1 BAB 14
Siklus ATP dan Bioenergetika Sel
Bioenergetika adalah bagian dari biokimia yang bersangkutan dengan transformasi dan penggunaan energi oleh sel hidup
Hukum Pertama dan Kedua Termodinamika
- Hukum Pertama
“Di dalam setiap perubahan fisik atau kimia, jumlah total energi pada lingkungan adalah tetap”.
Sederhananya, energi itu tidak dapat diciptakan dan tidak dapat dimusnahkan, hanya dapat diubah dari satu bentuk ke bentuk yang lainnya dan jumlah energinya tidak berubah.
- Hukum Kedua
“Semua perubahan fisik atau kimia cenderung berjalan menuju arah, sedemikian rupa, sehingga energi yang bermanfaat terurai secara tidak dapat balik (ireversibel) menjadi bentuk arah yang tidak teratur, yang disebut entropi. Penguraian ini terhenti pada titik keseimbangan. Pada saat ini, entropi yang terbentuk berada pada tingkat yang paling maksimum yang dimungkinkan oleh kondisi lingkungan yang bersangkutan”.
Terdapat dua jenis energi bermanfaat, yaitu energi bebas dan energi panas. Energi bebas adalah jenis energi yang melakukan kerja hanya pada suhu atau tekanan yang tetap. Energi panas adalah energi yang melakukan kerja hanya melalui suatu perubahan suhu tatau tekanan. Entropi adalah energi dalam keadaan acak atau tidak teratur (energi tidak berguna).
Aspek lain dari hukum termodinamika kedua ini adalah sistem dan lingkungan. Sistem adalah kumpulan benda-benda yang mengalami suatu proses kimia atau fisik tertentu. Sedangkan lingkungan adalah tempat saling bertukar
2 energi dengan sistem yang bereaksi. Sistem dan lingkungan bersama-sama menyusun suatu semesta(Gambar 14-1).
Semesta (sistem + lingkungan)
Perubahan dalam energi bebas, panas, dan entropi di dalam reaksi kimia pada suhu dan tekanan yang tetap, yang terdapat dalam sistem biologis, dihubungkan satu dengan yang lain secara kuantitatif oleh persamaan :
ΔG adalah perubahan energi bebas pada sistem yang sedang bereaksi.
ΔH adalah perubahan dalam kandungan panas sistem tersebut (entalpi). Jika sistem kehilangan atau membebaskan panas, ΔH bertanda negatif. Sedangkan jika sistem menyerap panas dari lingkungan, maka ΔH bertanda positif.
T adalah suhu absolut pada saat proses berlangsung.
ΔS adalah perubahan dalam entropi semesta, termasuk sistem yang sedang bereaksi.
Organisme hidup mempertahankan keteraturan internalnya dengan mengekstrak energi bebas dari makanan (atau sinar matahari) dari lingkungannya, dan mengenbalikan ke lingkungan jumlah energi yang sama, dalam bentuk yang kurang berguna, terutama dalam bentuk panas, yang menjadi terbaur (acak) ke tempat-tempat lain pada semesta. Organisme hidup secara terus menerus memberikan entropi kepada lingkungannya sebagai upah yang diperlukan untuk mempertahankan keteraturan internalnya sendiri.
Gambar 14-1
Pada keadaan standar, energi dapat dipertukarkan antara sistem dan lingkungannya, tetapi pertukaran energi harus mengikuti hukum-hukum termodinamika. Pertama, jumlah total energi didalam “semesta” yaitu sistem + lingkungannya harus tetap. Hukum kedua menyatakan bahwa jika suatu sistem mengalami bebrapa perubahan fisik atau kimia, entropi sistem harus meningkat, secara serentak, energi bebas sistem yang sedang bereaksi menurun. Bersamaan dengan perubahan itu, panas dapat mengalir dari sistem ke lingkunganya atau dari lingkungan ke sistem untuk memenuhi persamaan ΔG = ΔH – T ΔS
Lingkungan
Lingkungan sistem
3 Sel aerobik melakukan oksidasi glukosa (C6H12O6) menjadi CO2 dan H2O pada suhu dan tekanan tetap
C6H12O6 + 6O2 → 6CO2 + 6H2O
Keadaan dianggap baku jika suhu setempat 25°C atau 298°K dan tekanan 1 atm (760 mmHg). Dalam perhitungan termodinamika perubahan energi berikut ini terjadi pada oksidasi glukosa per mol.
ΔG = -686.000 kal/mol
ΔH = -673.000 kal/mol
ΔS = Δ Δ
= = + 44 kal/mol
Peningkatan di dalam ketidakteraturan atau entropi molekular, menyebabkan oksidasi glukosa.
Sel Membutuhkan Energi Bebas
Bentuk energi yang dapat dan harus digunakan oleh sel adalah energi bebas, yang dapat melakukan kerja pada suhu dan tekanan yang tetap. Sel heterotrof memperoleh energi bebas dari molekul nutrien kaya energi, dan sel fotosintetik memperoleh energi bebas dari radiasi solar yang diserap. Kedua jenis sel mengubah input energi bebas ini menjadi bentuk umum energi kimia, dan menggunakannya untuk menjalankan aktifitas sel melalui proses yang tidak melibatkan perbedaan suhu dan tekanan tetap.
Perubahan Energi Bebas Baku suatu Reaksi Kimia Dapat Dihitung
Setiap reaksi memiliki perubahan energi bebas baku (ΔG°). Di dalam reaksi yang mengandung lebih dari satu molekul pereaksi atau produk, seperti reaksi:
aA + bB cC + dD
(molekul yang bereaksi telah membebaskan panas)
(entropi semesta telah meningkat) (energi bebas sistem, yang merupakan molekul yang bereaksi telah menurun)
4 ΔG° = -2,303 RT log K’eq
a, b, c, dan d merupakan jumlah molekul A, B, C, dan D yang berpartisipasi. Tetapan kesetimbangan:
K’eq =
[ ]
Untuk menghitung perubahan energi bebas baku ΔG° menggunakan persamaan:
R = tetapan gas (1,987 kal/mol.K) T = suhu mutlak (298°K)
*K’eq> 1,0 , maka ΔG° = negatif (-) K’eq< 1,0 , maka ΔG° = positif (+)
ΔG°adalah perbedaan diantara kandungan enerdi-bebas pereaksi, dan energi bebas prosuk pada keadaan baku (298°K, 1 atm, pereaksi serta produk pada konsentrasi 1 M). ΔG° bertanda negatif jika produk mengandung energi bebas lebih sedikit dari pada pereaksi sehingga reaksi akan berjalan ke arah pembentukan produk pada keadaan baku. Jika ΔG° bertanda positif, berarti produk reaksi mengandung lebih banyak energi bebas dari pereaksi, sehingga reaksi akan berjalan ke arah kebalikannya.
ΔG° Memiliki Nilai Khas bagi Reaksi Kimia yang Berbeda
Jika perubahan energi bebas baku (ΔG°’) mempunyai tanda negatif, berarti reaksi berlangsung dengan hilangnya energi bebas. Jika perubahan energi bebas baku (ΔG°’) mempunyai tanda positif, berarti reaksi berlangsung dengan menyerap energi bebas.
Hidrolisis senyawa sederhana ester, amida, peptida, dan glikosida, serta penyusunan dan eliminasi berjalan dengan perubahan energi bebas baku yang relatif kecil, sedangkan hidrolisis anhidrida asam terjadi dengan penurunan energi baku yang relatif besar. Oksidasi senyawa organik menjadi CO2 dan H2O berjalan dengan penurunan energi bebas baku yang besar. Namun, perubahan energi bebas
5 baku tidak menyatakan berapa banyak energi bebas yang dapat dimanfaatkan secara biologi.
Terdapat Perbedaan Penting Diantara ΔGo’ dan ΔGo.
Perubahan energi bebas baku ΔG°’ menyatakan arah reaksi dan berapa jauh reaksi tertentu akan berjalan dalam mencapai keseimbangannya, jika reaksi ini terjadi pada kondisi baku (konsentrasi awal semua komponen 1,0 M; pH 7,0; dan suhu 25oC). ΔGo’merupakan suatu tetapan yang tidak dapat diubah.
Selanjutnya, ΔGo
setiap reaksi yang sedang menuju keseimbangan selalu beranda negatif dan menjadi lebih kecil (berkurang negatifnya) dengan melajunya reaksi, dan menjadi nol pada titik keseimbangan. Jadi, ΔGo
adalah perubahan energi bebas reaksi pada konsentrasi yang sesungguhnya, pada saat terjadi reaksi kimia tersebut.
ΔGo
dan ΔGo’ bagi setiap reaksi A + B → C + D dihubungkan oleh persamaan:
Dapat disimpulkan bahwa ΔGo
reaksi kimia yang sedang berlangsung bertanda negatif dan selalu meningkat menuju nilai nol, sedangkan ΔGo’ akan tetap konstan.
Nilai Energi Bebas Baku Reaksi-reaksi Kimia dapat Dijumlahkan
Sebagai contoh ada 2 buah reaksi kimia sebagai berikut: Glokusa 1-fosfat
Glukosa 6-fosfat
Fruktosa 6-fosfat
fosfoglukomutase
Glukosa fosfat isomerase
ΔGo’ = - 1,74 kkal/mol
6 Jumlah kedua reaksi:
Glukosa 1-fosfat fruktosa 6-fosfat Maka, energi bebas bakunya adalah:
∆G3 ’ = -1,74 + (+0,40) = -1,34 kkal/mol
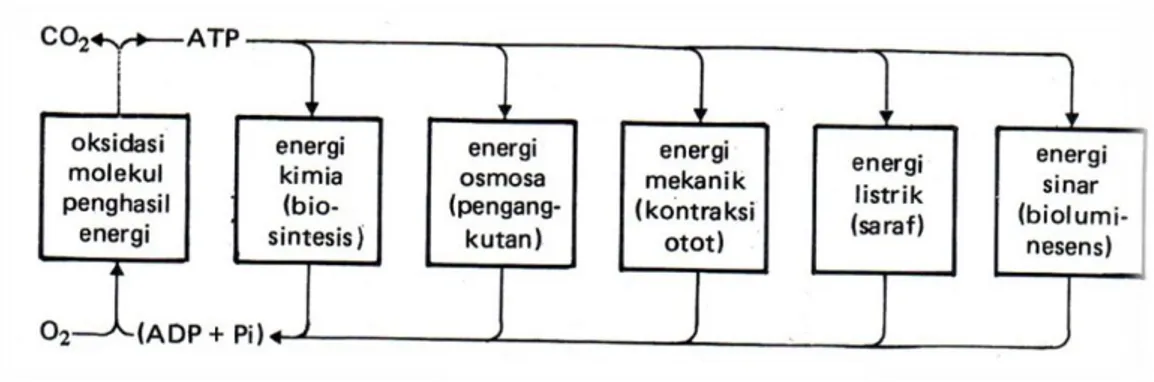
ATP Adalah Rantai Kimia Utama diantara Aktivitas Sel Penghasil Energi dan Aktivitas Sel yang Membutuhkan Energi
Prinsip dasar perubahan energi dalam sistem kimia dapat diamati dari siklus energi dalam sel. Sel heterotrof memperoleh energi bebas dari degradasi (katabolisme) molekul nutrien dan menggunakan energi ini untuk:
1. Melakukan sintesis biomolekul dari molekul pemula yang lebih kecil, 2. Menjalankan kerja mekanik,
3. Mengangkut biomolekul atau ion melalui membran menuju daerah berkonsentrasi lebih tinggi.
ATP berperan dalam katabolisme sebagai pembawa energi kimia dan memberikan sejumlah energinya kepada proses-proses yang memerlukan energi dengan memecah dirinya menjadi ADP dan fosfat.
Gambar Siklus ATP di dalam sel Kimiawi ATP Telah Dikenal Baik
ATP, ADP (adenosin difosfat) dan AMP (adenosin monofosfat) merupakan nukleotida. Di dalam ATP, ADP, dan AMP, basa yang terkandung adalah purin, adenin, dan gula 5-karbonnya adalah D-ribosa. Nukleotida melakukan berbagai fungsi sel, terutama sebagai unit pembangun DNA dan RNA, yakni berperan menjadi unsur penyandi. Di dalam sel normal yang melakukan
7 respirasi, ATP menyusun sampai 80% atau lebih dari jumlah ketiga adenin ribonukleotida.
Pada pH 7,0, ATP dan ADP terdapat sebagai anion ATP4- dan ADP3- yang bermuatan banyak karena hampir semua kandungan fosfat mengion sempurna pada pH ini. Namun dalam cairan intra sel yang mengandung Mg2+ berkonsentrasi tinggi, ATP dan ADP terdapat sebagai senyawa kompleks Mg ATP2- dan Mg ADP-. Dalam banyak reaksi enzimatik yang melibatkan ATP sebagai donor fosfat, bentuk aktifnya adalah MgATP2-. Di dalam sel, konsentrasi ATP biasanya relatif konstan dalam keadaan imbang, kecepatan pembentukan ATP diimbangi oleh kecepatan degradasinya. Jadi, gugus fosfat ujung pada pembentukan ATP mengalami penguraian dan penggantian secara terus-menerus dari pool fosfat anorganik selama metabolisme sel.
ATP Memiliki Energi Bebas Hidrolisis Baku yang Khas
ATP mengalami kehilangan gugus terminal fosfatnya jika terhidrolisis, dengan membentuk ADP dan fosfat anorganik
ATP + H2O ADP + Pi ∆G ’ = -7,3 kkal/mol
ATP disebut senyawa fosfat berenergi tinggi dan glukosa 6-fosfat sebagai senyawa fosfat berenergi rendah. ATP termasuk senyawa fosfat berenergi tinggi karena ketika ∆G ’ hidrolisis ATP mencapai -7,3 kkal/mol, sama dengan ∆G ’hidrolisis ADP menjadi AMP yang menandakan kedua gugus fosfat ujung ATP adalah gugus berenergi tinggi. Sedangkan ketika AMP dihidrolisis menghasilkan adenosin dan fosfat, ∆G ’nya hanya mencapai -3,4 kkal/mol saja sehingga pemecahan molekul ini termasuk ke dalam kelas berenergi rendah.
Mengapa ATP Memiliki Energi Bebas Hidrolisis Baku yang Relatif Tinggi? Terdapat tiga struktur yang menentukan energi bebas baku hidrolisis ATP yaitu:
1. Derajat ionisasi ATP dan produk hidrolisisnya.
2. Molekul ATP memiliki empat muatan negatif yang letaknya berdekatan, yang saling tolak-menolak ketika pH = 7,0.
8 3. Kedua produk hidrolisis ATP yaitu ADP3 -dan HPO42- merupakan hibrid resonansi, yaitu suatu bentuk stabil yang khusus, dengan elektron tertentu dalam konfigurasi yang memiliki jauh lebih sedikit energi dibandingkan dengan kedudukan aslinya di dalam molekul ATP.
Ikatan fosfat berenergi tinggi yang dilambangkan dengan ―~‖ menyatakan
bahwa ikatannya mengandung energi. Sebenarnya pemecahan ikatan justru membutuhkan input energi. Energi bebas yang ada pada hidrolisis suatu molekul organik bukan berasal dari penguraian ikatan khusus, tetapi lebih diakibatkan pada produk reaksi memiliki kandungan energi bebas yang lebih sedikit dibandingkan dengan pereaksi.
9 ATP Berperan Sebagai Senyawa Antara yang Berlaku Umum di dalam Reaksi Pemindahan Posfat
Reaksi-reaksi metabolisme berjalan melalui serangkaian reaksi enzimatis berurutan, yang dihubungkan oleh senyawa antara yang bersifat umum, di mana produk dari suatu reaksi menjadi substrat reaksi berikutnya.
A + B → C + D D + E → F + G
Reaksi-reaksi di atas dihubungkan oleh senyawa antara D. Agar energi kimia dapat berpindah dari satu reaksi kimia ke reaksi kimia yang lain, maka kedua reaksi harus memiliki senyawa antara. Jadi, komponen D pada kedua reaksi tersebut dapat berfungsi sebagai cara pemindahan energi dari reaksi pertama ke reaksi kedua.
Selama berlangsungnya reaksi katabolik yang memberikan energi, dihasilkan senyawa fosfat berenergi tinggi dengan menggunakan energi yang dibebaskan pada degradasi nutrien sel. Enzim kinase mengkatalisis pemindahan senyawa fosfat dari senyawa fosfat berenergi tinggi tersebut (X — P) kepada ADP membentuk ATP. Pada tahap kedua, kinase khusus lainnya mengkatalisa pemindahan senyawa fosfat terminal dari ATP ke suatu molekul penerima.
X — P + ADP → X + ATP ATP + Y → Y — P
Adanya ATP sebagai senyawa antara mengakibatkan pindahnya energi kimia dari X — P menuju Y melalui pemindahan gugus fosfat. ATP hampir selalu menjadi perantara bagi reaksi pemindahan gugus fosfat tersebut, karena sel tidak memiliki kinase yang dapat memindahkan senyawa fosfat secara langsung dari senyawa fosfat dengan energi super tinggi kepada senyawa penerima berenergi rendah.
10 Dua Senyawa Fosfat Berenergi Super-Tinggi Dihasilkan pada Pemecahan Glukosa Menjadi Laktat
Terdapat dua senyawa penting pemberi gugus fosfat kepada ADP, yaitu 3-fosfogliseroil fosfat dan fosfoenolfiruvat. Keduanya dibentuk pada reaksi pemecahan glukosa menjadi laktat yang menghasilkan energi. Proses ini disebut glikolisis.
m
Sebagian besar energi bebas yang dilepaskan pada degradasi glukosa menjadi laktat di simpan dalam kedua senyawa tersebut. Di dalam sel, senyawa fosfat berenergi tinggi tidak mengalami hidrolisis, namun gugus fosfat tersebut dipindahkan ke ADP menghasilkan ATP dengan katalis enzim kinase. Reaksi pemindahan gugus fosfat pada senyawa 3-fosfogliseroil fosfat adalah:
Glukosa Laktat 3-fosfogliseroil fosfat Fosfoenolfiruvat ATP ATP Senyawa antara
berenergi super tinggi Pembentukan 2 jenis senyawa fosfat berenergi super tinggi pada pemecahan glukosa menjadi laktat yang menghasilkan energi. Masing-masing dapat memberikan gugus fosfat kepada ADP, untuk membentuk ADP.
11 Hanya satu di antara dua gugus fosfat dari fosfogliseroil fosfat yang dipindahkan ke ADP, yaitu gugus fosfat yang terikat pada gugus karboksil. Gugus fosfat lainnya (pada karbon no. 3) merupakan gugus berenergi rendah.
Fosfoenolfiruvat, senyawa fosfat berenergi tinggi lainnya, yang dibentuk pada pemecahan glukosa menjadi piruvat juga memberikan gugus fosfatnya ke ADP melalui reaksi serupa, yang dikatalisis oleh piruvat kinase.
Pemindahan Gugus Fosfat dari ATP ke Suatu Molekul Penerima Dapat Memberikannya Tenaga
ATP dapat memindahkan gugus fosfat kepada berbagai molekul penerima untuk menghasilkan senyawa fosfat berenergi rendah, seperti senyawa fosfat dari alkohol. Terdapat heksokinase, yang mengkatalisa pemindahan gugus fosfat dari ATP ke D-glukosa.
Yang lain adalah gliserol kinase¸ yang mengkatalisis reaksi:
Glukosa 6-fosfat dan gliserol 3-fosfat berperan sebagai unit pembangun yang telah diaktifkan untuk sintesis molekul yang lebih besar. Sebagian dari energi bebas yang semula dihasilkan oleh degradasi glukosa menjadi laktat, yang disimpan dalam bentuk fosfogliseroil fosfat dan fosfoenolfiruvat, dapat dipindahkan ke gliserol, glukosa, atau penerima fosfat lainnya, di mana ATP berperan sebagai pembawa energi kimia antara dalam bentuk gugus fosfat.
12 ATP Dipergunakan untuk Memberikan Energi pada Kontraksi Otot
ATP memberikan energi kimia bagi dua bentuk utama kerja sel, yaitu kerja mekanik pada kontraksi otot dan kerja osmotik atau kerja melawan konsentrasi yang diperlukan untuk melakukan transport melawan konsentrasi gradien.
Di dalam sistem kontraktil sel otot kerangka terdapat dua jenis filament utama, yaitu filamen tebal yang terdiri dari serangkaian molekul miosin berbentuk batang yang tersusun secara parallel, dan filamen tipis yang terdiri dari dua untaian serabut aktin (F aktin) yang saling membelit terhadap sesamanya. Pada serabut otot, filamen tebal dan tipis secara teratur disusun sejajar, saling terjalin, dalam suatu unit berulang yang disebut sarkomer.
Selama kontraksi otot, filamen tipis di dalam tiap sarkomer, yang menyebabkan terjadinya pemendekan keseluruhan serat otot. Hidrolisis ATP menjadi ADP dan fosfat, memberikan energi kimia bagi gerakan meluncur filamen-filamen ini. Tiap-tiap molekul miosin pada filamen tebal memiliki kepala dengan ukuran besar, dan kepala tersebut sebenarnya merupakan enzim. Molekul ini menghidrolisis ATP bersamaan dengan berlangsungnya persinggung dan pelepasan berulang-ulang dengan filamen tipis, sehingga tenaga meluncur yang
13 menyebabkan filamen tebal bergerak di sepanjang filamen tipis menuju ke ujung sarkomer. Hidrolisis ATP dianggap terjadi bersama-sama dengan perubahan dalam bentuk atau konformasi kepala miosin, sehingga dihasilkan suatu tenaga mekanik. Dengan cara ini, miosin dan aktin berfungsi untuk mengubah energi kimia menjadi energi mekanik pada kontraksi otot.
Kontraksi dan relaksasi otot kerangka diatur oleh konsentrasi Ca2+ di dalam sitosol. Pada saat syaraf penggerak memberikan rangsangan terhadap serabut otot, Ca2+ dibebaskan dari tubul membran yang memanjang pada sel otot. Ca2+ yang dibebaskan berikatan dengan troponin, yang brfungsi sebagai pelatuk. Molekul ini mengalami perubahan konformasi yang menimbulkan aktivitas ATPase pada kepala molekul miosin pada filamen tebal, sehingga terjadi kontraksi. Otot relaksasi terjadi jika impul syaraf terhenti, dan Ca2+ diangkut dari sarkoplasma ke dalam sisterne pada sarkoplasmik reticulum melalui aktivitas pompa Ca2+ ATPase pada membran. Jadi, energi ATP diperlukan saat kontraksi dan relaksasi otot, di mana energi yang dibebaskan oleh hidrolisis ATP dapat digunakan untuk memompa ion lain melalui membran.
Fosfokreatin Merupakan Bentuk Cadangan Sementara dari Gugus Fosfat Berenergi Tinggi di dalam Otot
Fosfokreatin (disebut juga kreatin fosfat) berperan sebagai bentuk cadangan sementara gugus fosfat berenergi tinggi, di mana ∆Go’ hidrolisisnya sebesar -10,3 kkal/mol. Fosfokreatin dapat memindahkan gugus fosfatnya kepada ATP, dalam suatu reaksi yang dikatalisis oleh enzim kreatin kinase.
Fosfokreatin + ADP kreatin + ATP
Fosfokreatin berfungsi untuk mempertahankan konsentrasi ATP di dalam sel otot. Jika sebagian ATP pada sel otot digunakan untuk kontraksi, terjadi pembentukan ADP. Dengan bantuan kreatin kinase, fosfokreatin dengan cepat memberikan gugus fosfatnya ke ADP untuk mengembalikan tingkat normal ATP. Kandungan fosfokreatin otot kira-kira 3-4 kali lebih besar dari kandungan ATP, sehingga senyawa ini dapat menyimpan gugus fosfat dalam jumlah cukup untuk
14 mempertahankan tingkat ATP supaya tetap. Selama periode pengembalian selanjutnya, kreatin yang terakumulasi difosforilasi kembali menjadi fosfokreatin oleh ATP, karena reaksi kreatin kinase bersifat dapat balik.
Pada kebanyakan invertebrata, ototnya mengandung fosfoarginin, sebagai pengganti fosfokreatin sebagai bentuk cadanagn energi. Senyawa seperti fosfokreatin dan fosfoarginin yang berfungsi sebagai cadangan energi disebut fosfagen.
ATP juga Mengaktifkan Transport Aktif Melalui Membran
Energi ATP juga dapat digunakan untuk melaksanakan kerja konsentrasi, yaitu kerja yang diperlukan untuk mengangkut ion atau molekul melalui membran ke dalam ruang cair lain dengan dengan konsentrasi yang lebih tinggi. Untuk menghitung energi bebas yang diperlukan untuk mengangkut 1,0 mol senyawa terlarut nonionik dari satu ruangan ke ruangan lain dengan persamaan:
C1 : konsentrasi molar senyawa terlarut di dalam medium sekeliling C2 : konsentrasi molar di dalam sel
R : tetapan gas T : suhu absolut
Contoh: menentukan energi bebas yang diperlukan untuk mengangkut 1 mol glukosa, melawan ratusan kali gradien dari medium berkonsentrasi 1,0 mM ke dalam ruangan berkonsentrasi akhir 100 mM.
= 1360 (2,0)
15 = 2,72 kkal/mol
Karena energi bebasnya bernilai positif, maka transport 1 mol glukosa melawan seratus gradien membutuhkan sedikitnya 2,72 kkal/mol.
Transport aktif adalah trasnport senyawa terlarut melalui membran terhadap gradien konsentrasi. Membran sel darah mengandung enzim khusus, yang dinamakan ATPase pengangkut Na+ K+, yang berfungsi baik sebagai suatu enzim, maupun sebagai pompa molekuler. ATPase ini mengkatalisis hidrolisis ATP menjadi ADP dan fosfat, dan menggunakan energi bebas yang dibebaskan untuk memompa K+ ke dalam dan Na+ ke luar. Sebagai tahap pemberi energi dalam proses ini, gugus fosfat pada ujung ATP dipindahkan kepada molekul Na+ K+- ATPase. Gugus fosfat yang telah terikat pada enzim mengalami hidrolisis dan muncul sebagai fosfat anorganik di dalam sitosol pada saat molekul ini memberikan energi untuk mengangkut Na+ K+ pada arah yang berlawanan melalui membran, masing-masing melawan suatu gradien. ADP dan fosfat yang terbentuk lalu diubah kembali menjadi ATP oleh pemecahan glukosa yang menghasilkan Na+ K+- ATPase tidak menghidrolisis ATP, kecuali Na+ tersedia pada bagian dalam sel dan K+ tersedia di bagian luar. Na+ K+ - ATPase, yang memiliki 2 subunit dan 2 subunit β yang memanjang pada membran plasma. Molekul ini mengalami perubahan konformasi selama tahap pengangkutan ini. Pada ginjal, yang harus menghasilkan kelebihan Na+ di dalam urin dan menahan K+ ini dalam darah, hampir dua pertiga ATP yang dihasilkan oleh respirasi dipergunakan untuk mengangkut Na+ dam K+.
Berikut beberapa ATPase membaran pengankut kation.
Nama Jenis Sel Lokasi Fungsi
Na+ K+- ATPase Kebanyakan sel hewan
Membran plasma Mempertahankan konsentrasi tinggi K+di dalam sel
16 H+- ATPase Sel parietal pada
mukosa lambung
Membran plasma Sekresi H+ di dalam cairan lambung
H+- ATPase Sel hewan dan tumbuhan Sel tumbuhan Bakteri Membran sebelah dalam mitokondria Membran sebelah dalam kloroplas Membran plasma Berpartisipasi di dalam oksidatif dan fosforilasi fotosintetik ADP mejadi ATP C2+- ATPase Bakteri Sel hewan Membran plasma Retikulum sarkoplasmik Memompa Ca2+ keluar sel untuk mempertahankan Ca2+di dalam sitosol Memompa Ca2+ke dalam sisterna pada retikulum sarkoplasmik untuk membuat otot dalam keadaan istirahat
ATP dapat juga Diuraikan Menjadi AMP dan Pirofosfat
Meskipun secara umum menghasilkan ADP dan fosfat, pada beberapa reaksi sel pemakai ATP, kedua gugus fosfat pada ujung molekul ATP, yaitu gugus β dan γ, dilepaskan sebagai satu senyawa yaitu pirofosfat anorganik (dilambangkan sebagai PPi) dan adenosin monofosfat (AMP) sebagai produk lain
17 pada penguraian ATP. Contohnya adalah aktifasi enzimatik suatu asam lemak untuk membentuk ester koenzim A nya, suatu reaksi yang memberikan energi kepada asam dan mengubahnya menjadi suatu pemula aktif, asil lemak-KoA, bagi biosintesis lipid.
ATP + RCOOH + KoA-SH AMP + PPi +
Reaksi aktifasi ini berjalan dengan pelepasan pirofosfat dari ATP untuk menghasilkan pirofosfat dan AMP, berlainan dengan pemecahan ortofosfat pada umumnya dimana ATP kehilangan satu gugus ortofosfat seperti di dalam reaksi heksokinase.
ATP + D-glukosa ADP + D-glukosa 6-fosfat
Hidrolisi ATP untuk menghasilkan AMP + PPi
ATP + H2O AMP + PPi
Pirofosfat anorganik selanjutnya dihidrolisis oleh pirofosfatase menghasilkan dua molekul ortofosfat anorganik.
Pirofosfat + H2O AMP + 2Pi ΔGo’ = - 6,9 kkal/mol
Rekasi keseluruhan
Pirofosfat + H2O AMP + 2Pi ΔGo’ = -14,6 kkal/mol
Penggunaan 2 gugus fosfat dari ATP untuk mengaktifkan satu molekul prekursor terlihat seperti pemborosan energi gugus fosfat, tetapi hal ini merupakan cara penting untuk menjamin sempurnanya rekasi biosintetik tertentu.
ΔGo’
= -3,3 kkal/mol
Asam Lemak Asam Lemak - KoA
ΔGo’ = + 0,2 kkal/mol
18 Penggunaan menyimpang dari pembebasan pirofosfat dari ATP terjadi pada kunang-kunang yang menggunakannya sebagai sumber energi untuk memancarkan cahaya.
Pada kunang-kunang gabungan ATP dan energi oksidatif digunakan di dalam serangkaian reaksi yang mengubah energi kimia menjadi energi cahaya. Pertama, lusiferin diaktifkan oleh suatu reaksi enzimatik oleh ATP, dan pembentukan lusiferil adenilat. Senyawa ini sekarang bereaksi dengan molekul oksigen dan lusiferase menyebabkan reaksi dekarboksilasi oksidatif lusiferin menghasilkan oksilusiferin. Reaksi ini dengan tahap-tahap antaranya diikuti oleh pelepasan cahaya. Berikut siklus bioluminesen pada kunang-kunang.
Lusiferin dan lusiferase kunang-kunang dalam bentuk murni digunakan untuk mengukur sejumlah kecil ATP melalui pengukuran intensitas cahaya yang dihasilkan. ATP dapat diukur sampai sekecil beberapa pikomol (10-12 mol).
AMP dapat kembali ke siklus ATP melalui kerja enzim yang ada pada semua sel hewan yaitu adenilat kinase, yang mengkatalisis fosforilasi dapat balik AMP menghasilkan ADP.
ATP + AMP ADP + ADP
ADP yang dibentuk sekarang terfosforilasi kembali menjadi ATP.
Adenilat kinase memiliki fungsi penting lainnya. Jika enzim ini bekerja dengan arah sebaliknya.
19 2ADP ATP + AMP
Adenilat kinase membantu memindahkan gugus fosfat ujung dari suatu dari suatu molekul ADP ke molekul ADP lainnya, dan mengubah molekul kedua ini menjadi ATP. Pada otot yang berkontraksi, adenilat kinase membiarkan penggunaan kedua gugus fosfat γ dan pada ATP sebagai sumber energi. Akibatnya, adenilat kinase yang bekerja pada ADP dapat membantu menghasilkan fosfokreatin sebagai sumber ATP selama kontraksi otot.
Terdapat Nukleosida 5’-Trifosfat Berenergi Tinggi Lain Disamping ATP
Uridin trifosfat (UTP), guanosin trifosfat (GTP) dan sitidin trifosfat (CTP) merupakan analof ribonukleotida berfosfat bagi struktur ATP dan dengan ΔGo’ hidrolisis yang sama. Senyawa-senyawa ini terdapat di dalam semua sel, tetapi pada konsentrasi yang jauh lebih rendah dibandingkan ATP. Juga yang ditemukan di dalam sel pada konsentrasi rendah adalah senyawa-senyawa deoksiribonukleosida 5’-trifosfat deoksiadenosein 5’-trifosfat (dATP), 2’-deoksiguanosin 5’-trifosfat (dCTP) dan 2’-deoksitimidin 5’-trifosfat (dTTP).
Walaupun ATP merupakan pembawa gugus fosfat utama di dalam sel, jenis nukleosida 5’-trifosfat lainnya dikhususkan untuk berfungsi pada lintas biosintetik tertentu. Senyawa-senyawa ini memperoleh gugus fosfat terminalnya dari ATP dalam reaksi yang dikatalisis oleh enzim yang bergantung kepada Mg2+ yang dinamakan nukleosida difosfokinase, yang membantuk jenis reaksi balik berikut ini
ATP + UDP ADP + UTP ATP + GDP ADP + GTP ATP + CDP ADP + CTP GTP + UDP GDP + UTP
20 ATP + dCDP ADP + dCTP
GTP + dADP GDP + dATP Sistem ATP Berfungsi di dalam Keadaan Imbang Dinamik
Untuk mengetahui bahwa sistem ATP berfungsi untuk memindahkan energi fosfat di dalam sel utuh digunakan fosfor radioaktif (32P) sebagai pelacak dalam percobaan. Salah satu cara dengan menentukan kecepatan kembalinya gugus fosfat ujung ATP yang terdapat di dalam sel atau jaringan ke media dengan menggunakan fosfat anorganik yang dicirikan oleh 32P. Fosfat anorganik tersebut masuk ke dalam sel yang sedang melakukan aktivitas respirasi, dan kandungan ATP sel diisolasi dari contoh sel pada selang waktu pendek sesudah konsumsi fosfat tersebut. Walaupun konsentrasi ATP tidak berubah, gugus fosfat terminal dengan cepat menjadi fosfat radioaktif, menunjukkan bahwa gugus fosfat ujung atau γ cepat terurai dan cepat pula digantikan oleh fosfat radioaktif dari pool fosfat anorganik. Proses balik ini berlangsung sampai spesifik radioaktif gugus fosfat ujung dari ATP sama dengan fosfat anorganik pada pool. Kecepatan balik gugus fosfat terminal (ujung) ATP ternyata amat tinggi. Waktu paruh bagi pergantian gugus fosfat ujung ATP pada sel hati yang sedang melakukan respirasi hanya semenit atau 2 menit, di dalam sel bakteri anaerobik yang bernafas pada kecepatan yang jauh lebih tinggi daripada sel hewan, kecepatannya mencapai hanya dalam waktu detik. Sebaliknya, gugus fosfat `α pada ATP yang terikat langsung pada bagian D-ribosa molekul terurai dan terikat dengan amat lambat.
Walaupun kecepatan berbalik gugus fosfat ujung ATP amat tinggi di dalam sel hidup, konsentrasi total ATP tetap konstan, dan oleh karena itu kita memiliki suatu keadaan imbang dinamis. Pada keadaan ini, kecepatan penggunaan ATP untuk memberikan energi diimbangi oleh kecepatan refosforilasi ADP membentuk ATP bersamaan dengan proses oksidasi bahan bakar sel, sehingga ATP di bentuk dari ADP dan fosfat pada kecepatan yang cukup untuk mengatasi kebutuhan energi sel pada setiap waktu.