TANAMAN MAJA
Sebaran dan Aspek Historik Tanaman Maja
Maja atau bilwa, bael, Bengal quince, shirpal, bael fruits adalah sebutan lain bagi spesies Aegle marmelos(L.) Correa, sebagaimana disebutkan di India (Ghermandi 2002). Tanaman ini berasal dari India, namun sekarang banyak tersebar di Srilanka, Pakistan, Bangladesh, Myanmar, Thailand dan sebagian besar Asia Tenggara (Gambar 1).
Ket : menunjukkan wilayah penyebaran tanaman maja
Gambar 1. Peta penyebaran tanaman maja.
Tanaman maja juga tersebar di berbagai wilayah di Indonesia, karena iklim yang cocok untuk tanaman ini sehingga dapat tumbuh dengan baik. Tanaman maja banyak ditemukan di dataran rendah, dataran tinggi, bahkan di tanah berkapur seperti di Pulau Jawa (Banten, Bogor, Yogyakarta, Jawa Timur dan Madura), Pulau Sumatera (Aceh, Palembang, Bukit tinggi), Pulau Sulawesi (Makasar), Kepulauan Maluku dan NTT (Sumba Timur). Keberadaan tanaman ini tercatat sejak jaman kerajaan Majapahit, sebagaimana terbukti adanya berbagai nama daerah yang diawali dengan kata maja yang menunjukkan keterkaitan dengan keberadaan tanaman ini di berbagai daerah. Tanaman ini dikenal sebagai maja (Sunda), brenuk atau maos (Jawa), bila paek (Madura), wabiila (Sumba Timur), dilok (Timor) (Hariana 2002).
Karakteristik Tanaman Maja
Tanaman maja dapat tumbuh subur di atas tanah dengan pH 5 – 8. Di India tumbuhan ini dapat tumbuh subur pada lahan dimana tanaman buah lain tidak dapat bertahan di sana, bahkan menurut Singh & Malik (2000) dinyatakan bahwa tanaman ini dapat tumbuh di tanah rawa, alkali, bahkan pada tanah berbatu kapur di Florida Selatan. Tanaman ini dapat mencapai ketinggian 13 m dan buahnya dapat dipanen jika sudah berwarna hijau kekuningan. Karakteristik tanaman maja antara lain daun berbentuk memanjang, tulang daun menyirip, susunan daun berkarang, percabangan besar. Satu pohon dapat menghasilkan 800 buah, dan dapat dipanen satu kali dalam setahun. Dalam 1 musim panen rata-rata 150-200 buah berukuran besar (1-3 kg). Buah maja rasanya manis, harum, dan tajam ditenggorokan. Bunganya bertipe berumah dua, kelopak bunganya kecil, dengan 5 mahkota dan bagian luar berbentuk bulat, lonjong, dagingnya beraroma khas dan berlendir (Anonim 2006).
Gambar 2. Foto Tanaman maja (koleksi pribadi)
Klasifikasi Tanaman Maja ( Aegle marmelos. Corr )
Kingdom : Plantae
Subkingdom : Tracheobionta Superdivisi : Spermatophyta
Divisi : Angiospermae
Klas : Magnolliopsida
Ordo : Sapindales/ Rutaceales Famili : Rutaceae
Genus : Aegle
Kandungan Kimia Tanaman Maja
Tanaman maja baik berupa akar, batang, daun, buah maupun biji banyak digunakan oleh masyarakat sebagai bahan pengobatan tradisional seperti demam, sembelit, disentri, diare, hepatitis, TBC, radang selaput lendir hidung, gangguan otak, gangguan hati, gatal-gatal (kudis, borok, bisul, eksim), anti inflamasi, gangguan pendengaran, gangguan urinaria, dan abortiva (Kapoor 1990, Singh & Malik 2000).
Akar tanaman maja mengandung bahan : epoxyaurapten, 4-methoxy-1-
methyl-2-quinolon, 4-sitosten-3-one, 7-o-methylmarmin, aurapten, kumarin, decursinol, dictamin, imperatorin, integriquinolon, lupeol, marmesin, marmin, marginal, scopoletin, skimmianin, umbelliferon, xanthotoxin, γ fagarin (Anonim
2006). Sedangkan pada batang terkandung bahan-bahan : fagarine, marmin,
4-epi-lyoniresinol,3-ά-O-β-D-glucopyranoside, 6-hydroxy-1-methoxymethylanthraquinon dictamine, magnesium, marmasin, silicone, β sitosterol. Selain mengandung bahan
seperti tersebut di atas, pada daun dapat diekstraksi bahan lain seperti : minyak
essensial, pellandrene, aegelin, rutacin, p-cymene, cíñeole, cuminaldehyd, d-limonene, marmelosin, N-2-ethoxy-2-phenyl- ethylcinnamid, p-cymene, rutin, skimmianin, tannin, β sitosterol- β-d-glukosid, dan γ-sitosterol. Demikian juga pada
buah mengandung bahan seperti : aegelin, aegelenin, alanin, alloimperatorin, methyl
ether ά-amirin, arginin, asam aspartat, boron, kalsium, karoten, chlorin, cis linalol oxida, cuprum, cystine, dictamin, d-ά pellandrene, asam glutamic, glisin, histidin, imperatorin, ferrum, isoamil asetat, isoleusin, asam linoleat, asam linolenat, lysin, magnesium, mangaan, marmelosin, marmelid, marmelin, marmesin, methionin, niasin, asam palmitat, o-isopentinylharfordinol, o-methylharford, pectin, phenyl alanin, pospor, polisakarida, potasium, proantocyanidin, prolin, psoralen, riboflavin, scoparon, scopoletin, serin, skimmin, sodium, asam stearat, tannin, asam tartat, thiamin, threonin, thyrosin, umbelliferon, valin, vinyl-butanoat, xanthotoxol, seng, β-amyrin, β-karoten, β-sitosterol, γ-fagarin (Anonim 2006).
Bahan kimia Aegelin (C18 H18 O4), β-sitosterol dan Stigmasterol (C29 H50 O) (Joy et al. 2001) berdasarkan hasil analisa fitokimia Riyanto dan Mawardi (2000) tergolong ke dalam steroid. Bahan aktif yang berupa sitosterol pada daun maja memiliki kerangka dasar berbentuk gugus atom triterpen seperti cincin steroid, yakni
sistem cincin siklopentana perhidrofenantren yang tersusun dari rangkaian 4 cincin yakni 3 cincin berbentuk segienam (A, B, C) dan cincin segilima (D). Adapun persamaan struktur kimia fitosterol daun maja dan steroid estrogen serta progesteron dapat dilihat pada Gambar 3. Steroid pada tumbuhan (fitosterol) akan memberikan efek anti implantasi maupun estrogenik (Partodiharjo 1980). Fitosterol ini memberikan rasa pahit pada daun maja.
Gambar 3. Struktur kimia steroid progesteron, estrogen (Estriol, β-Estradiol), β-sitosterol dan stigmasterol (Guyton 1996 & Harborne 1987).
BIOLOGI UMUM TIKUS
Tikus putih (Rattus norvegicus), merupakan jenis hewan yang sering dipergunakan sebagai hewan percobaan dalam penelitian biologis maupun biomedis baik secara in-vitro maupun in-vivo. Tikus putih atau tikus albino merupakan hasil perkawinan secara selektif sehingga memiliki karakter yang stabil. Ada beberapa galur atau varietas tikus antara lain : galur Sprague-dawley memiliki kepala kecil dan ekor lebih panjang dibandingkan dengan badannya, galur Wistar ditandai dengan kepala besar dan ekornya lebih pendek, serta galur long evans memiliki ukuran tubuh lebih kecil dengan kepala dan tubuh bagian depan berwarna hitam (Baker 1979). Sebagai hewan percobaan, tikus memerlukan ruangan yang cukup cahaya, sirkulasi udara yang
HO R β-Sitosterol HO Stigmasterol A B C D A B C D
baik, suhu yang cocok, sehingga dapat bergerak bebas dalam kandang, serta membutuhkan makanan yang cukup baik kualitas maupun kuantitasnya (Smith & Mangkoewidjojo 1988).
Tikus betina mencapai usia dewasa kelamin setelah berumur 8 minggu, dengan berat badan berkisar antara 200 – 300gr. Pada umur tersebut tikus sudah siap untuk dikawinkan. Periode kebuntingan berkisar antara 21 – 22 hari. Masa produktifnya cukup panjang yaitu pada umur 2 – 14 bulan. Selama periode tersebut tikus mampu melahirkan lebih dari 10 kali kelahiran dan jumlah anak yang dilahirkan rata-rata 9 ekor bahkan mencapai 12 ekor per kelahiran (Malole & Pramono 1989). Jumlah anak tikus pada kebuntingan pertama umumnya lebih sedikit daripada kebuntingan ke 5 – 7 (masa yang stabil) (Fox 2007).
Tikus termasuk hewan poliestrus. Tikus memiliki post partum estrus dalam waktu 48 jam sesudah partus (melahirkan). Perkawinannya sering terjadi pada malam hari. Untuk mengetahui terjadinya perkawinan dapat dilihat dari sumbat vagina atau vaginal plug guna memeriksa ada tidaknya spermatozoa setelah terjadi kopulasi (Malole & Pramono 1989).
Siklus Reproduksi Tikus Betina
Siklus reproduksi hewan mamalia betina melibatkan berbagai organ reproduksi yaitu ovarium, uterus, vagina, dan kelenjar mammae. Setiap organ tersebut mengalami siklus yang teratur dan berlangsung secara sinkron. Hal ini berarti siklus yang satu menjadi indikasi siklus yang lain. Siklus tersebut meliputi siklus ovarium, siklus uterus, siklus vagina, siklus estrus dan siklus kelenjar susu (Bullock et al. 2004).
Siklus estrus merupakan sederetan proses perubahan kegiatan fisiologis pada organ-organ reproduksi dari awal berahi hingga berulang kembali ke berahi berikutnya. Berahi adalah saat dimana hewan betina bersedia menerima pejantan untuk kopulasi (Partodihardjo 1980). Pada tikus siklus estrus ini berlangsung 4 – 5 hari (Malole & Pramono 1989). Siklus estrus dibedakan dalam 2 tingkatan, yaitu fase folikuler dan fase luteal. Fase folikuler merupakan fase perkembangan folikel sampai pecahnya folikel de Graaf saat ovulasi. Sedangkan fase luteal merupakan fase setelah ovulasi yaitu periode sekresi progesteron dan fase terbentuknya corpus luteum
(Nalbandov 1990). Bila tidak terjadi fertilisasi maka siklus baru berikutnya akan segera dimulai, dimana korpus luteum akan mengalami regresi dan menghilang. Namun jika terjadi pembuahan dan kebuntingan, korpus luteum akan terus bertahan selama masa kebuntingan. Pada kenyataannya korpus luteum merupakan kelenjar yang menghasilkan hormon progesteron yaitu hormon essential untuk mempertahankan kebuntingan (Frandson 1992). Berdasarkan histologi vagina, satu siklus estrus terbagi menjadi 4 fase, yaitu fase proestrus, estrus, metestrus, dan diestrus. Fase folikuler dimulai dengan proestrus yang diikuti oleh estrus dan ovulasi, sedangkan fase luteal terdiri atas metestrus dan diestrus (Partodihardjo 1980).
Ovarium
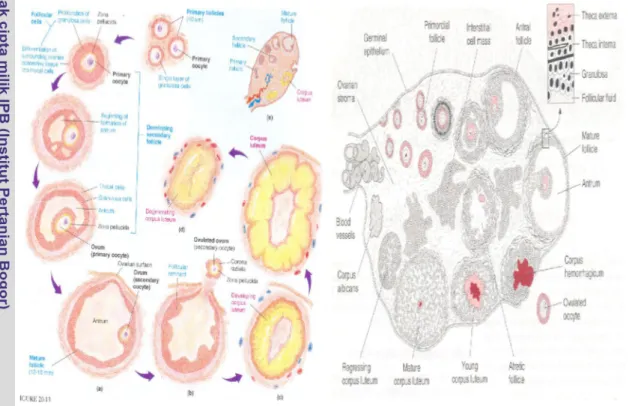
Tikus memiliki sepasang ovarium di dalam rongga abdomen, di bawah ginjal. Ovarium berbentuk bulat dengan permukaan yang berbenjol-benjol karena adanya sel-sel folikel dan korpus luteum (Gambar 4). Ovarium disel-selubungi oleh sel-selapis sel-sel epitel germinativum dan di bagian dalamnya terdapat tunika albugenia (Partodihardjo 1980 & Hafez 1993). Ovarium tersusun atas bagian korteks dan medulla. Bagian korteks terdiri atas stroma yang bersifat seluler dan mengandung folikel ovarium, korpus luteum dan sel interstitial serta pembuluh darah. Folikel terdiri atas oosit atau sel telur yang diselubungi oleh sel folikel yang merupakan hasil diferensiasi epitel germinativum. Folikel ovarium ada yang istirahat (primordial), dan ada yang sudah matang yaitu folikel de Graaf. Bagian medulla terdiri atas jaringan ikat fibroelastik yang penuh jaringan saraf, pembuluh darah dan limfe.
Ovarium memiliki 2 fungsi yaitu sebagai penghasil sel telur dan penghasil estrogen dan progesteron menurut Guyton (1996) & Lay Cock (1982). Proses pembentukan sel telur (Oogenesis) meliputi :
1. Proliferasi oogonium
Gamet yang berasal dari sel germinal primordial bermigrasi ke gonad dengan pergerakan amuboid melalui mesenterium dorsal ke gonad. Sel aktif bermitosis pada betina disebut Oogonium. Oogonia akan mengakhiri proliferasi setelah beberapa hari sebelum kelahiran (Ganong 2003).
2. Pertumbuhan Oosit
Pertumbuhan oosit dimulai dengan tumbuhnya ovum dan folikel secara cepat. Ketika antrum telah terbentuk, oosit tidak tumbuh lagi dan folikel mendapat pengaruh hormon dari hipofisis untuk melanjutkan tumbuh secara cepat. Pertumbuhan oosit ditandai dengan akumulasi kuning telur dalam sitoplasma, perkembangan zona pellucida, proliferasi mitosit dari epitel follikel dan jaringan di dekatnya (Hafez 2000).
Gambar 4. Ovarium, corpus luteum dan perkembangan folikel (Syahrum et al. 1994)
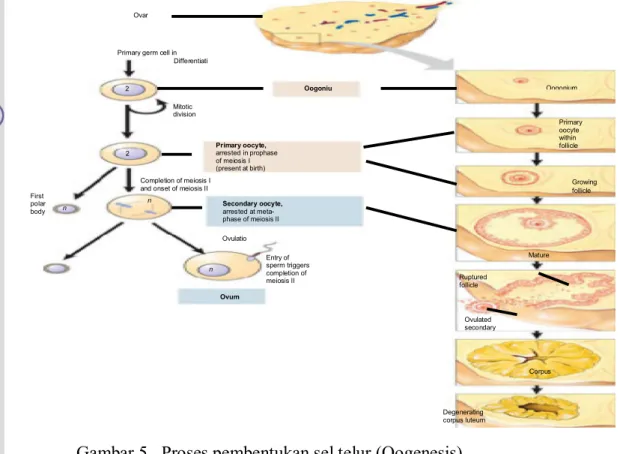
Oosit dikelilingi lapisan sel folikel granulosa. Oosit 1 mengalami meiosis 1 dan terjadi pada stadium akhir fetus. Oosit dilapisi beberapa lapis sel folikel yang disebut folikel primordial. Bila sel yang mengitari oosit ini lengkap maka disebut folikel primer. Pada proses ini oosit mengalami pembelahan meiosis 2 kali. Pada pembelahan meiosis 1, oosit primer menghasilkan 2 sel anak yaitu oosit sekunder dan badan kutub I yang masing-masing memiliki ½ perangkat kromosom (n) dari induknya 2n. Sedangkan pada pembelahan meiosis II, oosit sekunder menghasilkan 2 sel anak yaitu ootid dan badan kutub II. dan badan kutub I menghasilkan badan kutub III dan badan kutub IV. Masing-masing sel anak memiliki seperangkat kromosom.
Ovar
Primary germ cell in Differentiati Oogoniu m Oogonium Mitotic division Primary oocyte, arrested in prophase of meiosis I (present at birth) Completion of meiosis I
and onset of meiosis II
Primary oocyte within follicle Secondary oocyte, arrested at meta- phase of meiosis II First polar body Ovulatio n Entry of sperm triggers completion of meiosis II Ovum Growing follicle Mature Ruptured follicle Ovulated secondary Corpus Degenerating corpus luteum 2 n 2 n n n n
Badan kutub tersebut dilepas ke rongga perivitelin dan mengalami degenerasi (Hafez 2000).
Gambar 5. Proses pembentukan sel telur (Oogenesis) (Campbell et al. 2004).
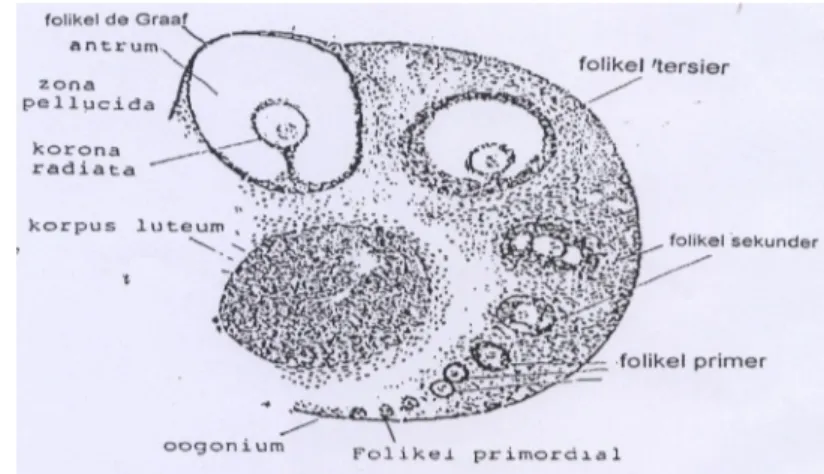
3. Perkembangan folikel
Tahap perkembangan folikel disebut fase folikuler. Fase ini dimulai dari
folikel primer. Selanjutnya sel folikel akan membelah diri untuk membentuk dinding
berlapis yang mengelilingi sel telur dan disebut folikel sekunder. Apabila sudah terbentuk rongga (antrum) diantara sel folikel dengan sel telur, maka folikel dikatakan sudah matang dan disebut folikel de Graaf. Folikel ini makin berkembang dan sel jaringan stroma bertambah banyak dan berdiferensiasi membentuk teka interna dan teka eksterna (Syahrum et al. 1994). Antrum berisi cairan dan semakin membesar sehingga sel telur terdesak ke tepi pada sel folikel. Sel telur bertambah besar yang dikelilingi membran granulosa, akhirnya sel folikel akan menempati seluruh tebal korteks. Hal ini mengakibatkan penipisan dan pecahnya dinding ovarium, sehingga sel telur yang diselubungi korona radiata dilepaskan ke dalam rongga peritoneum. Sedangkan pada tikus sel telur akan dilepas ke ruang periovarium (Rugs 1968). Proses
ini disebut ovulasi. Ovulasi pada tikus terjadi secara spontan selama fase estrus (Nalbandov 1990). Pada tikus, terdapat lebih dari satu folikel yang mengalami ovulasi dan menghasilkan 4 – 14 sel telur, yang memungkinkan kelahiran multiple (Smith & Mangkoewidjojo 1988).
Proses ovulasi diawali dengan perkembangan dan pematangan sel folikel dalam ovarium di bawah pengaruh FSH. Sel folikel yang sudah matang (folikel de Graaf) akan mensekresi estrogen yang menyebabkan kadar estrogen dalam darah meningkat, sehingga menekan sekresi FSH (mekanisme umpan balik negatif). Sebaliknya estrogen yang tinggi akan memacu sekresi LH sehingga terjadi ovulasi, setelah itu kadar estrogen dalam darah menurun (Syahrum et al. 1994). Folikel yang telah matang akan menempati daerah korteks dan menonjol ke permukaan ovarium. Pada permukaan yang menonjol terjadi penipisan jaringan. Cairan folikel makin banyak dan menyebabkan tekanan hidrostatik yang menyebabkan meningkatnya tekanan turgor jaringan. Di sisi lain kumulus ooforus mengalami desintegrasi sehingga ovum berada dalam keadaan bebas dalam cairan. Tegangan yang memuncak diikuti pecahnya selaput tipis ovarium pada stigma yang menyebabkan ovum lepas ke rongga peritonium (Ferin et al.1993). Skema proses pematangan folikel ovarium dapat dilihat pada Gambar 6 berikut :
Gambar 6. Ovarium tikus dengan tingkat perkembangan sel folikel (Hafez 1993).
Corpus Luteum
Setelah ovulasi terjadi, terbentuk lekukan pada permukaan ovarium yang telah melepaskan isinya kemudian terisi darah dan cairan limfe. Bagian folikel yang pecah kemudian bertaut kembali dan folikel pada saat ini dinamakan korpus hemoragicum.
Selanjutnya darah membeku diresorbsi dan terjadi lutenisasi sel-sel granulosa dan sel teka, sehingga terbentuk korpus luteum (Partodihardjo 1980). Pertumbuhan korpus luteum berlangsung melalui hipertrofi sel luteal. Pembentukan dan pertumbuhan korpus luteum dirangsang oleh LH. Sedangkan prolaktin (luteotropik hormon) berperan dalam memelihara fungsi korpus luteum agar tetap menghasilkan progesteron (Nalbandov 1990). Perkembangan selanjutnya dari korpus luteum tergantung pada terjadi tidaknya fertilisasi dan kebuntingan. Bila sel telur dibuahi dan terjadi kebuntingan maka korpus luteum akan tetap bertahan dan dikenal dengan korpus gravidatatum (CL kebuntingan) (Partodihardjo 1980). Apabila hewan betina tidak mengalami kebuntingan, maka korpus luteum mengalami regresi dan terjadi pematangan folikel yang baru. Regresi korpus luteum disertai munculnya sel tenun pengikat lemak dan struktur hialin diantara sel-sel luteum. Kondisi ini mempercepat regresi korpus luteum sampai sel luteum tidak terdapat lagi (Nalbandov 1990). Regresi korpus luteum berlangsung melalui kerja prostaglandin PGF2ά yang dihasilkan oleh uterus. PGF2ά akan merangsang vasokontriksi pembuluh darah pada korpus luteum sehingga suplai darah berkurang. Berkurangnya suplai darah menyebabkan luteolisis sel luteal (Sammuelson et al.1978).
Uterus
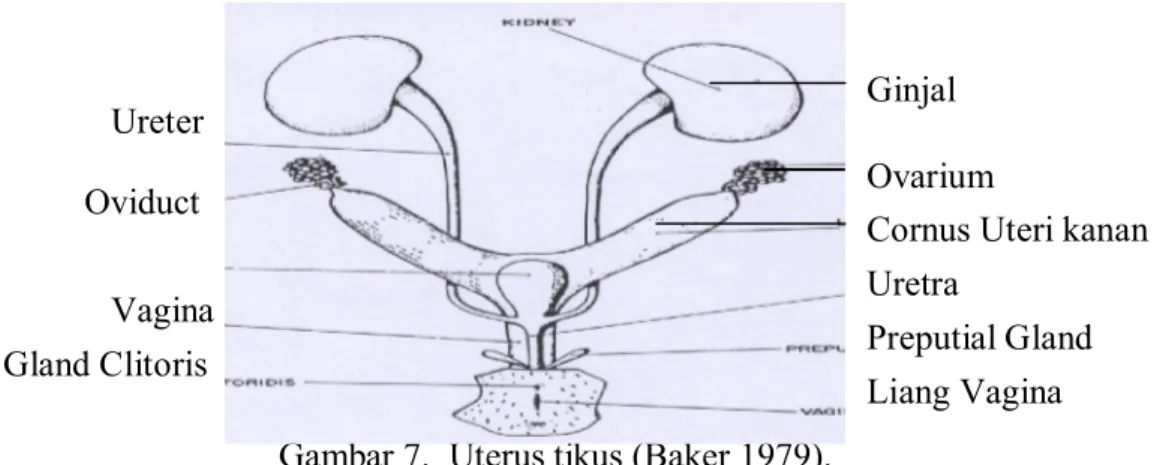
Uterus tikus merupakan tipe dupleks, yang terdiri atas dua tanduk (kornus uteri) dan satu badan yang bersatu membentuk huruf Y. Korpus uteri merupakan bagian pendek yang berbatasan dengan vagina. Sedangkan kornus uteri merupakan bagian uterus yang memanjang. Bagian proksimal berbatasan dengan tuba uterine, sedangkan bagian distalnya saling mendekat yang hanya dipisahkan oleh sekat tipis yang bermuara ke dalam lumen korpus uteri. Bagian uterus yang berbatasan langsung dengan vagina disebut serviks uteri (Baker 1979). Bagian-bagian uterus dapat dilihat pada gambar 7 berikut :
Gambar 7. Uterus tikus (Baker 1979).
Dinding uterus dari luar kedalam terdiri atas 3 lapisan yaitu membrana serosa, myometrium dan endometrium. Membrana serosa merupakan lapisan terluar. Myometrium merupakan lapisan otot yang tersusun atas pembuluh darah, limfe dan saraf. Endometrium merupakan lapisan dinding lumen yang terdiri atas epitel, kelenjar uterus dan tenunan pengikat. (Rugs 1968, Ross & Schreiber 1991).
Perubahan yang terjadi pada uterus selama siklus estrus disebut siklus uterus. Selama pertumbuhan folikel terjadi juga pertumbuhan dalam endometrium. Selama periode perkembangan korpus luteum, endometrium menyesuaikan diri untuk menerima kehamilan. Jika sel telur tidak dibuahi maka endometrium ke keadaan semula bersamaan dengan regresinya korpus luteum. Sedangkan jika terjadi pembuahan, maka endometrium dipertahankan pada keadaan yang terbaik untuk kehamilan (Sartono 1994).
Pada awal siklus estrus yaitu fase proestrus, folikel akan berkembang dan menghasilkan hormon estrogen untuk mempertahankan pertumbuhan maupun menyebabkan proliferasi endometrium. Dinding endometrium berangsur-angsur mengalami hiperemia, berproliferasi dan menebal dengan cepat. Kelenjar yang pendek menjadi bertambah panjang, dan pembuluh darah bertambah banyak (Bullock et al. 2004). Fase ini merupakan awal perkembangan folikel de Graaf. Fase proestrus ini merupakan periode terjadinya involusi fungsional korpus luteum serta pembengkakan praovulasi folikel (Mc Donald 1989). Selain itu pada tahapan ini juga terjadi vaskularisasi epitel vagina (Tolihere 1981). Vaskularisasi ini disebabkan oleh estrogen yang semakin tinggi. Di samping itu juga terdapatnya cairan yang terkumpul di uterus sehingga uterus sangat kontraktil (Turner & Bagnara 1976).
Ovarium
Cornus Uteri kanan Uretra Preputial Gland Liang Vagina Ureter Oviduct Vagina Gland Clitoris Ginjal
Pada fase estrus, estrogen meningkat merangsang sekresi kelenjar uterus dan merangsang serviks untuk berkontriksi, sehingga uterus menggelembung. Pada akhir fase ini, progesteron mulai dihasilkan oleh korpus luteum dan akan merangsang serviks untuk relaksasi. Hal ini menandai timbulnya hasrat kawin dan kopulasi mungkin terjadi pada tikus betina (Martin 1985). Fase ini berlangsung sekitar 12 jam (van Tienhoven 1983, Smith & Mangkoewidjojo 1988). Ciri khas pada fase ini hewan betina mau menerima pejantan untuk kopulasi. Hal ini disebabkan oleh pengaruh sekresi estrogen yang tinggi (Nalbandov 1990). Estrogen dari folikel de Graaf yang matang menyebabkan berbagai perubahan pada organ reproduksi seperti uterus tegang, mukosa vagina tumbuh cepat disertai sekresi lendir, terdapatnya sel-sel yang menumpuk dan menanduk. Sel-sel ini terkelupas ke dalam lumen vagina (Bullock et
al. 2004 & Rugs 1968).
Saat memasuki fase metestrus (setelah ovulasi), hasrat kawin menurun. Pada fase ini korpus luteum mulai tumbuh. Korpus luteum merupakan perubahan bentuk dari folikel de Graaf yang berubah fungsi setelah mengalami ovulasi. Uterus yang sudah berkembang mengalami oedema akibat pengaruh estrogen. Selanjutnya oleh pengaruh progesteron, kelenjar uterus aktif mensekresikan zat yang berguna untuk makanan dan proteksi terhadap embrio yang akan berimplantasi (Martin 1985 & Nelson 1983). Pembuluh darah bertambah panjang dan lebar. Apabila terjadi implantasi, mukosa endometrium makin menggelembung akibat pengaruh hormon estrogen dan progesteron. Mukosa endometrium menggelembung dan tempat implantasi zigot diketahui sebagai desidua (Nelson 1983). Pada akhir fase metestrus degenerasi epitel meningkat bahkan menghilang dan leukosit banyak terdapat pada dinding uterus (Bullock et al. 2004). Lapisan endometrium kembali ke fase istirahat dan dimulainya siklus baru yaitu fase diestrus. Pada fase ini hewan betina terlihat lebih tenang dan tidak ada aktivitas kelamin (van Tienhoven 1983). Pada fase ini korpus luteum menjadi matang dan pengaruh progesteron semakin nyata. Endometrium lebih menebal dan kelenjar membesar (Tolihere 1981), selaput mukosa vagina tipis, pucat, otot mengendur, uterus mengecil, anemik, dan agak kontraktil (Turner & Bagnara 1976). Fase ini berlangsung sekitar 60-70 jam (van Tienhoven
1983). Pada preparat ulas vagina terlihat banyaknya leukosit dan sel epitel berinti (Nalbandov 1990).
Vagina
Vagina merupakan saluran terpanjang yang terletak di bagian dorsal uretra dan bagian ventral rectum. Pembukaan vagina terjadi tidak lama setelah ovulasi pertama dan dipakai sebagai tanda pubertas (Hafez 2000). Perubahan karakteristik dinding epitel vagina selama estrus dapat diketahui melalui sediaan apus vagina yang merupakan petunjuk tahap-tahap dalam siklus estrus (Ganong 2003).
Siklus estrus merupakan cerminan bagi berbagai aktivitas yang saling berkaitan antara hipotalamus-hipofisis-ovarium. Selama siklus estrus terjadi perubahan baik pada organ reproduksi maupun perubahan tingkah laku seksual (Hafez 2000).
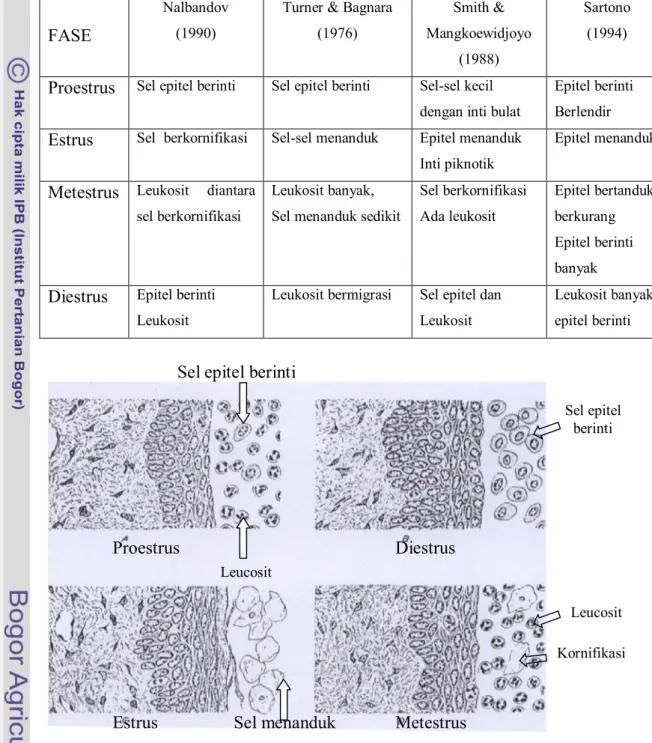
Meskipun peristiwa fisiologis yang utama pada siklus estrus terjadi pada ovarium, tetapi peristiwa tersebut tercermin pada vagina di bawah pengaruh hormon ovarium yaitu estrogen dan progesteron (Johnsons & Everith 1988). Perubahan siklus vagina tikus dapat dideteksi dengan menggunakan teknik preparat apus vagina. Gambaran perubahan yang terjadi di vagina dapat mencerminkan kejadian pada ovarium sehingga dapat digunakan untuk menentukan fase reproduksi pada tikus betina yang diperiksa (Sartono 1984). Apus vagina diperoleh dari sekret vagina yang diambil dengan menggunakan cotton bud pada 1/3 proksimal vagina (Hafez 2000). Adapun gambaran mikroskopis hasil ulas vagina berbagai fase siklus estrus tertuang pada Tabel 1 dan Gambar 8.
Tabel 1. Gambaran mikroskopis hasil ulasan vagina pada berbagai fase siklus estrus.
FASE
HASIL ULASAN VAGINA
Nalbandov (1990)
Turner & Bagnara (1976) Smith & Mangkoewidjoyo (1988) Sartono (1994)
Proestrus Sel epitel berinti Sel epitel berinti Sel-sel kecil dengan inti bulat
Epitel berinti Berlendir
Estrus Sel berkornifikasi Sel-sel menanduk Epitel menanduk Inti piknotik
Epitel menanduk
Metestrus Leukosit diantara sel berkornifikasi
Leukosit banyak, Sel menanduk sedikit
Sel berkornifikasi Ada leukosit Epitel bertanduk berkurang Epitel berinti banyak
Diestrus Epitel berinti Leukosit
Leukosit bermigrasi Sel epitel dan
Leukosit
Leukosit banyak, epitel berinti
Gambar 8. Irisan melintang dinding vagina tikus putih pada
berbagai fase siklus estrus (Turner & Bagnara, 1976).
Leucosit Kornifikasi Estrus Metestrus Sel epitel berinti Diestrus Proestrus Leucosit
Sel epitel berinti
Kelenjar Susu
Peristiwa fisiologis siklus estrus tikus tercermin pada siklus pertumbuhan dan regresi kelenjar susu meskipun sedikit berbeda apabila terjadi kebuntingan (Knobil & Neills 2006). Pada fase proestrus terlihat saluran panjang di sekitar kelenjar susu, terutama dekat puting. Selanjutnya saluran makin membesar pada fase estrus. Sedangkan pada fase metestrus pelebaran saluran ini menjadi berkurang dan menghilang dan tampak kembali pada fase diestrus. Siklus kelenjar susu dirangsang oleh hormon estrogen dan progesteron (Ganong 2003). Hormon estrogen mempengaruhi proliferasi saluran kelenjar susu, sedangkan hormon progesteron merangsang perkembangan lobulus.
Kopulasi
Kopulasi biasanya terjadi 3 jam pertama fase estrus. Pada fase estrus ini estrogen mempengaruhi substrat metabolik vagina sehingga memproduksi asam yang mudah menguap dan menyebabkan timbulnya daya tarik seks tikus betina (Smith & Mangkoewidjojo 1988). Terjadinya kopulasi ditandai dengan sumbat vagina (vaginal plug) pada liang vagina (Rugs 1968). Sumbat vagina merupakan air mani yang menggumpal yang berasal dari sekret kelenjar prostat tikus jantan. Kondisi ini dapat diamati kira-kira 16-48 jam setelah kopulasi (Hafez 2000).
Fertilisasi
Proses fertilisasi terjadi pada bagian ampula oviduk. Sel telur yang dilepas dari folikel saat ovulasi dikelilingi oleh sel kumulus ooforus, korona radiata, dan zona pellucida. Sebelum spermatozoa melakukan penetrasi terjadi pelepasan kumulus ooforus sehingga memudahkan penetrasi spermatozoa (Syahrum et al. 1994). Zona pellucida merupakan lapisan ekstraseluler yang mengandung asam dan enzim akrosim yang berasal dari akrosom sperma akan membantu sperma dalam menembus zona pellucida. Setelah berhasil menembus zona pellucida spermatozoa akan memasuki sitoplasma ovum secara perlahan sedangkan bagian tengah dan ekornya akan tetap tinggal di luar dan akhirnya terlepas (Hafez 2000). Sejalan dengan itu implantasi terjadi setelah 4-5 hari sejak kopulasi (Syahrum et al. 1994 & Hafez 2000).
Pengendalian Hormon terhadap Siklus Reproduksi
Seperti halnya mamalia lain, kunci siklus reproduksi tikus betina terletak pada hipotalamus yang berhubungan dengan kelenjar hipofisis. Siklus reproduksi berlangsung dengan bantuan hormon gonadotropin yang dihasilkan oleh kelenjar hipofisis bagian anterior melalui sistem portal hipotalamus – hipofisis. Hormon gonadotropin terdiri atas FSH (Follicle Stimulating Hormone), LH (Luteinizing Hormone), Prolactin dan LTH (Luteotropic Hormone). Sintesis dan sekresi FSH dan LH dirangsang oleh Gonadotropic Releasing Hormon (Gn RH) yang disekresi oleh hipotalamus. Hormon ini mulai bekerja saat hewan mencapai masa pubertas (kematangan sel kelamin) (Johnsons & Everith 1988). FSH dan LH dibutuhkan untuk perkembangan normal folikel di ovarium. Perkembangan awal sel folikel dikendalikan oleh FSH yang selanjutnya merangsang sel granulose dan sel teka ovarium untuk mensekresi estrogen. Sedangkan progesteron terdapat dalan jumlah sedikit pada awal perkembangan sel folikel tetapi produksi progesteron mulai meningkat di bawah pengaruh LH (Partodihardjo 1980 & Lay Cock 1982).
Perkembangan akhir sel folikel dikendalikan oleh LH dan selanjutnya LH mendorong pecahnya folikel dan ovulasi (Nalbandov 1990). Sedangkan prolaktin bersifat luteolitik pada siklus estrus dan berfungsi untuk mempertahankan korpus luteum serta merangsang korpus luteum untuk menghasilkan hormon progesteron (Nalbandov 1990). Selain itu hormon prolaktin berfungsi untuk laktasi dan perkembangan ovarium selama kehamilan (Knobil and Neills 2006).
Nasib folikel telur dalam ovarium tergantung pada hormon gonadotropin. Banyak folikel yang mengalami atresia karena FSH dan LH hanya mampu menumbuhkan dan mematangkan folikel telur dalam jumlah tertentu. Akan tetapi folikel atresia dapat dicegah bila konsentrasi FSH dan LH di dalam sirkulasi darah cukup memadai. Hormon ini terikat dengan reseptor FSH dan LH pada sel folikel ovarium. Disamping itu hormon gonadotropin mempengaruhi perubahan folikel pre antral menjadi folikel antral (Johnsons & Everith 1988).
Fungsi endokrin ovarium adalah menghasilkan estrogen dan progesteron (Guyton 1996). Sumber estrogen selain sel teka interna folikel ovarium juga korpus luteum (Ganong 2003). Fungsi estrogen merangsang perkembangan jaringan
reproduksi, pertumbuhan saluran kelamin, menyebabkan pemanjangan dan hipertrofi kelenjar uterus, serta menyebabkan kontraksi uterus. Selain itu estrogen merangsang proliferasi dan diferensiasi epitel vagina, merangsang perkembangan saluran kelenjar susu, menimbulkan hasrat kawin dan merangsang peregangan tulang pinggul (Martin 1985 & Ganong 2003).
Progesteron dihasilkan oleh korpus luteum di bawah rangsangan LH. Fungsi progesteron antara lain mempersiapkan endometrium untuk implantasi. Sedangkan progesteron dan estrogen dalam jumlah yang cukup berfungsi mempertahankan implantasi zigot (Knobil & Neills 2006).
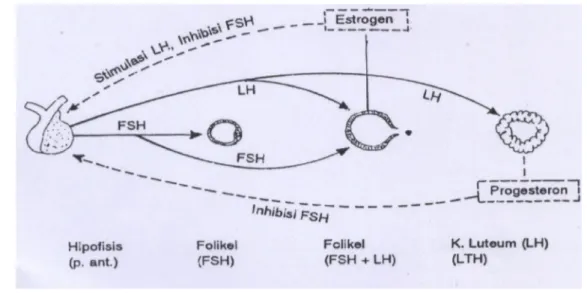
Pada permulaan fase folikuler, estrogen menghambat dan memberi umpan balik negatif terhadap sekresi FSH dan LH, tetapi pada akhir fase folikuler (sebelum ovulasi), peningkatan estrogen merangsang dan memberi umpan balik positif terhadap sekresi LH. Pada fase luteal jumlah estrogen dan progesteron yang tinggi menghambat dan memberi umpan balik negatif terhadap sekresi FSH dan LH. Adapun mekanisme umpan balik hormon pada poros hipothalamus-hipofisis-gonad tertera pada Gambar 9.
Gambar 9. Mekanisme umpan balik hormon pada poros
hipotalamus-hipofisis-gonad (Syahrum et al. 1994).
Adapun hubungan antara hipotalamus dan beberapa siklus reproduksi pada mamalia baik berupa siklus ovarium, siklus vagina, siklus endometrium dan hormon-hormon yang berperan dapat dilihat pada Gambar 10 berikut :
Gambar 10. Hubungan antara GnRH dan Hormon ovarium serta perubahan reproduksi pada mamalia betina poliestrus (Nelson 1983).
Peranan Hormon dalam Siklus Estrus
Pada akhir fase diestrus, korpus luteum yang mensekresi progesteron mengalami regresi. Regresi ini disebabkan pengaruh prostaglandin yang dihasilkan oleh uterus. Setelah produksi progesteron merendah, FSH-RH/LH-RH dilepas ke sistem porta hipofisa. Selanjutnya FSH-RH/LH-RH merangsang produksi dan pelepasan FSH yang disusul oleh produksi LH oleh hipofisa anterior. FSH merangsang folikel tertier pada ovarium untuk tumbuh menjadi folikel de Graaf. Lapisan sel teca interna dan sel granulosa pada folikel de graaf menghasilkan estrogen. Semakin masak dimensi folikel de Graaf semakin tinggi produksi produksi estrogen. Estrogen ini menyebabkan vaskularisasi alat kelamin. Setelah kadar estrogen dalam darah mencapai derajat ketinggian tertentu, maka terjadilah efek positif terhadap produksi dan pelepasan LH dari hipofisa anterior. Mekanisme ini disebut umpan balik (feed back) positif. Kadar LH dalam darah mendadak meningkat sehingga terjadi ovulasi. Ovulasi adalah peristiwa pecahnya dinding de Graaf dan keluarnya ovum (Nalbandov 1990).
Setelah ovulasi terjadi, kadar LH menurun dengan cepat, tetapi tidak kembali ke kadar dasar, melainkan cukup untuk merangsang sel teca interna untuk membentuk korpus luteum. Sejak terbentuknya korpus luteum, sel ini memproduksi hormon progesteron yang berfungsi meredakan aktivitas estrogen. Kecuali oleh LH, fungsi korpus luteum ditunjang oleh LTH. LTH berperan dalam merangsang korpus luteum untuk memproduksi progestin. Setelah folikel de Graaf pecah, produksi estrogen turun dengan cepat, hingga mencapai kadar dasar (kadar paling rendah dalam darah). Penurunan ini diikuti oleh kenaikan produksi FSH secara berangsur-angsur, FSH diperlukan oleh ovarium untuk merangsang pertumbuhan folikel. Folikel yang tumbuh secara berangsur-angsur mempertinggi kadar estrogen dalam darah. Setelah kadar estrogen dalam darah mencapai derajat ketinggian tertentu, maka terjadi rangsangan pada uterus untuk memproduksi prostaglandin. Prostaglandin ini menyebabkan korpus luteum beregresi dan progestin secara tajam menurun. Menurunnya progesteron dalam darah estrogen menjadi dominan pada alat reproduksi hingga terjadilah estrus (Partodihardjo 1980).
Mekanisme Antifertilitas terhadap Reproduksi Tikus
Suatu zat yang berkhasiat anti fertilitas dapat bekerja melalui beberapa mekanisme dan beberapa tempat kerja. Efek anti fertilitas suatu zat dapat terjadi di beberapa tempat atau pada tempat kerja yang sama tetapi dengan mekanisme berbeda. Sebagai contoh pada mamalia betina, aktivitas zat antifertilitas dapat terjadi di hipotalamus, hipofisis, ovarium, oviduk, uterus, dan vagina (Cheeke 1989).
Ada tiga macam istilah untuk aktivitas zat antifertilitas, yaitu zat yang bersifat sebagai kontrasepsi, abortivum dan intersepsi. Zat yang bersifat kontrasepsi berfungsi mencegah terjadinya ovulasi dan fertilisasi. Zat yang bersifat abortivum menyebabkan keluarnya fetus sebelum terjadinya implantasi sedangkan sebagai zat intersepsi, zat tersebut bekerja setelah terjadi fertilisasi yang akan mengganggu perjalanan menuju implantasi (Farnsworth et al. 1975 & Cheeke 1989).
Hewan betina yang tidak mengalami proses kebuntingan dapat disebabkan oleh beberapa faktor yaitu penghambatan proses ovulasi, fertilisasi dan implantasi (Suherman 2007). Salah satu penghambat proses ovulasi adalah penggunaan
obat-obatan atau zat kontrasepsi yang berupa fitosterol yang terkandung pada daun maja (β-sitosterol dan stigmasterol). Keadaan ini menyebabkan laju fertilitas dan tingkat kesuburan menjadi menurun.
Bahan aktif β-sitosterol dan stigmasterol tergolong steroid dan merupakan senyawa yang tidak larut dalam air, tetapi larut dalam pelarut organik seperti ether, etanol, kloroform, dan benzena (Harborne 1987). Steroid tumbuhan dapat bekerja sebagai kontrasepsi dengan menekan ovulasi pada sebagian besar siklus dan menghambat pelepasan gonadotropin (menghambat fungís ovarium) melalui mekanisme umpan balik (Nalbandov 1990). Selain itu steroid tumbuhan umumnya bersifat estrogenik sehingga akan mempengaruhi siklus menstruasi dan perkembangan folikel (Farnsworth et al. 1975). Steroid ini tentunya dapat turut meningkatkan kadar estrogenik dalam darah. Tingginya kadar estrogen dalam darah dapat menghambat hipofisis dalam mensekresi hormon FSH melalui umpan balik negatif (Ganong 2003, Johnsons & Everith 1988). Steroid tumbuhan memiliki kesamaan dengan steroid yang dihasilkan ovarium yaitu estrogen dan progesteron. Keduanya memiliki gugus utama berbentuk cincin siklopentana perhidrofenantrena (Tyler 1976 & Harborne 1987).
Berdasarkan sasaran aktivitas pada organ target, mekanisme zat antifertilitas dibedakan atas :
1. Hipotalamus-hipofisis
Mekanisme kerja steroid pada hipotalamus maupun hipofisis dapat menyebabkan aktivitas anti gonadotropin, artinya steroid tersebut akan secara langsung menghambat sekresi FSH dan LH yang dihasilkan oleh hipotalamus maupun hipofisis yang mengakibatkan terhambatnya produksi hormon estrogen dan progesteron. Estrogen dan progesteron diperlukan untuk memunculkan birahi normal pada hewan ovulator spontan, sehingga zat yang menekan gonadotropin mengakibatkan terhambatnya ovulasi dan menekan libido (Farnsworth et al. 1975). Selain itu juga akan menghambat induksi saraf ke hipotalamus yang mengatur sekresi GnRH pada hewan ovulator spontan yakni akan memberikan efek antifertilitas sebelum ovulasi (Farnsworth et al. 1975 & Hafez 2000).
2. Ovarium
Mekanisme kerja zat antifertilitas terhadap ovarium dapat ditunjukkan aktivitasnya pada penghambatan ovulasi dan steroidogenesis (Farnsworth et al. 1975 & Hafez 2000). Terhambatnya ovulasi ini sebagai akibat dari terhambatnya produksi estrogen dan progesteron seperti berkurangnya korpus luteum yang terbentuk berhubungan erat dengan jumlah ovum yang diovulasikan (Richards 1980). Selain itu juga akan menyebabkan penekanan jumlah dan perkembangan folikel yang mengakibatkan penurunan berat ovarium dan steroidogenesis (Zambrana 1971). 3. Oviduk
Mekanisme kerja zat antifertilitas dapat menyebabkan kekacauan fertilisasi dan kegagalan implantasi serta dapat menyebabkan sel telur yang sudah dibuahi akan mengalami regresi bila tiba terlalu dini di uterus (Farnsworth et al. 1975 & Hafez 2000).
4. Uterus
Mekanisme kerja zat antifertilitas pada uterus dapat bersifat interseptif maupun abortivum. Keseimbangan hormon estrogen dan progesteron pada tikus sangat diperlukan pada proses implantasi. Ada progesteron yang dilaporkan dapat menimbulkan gangguan keseimbangan proliferasi endometrium, sehingga mengganggu implantasi. Demikian juga antiestrogenik dapat menghambat implantasi pada hewan yang memerlukan estrogen pada proses implantasi (Farnsworth et al. 1975 & Hafez 2000). Mekanisme lain juga dapat berupa fetus yang mati atau resorbsi disebabkan karena terhambatnya aliran darah yang berasal dari placenta menuju fetus baik karena kekurangan oksigen maupun sari-sari makanan (Bronsons 1966). Selain itu resorbsi fetus dapat terjadi pada periode organogenesis yang terbukti dengan adanya bekas tapak implantasi pada uterus, maupun fetus yang melekat di daerah serviks akan diresorbsi sebagai benda asing oleh uterus (Rugs 1968).
5. Vagina
Mekanisme kerja zat antifertilitas di daerah vagina dapat bersifat spermisida (Farnsworth et al. 1975). Zat antifertilitas yang bersifat spermisida dapat mencegah terjadinya fertilisasi yang disebabkan tidak bergerak (imobilisasi) dan matinya spermatozoa (Syahrum et al. 1994).