HASIL DAN PEMBAHASAN
Ekstraksi dan Amplifikasi Sampel Daun
Ekstraksi dalam penelitian ini menggunakan metode CTAB yang telah dilakukan terhadap 30 sampel daun. Hasil elektroforesis rata-rata menunjukkan hasil pita DNA yang cukup jelas, namun ada juga beberapa yang kurang jelas. Settanni et al. (2006) menyatakan bahwa suatu sampel DNA dinyatakan berhasil diamplifikasi apabila hasil analisis elektroforesis menunjukkan terdapatnya pita tunggal DNA dengan
ukuran sesuai dengan penanda yang telah diketahui
sebelumnya.
Gambar 2. Hasil elektroforesis dengan visualisasi uv transiluminator dari optimalisasi DNA kemenyan toba.
Silalahi (2013) menyatakan bahwa di Provinsi Sumatera Utara terdapat 2 (dua) jenis pohon kemenyan paling dominan yaitu kemenyan toba (Styrax
sumatrana) dan kamenyan durame (Styrax benzoin)danpaling banyak di
Ekstraksi DNA merupakan langkah awal yang harus dikerjakan dalam rekayasa genetika sebelum melangkah ke proses selanjutnya. Prinsip dasar ekstraksi total DNA/RNA dari jaringan adalah dengan memecah dan mengekstraksi jaringan tersebut sehingga akan terbentuk ekstrak sel yang terdiri atas sel-sel jaringan, DNA, dan RNA. Kondisidaun cukup baik dan tidak busuk setelah sampai di Laboratorium dan dapat diisolasi.
Perusakan membran sel dilakukan dengan menambahkan buffer ekstraksi, yaitu buffer CTAB, sedangkan perusakan secara fisik dilakukan dengan penggerusan dalam mortal steril. Menurut Ardiana (2009) menyatakan bahwa penggunaan buffer CTAB adalah sebagai pengganti nitrogen cair untuk mengisolasi DNA sehingga dapat menghasilkan produk DNA yang berkualitas tinggi. Hasil ekstraksi daun kemudian dielektroforesis untuk dapat membaca pita DNA.
smeardi bawah pita DNA. Kesalahan selanjutnya dapat terjadi karena tidak
tepatnya jumlah konsentrasi pereaksi yang digunakan, dan tidak tepatnya pengaturan kondisi PCR.
Sekuensing DNA
Hasil sekuensing produk PCR dari total 30 sampel yang diamplifikasi menggunakan primer trnL-trnF tidak semua memiliki sekuen dan grafik kromatrogram yang cukup jelas. Namun sebanyak 26 sampel memiliki sekuen yang baik, dan 4 lainnya tidak dapat teridentifikasi dengan baik yaitu Styrax
sumatrana yang berasal dari Humbang Hasundutan (SS13HB, dan SS20HB), dan
dari Tapanuli Utara (SS23TU dan SS24TU) sehingga tidak diikutsertakan untuk tahapan analisa berikutnya.
Elektroforegram terdiri dari beberapa kurva dengan tinggi puncak dan warna yang berbeda tergantung dari jenis basa. Setiap jenis basa memiliki notasi yang berbeda-beda yaitu notasi A untuk adenin, notasi T untuk timin, notasi G untuk guanin, dan notasi C untuk sitosin. Seringkali terdapat lebih dari satu puncak yang bertumpuk pada satu posisi dengan tinggi puncak yang berbeda-beda atau terlalu rendahnya puncak yang dihasilkan dari suatu nukleotida yang dinotasikan dengan N. Hal ini disebabkan karena jumlah molekul DNA kloroplas yang sangat banyak dalam satu sel. Puncak yang rendah menunjukkan jumlah nukleotida yang jumlahnya sedikit atau minoritas, sedangkan puncak yang tinggi menunjukkan jumlah nukleotida yang banyak atau mayoritas.
Karakterisasi Sekuen
substitusi, inersi dan delesi. Variasi genetik tersebut membagi sekuen ke dalam empat haplotipe yang dapat dilihat dari polimorfisme basa pada sekuen. Haplotipe dasar diperoleh dari perbedaan basa nukleotida yang paling sedikit.
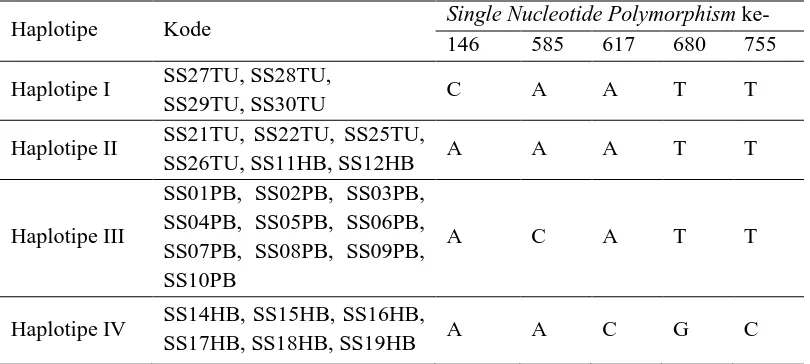
Tabel 1. Perbedaan jenis basa pada urutan nukleotida trnL-trnFStyrax sumatrana.
Haplotipe Kode Single Nucleotide Polymorphism ke- 146 585 617 680 755
Haplotipe I SS27TU, SS28TU,
SS29TU, SS30TU C A A T T
Haplotipe II SS21TU, SS22TU, SS25TU,
SS26TU, SS11HB, SS12HB A A A T T
Haplotipe IV SS14HB, SS15HB, SS16HB,
SS17HB, SS18HB, SS19HB A A C G C
Keterangan : Kode SS27TU adalah untuk sampel Styrax sumatrana yang berasal dari Tapanuli Utara pohon ke-27 dari total seluruh pohon sampel. Kode SS01PB adalah untuk sampel yang berasal dari Pakpak Bharat, sedangkan kode SS14HB adalah untuk sampel yang berasal dari Humbang Hasundutan.
Haplotipe I dianggap sebagai haplotipe dasar atau awal. Haplotipe II diperoleh dari adanya satu perbedaan polimorfisme basa ke-146 dengan haplotipe 1, yaitu A yang menggantikan C. Haplotipe III diperoleh dari adanya dua perbedaan polimorfisme basa ke-146 dan ke-558 dengan haplotipe 1, yaitu A dan C yang menggantikan C dan A pada haplotipe 1. Haplotipe IV diperoleh dari adanya tiga perbedaan polimorfisme basa ke-146, 617, dan 680 dengan haplotipe 1, yaitu A, C, G, yang menggantikan C, A, T.
Frekuensi mutasi paling tinggi dan mudah diamplifikasi maupun disekuen secara langsung karena ukurannya yang tidak terlalu panjang. Variasi terbanyak terjadi pada Styrax sumatrana yang berasal dari daerah bagian intron sekuen
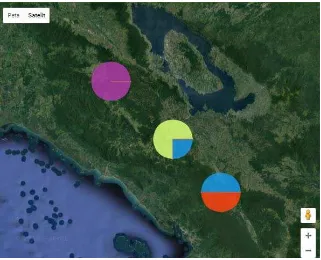
Gambar 3. Peta persebaran 4 haplotipe dari kemenyan toba di tiga daerah (Sumber: Phylogeoviz.org.)
Keterangan : Diagrampie berwarna merah untuk haplotipe I dan berwarna biru untuk haplotipe II pada daerah Tapanuli Utara; diagram pie berwarna biru untuk haplotipe II dan berwarna kuning untuk haplotipe VI pada daerah Humbang Hasundutan; dan diagram pie berwarna ungu untuk haplotipe III pada daerah Pakpak Bharat.
Untuk analisis mutasi dilakukan dengan mengurutkan nukleotida setiap sampel yang telah disejajarkan menggunakan software BioEdit. Penyejajaran dilakukan dengan tujuan untuk menentukan tingkat homologi dari urutan basa DNA sampel yang dianalisis. Sekuen yang dihasilkan memiliki variasi genetik karena beberapa nukleotida mengalami substitusi, inersi dan delesi. Pada hasil penyejajaran muncul gapyang ditandai dengan garis putus-putus. Gap menunjukkan terjadinya proses mutasi baik berupa delesi maupun inersi. Sedangkan substitusi ditandai dengan polimorfisme salah satu nukleotida seperti yang ditampilkan pada Tabel 1.
lampau dan/atau waktu sekarang. Waktu terjadinya mutasi tersebut secara tepat sulit ditentukan, tetapi berdasarkan sifat DNA kloroplas yang konservatif dan kecepatan mutasi yang rendah yaitu 3.2 x 10-5dan 7.9 x 10-5(Provan et al. 1999) boleh jadi peristiwa mutasi terjadi pada masa lampau. Gen yang mengalami mutasi satu basa dapat mengakibatkan perubahan asam amino yang dihasilkan sehingga berdampak pada perubahan fenotipe tanaman di lapangan.
Adanya mutasi genetik seperti substitusi, delesi, dan inersi dapat menghasilkan beberapa haplotipe. Haplotipe (haploid genotipe) adalah urutan DNA yang telah diwarisi dari satu induk. Jenis variasi yang paling umum di antara haplotipe yang dimiliki oleh individu dalam populasi adalah Single
Nucleotide Polymorphism(SNP), di mana nukleotida berbeda (alel) hadir di lokasi
tertentu (Gusfield & Orzack, 2001).
Genetika populasi Styrax sumatrana di daerah Humbang Hasundutan dan Tapanuli Utara lebih bervariasi yaitu memiliki 2 haplotipe dibandingkan dengan di daerah Pakpak Bharat yang hanya memiliki satu haplotipe. Hal ini dapat dilihat pada Gambar 4.4 yaitu peta persebaran 4 haplotipe dari kemenyan toba di tiga daerah. Diagram pie berwarna ungu untuk haplotipe III pada daerah Pakpak Bharat menunjukkan bahwa DNA Styrax sumatrana di daerah tersebut bersifat unik atau terisolasi karena hanya terdapat pada daerah Pakpak Bharat saja. Sedangkan pada daerah Humbang Hasundutan dan Tapanuli Utara terjadi sharing genetik.
II sebesar 50% dan merah untuk haplotipe I sebesar 50%. Hal ini disebabkan oleh beberapa faktor, menurut Hamrick dan Godt (1990) kondisi geografi merupakan faktor yang paling besar peranannya dalam variasi genetik (32%), diikuti mekanisme kehidupan (25%), serta sistem perkawinan dan penyebaran biji (±17%).
Kondisi geografis berpengaruh penting terhadap distribusi haplotipe. Pada penelitian ini, kondisi lingkungan dari ketiga lokasi berbeda-beda dilihat dari ketinggian tempat, jarak antar lokasi dan keadaan fisik lingkungan seperti gunung, sungai, dan bukit. Lokasi pengambilan sampel di Pakpak Bharat yaitu Desa Pardomuan, Kecamatan Sitellu Tali Urang Julu, tepatnya berada di bukit Lae Jerring. Bukit Lae Jerring terletak di ketinggian 1113 mdpl yang merupakan bagian dari punggung bukit barisan. Dapat dilihat pada peta bahwa terdapat hamparan bukit barisan yang tinggi nya lebih dari lokasi pengambilan sampel di Pakpak Bharat, sehingga menjadi faktor penghambat terjadinya gene flow dari daerah Humbang Hasundutan maupun Tapanuli Utara.
Menurut Henuhili (2008), isolasi genetik yang disebabkan oleh satu atau lebih mutasi hanya dapat timbul sesudah terjadinya isolasi geologi dalam waktu yang lama. Isolasi ini menghasilkan perbedaan yang nyata antara kedua kelompok populasi. Apabila dua populasi yang berbeda beradaptasi pada lingkungan yang berbeda, maka masing-masing populasi akan mengakumulasi perbedaan-perbedaan yang terjadi dalam kumpulan gen (perbedaan-perbedaan frekuensi alel dan genotip).
barisan di bagian sisi yang berlawanan dengan Pakpak Bharat yaitu dengan ketinggian 990 mdpl. Jarak dari Humbang Hasundutan ke Tapanuli Utara juga lebih dekat daripada Jarak Humbang Hasundutan ke Pakpak Bharat. Hal ini dapat menyebabkan terjadinya sharing genetik atau gen flow antara gen Styrax
sumatrana di Humbang Hasundutan dan Tapanuli Utara.
Dalam merupakan transfer dalam atau keluar suatu populasi dapat bertanggung jawab terhadap perubaha
Su, H et al. (2003) menyatakan bahwa terdapat sejumlah faktor-faktor yang memengaruhi aliran gen antara populasi-populasi yang berbeda. Salah satu faktor yang paling signifikan adalah mobilitas. Semakin besar mobilitas suatu individu, semakin besar potensi migrasi individu tersebut.Faktor-faktor lain yang dapat mempengaruhi laju aliran gen dapat mencakup jarak antara dua populasi, atau hambatan fisik tertentu seperti gunung, sungai, atau struktur buatan manusia tertentu.Pada tumbuhan, laju aliran gen tergantung pada efektivitas mekanisme penyebaran serbuk sari dan biji yang digunakan. Hal ini dapat terjadi karena adanya bantuan dari angin maupun hewan yang dapat membawa serbuk sari hingga jarak yang cukup jauh.
Analisis Filogenetik
Analisis filogenetik dilakukan dengan menggunakan software Mega 5, menggunakan metode Neighbour Joining (NJ). Analasis terhadap gen
trnL-trnFmelibatkan 57 data. Data tersebut antara lain adalah dari sampel sebanyak 26
gene of Styrax”. Data sekuens trnL-trnF Styrax yang diunduh antara lain Styrax
suberifolius, Styrax chinensis, Styrax gentryl, Styrax pentlandianus, Styrax
nunezii, Styrax latifolius, Styrax peruvianus, Styraz camporum, Styrax leprosus,
Styrax pohlii, Styrax obtusifolius, Styrax ferrugineus, Styrax rotundatus, Styrax
acuminatus, Styrax tomentosus, Styrax lanceolatus, Styrax glaber, Styrax
portoricensis, Styrax martii, Styrax laberi, Styrax ubargenteus, Styrax officinalis,
Styrax benzoin, Styrax aureus, Styrax japonicus, dan Styrax agrestis. Sampel dan
data dari NCBI di sejajarkan dengan menggunakan menu Align by Muscle.
Pohon filogenetik merupakan grafik yang digunakan untuk menggambarkan hubungan kekerabatan antar spesies yang terdiri atas sejumlah nodus dan cabang dengan hanya satu cabang yang menghubungkan dua nodus paling berdekatan.Setiap nodus mewakili unit-unit taksonomi dan setiap cabang mewakili hubungan antar unit yang menggambarkan hubungan keturunan dengan leluhur.
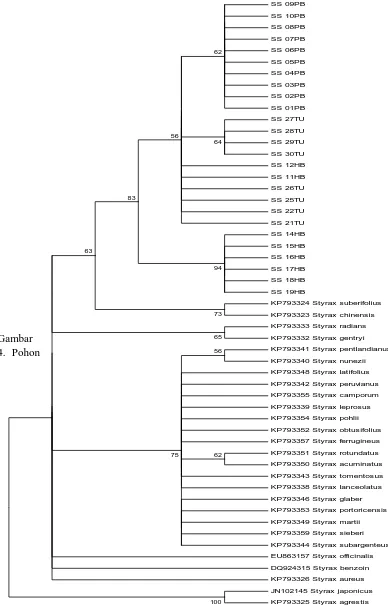
Pohon filogenetik yang dihasilkan oleh metode Neighbour Joining (NJ) menghasilkan hipotesis hubungan kekerabatan antarsampel berdasarkan jarak genetik pada gen trnL-trnF (Gambar 4). Pohon filogenetik yang telah direkonstruksi diuji secara statistik untuk meningkatkan nilai kepercayaan. Pada penelitian kali ini, pohon filogenetik diuji secara statistik menggunakan metode
bootstrap sebanyak 1000 ulangan.
sumatrana pohon ke 1 sampai 10 yang berasal dari Pakpak Bharat, Styrax
sumatrana pohon ke 27 sampai 30 tang berasal dari Tapanuli Utara dengan nilai
bootstrap 64%, Styrax sumatrana pohon ke 11 dan 12 dari Humbang Hasundutan
dan pohon ke 21, 22, 25, dan 26 yang berasal dari Tapanuli Utara dengan nilai
bootstrap 56%, dan Styrax sumatrana pohon ke 14 sampai 19 yang berasal dari
Humbang Hasundutan dengan nilai bootstrap 94%.
Dari pohon filogenetik dapat dilihat bahwa Styrax sumatrana memiliki monofiletik yang sama dengan Styrax suberifolius dengan Styrax chinensis yaitu dengan nilai bootstrap 63%. Sedangkan Styrax sumatrana dengan Styrax benzoin berjarak cukup jauh walaupun tumbuh dilokasi yang sama dengan lokasi pengambilan sampel.
Gambar 4. Pohon
filogenetik berdasarkan sekuens trnL-trnF.
Panjang sekuen trnL-trnF yang diperoleh antara lain 941 bp setelah di sejajarkan, hal ini sesuai dengan pernyataan Holt et al. (2005) yang menyatakan
bahwa plastid trnL (UAA) dan trnF (GAA) merupakan gen pengkode RNA transfer dan di antara kedua gen tersebut terdapat sekitar 1.000 bp sekuen daerah non-pengkode (intron dari trnL (UAA) dan intergenic spacer (IGS) dari
trnL-trnF (GAA).
Daerah noncoding merupakan daerah yang menunjukkan frekuensi mutasi paling tinggi sehingga dimungkinkan terdapat banyak perbedaan pada daerah tersebut. Menurut Taberlet (1991) daerah non-pengkode tersebut merupakan daerah yang menunjukkan frekuensi mutasi paling tinggi dan mudah diamplifikasi maupun disekuen secara langsung karena ukurannya yang tidak terlalu panjang.
Analisis filogenetik dilakukan untuk mengetahui kekerabatan antara Styrax
sumatrana dari tiga daerah di Sumatera Utara dan dengan spesies Styrax yang
lainnya yang diunduh dari NCBI. Hubungan kekerabatan dikonstruksi dengan menggunakan fragmen DNA kloroplas yaitu trnL-trnF (daerah dari trnL (UAA) 5’ ekson hingga trnF(GAA)). Pohon filogeni dibentuk dengan menggunakan metode Neighbour Joining (NJ) (Gambar 4).
Dari pohon filogenetik dapat dilihat bahwa sampel Styrax sumatrana bersifat monofiletik (mengelompok bersama berdasarkan asalnya). Menurut Hidayat et al. (2005), kelompok monofiletik merupakan kelompok yang anggotanya berasal dari satu nenek moyang. Anggota dalam kelompok monofiletik diasumsikan membawa sifat atau pola genetik yang sama.
sehingga semakin besar nilai bootstrap yang digunakan maka semakin tinggi tingkat kepercayaan topologi pohon hasil konstruksi tersebut.
Pohon filogenetik memberi informasi tentang pengklasifikasian populasi berdasarkan hubungan evolusionernya. Akar pada pohon menggambarkan titik percabangan pertama atau asal masing-masing populasi dengan asumsi bahwa laju evolusi berjalan konstan (Dharmayanti, 2011). Panjang cabang menggambarkan jumlah substitusi basa yang dapat berupa polimorfisme DNA atau haplotipe.
Analisis jarak antar genetik dilakukan dengan menggunakan Pairwise
Distance menggunakan model Kimura-2 parameter (Kimura, 1980). Nilai
similaritas = (1 – distance) x 100%. Hasil menunjukkan jarak genetik antara Styrax sumatrana dari Tapanuli Utara dengan Pakpak Bharat adalah 0,003 atau memiliki similaritas 99,7%, nilai similaritas Tapanuli Utara dengan Humbang Hasundutan adalah 99%, sedangkan nilai similaritas Humbang Hasundutan dengan Pakpak Bharat adalah 99,7%.
Jarak genetik Styrax sumatrana dengan Styrax chinensis dan Styrax suberifolius adalah 0,005 atau dengan similaritas 99,5%. Sedangkan Styrax sumatrana dengan Styrax benzoin memiliki similaritas 99,3%. Menurut Irawan et
al. (2016), semakin sedikit nilai jarak genetik antara dua organisme, semakin
KESIMPULAN DAN SARAN
Kesimpulan
Analisis sekuens dari kemenyan toba (Styrax sumatrana) dengan menggunakan trnL-trnF diperoleh panjang sekuen sebesar 941 bp setelah disejajarkan. Variasi genetik yang dihasilkan tersebut membagi sekuen ke dalam empat haplotipe yang dapat dilihat dari polimorfisme basa pada sekuen. Adanya aliran gen antara dua populasi yaitu Humbang Hasundutan dan Tapanuli Utara, sedangkan gen kemenyan toba di Pakpak Bharat terisolasi dan bersifat unik. Aliran gen dapat dipengaruhi oleh faktor geografis, hambatan fisik, serta jarak antar populasi. Dari pohon filogenetik dapat dilihat bahwa Styrax sumatrana memiliki monofiletik yang sama dengan Styrax suberifolius dengan Styrax
chinensis yaitu dengan nilai bootstrap 63%.
Saran