after chronic fluoxetine treatment in [45]. In short, tissue sections were post-fixed in 1% OsO4
adult rats [41]. for 30 min and then washed in 0.1 M phosphate buffer The purpose of the present study was to determine (3315 min). Sections were then free-floated in 3.5% whether SSRIs, administered to juvenile rats after the K Cr O2 2 7 solution for 90 min, mounted between two serotonergic system has developed, produce enduring microscope slides in a ‘sandwich’ assembly, and rapidly changes in CA1 dendritic spine density. The effects of two immersed in a 1% AgNO3 solution. The following day, SSRIs, fluoxetine and fluvoxamine, were compared with sections were rinsed in ddH O, dehydrated in 70% and2
those of desipramine (DMI), a norepinephrine-specific 100% ethanol, cleared with Histoclear, and mounted on uptake inhibitor. We report here that acute treatment with microscope slides with DPX.
fluvoxamine caused an increase in dendritic spine density
that could be observed 24 h later. Further, chronic fluox- 2.3. Analysis of dendritic spine density etine treatment prevented the age-dependent increase in
dendritic spine density that was observed in controls. Dendritic spines were counted on 12503camera lucida images that included all spines observable in each focal plane occupied by the dendrite. Cells were analyzed only if
2. Materials and methods they were fully impregnated (CA1: primary apical
Approximately 20 dendritic segments (10 in each cerebral hemisphere; 50–100 mm in length) within CA1 and 20 within dentate gyrus were examined for each experimental animal. Treatment conditions were coded throughout the entire process of cell identification, drawing, counting, and subsequent data analysis. Two-tailed Student’s t-tests were used to assess differences between experimental groups.
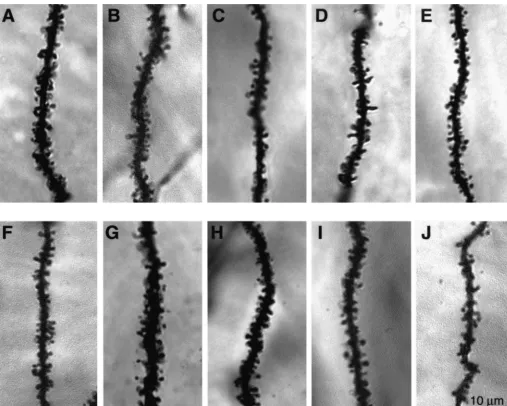
When significant changes in dendritic spine density were observed, camera lucida images and the Zeiss CLSM measurement program were used to quantify the number and length of secondary dendrites; this analysis was necessary since apparent changes in dendritic spine density can result from an increase or decrease in the length of dendrites and not the formation or loss of spines per se. Photomicrographs were obtained with a helium–neon 633 laser and a Zeiss 410 confocal laser scanning microscope.
3. Results
3.1. Acute treatment
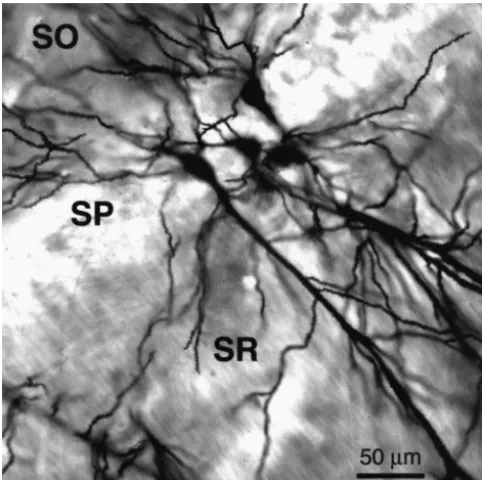
Fig. 1. Laser confocal photomicrograph illustrating Golgi-impregnated
pyramidal neurons within hippocampal area CA1. Secondary oblique In a 24-h period, single fluvoxamine injection increased
dendrites within stratum radiatum (SR) were analyzed in the present dendritic spine density along secondary dendrites by study. SO, stratum oriens; SP, stratum pyramidale; SR, stratum radiatum.
18.4% in stratum radiatum of CA1 (t523.79; P50.013) and 11.6% in dentate gyrus (t522.72; P50.042) com-CA1, secondary dendrites were defined as those branches pared to non-handled controls (Table 1A,B). There was projecting directly from the primary apical dendrite exclu- also a significant increase in spine density in the same two sive of tertiary daughter branches. In addition, spines were subregions when compared to saline-injected animals; counted along the length of secondary dendrites of granule dendritic spine density was 17.4% greater in CA1 (t5
cells in the dentate gyrus to determine if effects were 26.39; P50.001) and 16% greater in dentate gyrus (t5
limited to CA1. Most of the secondary dendrites in dentate 23.02; P50.023) (Table 1A,B). There was no significant gyrus were in the outer half of the molecular layer. difference in spine density between saline-injected and
Table 1
Dendritic spine densities (number / 10mm segment6S.E.M.) in the area CA1 (A) and (B) dentate gyrus following acute and chronic antidepressant treatment*
Treatment Single injection Daily injection for Daily injection for 21 days
(PN 22) 21 days (PN 42) followed by 21-day washout (PN 62)
A. CA1
a a
Unhandled 14.8660.04 (n54) 20.2660.81 (n54) 20.1560.54 (n54)
b b
Saline 15.5360.22 (n54) 18.2060.73 (n54) 19.2360.84 (n54)
c d
Fluoxetine 14.4360.09 (n54) 15.0960.32 (n54) 15.3860.39 (n54)
f e
Fluvoxamine 17.6060.34 (n54) 17.5360.18 (n54) 18.2560.44 (n54)
e
DMI 14.3660.36 (n54) 17.2760.28 (n56) 17.5960.51 (n56)
B. Dentate gyrus
Unhandled 14.4960.88 (n54) 15.2760.51 (n54) 15.6860.46 (n54)
Saline 13.9460.61 (n54) 17.0661.04 (n53) 14.8560.86 (n54)
Fluoxetine 14.2660.22 (n54) 15.6660.91 (n54) 15.8560.13 (n53)
f
Fluvoxamine 16.3360.34 (n54) 14.8261.08 (n54) 15.9160.39 (n54)
DMI 15.2360.57 (n54) 15.4960.39 (n56) 15.8660.30 (n56)
*
The number of animals per condition is indicated by n. For each animal, spine density on approximately 20 secondary dendritic segments of at least 50 a
mm in length was quantified. Superscript letters in table indicate significance: P,0.05 when compared to mean spine density of unhandled animals at PN
b c
22; P,0.05 when compared to mean spine density of saline-injected animals at PN 22; P,0.05 when compared to both unhandled and saline-injected
d e
controls at PN 42; P,0.05 when compared to both unhandled and saline-injected controls at PN 62; P,0.05 when compared to unhandled controls at f
non-injected animals in CA1 or the dentate gyrus in either variance in the data and differences between and among the acute or chronic conditions (Fig. 2 and Table 1A,B). treatment groups were not due to outlying data points. This Fluvoxamine-injected animals showed significantly more was true for all of the spine density data in this study. spines per unit length in CA1 compared to control animals The number and length of secondary dendrites was (Fig. 2F,G); this fluvoxamine-induced increase in spine analyzed to determine whether these factors affected the density occurred in the absence of change in length of apparent increase in spine density following acute antide-individual dendrites (Table 2A). Acute fluoxetine, DMI, or pressant treatment. Acute treatment with each drug did not saline treatment produced no significant changes in den- alter the average length of individual secondary dendrites dritic spine density in either hippocampal subregion when in CA1 or dentate gyrus (Table 2A,B). In CA1, however, compared to non-handled controls (Table 1A,B). The small the total number of secondary dendrites was increased S.E.M. values shown in the tables reflect the small following acute fluoxetine (t524.05; P50.0008) and DMI
Table 2
Summary of the number of secondary dendritic branches, average length per dendrite, and summed dendritic length (6S.E.M.) in CA1 (A) and the dentate a
gyrus (B) following acute antidepressant treatment
Acute treatment (PN 22) Total no. of Secondary dendrite Summed dendritic
secondary dendrites average length (mm) length (mm)
A. CA1
Control (n58) 16.6060.65 87.6663.13 1448664
a b
Fluoxetine (n54) 20.9060.84 86.3363.09 1722687
b
Fluvoxamine (n54) 18.7161.23 89.1664.32 16426208
a b
DMI (n54) 21.6261.19 81.4163.33 1676676
B. Dentate gyrus
Control (n58) 8.560.37 101.63612.22 931633
Fluoxetine (n54) 9.060.38 110.8564.84 987626
Fluvoxamine (n54) 8.460.60 104.0163.43 870658
DMI (n54) 9.460.87 102.9365.51 9646103
a
Summed dendritic length refers to total dendritic length of all secondary dendrites located in stratum radiatum on the neurons analyzed for spine density. The number of animals per condition is indicated by n. Ten cells per treatment condition were quantified. Data from unhandled and saline-injected animals were pooled because they did not differ from one another in terms of number of secondary dendrites, length of secondary dendrites, sum dendritic length,
a
or dendritic spine density. Superscripts in table indicate significance: P,0.05 when total number of secondary dendrites is compared to control values at b
PN 22; P,0.05 when summed dendritic length is compared to control values at PN 22.
(t523.69; P50.004) (Table 2A). There was also an vidual neurons in dentate gyrus was not affected by any increase in the total number of secondary dendrites in CA1 acute treatment (Table 2B).
following acute fluvoxamine, but this change was not
statistically significant (Table 2A). Consistent with the 3.2. Chronic treatment increased number of secondary dendrites, the sum of the
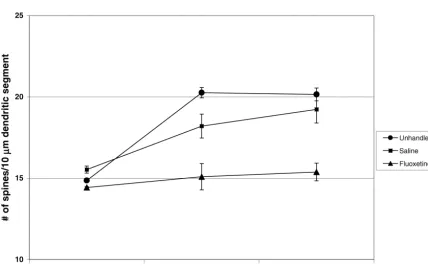
secondary dendritic lengths for individual neurons in CA1 From PN 22 to 42, unhandled controls showed a 27% was increased by fluoxetine (16%; t52.55, P50.02), increase (t55.68; P50.001) in dendritic spine density and fluvoxamine (12%; t523.11, P50.02), and DMI (11%; saline-injected controls showed a 15% increase (t524.73;
t52.29, P50.004) (Table 2A). The total number of P50.009) (Fig. 3 and Table 1A) in CA1. Dendritic spine secondary dendrites and sum dendritic length for indi- density in CA1 of saline-injected animals was not
nificantly different from that in non-injected animals at PN and Table 1A). When compared to unhandled controls, 22 or 42. (Fig. 2A,B and Table 1A). Animals exposed to animals exposed to a 3-week DMI treatment followed by a fluoxetine for 3 weeks and killed 24 h after the last 3-week washout period had 12.7% fewer spines per injection, however, had 17.1% fewer dendritic spines per dendritic length (t50.71; P50.005) in CA1 (Table 1A). In dendritic length in CA1 when compared to saline-injected addition, animals treated for 21 days with fluvoxamine and rats (t54.48; P50.011) and 25.5% fewer compared to left undisturbed for 3 weeks had 9.4% fewer spines per non-injected animals (t56.86; P50.002) (Fig. 2A–C and dendritic length (t52.96; P50.025) compared to unhand-Table 1A). At PN 42, dendritic spine density in fluoxetine- led control animals (Table 1A). Total number of secondary treated animals was similar to that found at PN 22 (Fig. 3 dendrites, average length per secondary dendrite, and and Table 1A). In CA1, spine density was 13.8% (t53.87; summed dendritic length per neuron were examined to
P50.031) lower following chronic fluvoxamine treatment determine whether the observed changes in spine density and 14.8% (t53.95; P50.017) lower following chronic reflected changes in these dendritic factors. None of the DMI treatment when compared to unhandled rats (Figs. chronic antidepressant treatments or washout periods af-2A,D,E and Table 1A). Total number of secondary den- fected dendritic length in stratum radiatum of CA1 (Table drites, average length per secondary dendrite, and summed 3B). In dentate gyrus, there were no significant differences dendritic length per neuron were examined to determine in secondary dendritic length, number or spine density whether the observed changes in spine density reflected among animals administered fluoxetine, fluvoxamine, changes in these dendritic factors. None of the chronic DMI, or saline for 21 days followed by a washout period antidepressant treatments affected dendritic length in (Table 1B).
stratum radiatum of CA1 (Table 3A).
In the dentate gyrus, spine density did not differ between 3.4. Body weight unhandled and saline-injected animals at PN 22 or 42
(Table 1B). In addition, chronic antidepressant treatment Although SSRIs produce anorectic effects in rats [7], did not affect dentate gyrus spine density (Table 1B). there was no significant difference in body weight among the experimental groups examined in this study (data not 3.3. Chronic treatment followed by a 21-day washout shown).
period
In CA1, unhandled and saline-injected animals show no 4. Discussion
further increase in spine density from PN 42 to 62 (Fig. 3
and Table 1A). Rats treated for 3 weeks with fluoxetine The main findings of the present study are that dendritic and left undisturbed for 3 weeks had 20% fewer spines per spine density in CA1 and the dentate gyrus is increased unit dendritic length (t54.81; P50.009) in CA1 compared following acute fluvoxamine treatment, and spine density to saline-injected controls, and 23.6% fewer (t58.27; P5 in CA1, but not in dentate gyrus, fails to achieve young 0.0004) compared to unhandled controls (Fig. 2H–J and adult levels following chronic fluoxetine treatment. This Table 1A). At PN 62, dendritic spine density in fluoxetine- immature level of spine density persists for at least 3 treated animals was similar to that found at PN 22 (Fig. 3 weeks. In addition, acute, but not chronic, SSRI treatment
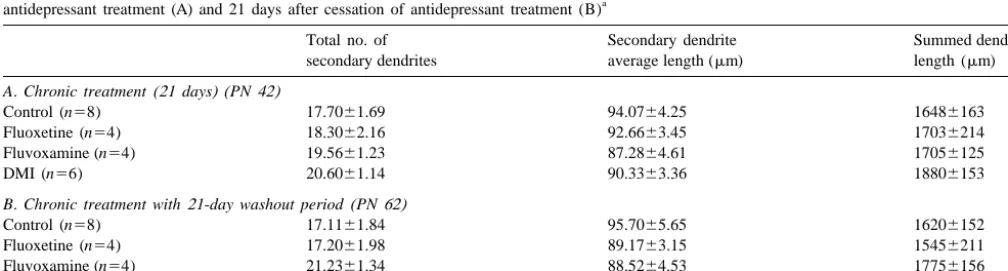
Table 3
Summary of the number of secondary dendritic branches, average length per dendrite, and summed dendritic length (6S.E.M.) in CA1 following 21-day a
antidepressant treatment (A) and 21 days after cessation of antidepressant treatment (B)
Total no. of Secondary dendrite Summed dendritic
secondary dendrites average length (mm) length (mm)
A. Chronic treatment (21 days) (PN 42)
Control (n58) 17.7061.69 94.0764.25 16486163
Fluoxetine (n54) 18.3062.16 92.6663.45 17036214
Fluvoxamine (n54) 19.5661.23 87.2864.61 17056125
DMI (n56) 20.6061.14 90.3363.36 18806153
B. Chronic treatment with 21-day washout period (PN 62)
Control (n58) 17.1161.84 95.7065.65 16206152
Fluoxetine (n54) 17.2061.98 89.1763.15 15456211
Fluvoxamine (n54) 21.2361.34 88.5264.53 17756156
DMI (n56) 20.5061.95 88.4364.17 17516131
a
produced an increase in total dendritic length in CA1 but BDNF [35,88]. Antidepressant treatment did not affect not in the dentate gyrus. secondary dendritic length or number in the dentate gyrus. The increase in spine density produced by fluvoxamine was not replicated by fluoxetine treatment. The disparity in 4.1. Acute antidepressant treatment the effects of fluoxetine and fluvoxamine (both potent SSRIs) may be due to differences in the pharmacological Antidepressant drug treatment (and subsequent alteration profile of each drug. When compared to fluoxetine, fluvox-of hippocampal serotonin and NE) was initiated on PN 21 amine is more selective, less potent, has a shorter elimina-for several reasons. In rat, serotonergic projections to the tion half-life, and is less heavily bound to plasma proteins hippocampus proliferate during the first 3 postnatal weeks [43,86]. These differences in the pharmacological prop-and reach the adult state at the end of the third week. erties of each drug may contribute to the different effects During the first 3 postnatal weeks, serotonin assumes a of the drugs on dendritic spine density.
regulatory role in neuronal migration and differentiation
during pre- and postnatal development [17–19,23,50,81– 4.2. Chronic antidepressant treatment 86]. For example, serotonin re-uptake transporters have
been implicated in the mechanism underlying neuronal The present study demonstrates that 21-day fluoxetine migration, and are present on serotonergic axonal growth treatment in juvenile rats prevents the age-associated cones as early as embryonic day 15 [31,44]; these re- elevation in dendritic spine density observed in control uptake sites are capable of binding SSRIs [44]. Binding of animals. Analysis of fluoxetine-treated animals showed SSRIs to re-uptake sites prior to PN 21 may therefore have that the number of spines per unit length is lower with no a profound effect on neuronal development. Dendritic significant overall change in dendritic length. Thus, it is spine formation on CA1 pyramidal cells also begins during reasonable to conclude that chronic fluoxetine treatment the first 3 postnatal weeks with growth continuing into affects spine formation and retention. One possibility is young adulthood (approximately PN 42) [69]. that new spines are formed from PN 21 to 42 (as seen in A single injection of fluvoxamine induced a significant control animals) but not retained. A second possibility is increase in dendritic spine density on pyramidal cells in the that fluoxetine prevents the formation of new spines during stratum radiatum sector of CA1 and on dentate granule this period; the number of spines observed following the cells. To our knowledge, although single SSRI injections 3-week fluoxetine treatment was markedly similar to that elevate serotonin levels in the hippocampus [72], the effect observed in 21-day-old control animals.
of this serotonin elevation on dendrites in juvenile rat brain The mechanism underlying the lower spine density in has not been widely investigated. Chronic alteration of CA1 is currently unknown. This issue is complicated by hippocampal serotonin level, however, impacts dendritic inconsistencies in the literature on serotonin transmission morphology in both juvenile and adult brains and chronic fluoxetine administration. Chronic fluoxetine [53,54,62,90]. Evidence that 5-HT maintains dendritic treatment has been reported to decrease whole brain spines comes from studies demonstrating a significant concentrations of both serotonin and its major metabolite, reduction in hippocampal spine density following removal 5-hydroxyindoleacetic acid (5-HIAA) [8,40,78]. In hip-of serotonergic innervation [12,53,55,62,89]. For example, pocampus, baseline serotonin levels remain reduced as the removal of serotonergic innervation during neonatal long as 7–14 days after termination of chronic fluoxetine development results in a decrease in the 5-HT1A receptor- treatment [78]. Conversely, repeated fluoxetine treatment mediated release of S-100b growth factor from astroglial was reported to induce a 3-fold increase in baseline cells; this loss of growth factor is believed to mediate the serotonin levels in the hippocampus [48]. Fluoxetine-in-spine loss [23,80–85]. The observed increase in hippocam- duced increases in extracellular serotonin are believed to pal spine density following a single fluvoxamine injection result from somatodendritic and axon terminal autoreceptor may be the result of an enhanced release of S-100B. activity. Initially, the fluoxetine-induced increase in ex-The number and length of secondary dendrites were tracellular serotonin levels is offset by 5-HT1A autorecep-analyzed in CA1 and dentate gyrus to determine if the tor activity [4,34,48,68]; these autoreceptors become de-apparent increase in dendritic spine density was simply the sensitized following chronic treatment [4,21,22]. Repeated result of shortened secondary dendrites. The data show, fluoxetine treatment has also been reported to progressive-however, that all three antidepressants increased dendritic ly increase extracellular 5-HT levels with subsequent length in CA1. In addition, all three antidepressants challenge injections [67]. These studies demonstrate that increased the number of secondary dendritic branches in the effect of chronic fluoxetine administration on hip-CA1 but the fluvoxamine-related change was not statisti- pocampal serotonin levels remains unclear.
density and dendritic atrophy [51,53,54,56,57]. Dendritic immature spine density persists and whether this effect is atrophy in the hippocampus is produced by repeated stress present in animals treated as adults.
and is accompanied by elevated serotonin levels [11], an The lower spine density observed at PN 62 may be effect that is attenuated by the serotonin uptake enhancing associated with an enduring alteration in serotonergic drug tianeptine [56]. In other words, reducing the elevated transmission in hippocampus. Animals chronically treated levels of extracellular serotonin prevents or reverses stress- with serotonergic agents either perinatally or as adults have induced atrophy. In the same stress paradigm, dendritic shown enduring changes. Offspring of rats administered spine number per unit length of dendrite actually in- fluoxetine from embryonic days 13 to 20 had reduced creased, suggesting that spines were retained even as serotonin content in frontal cortex as juveniles and in the dendritic length decreased. On the other hand, dramatic midbrain as adults [6]. In addition, reduced or blocked reduction of serotonin levels in the adult hippocampus can serotonergic transmission during pre- and postnatal de-impair memory acquisition and decrease synaptic density velopment results in reduced synapse number and smaller [2,53,54]; since most synapses are on dendritic spines, this hippocampal volume during adulthood [89]. Further, adults very likely involves a reduction in spine density as well treated with fluoxetine for 21 days (30 mg / kg) displayed a [32]. Lower spine density may be the result of a significant significant reduction in hippocampal serotonin up to 14 loss of the trophic factor S-100 which is serotonergically days after cessation of drug treatment [78]. Thus, it is regulated [23,80–85]. Taken together, these data suggest reasonable to suggest that the enduring lower spine density that alteration of serotonin neurotransmission in the raphe- results from a persistent fluoxetine-induced reduction in hippocampal system can substantially impact the number 5-HT levels.
of spines within hippocampal subfields.
Chronic fluoxetine initiates a number of intracellular 4.4. Unhandled vs. DMI- or fluvoxamine-treated animals changes that may be, in part, responsible for arresting the
development of dendritic spines. For example, in CA1, Compared to unhandled controls, animals chronically chronic fluoxetine treatment increases activity along 5-HT treated with DMI or fluvoxamine had lower dendritic spine and NE receptor-mediated cascades [21,22,61]. These density in CA1 when analyzed 1 or 21 days after the last receptor subtypes can increase cAMP levels that activate injection. Compared to saline injected controls, however, cAMP-dependent protein kinase (PKA) which in turn there were no significant changes. The lower dendritic phosphorylates cAMP response element binding protein spine density likely reflects an additive effect of the drug (CREB) [28,58]. Specific proteins whose expression is and the injection per se. These data suggest that the DMI increased by CREB include brain derived neurotrophic and fluvoxamine may affect spine density if delivered factor (BDNF) and its associated receptor trkB [14]. through a less stressful route of administration such as Expression of BDNF and trkB is enhanced in CA1 osmotic mini-pump or oral ingestion.
following chronic fluoxetine administration in adult rats It has been reported that spine density in CA1 increases [21,22,61]. In cultured rat hippocampal neurons, BDNF in response to both i.p. saline and clomipramine injection attenuates estradiol-induced increases in dendritic spine [37]. This effect was limited to basal dendrites in stratum density [60]. Further, elevated BDNF in ferret cerebral oriens only, and was interpreted as a result of stress. These cortex results in destabilization of newly formed spines results are not directly comparable to those of the present [35]. In the present study, it is quite possible that a study for several reasons: animal age (juvenile vs. adult) fluoxetine-induced increase in BDNF may have destabil- and strain (Sprague–Dawley vs. Wistar, respectively), and ized new spines thus arresting spine density at the PN 21 type of antidepressant administered (fluoxetine vs. clomi-level in 42-day-old animals. pramine).
4.3. Chronic antidepressant treatment with 21-day 4.5. Conclusions
washout
re-creased density of multiple-head dendritic spines on medium-sized
tention of new spines which typically occurs during the
spiny neurons of the striatum in rats reared in a complex
environ-second postnatal month in the rat hippocampus.
ment, Neurobiol. Learn. Mem. 66 (1996) 93–96.
[14] D.F. Condorelli, P. Dell’Albani, G. Mudo, T. Timmusk, N. Bel-lurado, Expression of neurotrophins and their receptors in primary astroglial cultures: induction by cAMP elevating agents, J.
Neuro-Acknowledgements
chem. 63 (1994) 509–516.
[15] E.F. da Cruz e Silva, O.A.B. da Cruz e Silva, C.T.B.V. Zaia, P.
We acknowledge Drs. Eric Nestler and Ronald Duman Greengard, Inhibition of protein phosphatase 1 stimulates secretion for their advice and gift of fluoxetine and Charles Badland, of Alzheimer amyloid precursor protein, Mol. Med. 1 (1995) 535–
541.
Elizabeth Sullivan, and Amanda Clark for their technical
[16] N.L. Desmond, W.B. Levy, Changes in the numerical density of
assistance. This work was supported by a grant from NIH
synaptic contacts with long-term potentiation in the hippocampal
RO3MH58834-01 awarded to C.O. and a Florida State
dentate gyrus, J. Comp. Neurol. 253 (1986) 466–475.
University Dissertation Grant awarded to S.N. [17] A. Dinopoulos, I. Dori, J.G. Parnavelas, Serotonergic innervation of
the mature and developing lateral septum of the rat: a light and electron microscopic immunocytochemical analysis, Neuroscience 55 (1993) 209–222.
References [18] A. Dinopoulos, I. Dori, J.G. Parnavelas, The serotonin innervation
of the basal forebrain shows a transient phase during development, Brain Res. Dev. Brain Res. 99 (1997) 38–52.
[1] R.A. Altschuler, Morphometry of the effect of increased experience
and training on synaptic density in area CA3 of the rat hippocampus, [19] I. Dori, A. Dinopoulos, M.E. Blue, J.G. Parnavelas, Regional J. Histochem. Cytochem. 27 (1979) 1548–1550. differences in the ontogeny of the serotonergic projection to the
cerebral cortex, Exp. Neurol. 138 (1996) 1–14. [2] E.C. Azmitia, V.J. Rubinstein, J.A. Strafaci, J.C. Rios, P.M.
Whitaker-Azmitia, 5-HT1A agonist and dexamethasone reversal of [20] R.S. Duman, G.R. Heninger, E.J. Nestler, Molecular psychiatry. para-chloroamphetamine induced loss of MAP-2 and synaptophysin Adaptations of receptor-coupled signal transduction pathways under-immunoreactivity in adult rat brain, Brain Res. 677 (1995) 181–192. lying stress- and drug-induced neural plasticity, J. Nerv. Ment. Dis.
182 (1994) 692–700. [3] S.G. Beck, S. Birnsteil, K.C. Choi, W.A. Pouliot, Fluoxetine
selectively alters 5-hydroxytryptamine1A and gamma-aminobutyric [21] R.S. Duman, G.R. Heninger, E.J. Nestler, A molecular and cellular acid receptor-mediated hyperpolarization in area CA1, but not areaB theory of depression, Arch. Gen. Psychiatry 54 (1997) 597–606. CA3, hippocampal pyramidal cells, J. Pharmacol. Exp. Ther. 281 [22] R.S. Duman, V.A. Vaidya, M. Nibuya, S. Morinobu, L. Rydelek (1997) 115–122. Fitzgerald, Stress, antidepressant treatments, and neurotrophic fac-[4] P. Blier, Y. Chaput, C. deMontigny, Long-term 5-HT re-uptake tors: molecular and cellular mechanisms, Neuroscientist 1 (1995)
blockade, but not monoamine oxidase inhibition, decreases the 351–360.
function of terminal 5-HT autoreceptors: an electrophysiological [23] K.M. Faber, J.H. Haring, Synaptogenesis in the postnatal rat fascia study in the rat brain, Naunyn Schmiedeberg’s Arch. Pharmacol. dentata is influenced by 5-HT1areceptor activation, Dev. Brain Res.
337 (1988) 246–254. 114 (1999) 245–252.
[5] J.W. Brock, C. Prasad, Alterations in dendritic spine density in the [24] P.M. Field, G. Raisman, The density of reinnervation of adult rat rat brain associated with protein malnutrition, Dev. Brain Res. 66 superior cervical sympathetic ganglionic neurons is limited by the (1992) 266–269. number of available postsynaptic sites, Brain Res. 360 (1985) [6] T.M. Cabrera-Vera, F. Garcia, W. Pinto, G. Battaglia, Effect of 398–402.
prenatal fluoxetine (Prozac) exposure on brain serotonin neurons in [25] E. Fifkova, A. Van Harreveld, Long-lasting morphological changes prepubescent and adult male rat offspring, J. Pharmacol. Exp. Ther. in dendritic spines of dentate granular cells following stimulation of 280 (1997) 138–145. the entorhinal area, J. Neurocytol. 6 (1977) 211–230.
[7] S. Caccia, Anorectic activity of fluoxetine and norfluoxetine in rats: [26] J.R. Fozard, Neuronal 5-HT receptors in the periphery, Neuro-relationship between brain concentrations and in-vitro potencies on pharmacology 23 (1984) 1473–1486.
monoaminergic mechanisms, J. Pharm. Pharmacol. 44 (1992) 250– [27] Y. Geinisman, F. Morrell, L. de Toledo-Morrell, Perforated synapses 254. on double-headed dendritic spines: a possible structural substrate of [8] S. Caccia, Effects of short- and long-term administration of fluox- synaptic plasticity, Brain Res. 480 (1989) 326–329.
etine on the monoamine content of rat brain, Neuropharmacology 31 [28] A. Ghosh, M.E. Greenberg, Calcium signaling in neurons: molecular (1992) 343–347. mechanisms and cellular consequences, Science 268 (1995) 239– [9] A.K. Cadogan, C.A. Marsden, I. Tulloch, D.A. Kendall, Evidence 247.
that chronic administration of paroxetine or fluoxetine enhances [29] W.T. Greenough, A. Alcantara, N. Hawrylak, B.A. Anderson, T. 5-HT2 receptor function in the brain of the guinea pig, Neuro- Karr, I.J. Weiler, Determinants of brain readiness for action: pharmacology 32 (1993) 249–256. Experience shapes more than neuronal form, Brain Dysfunct. 5 [10] M.O. Carruba, G.B. Picotti, F. Zambotti, P. Mantegazza, Effects of (1992) 129–149.
mazindol, fenfluramine, and chlorimipramine on the 5-hydroxy- [30] Q. Gu, W. Singer, Involvement of serotonin in developmental tryptamine uptake and storage mechanisms in rat brain: similarities plasticity of kitten visual cortex, Eur. J. Neurosci. 7 (1995) 1146– and differences, Naunyn-Schmiedeberg’s Arch. Pharmacol. 300 1153.
(1977) 227–232. [31] S.R. Hansson, E. Mezey, B.J. Hoffman, Serotonin transporter [11] F. Chaouloff, Physiopharmacological interactions between stress messenger RNA in the developing rat brain: early expression in hormones and central serotonergic systems, Brain Res. Brain Res. serotonergic neurons and transient expression in non-serotonergic Rev. 18 (1993) 1–32. neurons, Neuroscience 83 (1998) 1185–1201.
[12] L. Cheng, K. Hamaguchi, M. Ogawa, S. Hamada, N. Okado, PCPA [32] K.M. Harris, S.B. Kater, Dendritic spines: cellular specializations reduces both monoaminergic afferents and non-monoaminergic imparting both stability and flexibility to synaptic function, Annu. synapses in the cerebral cortex, Neurosci. Res. 19 (1993) 111. Rev. Neurosci. 17 (1994) 341–371.
systems and the regulation of dendritic growth and spine formation, [54] M. Matsukawa, M. Ogawa, K. Nakadate, T. Maeshima, Y. Ichitani, J. Comp. Neurol. 281 (1989) 13–22. N. Kawai, N. Okado, Serotonin and acetylcholine are crucial to maintain hippocampal synapses and memory acquisition in rats, [34] I. Hervas, F. Artigas, Effect of fluoxetine on extracellular
5-hy-Neurosci. Lett. 230 (1997) 13–16. droxytryptamine in rat brain. Role of 5-HT autoreceptors, Eur. J.
Pharmacol. 358 (1998) 9–18. [55] C. Mazer, J. Muneyyirci, K. Taheny, N. Raio, A. Borella, P.M. ¨ Whitaker-Azmitia, Serotonin depletion during synaptogenesis leads [35] H.W. Horch, A. Kruttgen, S.D. Portbury, L.C. Katz, Destabilization
to decreased synaptic density and learning deficits in the adult rat: a of cortical dendrites and spines by BDNF, Neuron 23 (1999)
possible model of neurodevelopmental disorders with cognitive 353–364.
deficits, Brain Res. 760 (1997) 68–73. [36] C.H. Horner, Plasticity of the dendritic spine, Prog. Neurobiol. 41
[56] B.S. McEwen, C.D. Conrad, Y. Kuroda, M. Frankfurt, A.M. (1993) 281–321.
Magarinos, C. McKittrick, Prevention of stress-induced morphologi-[37] C.H. Horner, M. O’Regan, E. Arbuthnott, Neural plasticity of the
cal and cognitive consequences, Eur. Neuropsychopharmacol. 7 hippocampal (CA1) pyramidal cell — quantitative changes in spine
(Suppl. 3) (1997) S323–S328. density following handling and injection for drug testing, J. Anat.
[57] C.R. McKittrick, B.S. McEwen, Regulation of serotonergic function 174 (1991) 229–238.
in the CNS by steroid hormones and stress, in: T.W. Stone (Ed.), [38] C.H. Horner, M. O’Regan, E. Arbuthnott, A quantitative study of
CNS Neurotransmitters and Neuromodulators: Neuroactive Steroids, the perikaryon and the basal dendritic tree in rat hippocampus
CRC Press, Boca Raton, FL, 2000, pp. 37–76. (CA1) pyramids following different experimental procedures, Ir. J.
[58] T.E. Meyer, HabenerJ.F., Cyclic adenosine 39,59-monophosphate Med. Sci. 162 (1993) 258–262.
response element binding protein (CREB) and related transcription-[39] K. Hosoda, R.S. Duman, Regulation of beta 1-adrenergic receptor
activating deoxyribonucleic acid-binding proteins, Endocr. Rev. 14 mRNA and ligand binding by antidepressant treatments and
norepi-(1993) 269–290. nephrine depletion in rat frontal cortex, J. Neurochem. 60 (1993)
[59] M.B. Moser, M. Trommald, P. Andersen, An increase in dendritic 1335–1343.
3
spine density on hippocampal CA1 pyramidal cells following spatial [40] P.D. Hrdina, Regulation of high- and low-affinity [ H]imipramine
learning in adult rats suggests the formation of new synapses, Proc. recognition sites in rat brain by chronic treatment with
antidepres-Natl. Acad. Sci. USA 91 (1994) 12673–12675. sants, Eur. J. Pharmacol. 138 (1987) 159–168.
[60] D.D. Murphy, N.B. Cole, M. Segal, Brain-derived neurotrophic [41] P.D. Hrdina, T.B. Vu, Chronic fluoxetine treatment upregulates 5-HT
factor mediates estradiol-induced dendritic spine formation in uptake sites and 5-HT receptors in rat brain: an autoradiographic2
hippocampal neurons, Proc. Natl. Acad. Sci. USA 95 (1998) 11412– study, Synapse 14 (1993) 324–331.
11417. [42] E.C. Hwang, I. Magnussen, M.H. Van Woert, Effects of chronic
[61] M. Nibuya, E.J. Nestler, R.S. Duman, Chronic antidepressant fluoxetine administration on serotonin metabolism, Res. Commun.
administration increases the expression of cAMP response element Chem. Pathol. Pharmacol. 29 (1980) 79–98.
binding protein (CREB) in rat hippocampus, J. Neurosci. 16 (1996) [43] J. Hyttel, Pharmacological characterization of selective serotonin
2365–2372. reuptake inhibitors (SSRIs), Int. Clin. Psychopharmacol. 9 (Suppl.
1) (1994) 19–26. [62] N. Okado, L. Cheng, Y. Tanatsugu, S. Hamada, K. Hamaguchi, [44] N. Ivgy-May, H. Tamir, M.D. Gershon, Synaptic properties of Synaptic loss following removal of serotoninergic fibers in newly
serotonergic growth cones in developing rat brain, J. Neurosci. 14 hatched and adult chickens, J. Neurobiol. 24 (1993) 687–698. (1994) 1011–1029. [63] A. Peters, S.L. Palay, H.F. de Webster, The Fine Structure of the [45] P.N. Izzo, A.M. Graybiel, J.P. Bolam, Characterization of substance Nervous System: The Neurons and Supporting Cells, Saunders,
P- and [Met]enkephalin-immunoreactive neurons in the caudate Philadelphia, PA, 1976.
nucleus of the cat and ferret by a single section golgi procedure, [64] T.E. Robinson, B. Kolb, Persistent structural modifications in Neuroscience 20 (1987) 577–587. nucleus accumbens and prefrontal cortex neurons produced by [46] J.M. Kerns, D.K. Sierens, L.C. Kao, H.L. Klawans, P.M. Carvey, previous experience with amphetamine, J. Neurosci. 17 (1997)
Synaptic plasticity in the rat striatum following chronic haloperidol 8491–8497.
treatment, Clin. Neuropharmacol. 15 (1992) 488–500. [65] T.E. Robinson, B. Kolb, Alterations in the morphology of dendrites [47] C. Koch, A. Zador, The function of dendritic spines: Devices and dendritic spines in the nucleus accumbens and prefrontal cortex subserving biochemical rather than electrical compartmentalization, following repeated treatment with amphetamine or cocaine, Eur. J. J. Neurosci. 13 (1993) 413–422. Neurosci. 11 (1999) 1598–1604.
[48] D.S. Kreiss, I. Lucki, Effects of acute and repeated administration of [66] M. Rocha, M. Sur, Rapid acquisition of dendritic spines by visual antidepressant drugs on extracellular levels of 5-hydroxytryptamine thalamic neurons after blockade of N-methyl-D-aspartate receptors, measured in vivo, J. Pharmacol. Exp. Ther. 274 (1995) 866–876. Proc. Natl. Acad. Sci. USA 92 (1995) 8026–8030.
[49] D.A. Lewis, L.A. Glantz, Specificity of decreased s pine density on [67] J.J. Rutter, C. Gundlah, S.B. Auerbach, Increase in extracellular layer III pyramidal cells in schizophrenia (Abstract), Schizophr. Res. serotonin produced by uptake inhibitors is enhanced after chronic 24 (1997) 39. treatment with fluoxetine, Neurosci. Lett. 171 (1994) 183–186. [50] H.G. Lidov, M.E. Molliver, Immunohistochemical study of the [68] J.J. Rutter, C. Gundlah, S.B. Auerbach, Systemic uptake inhibition
development of serotonergic neurons in the rat CNS, Brain Res. decreases serotonin release via somatodendritic autoreceptor
activa-Bull. 9 (1982) 559–604. tion, Synapse 20 (1995) 225–233.
[51] A.M. Magarinos, A. Deslandes, B.S. McEwen, Effects of antide- [69] P.A. Schwartzkroin, D.D. Kunkel, L. Mathers, Development of pressants and benzodiazepine treatments on the dendritic structure of rabbit hippocampus: anatomy, Brain Res. 254 (1981) 453–468. CA3 pyramidal neurons after chronic stress, Eur. J. Pharmacol. 371 [70] M. Segal, Morphological alterations in dendritic spines of rat (1999) 113–122. hippocampal neurons exposed to N-methyl-D-aspartate, Neurosci. [52] I. Malagie, A.C. Trillat, C. Jacquot, A.M. Gardier, Effects of acute Lett. 193 (1995) 73–76.
fluoxetine on extracellular serotonin levels in the raphe: an in vivo [71] F.J. Seil, F.P. Eckenstein, P.J. Reier, Induction of dendritic spine microdialysis study, Eur. J. Pharmacol. 286 (1995) 213–217. proliferation by an astrocyte secreted factor, Exp. Neurol. 117 [53] M. Matsukawa, K. Nakadate, N. Okado, Synaptic loss following (1992) 85–89.
removal of serotonergic and / or noradrenergic fibers in visual cortex [72] D.B. Shachar, E. Klein, A. Tabak, J.P.M. Finberg, Effect of single of the developing and adult rat, Soc. Neurosci. Abstr. 21 (1995) and repeated administration of fluvoxamine on noradrenaline release
[73] O. Steward, S.L. Vinsant, The process of reinnervation in the dentate [82] P.M. Whitaker-Azmitia, E.C. Azmitia, Stimulation of astroglial gyrus of the adult rat: a quantitative electron microscopic analysis of serotonin receptors produces culture media which regulates growth terminal proliferation and reactive synaptogenesis, J. Comp. Neurol. of serotonergic neurons, Brain Res. 497 (1989) 80–85.
214 (1983) 370–386. [83] P.M. Whitaker-Azmitia, E.C. Azmitia, Astroglial 5-HT1a receptors [74] M.G. Stewart, M. Lowdnes, A. Hunter, T. Doubell, Memory storage and S-100 beta in development and plasticity, Perspect. Dev.
in chicks involves an increase in dendritic spine number and Neurobiol. 2 (1994) 233–238.
synaptic density, Brain Dysfunct. 5 (1992) 50–64. [84] P.M. Whitaker-Azmitia, R. Murphy, E.C. Azmitia, Stimulation of [75] M. Sunanda, M.S. Rao, T.R. Raju, Effect of chronic restraint stress astroglial 5-HT1Areceptors releases the serotonergic growth factor, on dendritic spines and excrescences of hippocampal CA3 pyrami- protein S-100, and alters astroglial morphology, Brain Res. 528 dal neurons — a quantitative study, Brain Res. 694 (1995) 312–317. (1990) 155–158.
[76] M. Trommald, G. Hulleberg, P. Andersen, Long-term potentiation is [85] P.M. Whitaker-Azmitia, A.V. Shemer, J. Caruso, L. Molino, E.C. associated with new excitatory spine synapses on rat dentate granule Azmitia, Role of high affinity serotonin receptors in neuronal cells, Learn. Mem. 3 (1996) 218–228. growth, Ann. N.Y. Acad. Sci. 600 (1990) 315–330.
[77] M. Trommald, T.W. Vaaland, T.W. Blackstad, P. Andersen, Dendritic [86] D.T. Wong, F.P. Bymaster, E.A. Engelman, Prozac (fluoxetine, Lilly spine changes in rat dentate granule cells associated with long-term 110140), the first selective serotonin uptake inhibitor and an potentiation, in: A. Guidotti (Ed.), Neurotoxicity of Excitatory antidepressant drug: twenty years since its first publication, Life Sci. Amino Acids, Raven Press, New York, 1990, pp. 163–174. 57 (1995) 411–441.
[78] J.H. Trouvin, A.M. Gardier, E. Chanut, N. Pages, C. Jacquot, Time [87] C.S. Woolley, B.S. McEwen, Estradiol mediates fluctuation in course of brain serotonin metabolism after cessation of long-term hippocampal synapse density during the estrous cycle in the adult fluoxetine treatment in the rat, Life Sci. 52 (1993) L187–L192. rat, J. Neurosci. 12 (1992) 2549–2554.
[79] J.K. Wamsley, W.F. Byerly, R.T. McCabe, E.J. McConnell, T.M. [88] G.Y. Wu, D.J. Zou, I. Rajan, H. Cline, Dendritic dynamics in vivo Dawson, B.I. Grosser, Receptor alterations associated with change during neuronal maturation, J. Neurosci. 19 (1999) 4472– serotonergic agents: an autoradiographic analysis, J. Clin. Psychiatry 4483.
48 (1987) 19–25. [89] W. Yan, C.C. Wilson, J.H. Haring, 5-HT1a receptors mediate the [80] M. Werner, H. Hatt, K. Gottman, Synapse formation and mor- neurotrophic effect of serotonin on developing dentate granule cells,
phological differentiation of neuron types in embryonic rat dentate Brain Res. Dev. Brain Res. 98 (1997) 185–190.
gyrus explants in vitro, Brain Res. Dev. Brain Res. 105 (1998) [90] W. Yan, C.C. Wilson, J.H. Haring, Effects of neonatal serotonin 9–23. depletion on the development of rat dentate granule cells, Brain Res. [81] P.M. Whitaker-Azmitia, Role of serotonin and other neurotransmitter Dev. Brain Res. 98 (1997) 177–184.