Bab II
Tinjauan Pustaka
II.1 α-Amilase
α-Amilase (1,4-α-D-glukan-glukanhidrolase, E.C. 3.2.1.1) adalah endoenzim yang mengkatalisis hidrolisis ikatan α-1,4-glikosidik pada amilosa atau amilopektin menghasilkan α-limit dekstrin, oligosakarida linier, dan sejumlah kecil maltosa serta glukosa (Gambar II.1). Sebagian besar α-amilase merupakan metaloenzim, yang memerlukan ion kalsium (Ca2+) untuk aktivitasnya, integritas struktural, dan stabilitas. Keluarga α-amilase bisa dibagi menjadi dua kelompok, yaitu enzim yang menghidrolisis pati dan mengubah pati, atau enzim transglikosilasi (Sivaramakrishnan, 2006).
Gambar II.1 Proses hidrolisis ikatan α-1,4-glikosidik pada amilopektin
α-Amilase termasuk kedalam bagian kelas 13-glikosilhidrolase. Enzim-enzim ini bisa dibagi menjadi empat kelompok, yaitu endoamilase, eksoamilase, enzim debranching, dan transferase.
1. Endoamilase memotong ikatan internal α-1,4 menghasilkan produk α -anomerik.
2. Eksoamilase memotong ikatan α-1,4 atau α-1,6 pada residu glukosa bagian luar menghasilkan produk α- atau β-anomerik.
α-1,4-glikosidik
α-limit dekstrin
Oligosakarida linier
glukosa
transfer dari donor ke akseptor glikosidik membentuk ikatan glikosidik baru (Sivaramakrishnan, 2006).
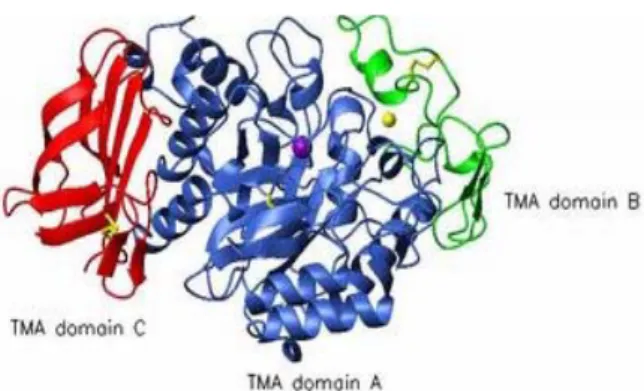
Gambar II.2 Struktur tiga dimensi α-amilase
α-Amilase merupakan enzim multidomain yang terdiri dari tiga domain utama, yaitu domain A, B, dan C (Gambarb. II.2). Domain A adalah N-terminal dari struktur TIM barrel, domain B terdiri dari loop panjang yang menonjol diantara β-strand 3 dan α-heliks 3 dan domain C dengan struktur β-sheet berhubungan dengan domain A. (β/α)8 Barrel terdiri dari delapan β-strand yang dikelilingi oleh delapan α-heliks (Gambar II.3). TIM barrel terdiri dari empat daerah lestari yang berhubungan dekat dengan sisi aktif, yang ada pada semua α-amilase, yaitu (i) ujung C-terminal β-strand 3 dan residu histidin yang berinteraksi dengan residu glukosa pada substrat, (ii) β-strand 4 dengan residu aspartat yang beraksi sebagai nukleofil selama katalisis, (iii) β-strand 5 dengan residu asam glutamat yang beraksi sebagai donor/akseptor proton, dan (iv) β-strand 7 dengan residu histidin dan residu aspartat yang bisa membentuk ikatan hidrogen dengan residu glukosa pada substrat (Sivaramakrishnan et al., 2006).
A B
Gambar II.3 Struktur (A) dan topologi (B) (β/α)8 atau TIM barrel. = α-heliks, = β-strand.
Konsep pengelompokan enzim α-amilase diusulkan pada tahun 1992. Berdasarkan definisi, anggota keluarga α-amilase mempunyai sifat: (i) harus beraksi pada rantai α-glukosidik dan menghidrolisisnya untuk menghasilkan monosakarida α-anomerik dan oligosakarida atau membentuk rantai α-glukosidik melalui transglikosilasi; (ii) mempunyai empat daerah lestari dalam struktur primernya yang terdiri dari sisi katalitik dan bagian pengikat substrat yang penting (Tabel II.1); (iii) mempunyai residu Asp, Glu, Asp sebagai sisi katalitik berdasarkan Asp206, Glu230, dan Asp297 pada Taka amilase A; dan (iv) mempunyai domain katalitik (β/α)8 atau TIM barrel (Gambar. II.3) (Sivaramakrishnan et al., 2006).
Tabel II.1. Empat daerah lestari dan berhubungan dengan β-sheets yang ditemukan dalam urutan asam amino enzim keluarga α-amilase.
Amilomaltase Amilosukrase CGTase CMDase BE Isoamilase M. amilase Pullulanase Sukrosa pase BLamilase I β2 II β4 III β5 IV β7
Bagian yang diberi warna adalah residu asam amino katalitik yang lestari (van der Maarel, et al., 2002).
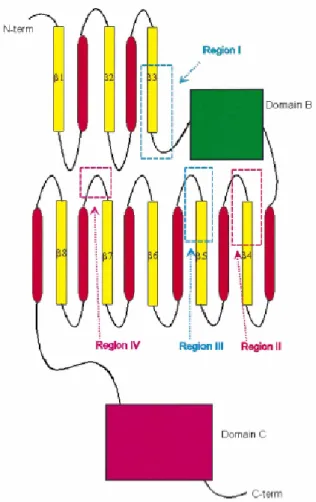
Gambar II.4 Diagram topologi α-amilase (Nielsen et al., 2000).
Dalam database CAZy, α-amilase dikelompokkan dengan jenis yang berbeda pada glikosil hidrolase dalam keluarga 13. Urutan asam amino α-amilase mengandung paling sedikit empat pola yang lestari (momor I-IV), yang ditemukan dalam TIM-barrel pada β-strands 3, 4 dan 5 dan dalam loop yang menghubungkan β-strand 7 dengan α-heliks 7 (Gambar II.4).
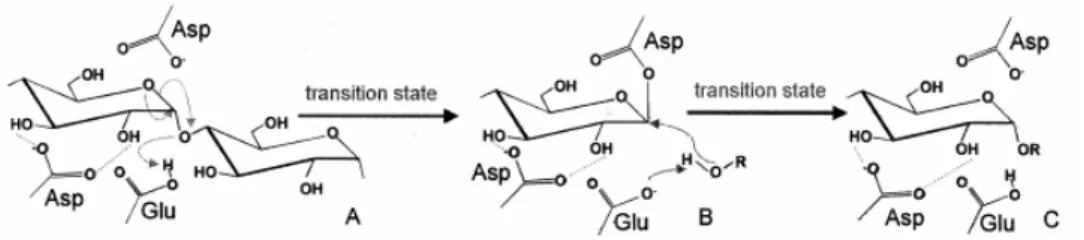
Pada umumnya mekanisme katalitik keluarga α-amilase adalah α-retaining double displacement. Mekanisme ini melibatkan dua residu katalitik dalam sisi aktif; yaitu asam glutamat sebagai katalis asam/basa dan aspartat sebagai nukleofil (Gambar II.5). Mekanisme ini melibatkan lima tahap: (i) setelah substrat terikat dalam sisi aktif, asam glutamat dalam bentuk asam mendonorkan sebuah proton pada oksigen ikatan glikosidik, yaitu oksigen diantara dua molekul glukosa pada
subsite – 1 dan + 1 dan nukleofil aspartat menyerang C1 pada glukosa pada subsite – 1; (ii) ion oksokarbonium pada keadaan transisi selanjutnya terbentuk melalui pembentukan intermediet kovalen; (iii) molekul glukosa diprotonasi pada subsite + 1 meninggalkan sisi aktif sementara molekul air atau molekul glukosa baru bergerak ke arah sisi aktif dan menyerang ikatan kovalen diantara molekul glukosa pada subsite – 1 dan aspartat; (iv) ion oksokarbonium pada keadaan transisi dibentuk kembali; (v) katalis basa glutamat menerima hidrogen dari air yang masuk atau molekul glukosa yang masuk pada subsite + 1, oksigen pada air yang masuk atau molekul glukosa yang masuk pada subsite + 1 menempati ikatan oksokarbonium di antara molekul glukosa pada subsite – 1 dan aspartat membentuk gugus hidroksil baru pada posisi C1 pada glukosa pada subsite – 1 (hidrolisis) atau ikatan glikosidik baru di antara glukosa pada subsite – 1 dan + 1 (transglikosilasi) (van der Maarel et al., 2002).
Gambar II.5 Mekanisme double displacement dan pembentukan intermediet kovalen dengan mempertahankan aksi glikosilhidrolase.
Sisi aktif enzim pada keluarga α-amilase bisa diumpamakan dengan sejumlah subsite, setiap subsite dapat berinteraksi dengan satu residu glukosa pada substrat (Gambar II.6). Masing-masing subsite terdiri dari rantai samping residu asam amino yang terletak pada loop dalam struktur enzim yang menghubungkan ujung C-terminal β-strands dengan ujung N-terminal heliks yang berdekatan pada domain katalitik (β/α)8-barrel.
Gambar II.6 Skema susunan subsite dengan penempatan oligosakarida pada subsite –5 sampai +3. Pemutusan terjadi antara subsite -1 dan +1, yang ditandai oleh tanda panah. Ujung pereduksi terikat pada subsite +3.
Dalam TAKA-amilase A, tiga residu asam, yaitu satu glutamat dan dua aspartat ditemukan pada pusat sisi aktif, mutasi yang dilakukan menunjukkan bahwa residu-residu ini sangat penting untuk katalisis. Residu asam glutamat merupakan proton donor, sementara salah satu dari dua asam aspartat yang ditunjukkan oleh urutan asam amino keluarga α-amilase bertindak sebagai nukleofil. Peran asam aspartat kedua belum ditentukan, tetapi diperkirakan terlibat dalam menstabilkan keadaan transisi ion oksokarbonium dan juga mempertahankan keadaan protonasi pada aktivitas asam glutamat (MacGregor et al., 2001).
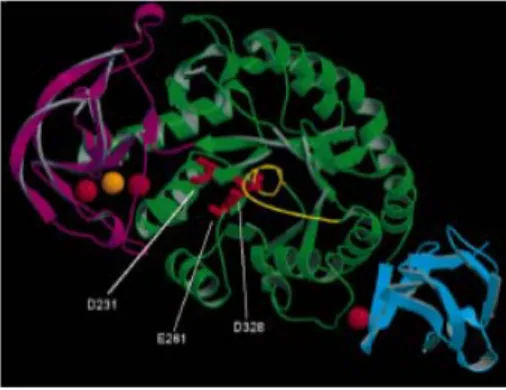
Gambar II.7 Susunan domain dan letak asam amino pada α-amilase. Domain A ditunjukan dengan warna hijau, domain B ditunjukan dengan warna magenta, dan domain C ditunjukan dengan warna hijau. Ion kalsium dan ion natrium ditunjukkan dengan bulatan berwarna merah dan oranye secara berturut-turut. Residu Asp231, Glu261 dan Asp328 pada sisi aktif ditunjukan dengan warna merah (Nielsen et al., 2000).
Laju hidrolisis pati oleh α-amilase bergantung pada beberapa kondisi seperti suhu, pH, substrat alami, konsentrasi substrat, konsentrasi enzim, adanya ion Ca2+ dan unsur-unsur penstabil lainnya (Sivaramakrishnan et al., 2006). Stabilitas α -amilase terutama dipengaruhi oleh suhu dan pH. Pada umumnya α--amilase stabil pada rentang pH antara 4 sampai 11 (Fogarty et al., 1979; Vihinen et al., 1989; Hamilton et al., 1999; Saito et al., 1973; Khoo et al., 1994 dalam Reddy et al., 2003) dan suhu optimum aktivitas α-amilase tergantung pada pertumbuhan mikroorganisme (Vihinen et al., 1989 dalam Reddy et al., 2003). Selain ion kalsium, ditemukan juga ion klorida yang berfungsi untuk meningkatkan efisiensi katalitik pada sisi aktif dengan meningkatkan pKa dari residu donor hidrogen pada sisi aktif (Nielsen, et al., 2001) dan ion seng yang berfungsi untuk meningkatkan aktivitas katalitik pada suhu tinggi dan stabilitas struktur (Linden, et al., 2003).
α-Amilase digunakan untuk menghidrolisis pati pada proses likuifaksi pati yang mengubah pati menjadi fruktosa dan sirup glukosa. Tahap pertama adalah perubahan pati menjadi maltodekstrin. Likuifaksi pati yang berlangsung secara bersamaan dengan pemasukan energi sangat diperlukan untuk menurunkan viskositas 40% pati terlarut. Sekarang ini, proses tersebut berlangsung pada pH 6, tetapi sangat diharapkan pemakaian pH yang lebih rendah untuk mengurangi pembentukan produk samping.
II.2 Pati
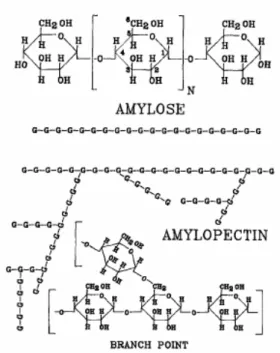
Pati merupakan sistem biopolimer yang terdiri dari dua polisakarida, yaitu amilosa (20-30%) yang mempunyai ikatan α1→4, dan amilopektin (70-80%) yang mempunyai ikatan α1→4 seperti pada amilosa juga mempunyai beberapa titik cabang α1→6 (Gambar II.8). Kedua polisakarida tersebut tersusun dari monomer glukosa.
Gambar II.8 Komponen pati jagung (G adalah unit anhidroglukosa) (Swanson et al., 1993).
Pati dibentuk melalui jalur biologi yang melibatkan fotosintesis. Pati dihasilkan sebagai granula yang disimpan pada plastid tanaman tingkat tinggi dan merupakan sumber energi utama pada tanaman.
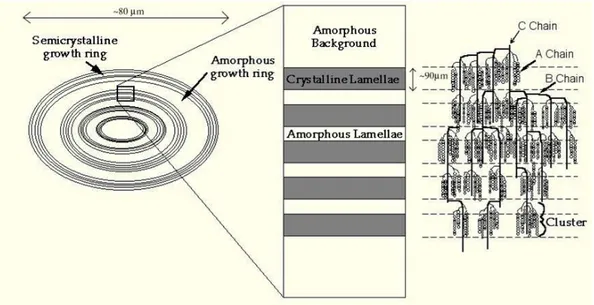
Pada pati dengan bentuk native-nya, amilosa dan amilopektin tersusun dari granula sebagai lapisan semi-kristalin dan amorf yang membentuk cincin (Gambar II.9). Lapisan semi-kristalin mengandung susunan daerah yang terdiri dari double heliks yang dibentuk melalui cabang amilopektin pendek, yang selanjutnya tersusun menjadi struktur kristal yang diketahui sebagai lamellae kristal. Bagian amorf pada lapisan semi-kristal dan lapisan amorf terdiri dari amilosa dan cabang amilopektin yang tidak tersusun.
Gambar II.9 Struktur granula pati dengan bagian amorf dan semi-kristalin yang membentuk cincin.
Dalam bakteri, amilopektin ekuivalen ditemukan dalam bentuk glikogen, yang mempunyai struktur yang sama dengan amilopektin. Perbedaan utama terletak dalam rantai samping: dalam glikogen, lebih pendek dan jumlahnya dua kali lebih besar. Sejumlah besar bakteri menggunakan enzim ekstraseluler atau intraseluler yang dapat mengkonversi pati atau glikogen dan dengan demikian menyediakan sumber energi dan karbon (van der Maarel et al., 2002).
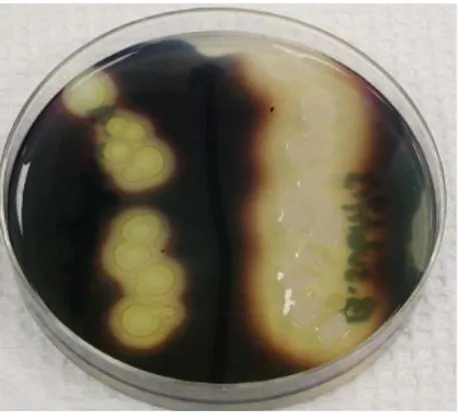
Pati pada media agar digunakan untuk menguji kemampuan suatu organisme dalam menghasilkan eksoenzim tertentu, termasuk α-amilase dan oligo-1,6-glukosidase, yang menghidrolisis pati. Molekul pati terlalu besar untuk masuk ke dalam sel bakteri, sehingga beberapa bakteri mensekresikan eksoenzim untuk mendegradasi pati menjadi subunit yang kemudian bisa digunakan oleh organisme.
Ketika tidak ada perubahan warna yang terjadi pada media ketika organisme menghidrolisis pati, kemudian ditambahkan iodin pada plate setelah diinkubasi. Iodin akan menjadi biru, ungu, atau hitam (tergantung pada konsentrasi iodin)
Gambar II.10 Iodin setelah ditambahkan pada media agar mengandung pati.
II.3 α-Amilase bakteri laut
Bakteri dan mikroorganisme lainnya banyak terdapat di lingkungan laut, yang secara taksonomi bermacam-macam jenisnya, secara biologi merupakan makhluk hidup yang aktif, dan semua habitat laut berkoloni, dari laut dalam sampai paling dangkal (Austin, 1988; Rheinheimer, 1992 dalam Kelman et al., 2006). Laut memiliki karakteristik lingkungan yang unik, yaitu lingkungan dengan kadar garam tinggi dan suhu rendah. Berdasarkan keunikan ini, bakteri laut mempunyai karakteristik yang berbeda, yaitu merupakan jenis halobakteri yang tahan terhadap kadar garam tinggi dan stabil terhadap penyimpanan pada suhu kamar.
Penelitian terhadap α-amilase dari bakteri laut telah banyak dilakukan, diantaranya penentuan struktur kristal α-amilase psikrofilik dari Alteromonas haloplanctis (Aghajari etal., 1998); kloning, sekuensing, dan ekspresi gen pengkode α-amilase ekstraseluler dari Pyrococcus furiosus (Dong et al., 1997); penentuan struktur α-amilase baru AmyC dari Thermotoga maritima (Dickmanns et al., 2006); dan pemurnian dan karakterisasi α-amilase ekstraseluler dari Vibrio sp. (Najafi et al., 2005).
Struktur kristal α-amilase dari Alteromonas haloplanctis (AHA) dalam keadaan nativenya pada resolusi 2,0 Å sangat baik membentuk kompleks dengan Tris pada resolusi 1,85 Å. Sisi aktifnya dikarakterisasi oleh residu asam, yaitu Asp174, Glu200, dan Asp264. Struktur kristal AmyC dari Thermotoga maritima ditentukan pada resolusi 2,2 Å. AmyC menunjukkan fitur yang mengkarakterisasi
α-amilase, seperti distorsi struktur TIM-barrel yang dibentuk oleh tujuh β-strand dan α-heliks (domain A), dan dua tambahan domain. Domain B mengandung tujuh heliks yang diinsersikan pada TIM-barrel setelah β-sheet 2, dan domain C, bagian lima-heliks pada C-terminal. Struktur α-amilase dari AmyC ini memiliki beberapa perbedaan dengan struktur α-amilase pada umumnya, yang mempunyai struktur TIM-barrel yang tersusun dari delapan α-heliks dan delapan β-sheet, serta domain B yang terbentuk dari loop yang panjang.
α-Amilase ekstraseluler dari Pyrococcus furiosus dikode oleh 460 residu rantai polipeptida tunggal, yang terdiri dari 26 residu sinyal peptida. Protein rekombinan ini merupakan homodimer dengan massa molekul 100 000 yang diperkirakan melalui filtrasi gel, mempunyai suhu optimum 100°C dan pH 5,5-6,0 dan tidak memerlukan Ca2+ untuk aktivitas atau termostabilitasnya. α-Amilase ekstraseluler dari Vibrio sp. mempunyai massa molekul 52, 480 kDa, aktivitas maksimum pada suhu 55-60°C dan pH 6,5. Berbeda dengan α-amilase ekstraseluler dari Pyrococcus furiosus, α-amilase dari Vibrio sp. memerlukan ion logam, seperti Ca2+ untuk aktivitas enzimnya.
II. 4 Bakteri genus Vibrio
Bakteri dengan genus Vibrio termasuk kedalam kerajaan Bakteria, divisi Proteobacteria, kelas Gamma Proteobacteria, ordo Vibrionales, famili Vibrionaceae. Keluarga Vibrionaceae termasuk beberapa spesies yang menyebabkan infeksi sistem pencernaan pada manusia dan hewan. Beberapa spesies Vibrionaceae secara luas terdistribusi pada lingkungan, yang berperan dalam siklus senyawa organik dan anorganik. Spesies Vibrionaceae juga telah
Organisme dengan bentuk kurva seringkali diklasifikasikan sebagai “vibrios” atau “spesies Vibrio”, diantaranya V. succinogenes, V. fetus, dan V. psychroerythrus Beberapa spesies Vibrionaceae mempunyai flagela polar ketika tumbuh dalam media cair, tetapi mensintesis flagela peritrichous ketika tumbuh pada media padat (Farmer, 2006). Anggota keluarga Vibrionaceae diketahui menghasilkan beberapa ekstraseluler protein.
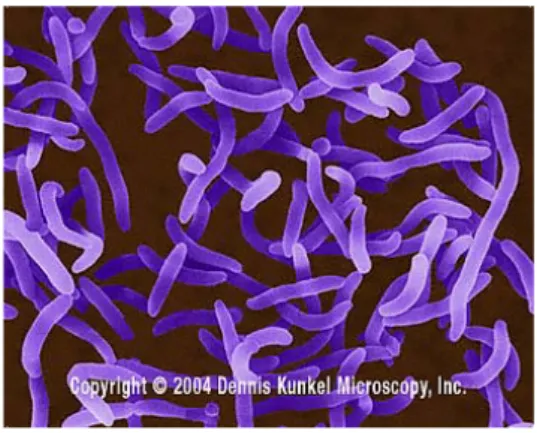
Bakteri dengan genus Vibrio bersifat gram negatif dan sebagian besar halofilik, mempunyai sel berbentuk batang yang melengkung berukuran antara 2-3 µm (Gambar II.11). Vibrio adalah fakultatif anaerobik yang memberikan uji positif untuk oksidase dan tidak membentuk spora. Semua genus Vibrio adalah motil dan mempunyai flagela polar pada ujung selnya untuk bergerak, tumbuh dengan baik pada media pepton.
Gambar II.11 Vibrio cholerae
Banyak spesies dari Vibrio yang sensitif terhadap pH asam, tapi tahan terhadap pH basa. Vibrio bisa mengalami metabolisme respiratori dan juga fermentatif dan merupakan organisme heterotropik yang memperoleh makanan dari hubungan mutualistik, parasitik, atau patogenik dengan organisme lain.
Beberapa spesies dari genus Vibrio telah banyak dipelajari, diantaranya adalah V. cholerae, V. alginolyticus, V. fischeri, V. harveyi, V. natriegens, V. parahaemolyticus, dan V. vulnificus.
Bakteri laut galur
Vibrio, Vibrio sp. galur 60 ditemukan bisa menghasilkan beberapa ekstraseluler protein termasuk protease, amilase, Dnase, dan hemaglutinin (Ichige et al., 1988).Salah satu spesies dari genus Vibrio yang telah diketahui mempunyai α-amilase adalah Vibrio sp. yang berasal dari laut India dengan massa molekular α-amilase sebesar 52,480 kDa, aktivitas maksimum pada suhu 55-60°C dan pH optimum 6,5 (Najafi et al., 2005).
II.5 Kloning Gen
Kloning gen merupakan cara untuk memperbanyak gen yang diinginkan. Hal ini dapat dilakukan dengan beberapa pendekatan, diantaranya dengan pembuatan perpustakaan genom cDNA dan amplifikasi gen dengan proses PCR.
Untuk mengklon fragmen DNA pada sel inang diperlukan suatu vektor. Ada beberapa jenis vektor (Tabel II.2) yang masing-masing berfungsi sebagai kromosom yang dapat menerima fragmen DNA sisipan dan mereplikasi secara bebas pada sel inang.
dibawa
Plasmid DNA sirkular
double-stranded E. coli Sampai 15 kb
Perpustakaan cDNA, subkloning Lambda
Bakteriofage Virus (DNA linier) E. coli Sampai 25 kb
Perpustakaan genom dan cDNA
Kosmid DNA sirkular
double-stranded E. coli 30-45 kb Perpustakaan genom Bakteriofage P1 Virus (DNA sirkular) E. coli 70-90 kb Perpustakaan genom BAC Bacterial Artificial
Chromosome E. coli 100-500 kb
Perpustakaan genom YAC Yeast Artificial
Chromosome Ragi
250-2000 kb (2 megabasa)
Perpustakaan genom
Vektor-vektor yang dapat digunakan berbeda antara satu dengan yang lainnya dalam sifat-sifat biologinya, kapasitas dalam membawa insert, dan jenis inang yang bisa membawanya. Vektor yang paling sederhana adalah DNA double-stranded dengan bentuk lingkaran kecil yang diketahui sebagai plasmid yang bisa mereplikasi dalam sitoplasma beberapa jenis sel bakteri, secara bebas pada kromosom bakteri. Plasmid yang banyak digunakan mempunyai beberapa sisi pengenalan, salah satunya adalah beberapa enzim restriksi yang berbeda; contohnya sisi pengenalan EcoRI dan sisi pengenalan HpaI. Hal ini memberikan fleksibilitas dalam memilih enzim-enzim yang bisa digunakan untuk memotong DNA yang mengandung fragmen, atau fragmen-fragmen tertentu yang diinginkan.
Plasmid yang membawa fragmen sisipan diketahui sebagai plasmid rekombinan. Setiap vektor plasmid juga membawa origin of replication dan gen yang resisten pada antibiotik tertentu. Origin of replication memungkinkan plasmid untuk mereplikasi secara bebas dalam bakteri. Gene yang resisten pada antibiotik tertentu dapat digunakan untuk menyeleksi sel bakteri yang mengandung plasmid.
Vektor plasmid dengan ukuran 2-4 kb dapat membawa DNA sisipan sampai 15 kb. Untuk fragmen DNA sisipan dengan ukuran yang lebih besar dapat digunakan beberapa vektor lain, seperti lambda bakteriofage (λ). λ fage adalah virus DNA
double-stranded yang menginfeksi E. coli. Kromosom λ mempunyai panjang 48,5 kb yang bisa menerima fragmen sisipan dengan panjang sampai 25 kb, yang menempati urutan viral nonesensial. Urutan viral yang berada pada kedua sisi insert melambangkan “tangan” vektor yang mengandung gen-gen yang diperlukan untuk membentuk keseluruhan partikel virus; partikel-partikel ini menginfeksi sel inang dengan efisiensi laju yang sangat tinggi dan selanjutnya bertambah secara terus-menerus. DNA yang disisipkan di antara dua “tangan” vektor λ akan masuk ke dalam partikel virus λ dan diamplifikasi didalam sel inang.
Vektor-vektor dengan kapasitas yang paling besar adalah artificial chromosomes : molekul DNA rekombinan yang dibentuk melalui kombinasi replikasi kromosomal dan segregasi elemen dengan DNA sisipan. Bacterial artificial chromosome (BAC) bisa mengakomodasi DNA sisipan dengan ukuran 150 kb, sedangkan yeast artificial chromosome (YAC) bisa mengakomodasi DNA sisipan dengan ukuran 2000 kb. Tetapi terdapat kerugian jika menggunakan BAC dan YAC sebagai vektor karena membuat lebih banyak pekerjaan untuk megkarakterisasi fragmen sisipan dengan ukuran besar yang dibawanya. Peneliti harus beberapa kali melakukan subklon fragmen restriksi yang lebih kecil dari koloni BAC atau YAC agar dapat melihat lebih dekat bagian-bagian tertentu pada DNA genom.
Beberapa vektor yang umum digunakan seperti pUC19 biasanya dapat mendeteksi adanya fragmen DNA yang diklon, berdasarkan pada hilangnya fenotip tertentu. Vektor ini banyak digunakan untuk mendeteksi gen pengkode β-galaktosidase pada E. coli, yang dapat dengan mudah dideteksi melalui kemampuan enzim yang mengkodenya untuk menghidrolisis substrat yang memberikan warna, yaitu X-gal (5-bromo-4-kloro-3-indolil-β-d-galaktosida) menjadi produk yang berwarna biru.
Dalam beberapa penelitian, hanya informasi pada urutan gen pengkode yang merupakan penelitian menarik, dan memberikan keuntungan yang lebih dalam membatasi analisis terhadap ekson gen tanpa menentukan struktur intron. Urutan pengkode dengan jumlah yang sangat kecil pada DNA genom dalam eukariot
jaringan, atau organ. Karena DNA tersebut diperoleh dari transkrip RNA, maka urutan-urutan ini hanya membawa informasi mengenai ekson.
Untuk menghasilkan koloni-koloni DNA dari urutan mRNA, bergantung pada serangkaian reaksi in vitro yang menyerupai beberapa tahapan dalam siklus hidup virus, yaitu retrovirus. Setelah menginfeksi sel, retrovirus menggunakan reverse transcriptase untuk menyalin single-strand RNA menjadi strand DNA komplementer, atau cDNA. Reverse transcriptase ini bisa juga berfungsi sebagai DNA-dependent DNA polimerase yang membuat strand kedua DNA komplementer untuk strand cDNA yang pertama (ekuivalen dengan urutan templat RNA asli).
Keuntungan utama perpustakaan genom adalah bahwa koloni-koloni genom didalamnya menunjukkan semua bagian DNA yang sama dan menunjukkan apakah genom utuh terdapat pada bagian dalam setiap koloni. Keuntungan utama perpustakaan cDNA adalah bahwa koloni-koloni cDNA menunjukkan bagian genom yang mengandung ingormasi yang digunakan dalam membuat protein dalam jaringan tertentu, yang ditentukan dari mRNA gen-gen yang dilibatkan.
Jika urutan nukleotida gen yang diinginkan sudah diketahui, dapat dilakukan amplifikasi gen dengan proses PCR (Polymerase Chain Reaction). PCR merupakan teknik perbanyakan DNA secara in vitro, yang memungkinkan adanya amplifikasi antara dua bagian DNA yang diketahui, hanya di dalam tabung reaksi, tanpa perlu memasukkannya ke dalam sel inang. Setiap siklus pada proses PCR terdiri dari tiga tahap utama, yaitu denaturasi, annealing, dan polimerisasi.
Untuk memperoleh koloni yang membawa gen yang diinginkan dari perpustakaan genom, ada beberapa sistem penapisan yang dapat digunakan, diantaranya sistem penapisan berdasarkan komplementasi dan uji aktivitas. Sistem penapisan lain
yaitu dengan hibridisasi atau diklon langsung setelah dilakukan amplifikasi gen yang diinginkan dengan PCR. Kloning gen yang menggunakan sistem penapisan dengan hibridisasi