Virus Influenza Tipe A
Penyakit Avian Influensa (AI) disebabkan oleh virus influensa tipe A yang merupakan virus RNA dari famili Orthomyxoviridae dengan genus Orthomyxovirus. Berbentuk spheris atau longitudinal, asam nukleatnya berantai tunggal dengan kutub negatif, sepanjang kurang lebih 13.588 nukleotida yang tersusun dalam 8 segmen gen yang mengkode sekitar 10 jenis protein, yakni HA (Hemaglutinin), NA (Neuraminidase), NP (Protein Nukleokapsid), M1 dan M2 (Protein matriks), NS1 dan NS2 (Protein nonstruktural), PA (Polimerase protein A), PB1 dan PB2 (Polimerase protein B1 dan B2) (Ghedin et al. 2005).
Daerah eksternal Hemaglutinin terdiri dari oligosakharida yang menyalurkan derivat neuroaminic, berbentuk seperti tombol dan berkaitan dengan kemampuannya melekat pada reseptor sel (Watowich et al. 1994). Neuraminidase berperan untuk menghidrolisis ikatan antara galaktosa dan N–acetylneraminic pada rantai ujung oligosakharida–glikoprotein. Fungsi NA ini harus berada dalam keseimbangan dengan HA. Hal ini agar aktivitas enzimatik dalam melepaskan asam sialat dari sel yang terinfeksi tidak menyebabkan penurunan efisiensi infeksi sel berikutnya. Fungsi lain dari NA adalah untuk melepaskan partikel virus yang sudah selesai replikasi dalam sel, mencegah virion yang sudah terbentuk tersebut menempel kembali pada reseptor asam sialat melalui tonjolan HA. Oleh karena itu efisiensi replikasi virus AI sangat tergantung pada kerjasama protein HA dan NA dari virus (Suzuki et al. 2000).
Polimerase RNA-RNA aktif, yang bertanggung jawab untuk replikasi dan transkripsi, dibentuk dari PB2, PB1, dan PA. Polimerase tersebut memiliki aktivitas endonuklease dan diikat ke RNP. Protein NS1 dan NS2 memiliki fungsi pengaturan untuk mendorong sintesis komponen-komponen virus dalam sel terinfeksi. (Gurtler 2006)
Protein matriks mempunyai peran dalam penyusunan virion virus AI. Bersama dengan protein HA dan NA protein M2 menyusun struktur amplop virus dan berperan sebagai saluran ion. Protein M1 tidak hanya sebagai komponen
struktural virus, tetapi juga berperan pada awal infeksi dalam pemisahan protein M1 dari RNP untuk masuk ke dalam sitoplasma sel tropisma (Reid et al. 2002).
Virus influensa memiliki 2 antigen utama yaitu Hemaglutinin (HA atau H) dan Neuraminidase (NA atau N) (Horimoto & Kawaoka 2005). Subtipe virus influensa tipe A dinamai berdasarkan kedua antigen utama tersebut (HA dan NA). Berdasarkan sifat antigenisitas dari glikoprotein, virus influensa A telah teridentifikasi ada 16 subtipe HA (H1-H16) dan 9 subtipe NA (N1-N9). (CDC 2008). Klasifikasi tersebut ditetapkan melalui analisis filogenetik terhadap nukleotida dan penetapan urutan (sequences) gen-gen HA dan NA melalui cara deduksi asam amino (Fouchier et al. 2005).
Virus AI juga bersifat dinamis dan dapat berkembang secara terus menerus. Variasi antigenik pada virus AI dapat terjadi melalui 2 cara yakni drift dan shift
Pada antigenic drift, prosesnya terjadi secara berlahan-lahan dengan menghasilkan sedikit perubahan pada struktur antigen permukaan HA dan atau NA, sehingga dihasilkan virus strain baru (CDC 2005b). Determinan antigenik dari glikoprotein permukaan HA dan NA dipengaruhi oleh mekanisme yang dipicu oleh kekebalan (Fergusson et al 2003). Mutasi yang menonjol di tempat antigenik menurunkan atau menghambat ikatan antibodi-antibodi penawar (neutralising antibody), dengan demikan memudahkan subtipe baru untuk menyebar dalam populasi yang tidak punya kekebalan.
Antigenic shift menunjukkan adanya perubahan mendadak dan mendalam dalam determinan antigenik, yaitu pertukaran subtipe HA dan atau NA, di dalam satu siklus tunggal replikasi. Hal ini terjadi dalam sebuah sel yang secara bersamaan terinfeksi oleh dua atau lebih virus influensa A dari subtipe yang berbeda. Karena distribusi segmen genomik virus yang sudah tereplikasi ke dalam progeni yang baru tumbuh berlangsung tanpa tergantung kepada subtipe asal dari tiap segmen itu, dapat muncul progeni yang berkemampuan untuk bereplikasi yang membawa informasi genetik dari virus induk yang berbeda-beda (WHO 2005b).
Antigenic shift dapat timbul melalui tranmisi secara langsung dari unggas ke manusia atau melalui percampuran antara gen virus influensa tipe A pada
manusia dan gen virus influensa tipe A pada hewan untuk membentuk virus subtipe A melalui sebuah proses yang disebut genetic reabsorbment (CDC 2005a).
Virus Influensa A mempunyai selubung yang mengandung glikoprotein dan dilapisi oleh lapisan lemak ganda. Virus ini tetap hidup dalam air pada suhu 200 C selama 4 hari, pada suhu 00 C dapat bertahan hidup lebih dari 30 hari dan masih tetap infektif dalam feses pada suhu 40 C selama 30-35 jam serta mampu hidup selama 7 hari pada suhu 200 C. Virus AI sensitif pada panas dan mati pada temperatur 560 C selama tiga jam atau pada temperatur 600 C selama 30 menit atau lebih. Virus tahan pada kondisi pH asam, kondisi lingkungan yang kering atau pada kondisi yang nonisotonik. Selubung virus dapat dirusak oleh deterjen dan infektivitasnya mampu dirusak dengan cepat oleh formalin, beta-propiolakton, agen yang bersifat oksidan, asam encer, eter, deoksikolat, hidroksilamin, Na-dedoksisulfat dan ion-ion ammonium serta senyawa iodium (Tabbu 2000).
Virus Influenza A Subtipe H5N1
Semua virus AI yang ada pada burung, virus H5N1 yang paling menjadi perhatian karena pengaruhnya terhadap kesehatan manusia dengan dua alasan; pertama, virus H5N1 sampai sejauh ini adalah penyebab paling banyak kasus kesakitan yang sangat parah dengan sejumlah kematian yang cukup besar. Virus ini telah menembus batas spesies untuk menginfeksi manusia; yang kedua adalah dampak selanjutnya terhadap kesehatan manusia, dimana jika virus H5N1 diberi kesempatan yang cukup maka akan mengembangkan karakter yang diperlukannya untuk menyebabkan pandemi influensa (WHO 2006).
Virus H5N1 adalah subtipe dari spesies virus influensa A. Strain virus HPAI A (H5N1) yang pertama kali diketahui telah membunuh dua flok ayam di peternakan Skotlandia tahun 1959, tetapi strain virus tersebut sangat berbeda dengan strain virus HPAI A yang ada saat ini. Studi AI H5N1 sejak tahun 1997 menunjukkan bahwa virus ini terus berkembang dengan mengalami perubahan pada antigenisitas dan susunan gen internalnya sehingga terjadi peningkatan patogenisitas dengan kemampuannya menginfeksi, menyebabkan terjadi perluasan jenis induk semang seperti pada spesies burung, tikus dan cerpelai serta bangsa
kucing dengan mengakibatkan infeksi sistemik dan virus ini juga dapat menjadi lebih stabil dilingkungannya (Perdue & Swayne 2005).
Penemuan lain menunjukkan virus H5N1 menginfeksi babi di Cina dan Vietnam, menginfeksi kucing (pada percobaan infeksi pada kucing rumah) di Netherland, diisolasi dari kucing rumah di Jerman, Thailand, Irak dan Rusia, menginfeksi anjing (isolasi virus H5N1 pada anjing mati di Thailand), isolasi virus H5N1 pada harimau dan leopard di kebun binatang Thailand. Selanjutnya virus H5N1 juga menginfeksi sejenis cerpelai di Jerman serta pada kucing liar di Vietnam. Strain virus H5N1 terus berkembang dan dapat beradaptasi, sehingga mamalia lain juga dapat terinfeksi (CDC 2008b).
Virus H5N1 telah bermutasi ke dalam berbagai strain dengan profil patogenik yang berbeda. Beberapa bersifat patogen terhadap spesies tertentu, tetapi yang lain tidak. Setiap variasi genetik spesifik yang diketahui dapat dilacak dengan isolat virus dari kasus infeksi tertentu. Strain virus HPAI A yang dominan pada tahun 2004, berkembang dari virus tahun 1999 sampai 2002 untuk menciptakan genotip Z (Perdue & Swayne 2005).
Genotip Z muncul tahun 2002 melalui pengelompokan dari genotip awal virus HPAI H5N1 (Li et al. 2004) yang menginfeksi burung di china tahun 1996, dan pertama menginfeksi manusia di Hongkong tahun 1997 (WHO 2006). Genotip Z endemik pada burung-burung di Asia Tenggara dimana telah membentuk sedikitnya dua clades yang dapat menginfeksi manusia dan telah menyebar di dunia pada populasi burung. Mutasi yang terjadi diantara genotip ini telah meningkatkan patogenitasnya (Chen et al 2004).
Isolat virus H5N1 yang diperoleh dari Hongkong tahun 1997 sampai 2001 menunjukkan bahwa virus ini tidak menyebabkan gejala penyakit yang nyata pada burung. Namun pada tahun 2002 isolat baru virus H5N1 yang muncul pada populasi burung di Hongkong, virus ini menyebabkan penyakit akut, termasuk disfungsi neurologis dan kematian pada itik (Sturm-Ramirez et al. 2004).
Penelitian terhadap virus H5N1 yang diisolasi dari manusia yang terinfeksi pada tahun 1997, menunjukkan bahwa mutasi genetik pada posisi 627 dari gen PB2 yang mengkode ekspresi polymesase basic protein (Glu627Lys) telah menghasilkan highly cleavable hemagglutininglycoprotein yang merupakan faktor
virulensi yang dapat meningkatkan aktivitas replikasi virus H5N1 dalam sel hospesnya (Hatta et al. 2001). Mutasi dari virus H5N1 kemungkinan besar dapat menghasilkan varian virus H5N1 baru yang dapat mengenali reseptor spesifik yang ada pada sel manusia (natural human α 2-6 glycan), sehingga bila ini terjadi maka penularan virus H5N1 dari manusia ke manusia dapat terjadi dengan mudah (Stevens et al. 2006).
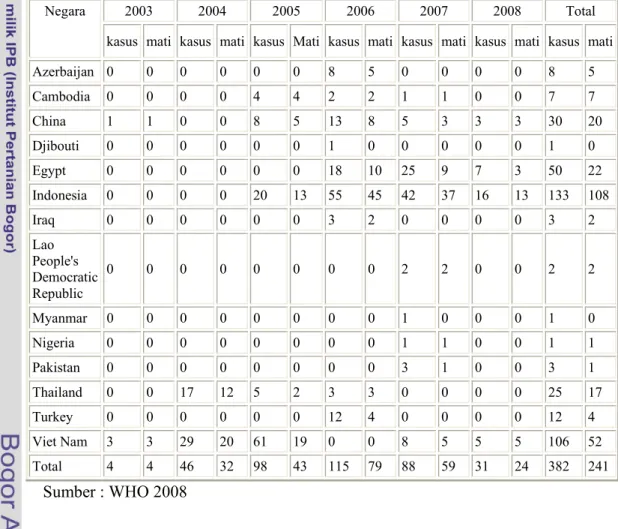
Tabel 1. Jumlah kumulatif kasus Avian Influensa A (H5N1) pada manusia yang dilaporkan dan dikonfirmasi ke WHO
Sumber : WHO 2008
Virus H5N1 memungkinkan untuk menyebabkan lebih dari satu pandemi influenza karena diduga terus bermutasi pada burung tanpa peduli apakah manusia mampu mengembangkan imunitas turunan terhadap strain-strain virus yang baru tersebut. Analisa genetik virus H5N1 menunjukkan bahwa keturunan genetiknya
Negara
2003 2004 2005 2006 2007 2008 Total kasus mati kasus mati kasus Mati kasus mati kasus mati kasus mati kasus mati
Azerbaijan 0 0 0 0 0 0 8 5 0 0 0 0 8 5 Cambodia 0 0 0 0 4 4 2 2 1 1 0 0 7 7 China 1 1 0 0 8 5 13 8 5 3 3 3 30 20 Djibouti 0 0 0 0 0 0 1 0 0 0 0 0 1 0 Egypt 0 0 0 0 0 0 18 10 25 9 7 3 50 22 Indonesia 0 0 0 0 20 13 55 45 42 37 16 13 133 108 Iraq 0 0 0 0 0 0 3 2 0 0 0 0 3 2 Lao People's Democratic Republic 0 0 0 0 0 0 0 0 2 2 0 0 2 2 Myanmar 0 0 0 0 0 0 0 0 1 0 0 0 1 0 Nigeria 0 0 0 0 0 0 0 0 1 1 0 0 1 1 Pakistan 0 0 0 0 0 0 0 0 3 1 0 0 3 1 Thailand 0 0 17 12 5 2 3 3 0 0 0 0 25 17 Turkey 0 0 0 0 0 0 12 4 0 0 0 0 12 4 Viet Nam 3 3 29 20 61 19 0 0 8 5 5 5 106 52 Total 4 4 46 32 98 43 115 79 88 59 31 24 382 241
dapat dengan mudah menjadi jauh lebih mematikan (Webster & Govorkova 2006).
Patogenesis
Virus AI terbagi atas dua grup berdasarkan pathogenitasnya yang terdiri atas virus influenza unggas berpatogenitas rendah (Low Pathogenic Avian Influenza, LPAI), menyebabkan penyakit yang ringan yang mungkin tidak terdapat atau tidak menunjukkan gejala pada beberapa spesies burung ; dan yang kedua adalah virus influensa unggas yang sangat patogen (Highly Pathogenic Avian Influenza, HPAI) menyebar dengan cepat dapat menyebabkan serangan penyakit yang serius dengan tingkat kematian yang tinggi (mencapai 90 sampai 100%) (Swayne & Suarez 2003).
Patogenesis virus AI dipengaruhi antara lain oleh tropisme (reaksi ke arah atau menjauhi stimulus), efektivitas replikasi dan mekanisme penghindaran imunitas. Selain itu faktor spesifik pada tiap spesies berperan juga terhadap hasil suatu infeksi, yang terjadi setelah penularan antar spesies (Swayne & Suarez 2000).
Patogenesis dan virulensi virus AI ditentukan oleh beberapa faktor yang saling berinteraksi (Behrens & Stoll 2006):
a. Faktor hospes :
• Keberadaan reseptor target pada sel hospes
• Ketersediaan enzim-enzim pada sel hospes yang penting untuk masuknya virus dan replikasi
• Keadaan kekuatan sistem imun pada hospes individu
• Imunitas spesifik terhadap epitope virus tertentu pada hospes individu dan populasi target
• Kemampuan sistem imun untuk mengendalikan replikasi virus yang efektif tanpa menimbulkan kerusakan kolateral yang serius karena respons inflamasi bagi hospes
b. Faktor virus
• Kemampuan untuk berikatan dengan sel hospes • Kemampuan pelepasan virus
• Restriksi efek sitopatogenik untuk memberikan keseimbangan yang cocok antara replikasi virus dan pengendalian oleh hospes
• Melepaskan diri dari pengawasan sistem imun dengan evolusi antigen yang bervariasi yang digerakkan oleh tekanan selektif respon imun. • Melepaskan diri dari pengawasan sistem imun lewat cara rekombinasi
dengan jenis virus berbeda dari penyakit zoonotik
• Modulasi respons imun untuk melemahkan mekanisme pertahanan hospes yang efektif.
Biasanya virus-virus H5 dan H7 bertahan stabil dalam bentuk yang berpatogenitas rendah dalam reservoir alaminya. Selanjutnya virus ditularkan melalui berbagai jalan ke peternakan unggas, mengalami masa sirkulasi yang bervariasi dan mengalami adaptasi dalam populasi unggas yang rentan seperti ayam dan kalkun. Selanjutnya spesies unggas tersebut menjadi sebab dari terjadinya beberapa siklus penularan. Virus influenza A subtipe H5 dan H7 dapat pula berubah melalui mutasi insersi menjadi bentuk yang sangat patogen (Rohm et al. 1995). Bentuk AI yang sangat patogen sampai saat ini secara eksklusif ditimbulkan oleh subtipe H5 dan H7, mampu menimbulkan penyakit sistemik yang ganas dan mematikan secara cepat, virus jenis HPAI tersebut dapat muncul secara tidak terduga dan sebagai tipe yang sama sekali baru dalam unggas yang terinfeksi (Perdue & Suarez 2000).
Infeksi virus H5N1 dimulai pada saat virus memasuki sel hospes setelah terjadi penempelan spikes virion virus influenza A, kemudian terjadi interaksi antara glikoprotein Hemaglutinin dengan reseptor yang mengandung sialic acid (SA) yang ada pada permukaan sel hospesnya. Setelah berhasil melekat pada reseptor yang sesuai, virion masuk dan menyatu ke dalam sebuah ruang endosom. Dalam ruang ini virus tersebut mengalami degradasi dengan cara menyatukan membran virus dengan membran endosom: dimediasi oleh pemindahan proton melalui terowongan protein dari matriks 2 virus, pada pH di endosom sekitar 5,0. Selanjutnya akan terjadi serangkaian penataan ulang protein matriks 1 dan komplek glikoprotein homotrimerik HA, terbukalah sebuah bidang yang sangat lipofilik dan fusogenik dari setiap monomer HA yang masuk ke dalam membran endolisomal, dimulailah terjadinya fusi antara membran virus dengan membran
lisomal (Wagner 2005). Berikutnya kedelapan segmen RNA genomik dari virus yang terbungkus dalam lapisan pelindung dari protein nukleokapsid dilepaskan ke dalam sitoplasma yang selanjutnya disalurkan ke nukleus untuk melakukan transkripsi mRNA virus dan replikasi RNA genomik melalui proses yang rumit yang diatur oleh faktor virus dan faktor sel (Whitaker et al. 1996).
Sesungguhnya ada perbedaan antara molekul reseptor yang ada pada manusia dengan reseptor yang ada pada unggas. Virus AI dapat mengenali dan terikat pada reseptor yang hanya terdapat pada jenis unggas yaitu oligosakharida yang mengandung N-acethylneuraminic acid α-2,3-galactose (SA α-2,3- Gal). Sedangkan reseptor yang ada pada permukaan sel manusia adalah SA α- 2,6-galactose (SA α-2,6-Gal), sehingga sebenarnya virus AI yang menyerang unggas tidak bisa menginfeksi manusia karena perbedaan reseptor spesifiknya. Tetapi bila terjadi perubahan pada 1 asam amino saja pada konfigurasi reseptornya maka reseptor yang ada pada manusia dapat dikenali oleh virus HPAI-H5N1 (Stevens et al. 2006).
Mutasi genetik virus AI seringkali terjadi sesuai dengan kondisi dan lingkungan replikasinya. Mutasi gen ini tidak saja untuk mempertahankan diri akan tetapi juga dapat meningkatkan sifat patogenisitasnya (Hatta et al. 2001).
Virus H5N1 diketahui telah mampu menembus barier spesies. Diawali pada kasus yang terjadi pada tahun 1997 di Hongkong yang menyebabkan 18 orang terinfeksi dan 6 diantaranya meninggal. Tindakan cepat dengan memusnahkan semua ayam di wilayah Hongkong berhasil menghentikan penyebaran virus tersebut secara sementara. Rekombinasi genetik dapat terjadi melalui hewan perantara, seperti babi atau di dalam diri manusia sendiri ketika terinfeksi oleh keduanya pada saat bersamaan. Akibat proses rekombinasi tersebut bisa muncul subtipe baru yang telah berubah sifat antigeniknya sehingga mampu dengan mudah menginfeksi manusia (Sturm-Raminez et al. 2004).
Penularan dan Penyebaran Penyakit
Penularan atau tranmisi virus AI secara umum dapat terjadi melalui inhalasi, kontak langsung, atau pun kontak tidak langsung (Bridges et al. 2003).
Menurut Soejoedono dan Handharyani (2005), penyakit AI dapat ditularkan dari unggas ke unggas lain dan dari peternakan ke peternkan lainnya dengan cara : - kontak langsung dari unggas terinfeksi dengan hewan yang peka, - melalui lendir yang berasal dari hidung dan mata,
- melalui kotoran (feses) unggas yang terserang AI,
- melalui manusia lewat sepatu dan pakaian yang terkontaminasi dengan virus,
- melalui pakan, air, dan peralatan kandang yang terkontaminasi,
- melalui udara karena memiliki peran penting pada penularan dalam satu kandang tetapi memiliki peran terbatas dalam penularan antar kandang, - melalui unggas air, yang dapat berperan sebagai sumber (reservoir) virus
yang terdapat dalam saluran intestinal dan dilepaskan lewat kotorannya. Virus H5N1 mudah berpindahdiantara burung yang memfasilitasi potensi penyebaran global virus H5N1. Sementara virus ini bermutasi dan mengelompok kembali, menciptakan variasi yang menginfeksi spesies yang belum jelas diketahui sebagai pembawa virus (Shinya et al. 2006). Burung dapat menyebarkan virus untuk waktu yang lama sebelum ia mati, hal ini memungkinkan untuk meningkatkan kemampuannya dalam menularkan virus (Chen et al. 2004).
Sekali virus menginfeksi kawanan unggas, virus LPAI tidak harus mengalami suatu fase adaptasi pada spesies unggas tersebut sebelum dikeluarkan lagi dalam jumlah yang cukup besar untuk dapat menular secara horisontal ke unggas lain, baik dalam kawanan sendiri ataupun ke kawanan yang lain. Demikian pula sekali virus HPAI berkembang dari kawanan unggas yang terinfeksi virus LPAI, ia dapat menular dengan cara yang sama. Pasar unggas yang menjual unggas dalam jumlah besar dan unggas yang ditempatkan secara saling berdesakan, merupakan multiplikator penyebaran penularan (Bulaga et al. 2003).
Avian Influenza pada burung kadang kala juga memanifestasikan gejala penyakit yang ringan dan tidak mematikan, sehingga itik liar dapat membawa virus dalam jarak yang jauh, dan membuang virus tersebut melaui kotorannya. Unggas yang terinfeksi oleh influenza A atau virus H5N1 dapat mengeluarkan virus dengan jumlah besar dalam kotorannya.
Melalui perdagangan internasional unggas hidup, virus AI dapat menyebar dari satu negara ke negara lain. Jenis burung kelana, termasuk unggas air liar, unggas laut dan unggas darat, mereka dapat membawa virus dalam jarak jauh. Burung yang terinfeksi virus akan mengeluarkan virus ini melalui saliva (air liur), cairan hidung, dan kotoran. Penyakit ini juga dapat menular melalui udara yang tercemar virus H5N1 yang berasal dari kotoran atau sekreta burung/unggas yang menderita AI. Penularan AI melalui feses ke oral adalah cara penularan yang paling sering terjadi (CDC 2008a) Resiko Penularan dari burung liar ke unggas peliharaan terutama terjadi kalau unggas peliharaan tersebut dibiarkan bebas berkeliaran, menggunakan air yang juga digunakan oleh burung liar, atau makan dan minum dari sumber yang tercemar kotoran burung liar pembawa virus (Henzler et al. 2003).
Burung-burung peliharaan dapat terinfeksi dengan virus AI melalui kontak langsung dengan unggas air terinfeksi atau unggas terinfeksi lainnya melalui kontak dengan permukaan seperti tanah, peralatan atau kandang atau bahan-bahan (seperti air atau makanan yang terkontaminasi oleh virus). Manusia dan kendaraan dapat menjadi vektor dari penyebaran virus AI dari satu peternkan ke peternakan lainnya (CDC 2008a).
Kebanyakan sumber utama infeksi virus H5N1 pada manusia adalah melalui kontak langsung dengan burung sakit yang mati, termasuk pada kegiatan penyembelihan, penghilangan bulu, penjagalan dan proses pengolahan untuk konsumsi dari burung yang terinfeksi. Pada sedikit kasus, kontak dengan feses unggas disaat anak-anak bermain di area peternakan unggas merupakan salah satu penyebab infeksi. Berenang ditempat dimana burung mati dibuang atau airnya telah terkontaminasi feses burung atau unggas terinfeksi juga salah satu sumber dari penularan. Pada beberapa kasus infeksi, sumber penularan belum dapat diidentifikasi, mungkin melibatkan faktor lingkungan yang belum diketahui. Burung peliharaan seperti merpati, kotoran unggas yang masih mentah yang digunakan sebagai pupuk juga berperan (WHO 2006)
Virus AI dapat ditularkan terhadap manusia dengan 2 jalan. Pertama kontaminasi langsung dari lingkungan burung terinfeksi yang mengandung virus kepada manusia, cara lain adalah lewat perantara binatang babi. Penularan diduga
terjadi dari kotoran secara oral atau melalui saluran pernapasan. Kekhawatiran yang muncul di kalangan para ahli genetika adalah bila terjadi rekombinasi genetik (genetic reassortment) diantara virus Influenza burung dan virus influenza manusia, sehingga dapat menular antara manusia ke manusia.
Menurut Herman & Strock (2005). Ada dua kemungkinan yang dapat menghasilkan subtipe baru dari H5N1 yang dapat menular antara manusia ke manusia, yaitu :
1. virus dapat menginfeksi manusia dan mengalami mutasi sehingga virus tersebut dapat beradaptasi untuk mengenali linkage RNA pada manusia, atau virus burung tersebut mendapatkan gen dari virus influenza manusia sehingga dapat bereplikasi secara efektif di dalam sel manusia. Subtipe baru virus H5N1 ini bermutasi sedemikian rupa untuk membuat protein tertentu yang dapat mengenali reseptor yang ada pada manusia, untuk jalan masuknya ke dalam sel manusia.
2. Kedua jenis virus, baik virus avian maupun human influenza tersebut dapat secara bersamaan menginfeksi manusia, sehingga terjadi “mix” atau rekombinasi genetik, sehingga menghasilkan strain virus baru yang sangat virulen bagi manusia .
Dari hasil penelitian yang dilakukan dengan cara memberi makan binatang seperti kucing, macan, ataupun macan tutul dengan unggas yang terinfeksi dengan H5N1 terbukti bahwa binatang pemakan daging tersebut dapat mengalami kelainan paru berupa pneumonia, severe diffuse alveolar damage, dan menyebabkan kematian (Keawcharoen et al. 2004)
Virus AI Pada Unggas Air
Burung-burung air liar, terutama yang termasuk dalam golongan Anseriformis (itik dan angsa) dan Charadiformis (burung camar dan burung-burung pantai), adalah pembawa (carrier) seluruh varietas subtipe dari virus influensa A, dan menjadi sangat mungkin merupakan reservoir alami untuk semua jenis virus influensa (Krauss et al. 2004).
Beberapa jenis burung khususnya unggas air berperan sebagai reservoir alami untuk virus AI dengan membawa virus tersebut dalam intestinumnya dan
mengeluarkannya melalui fesesnya namun kebanyakan dari burung-burung liar tersebut tidak menunjukkan sakit sehingga mereka dapat menyebarkan virus AI pada burung-burung lainnya (CDC 2008a). Dalam banyak kejadian itik piara dan itik liar (wild waterfowl) sering kali ditemukan tahan terhadap virus tersebut sehingga justru menjadi pembawa (carrier) yang menyebarkan virus ke ayam atau menyebarkan virus AI melintasi benua (Chen et al. 2005).
Cara pemeliharaan unggas air di Indonesia dan dibeberapa negara Asia Tenggara umumnya serupa, yakni : (a) dibiarkan berkeliaran di halaman bercampur dengan unggas lain apabila jumlahnya tidak banyak, (b) dikandangkan sementara dan dibawa ke sawah ketika musim panen padi usai, umumnya dalam jumlah puluhan sampai ratusan, (c) terus-menerus dikandangkan dan diberi makanan khusus, terutama untuk jenis itik unggul dalam jumlah banyak. Tipe pemeliharaan (a) dan (b) yang mempunyai potensi tinggi untuk menyebarkan virus AI ke daerah yang lebih luas. Di Indonesia itik piara tipe (b) kadang-kadang dibawa dengan kendaraan roda empat ke daerah yang cukup jauh untuk mengais sisa-sisa panen padi di sawah. Apabila diantara itik tadi membawa virus H5N1, tinja itik dapat mencemari saluran air, petani yang memanfaatkan saluran air tersebut untuk keperluan sehari-hari menjadi kelompok orang yang rawan tertular virus AI. Untuk itik piara tipe (a) karena tidak dikandangkan, tinjanya dapat mencemari kolam air di sekitar rumah dan halaman (Anonim 2002).
Dilaporkan pertama kali virus H5N1 ditemukan pada angsa terjadi di Guangdong China tahun 1996, yang menyebar pada itik-itik di Provinsi Coastall China Selatan dan di pasar-pasar unggas hidup di Hongkong. Dari kasus tersebut selanjutnya dilaporkan sekitar 18 orang terinfeksi dan 6 diantaranya meninggal (CDC 1998). 22 Desember 2003, virus AI menyerang unggas di Korea Selatan. Kasus AI yang pertama di Korsel ini, ditemukan di peternakan itik dekat Kota Eumseong Korea Selatan
Di Jepang virus HPAI H5N1 diisolasi dari daging itik yang diimpor dari Provinsi Shandong China Utara (Mase et al. 2005).Tahun 2001 virus HPAI H5N1 diisolasi dari daging itik yang diimpor Korea dari China. Dari hasil penelitian mengenai karakteristik dan pathogenitas dari virus AI H5N1 yang diisolasi dari daging itik tersebut ternyata virus ini menyebabkan kematian pada ayam dan juga
pada tikus. Sedangkan percobaan yang dilakukan pada itik tidak ada kematian namun titer virus relatif tinggi ditemukan pada otot kerangka dan jaringan otak pada itik (Tumpey et al. 2002).
Berdasarkan penelitian, itik piara yang ditulari virus H5N1 hasil isolasi tahun 2004 terbukti mengeluarkan atau mengekspresikan lebih banyak virus, dan dalam tempo lebih lama lewat tinja dibandingkan dengan itik yang ditulari dengan isolat virus tahun 2003. Disebutkan jumlah virus AI yang dikeluarkan itik piara tersebut mendekati jumlah virus yang dikeluarkan oleh ayam yang sedang terserang parah atau hampir mati oleh AI. Namun itik yang ditulari secara buatan tersebut tidak menunjukkan gejala klinik (Anonim 2008b). Penemuain ini menunjukkan bahwa adanya jalur penyebaran penyakit kepada hewan lain dan juga ke manusia (CDC 2008a)
Pada akhir tahun 2002 dilaporkan adanya wabah yang disebabkan oleh virus HPAI H5N1 yang menyebabkan kematian diantara burung-burung migrasi liar dan unggas air yang menetap di suaka margasatwa Hongkong. Menyebabkan infeksi sistemik pada itik dengan titer virus tinggi dan pathologi pada berbagai macam organ khususnya pada otak, itik mengalami penyakit yang akut termasuk menderita disfungsi neurologis. Virus juga diisolasi dalam titer yang tinggi pada air minum dan juga dari peralatan yang berkontak (Sturm-Ramirez et al. 2004).
Laporan kejadian AI di Thailand pada tahun 2005, diawali dengan penggembalaan itik sekitar 3000-5000 itik di area pertanian, walaupun tidak ada kontak antara itik yang digembalakan dan ayam peliharaan, namun dua minggu setelah kedatangan itik-itik tersebut, ayam di area tersebut banyak yang mati dan manusia yang berkontak langsung dengan ayam yang mati, meninggal terinfeksi virus H5N1 (Songserm et al. 2006).
Vaksinasi AI
Vaksinasi dalam dunia kedokteran hewan ditujukan untuk mencapai : perlindungan terhadap timbulnya penyakit secara klinis, perlindungan terhadap serangan virus yang virulen, perlindungan terhadap ekskresi virus, pembedaan secara serologik antara hewan yang terinfeksi dari hewan yang divaksinasi
(dikenal sebagai differentiantion of infected from infected from vaccinated animals, atau prinsip DIVA) (Lee & Suarez 2005).
Beberapa manfaat yang dapat diperoleh dengan menjalankan program vaksinasi AI (Indartono & Slamet 2005) :
• Mencegah dan menghindari terjadinya kerugian ekonomi akibat penyakit tersebut, yakni kerugian akibat terjadinya kematian, penurunan produksi, penurunan berat badan dan keterlambatan replacement
• Menurunkan dan menekan jumlah virus AI yang dapat disebarkan oleh hewan terinfeksi AI
• Menurunkan virulensi virus AI yang patogen di lapangan, menjadi lebih rendah tingkat keganasannya
• Mencegah dan menghindari timbulnya varian-varian baru akibat mutasi yang dapat membahayakan kesehatan manusia
Untuk kepentingan praktikal, beberapa persyaratan harus dipenuhi pada vaksin AI (Lee & Suarez 2004):
• Karena berpotensi menimbulkan perubahan struktur genetis, juga pada kasus subtipe H5 dan H7, resiko mutasi spontan yang mengakibatkan peningkatan patogenik, vaksin tidak boleh diubah dengan replikasi kompeten virus influenza. Oleh karena itu penggunaan vaksin dengan virus hidup tidak dipakai lagi
• Perlindungan terhadap HPAI pada unggas terutama tergantung kepada antibodi yang spesifik untuk HA tertentu. Oleh karena itu virus untuk vaksin harus berasal dari subtipe HA yang sama dengan virus liar yang ada di sana. Kecocokan ideal antara vaksin dan badan virus yang disyaratkan bagi vaksin untuk manusia, tidak menjadi keharusan bagi vaksin unggas. Pembangkitan imunitas reaktif-silang homosubtipik pada unggas mungkin sudah menjadi perlindungan yang memadai karena pada saat ini jarang dijumpai adanya pembentukan antigen yang dipicu vaksin pada virus influenza unggas, akibat tidak adanya upaya vaksinasi yang meluas.
• Strategi penandaan (DIVA) harus digunakan (Suarez 2005) atau sebagai gantinya, digunakan unggas yang tidak divaksinasi sebagai penanda untuk monitoring
Resiko hewan yang divaksinasi untuk terkena infeksi virus virulen dan mengeksresikannya biasanya dapat diturunkan namun tidak sepenuhnya mencegah. Hal ini dapat menimbulkan masalah epidemiologik yang signifikant di daerah endemik yang sudah mendapat vaksinasi secara luas, unggas yang sudah divaksinasi yang terlihat sehat dapat juga terkena infeksi dan mengeluarkan virus liar dibalik perlindungan vaksin (Lee & Suarez 2004).
Prinsip dasar pemakaian vaksin AI adalah virus vaksin harus homolog dengan sub tipe H atau subtipe H dan N virus asal lapang. Menurut regulasi OIE, vaksin harus mempunyai komposisi genetik yang stabil, proses inaktivasi sempurna (uji laboratorik), bebas pencemaran agen infeksius lainnya, mengandung konsentrasi antigen yang tinggi, menggunakan adjuvant berkualitas tinggi dan mempunyai tingkat keamanan, potensi serta efektifitas yang tinggi (uji laboratorik dan uji lapang) (Suarez 2005)
Vaksinansi dengan strain virus homolog telah terbukti menurunkan angka kematian dan memperpendek viral shedding. Utamanya ditujukan pada gen H yang menyandi antigen yang menginduksi pembentukan antibodi netralisasi (Asmara 2006).
Imunohistokimia
Imunohistokimia (IHC) telah ada sejak tahun 1930-an tetapi pada tahun 1940 studi tentang IHC baru dilaporkan (Anonim 2008a). Nama IHC berasal dari kata immuno dalam hubungannya dengan antibodi yang digunakan dalam prosedur, dan histo yang berarti jaringan. IHC adalah proses memisahkan protein pada sel dalam suatu jaringan dengan menggunakan prinsip ikatan spesifik antibodi pada antigen pada jaringan biologik, IHC dilakukan untuk membantu mendiagnosa agen etiologi dari proses infeksi. Keberadaan reaksi warna (jika itu spesifik berdasarkan pemakaian kontrol menunjukkan adanya komponen agen infektius yang diuji (Ramos-Vara 2005).
Pada reaksi imunohistokimia ini sifatnya adalah spesifik karena bahan yang dideteksi akan direaksikan dengan antibodi spesifik yang dilabel dengan suatu enzym. Enzym yang digunakan untuk melabel antibodi tersebut dapat berupa enzym : peroksidase, alkali fosfatase dan b-galaktosidase. Untuk menandai
adanya suatu reaksi enzimatik maka digunakan suatu indikator warna (chromogen) (Sudiana 2005).
Antibodi yang digunakan untuk deteksi spesifik dapat berupa poliklonal atau monoklonal. Antibodi monoklonal umumnya dianggap dapat menunjukkan lebih terperinci. Antibodi monoklonal, adalah antibodi yang memiliki idiotipe dan isotipe yang sama. Idiotipe adalah bagian antibodi yang menentukan spesifisitasnya (antigen binding surface), sedangkan isotipe adalah bagian antibodi yang menentukan kelas-sub kelas dari antibodi maupun yang menentukan tipe-subtipe dari suatu antibodi. Dapat dikatakan bahwa antibodi monoklonal adalah suatu antibodi yang memiliki antigen binding surface yang sama, kelas-sub kelas maupun tipe-kelas-sub tipe yang sama (Sudiana 2005). Antibodi poliklonal dibuat dengan menginjeksi hewan dengan antigen peptida, dan setelah respon imun sekunder dirangsang selanjutnya diisolasi antibodinya dari keseluruhan serum. Jadi antibodi poliklonal adalah pencampuran dari berbagai antibodi yang mengenal sejumlah epitopes.
Imunohistokimia adalah metode alternatif yang sangat baik disamping spesifik dan sensitif, relatif cepat, tidak mahal dan sedikit menggunakan laboratorium. IHC telah menjadi metode baik dan terpercaya untuk diagnosa rutin dan aktifitas penelitian (Ramos-Vara et al. 1999).
Imunohistokimia telah menjadi tehnik yang sangat penting dan secara luas dipakai pada laboratorium penelitian medis dan juga diagnosa klinika. Banyak sekali metode IHC yg bisa digunakan untuk melokalisasi antigen. Pemilihan metode yang sesuai harus didasari parameter-parameter seperti tipe spesimen yg diselidiki, dan tingkat sensitifitas yang dibutuhkan (Anonim 2008a).
Terdapat dua metode dasar IHC untuk mengetahui adanya antigen pada jaringan, yaitu : pertama, metode langsung adalah metode dengan satu tahap pewarnaan. Melibatkan antibodi berlabel yang bereaksi dengan antigen dalam bagian jaringan. Tehnik ini hanya menggunakan satu macam antibodi, sehingga prosedurnya pendek dan cepat. Walaupun demikian metode ini tidak sensitif karena hanya sedikit signal yang ditampilkan ; kedua, metode tidak langsung melibatkan antibodi primer tidak berlabel ( lapisan 1) yang bereaksi dengan antigen jaringan, dan antibodi sekunder berlabel (lapisan 2) yang bereaksi dengan
antibodi primer. (Antibodi sekunder harus melawan IgG spesies hewan dimana antibodi primer berasal). Pada metode tidak langsung selain lebih sensitif juga relatif lebih sedikit jumlah antibodi sekunder yang diperlukan sebagai ukuran konjugasi yang perlu dihasilkan (Anonim 2007).