TINJAUAN PUSTAKA
Gambaran Umum Monyet Ekor Panjang (Macaca fascicularis)
Monyet ekor panjang/MEP dikenal juga sebagai long-tailed macaque, monyet cynomolgus, Macaca irus, monyet jawa dan monyet pemakan kepiting
(crab eating monkey). Monyet ekor panjang merupakan spesies dengan
populasi terbanyak di seluruh dunia, yaitu sekitar 20 juta ekor (Whitney 1995; Soehartono dan Mardiastuti 2002).
Monyet ekor panjang memiliki tubuh ramping dan berekor panjang, berkisar 60 cm. Monyet ini memiliki dimorfisme seksual, dengan bobot individu jantan berkisar antara 5-7 kg, lebih besar bila dibandingkan dengan individu betina, yaitu 3-4 kg. Secara morfologis, MEP jantan dan betina dewasa kelihatan sama, yaitu memiliki rambut kepala berwarna abu-abu sampai kecoklatan, sedangkan rambut bayi MEP berwarna hitam. Hal ini memperlihatkan perbedaan yang mencolok dengan warna rambut MEP dewasa. Dalam beberapa minggu setelah lahir, warna rambut bayi akan berubah menjadi coklat dan keabuan, yang menyerupai warna rambut MEP dewasa (Soehartono dan Mardiastuti 2002).
Menurut Rowe (1996), periode bayi berlangsung antara umur 6-12 bulan dengan masa sapih antara umur 12-24 bulan dan masa puber pada umur 42-54 bulan. Individu betina memasuki masa dewasa kelamin pada umur 51.6 bulan, sedangkan individu jantan pada umur 50.4 bulan. Siklus estrus MEP betina adalah 28 hari, dengan lama kebuntingan berkisar antara 160-170 hari. Jarak antara kebuntingan berkisar 12-24 bulan, dengan rata-rata berlangsung selama 13 bulan. Menurut Whitney (1995), masa hidup (life-span) MEP mencapai 22-25 tahun.
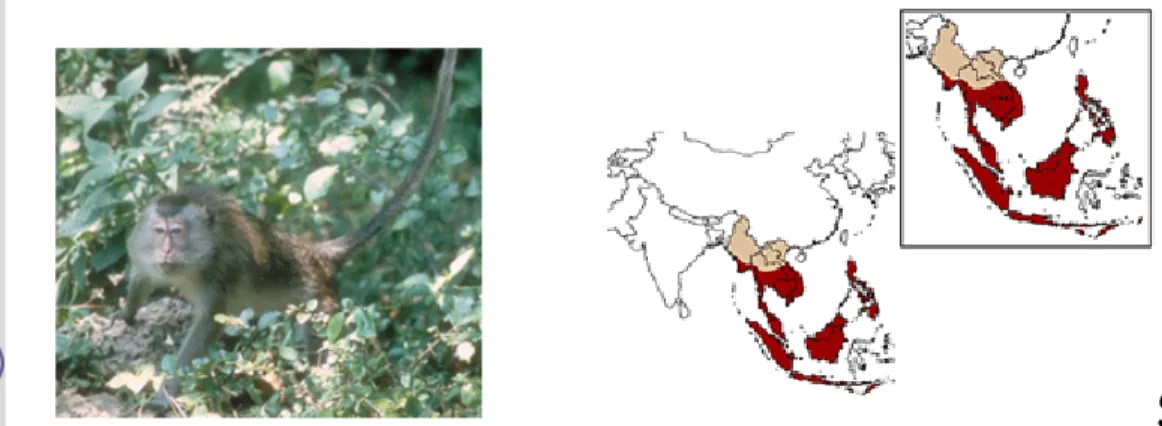
Penyebaran MEP di alam meliputi kawasan Asia Tenggara, antara 20 LU dan 10 LS serta 92 BT dan 127 BT. Satwa ini menempati wilayah geografis yang luas, mulai dari Myanmar, Thailand, Kamboja, Laos, Vietnam, Malaysia, Indonesia dan Filipina. Penyebaran satwa primata ini di Indonesia dimulai dari Pulau Sumatera hingga Pulau Timor (Soehartono dan Mardiastuti 2002), namun tidak ditemukan di Pulau Sulawesi (Gambar 1) (Napier dan Napier 1967).
Gambar 1 Monyet ekor panjang (M. fascicularis) dan peta penyebarannya (merah) (Sumber : Lang 2006).
Monyet ekor panjang sejak tahun 1977 termasuk ke dalam daftar Appendix II pada Convention on International Trade in Endangered Species of
Wild Fauna and Flora (CITES). Dari status tersebut, berarti MEP dapat
dimanfaatkan dan diperdagangkan, sejauh merupakan hasil penangkaran, karena populasinya masih cukup banyak (Soehartono dan Mardiastuti 2002).
Klasifikasi MEP (Whitney 1995) adalah sebagai berikut:
Filum : Chordata
Kelas : Mammalia
Ordo : Primata
Sub ordo : Anthropoidea
Infra ordo : Catarrhini
Super famili : Cercopithecoidea
Famili : Cercopithecidae
Sub famili : Cercopithecinae
Genus : Macaca
Spesies : Macaca fascicularis
Satwa primata berperan penting sebagai hewan model dalam penelitian biomedis, karena secara anatomi dan fisiologi memiliki banyak kemiripan dengan manusia dibandingkan dengan hewan model lainnya. Kemiripan itu disebabkan oleh hubungan filogenetik, sehingga menyebabkan satwa primata memiliki nilai ilmiah yang tinggi bagi penelitian di bidang biomedis (Vandeberg 1995).
Beberapa spesies primata non manusia yang sering digunakan sebagai hewan model adalah monyet rhesus (M. mullata), MEP (M. fascicularis), babun savanna
(Papio cynocephalus) dan monyet vervet (Cercopithecus aethiops) (Soehartono Anatomi dan Fungsi Hipofise
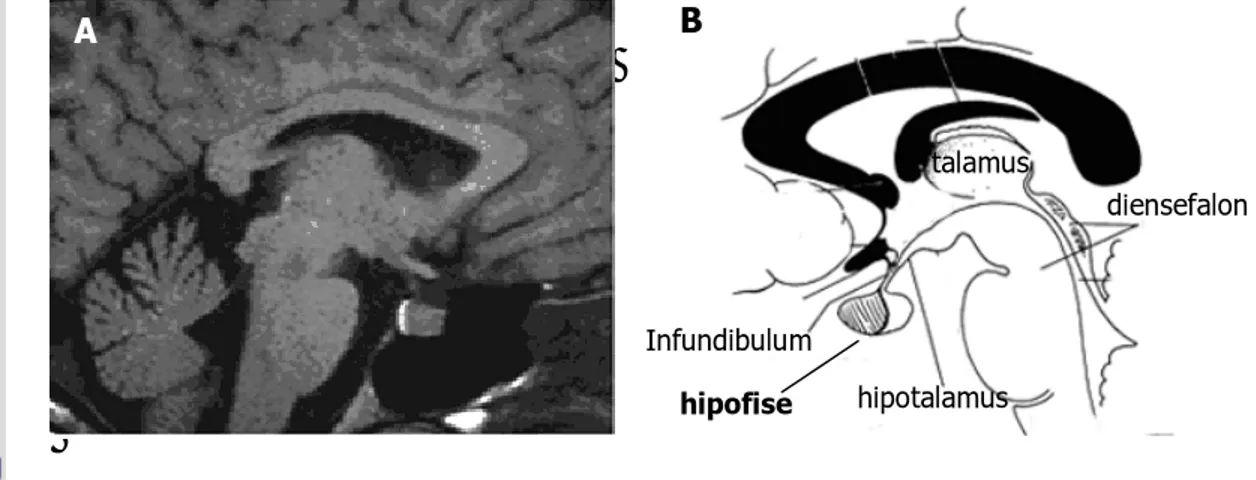
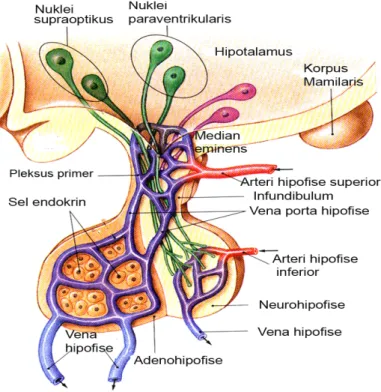
Hipofise seringkali disebut dengan istilah ‘the master of endocrine gland’ pada hewan vertebrata, karena menghasilkan sejumlah hormon penting yang berperan dalam proses fisiologi tubuh. Hipofise terletak di ventral hipotalamus, berbentuk ‘ellipsoidal’ dengan ukuran yang bervariasi, tergantung pada spesies hewan. Organ ini berada di fossa hipofisial atau sella tursica os sphenoid, dilapisi oleh selaput otak (duramater) dan dihubungkan dengan hipotalamus melalui tangkai hipofise atau infundibulum (Gambar 2) (Dyce et al. 1996). Sebagai kelenjar endokrin utama, organ ini berperan penting sebagai penghubung dalam sistem neuroendokrin, namun kelenjar ini memiliki kemampuan yang kecil untuk berfungsi secara bebas (Turner dan Bagnara 1976).
Gambar 2 Magnetic resonance image (MRI) hipotalamus dan hipofise manusia. A. Menunjukkan posisi hipofise di ventral otak tengah (diensefalon) yang berhubungan erat dengan hipotalamus, B. Skema dari gambar A (Sumber: Lechan dan Tony 2006).
A hipofise talamus hipotalamus diensefalon B Infundibulum
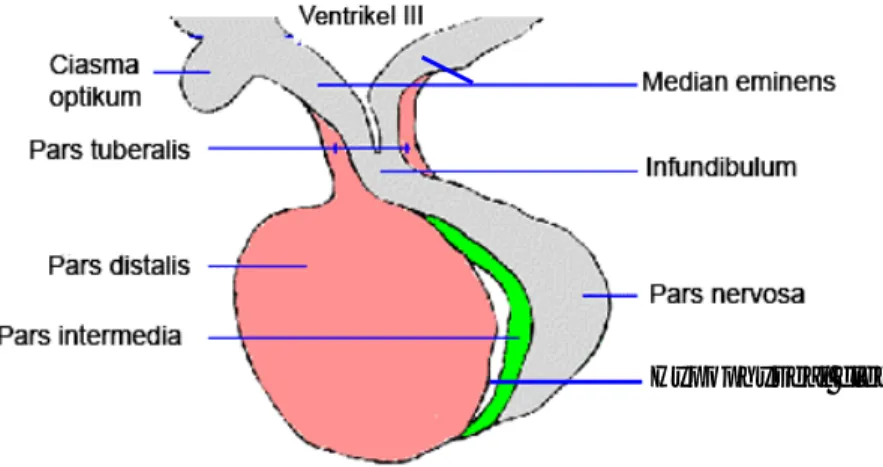
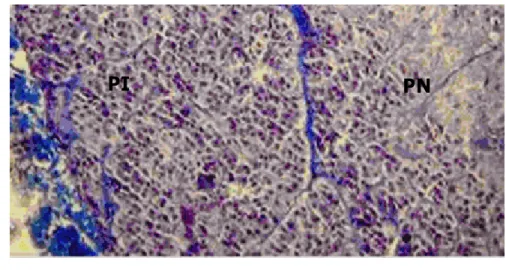
Hipofise terbagi atas dua bagian, yaitu adenohipofise dan neurohipofise (Gambar 3). Adenohipofise terdiri atas pars distalis (PD) yang merupakan bagian terbesar dari adenohipofise, pars tuberalis (PT) dan pars intermedia (PI) (Brown 1994). Neurohipofise terdiri atas median eminens, infundibulum dan pars nervosa (PN) (Kent dan Carr 2001). Gabungan PD dan infundibulum membentuk batang hipofise (Banks 1993).
Gambar 3 Hipofise dan bagian-bagiannya. Pars intermedia (warna hijau), berbatasan langsung dengan pars nervosa, dengan pars distalis dipisahkan oleh hypophyseal cleft (Sumber: KUL 2000).
Secara histologi, hipofise terdiri atas beberapa jenis sel, dimana masing-masing sel tersebut menghasilkan hormon yang berbeda. PD adenohipofise merupakan bagian yang memiliki jenis sel terbanyak yang menghasilkan enam jenis hormon. Sel-sel yang berada di PD terdiri atas dua kelompok, yaitu sel kromofilik dan sel kromofobik. Berdasarkan afinitas terhadap zat warna, sel kromofilik dibagi menjadi sel-sel asidofil dan basofil (Banks 1993). Sel-sel asidofil terdiri atas somatotrop dan laktotrop, sedangkan sel-sel basofil terdiri atas tirotrop, gonadotrop, kortikotrop dan melanotrop (Brown 1994). Sel-sel tersebut mensintesis hormon-hormon hipofise yang berperan dalam pengaturan kerja target organ seperti gonad, kelenjar adrenal, kelenjar air susu, uterus, ginjal dan jaringan tubuh lainnya (Frandson dan Whitten 1991).
Diantara sel-sel penghasil hormon, yaitu pada jaringan interstisial adenohipofise, terdapat sel folliculo-stellate. Sel ini memiliki penjuluran sitoplasmik yang menyebar di antara sel-sel sekretori hormon adenohipofise. Sel
folliculo-stellate berperan sebagai regulator terhadap lingkungan jaringan
interstisial hipofise dan membentuk komunikasi yang bersifat parakrin dengan sel-sel penghasil hormon (Allaerts dan Vankelecom 2005).
Perkembangan Hipofise
Hipofise merupakan suatu struktur ektodermal yang berasal dari dua sumber jaringan yang berbeda pada masa embrional. Adenohipofise primordial berasal dari penonjolan rongga mulut primitif (stomodeum) yang tergolong ektoderm umum, sedangkan neurohipofise berasal dari diensefalon yang tergolong ektoderm syaraf (Latshaw 1987; Hadley 1992).
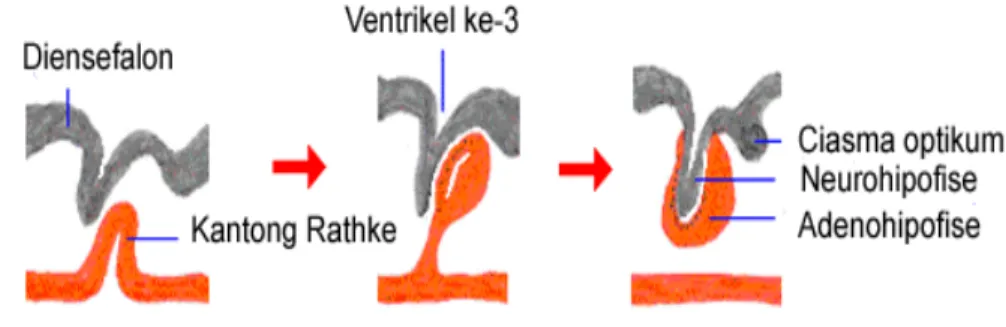
Perkembangan hipofise terjadi pada awal kebuntingan dan prosesnya berbeda pada setiap spesies hewan. Perkembangan adenohipofise berawal dari evaginasi atap rongga mulut ke arah dorsal yang membentuk kantong Rathke. Pada saat yang bersamaan terjadi penjuluran ektoderm syaraf, berupa evaginasi diensefalon otak ke arah ventral. Akibatnya, otak mengalami perluasan ke arah ventral membentuk neurohipofise (Gambar 4) (Latshaw 1987; Hill 2006).
Gambar 4 Tahapan perkembangan hipofise. Hipofise berasal dari dua jaringan berbeda, stomodeum (merah) dan diensefalon (abu-abu) (Sumber: Bowen 2006).
Perkembangan hipofise pada manusia terjadi pada minggu ke-4 kehamilan, diawali dengan terbentuknya kantong Rathke (yang berasal dari divertikulum atap rongga mulut). Pada minggu ke-5 terjadi elongasi divertikulum diensefalon (infundibulum). Adenohipofise terbentuk pada minggu ke-6, diikuti dengan proliferasi dinding anterior membentuk PD, sedangkan dinding posterior
mengalami sedikit perkembangan membentuk PI. PT terbentuk dari pertumbuhan bagian rostral yang mengelilingi sel bakal infundibular. Pada tahap selanjutnya terbentuk neurohipofise yang terdiri atas infundibulum, median eminens dan PN (Hill 2006).
Menurut Sasaki dan Nishioka (1998), perkembangan hipofise pada anjing beagle dimulai pada umur kebuntingan 25 hari. Pada umur tersebut, kantong Rathke (adenohipofise primordium) yang berasal dari jaringan epithelium rongga mulut mulai terbentuk. Memasuki usia kebuntingan 38 hari, PI telah terbentuk dan hipofise fetus anjing beagle secara morfologis telah menyerupai hipofise anjing dewasa.
Berdasarkan analisis imunohistokimia oleh Sasaki et al. (1992) terhadap hipofise babi, diketahui sel-sel hipofise yang pertama menghasilkan hormon di bagian PD dan PI adalah sel adrenocorticotropichormone (ACTH). Sel ini ditemukan pada fetus yang berumur 40 hari, diikuti dengan sel growth hormone (GH) dan luteinizing hormone (LH) pada fetus umur 60 hari dan sel prolactin (PRL) pada fetus umur 105 di bagian PD. Sel imunoreaktif ACTH (sel-ir-ACTH) di PI terlihat lebih banyak dan dengan intensitas lebih kuat dibandingkan dengan PD.
Keberadaan sel ir-ACTH sebagai sel pertama yang terbentuk di PD hipofise babi, ditemukan pula pada fetus anjing, manusia dan tikus, sehingga diduga ACTH berperan penting pada awal kehidupan fetus dibandingkan hormon-hormon adenohipofise lainnya. Peran ACTH lainnya adalah berhubungan dengan perkembangan duodenum, penyimpanan glikogen di hati dan inhibisi adrenal pada aktivitas adenohipofise selama kehidupan fetus (Sasaki dan Nishioka 1998).
Vaskularisasi Hipofise
Menurut Sasaki et al. (1992), pada fetus babi umur 40 hari dan fetus
anjing beagle umur 30 hari, pembuluh kapiler hipofise ditemukan di antara jaringan parenkim kantong Rathke. Sedangkan menurut Sasaki dan Nishioka (1998), pada fetus anjing beagle umur 38 hari, pembuluh kapiler terlihat di PD yang menerima darah dari vena porta hipofise. Namun, pleksus primer di median eminens belum ditemukan. Pleksus ini baru terlihat pada fetus umur 52 hari, di mana bagian ini telah berhubungan dengan vena porta hipofise.
Sistem portal hipofise merupakan suatu sistem vaskularisasi yang berperan dalam mekanisme neurohumoral yang mengatur fungsi tropik hipofise. Sistem tersebut membawa substansi yang berasal dari nervus terminal median
eminens ke hipofise (Halasz 2000). Hubungan vaskularisasi antara hipotalamus dan adenohipofise relatif kompleks dibandingkan dengan neurohipofise. Aliran darah menuju adenohipofise berasal dari arteri hipofise superior. Dari pembuluh ini darah dibawa ke median eminens hipotalamus membentuk suatu jalinan pembuluh kapiler yang disebut pleksus primer. Pada bagian ini, hormon hipotalamus dilepaskan dan dibawa ke pars tuberalis oleh vena porta hipofise dan bermuara di pleksus sekunder, yaitu jalinan pembuluh kapiler yang terdapat di adenohipofise (Gambar 5). Pada pleksus sekunder ini beberapa hormon hipotalamus merangsang sel-sel hipofise untuk menghasilkan hormon yang selanjutnya dilepaskan ke sirkulasi darah umum. Vaskularisasi neurohipofise berasal dari arteri hipofise inferior. Pada arteri ini dilepaskan pula hormon-hormon hipotalamus yang berasal dari nuklei supraoptik (SON) dan nuklei paraventrikular (PVN). Sebelum memasuki sirkulasi darah umum, hormon-hormon tersebut disimpan di pars nervosa neurohipofise (Brown 1994).
Gambar 5 Sistem vaskularisasi hipofise. Sistem portal hipofise (biru) mengalirkan darah dari hipotalamus dan menyalurkan hasil sintesis hormon-hormon adenohipofise melalui vena hipofise ke organ target (Sumber: Martini 2006)
Hormon-hormon Hipofise
Sebagai kelenjar endokrin terbesar, hipofise menghasilkan sejumlah
hormon penting, yang sebagian besar dihasilkan oleh PD adenohipofise (6 hormon), PI (2 hormon) dan PN (2 hormon). Hormon-hormon PD terdiri atas:
growth hormone (GH), adrenocorticotropic hormone (ACTH), thyroid stimulating hormone (TSH), follicle stimulating hormone (FSH), luteinizing hormone (LH),
dan prolactin (PRL). PI menghasilkan melanocyte stimulating hormone (MSH) dan β-endorphin (tabel 1), sedangkan PN melepaskan hormon oxytocin dan
vasopressin. Pada dasarnya kedua hormon yang dilepaskan oleh neurohipofise
merupakan hormon yang disekresikan oleh sel neurosekretori hipotalamus yang terdapat di nuklei paraventrikular (PVN) dan nuklei supraoptik (SON), kemudian dialirkan melewati infundibulum menuju PN neurohipofise. Hormon oxytocin dan
vasopressin berada di akson sel neurosekretori dan disimpan di nervus terminal
dan akhirnya dilepaskan ke arteri hipofise anterior sebelum memasuki sirkulasi darah umum (Brown 1994). Hormon-hormon yang disekresikan oleh sel-sel adenohipofise dialirkan melalui vena hipofise ke sirkulasi sistemik menuju organ target seperti gonad, kelenjar adrenal dan tiroid (Fink 2000).
Tabel 1 Jenis sel adenohipofise dan hormon-hormon yang disekresikan
Tipe sel Nama Sel Jenis Hormon
Asidofil Somatotrop Growth hormone (GH) a
Laktotrop Prolaktin (PRL)a
Basofil Tirotrop
Gonadotrop Kortikotrop
Melanotrop
Tiroid stimulating hormon (TSH)a
Luteinizing hormone (LH) dan folikel
stimulating hormon (FSH)a
Adrenokortikotropik hormon (ACTH)a
Melanosit stimulating hormon (MSH)b
Kromofob Sel-sel yang tidak atau memiliki sedikit hormon dan dapat
berbentuk asidofil atau basofila, b
Keterangan: a dihasilkan oleh PD, b dihasilkan oleh PI, a, b dihasilkan oleh PD dan PI
(Sumber: Aron et al. 1997)
Hubungan antara hipotalamus dan adenohipofise pada proses transmisi sinyal diperantarai oleh chemical messengers (neurohormon) yang berperan sebagai faktor pelepas dan penghambat sekresi hormon adenohipofise. Kedua faktor tersebut disintesis oleh sel-sel syaraf di nuklei hipotalamus dan dibawa ke adenohipofise melalui sistem portal hipofise (Fink 2000).
Pars Intermedia Hipofise
Perkembangan pars intermedia
Pada masa perkembangan embrional, PI berasal dari hubungan antara infundibulum dan perluasan adenohipofise (Hadley 1992). Pada anjing beagle, PI terbentuk dari dinding atas dan bagian dorsal dinding anterior kantong Rathke. Bagian ini terlihat jelas saat fetus berumur 38 hari (Sasaki dan Nishioka 1998). Proliferasi sel-sel di bagian kaudal kantong Rathke yang membentuk PI pada periode fetus relatif sedikit bila dibandingkan dengan bagian hipofise lainnya (Lathsaw 1987). PI termasuk bagian adenohipofise yang berhubungan erat dengan PN membentuk lobus neurointermedia dan terpisah dari PD oleh celah hipofise (hypophyseal cleft). Hal ini ditemukan pada beberapa spesies ikan bertulang seperti Cyprinus caprio, Parasilurus asotus dan Fugu rubripes serta pada elasmobrachii (Fujita et al. 1988) dan sebagian besar spesies mamalia termasuk fetus manusia (Bowen 2006).
Menurut Hadley (1992) dan Bowen (2006), pada setiap spesies hewan, PI memiliki ukuran dan letak yang bervariasi. Pada hewan tertentu seperti
cetacea (lumba-lumba dan paus) serta aves, PI tidak ditemukan di hipofise.
Pada manusia, PI berkembang dengan baik pada periode fetus, mengalami rudimenter setelah fetus lahir dan akhirnya menghilang pada saat dewasa. Pada veretebrata tingkat rendah seperti reptil, amfibi dan ikan, PI berkembang dengan baik. Hal ini berhubungan erat dengan kemampuan adaptasi hewan-hewan tersebut dengan lingkungan, terutama kemampuan dalam mengubah warna kulit dan beberapa fungsi fisiologis lainnya.
Vaskularisasi pars intermedia
Menurut Bennet dan Whitehead (1983), vaskularisasi PI relatif lebih sedikit dibandingkan dengan bagian hipofise lainnya. Vaskularisasi berasal dari sistem portal hipofise yang selanjutnya memasuki jaringan parenkim PI. Sistem portal hipofise juga berperan sebagai jalur sekresi produk sel serta faktor regulasi dari dan menuju hipofise.
Struktur pars intermedia
Sel-sel yang menyusun PI terdiri atas tiga jenis sel, yaitu sel-sel basofil (melanotrop dan kortikotrop), kromofob (sel yang sedikit menyerap zat warna) dan sel-sel koloid (colloid-filled cysts). Sel utama yang ditemukan dengan jumlah terbesar di jaringan PI adalah melanotrop yang merupakan sel granular sekretori. Sel tersebut telah ditemukan pada PI hipofise sapi, anjing, kelinci dan tikus. Selain melanotrop, ditemukan juga kortikotrop dalam jumlah sedikit dan sel-sel
folliculo-stellate yang tersebar di jaringan interstitial PI dan mendapat inervasi
syaraf dari neuron yang berasal dari hipotalamus (Bennet dan Whitehead 1983, Bowen 2006). Menurut Fujita et al. (1988), sel-sel endokrin PI memiliki granul sekretori yang mengandung prekursor peptida kortikotropin dan β-lipotropin serta sel-sel imunoreaktif MSH (sel-sel ir-MSH).
Gambar 6 Sel-sel basofil PI (melanotrop dan kortikotrop) hipofise kambing. PI berbatasan langsung dengan PN membentuk lobus neurointermedia (Sumber: Charlotte 2002).
Berdasarkan penelitian yang dilakukan pada hipofise ikan teleost
(Poecilla latipinna), ditemukan dua tipe sel penyusun PI. Sel tipe pertama adalah
sel pars intermedia positive hematoxylin (PIPbH) yang bereaksi positif terhadap hematoksilin, sedangkan sel tipe kedua adalah sel pars intermedia positive
periodic acid Schiff (PIPAS) yang bereaksi positif terhadap periodic acid Schiff.
Sel PIPbH diketahui sebagai sel yang mensekresikan MSH (Hadley 1992). Fungsi pars intermedia
Menurut Tsatmali et al. (2002), melanotrop PI yang tergolong basofilik
(Gambar 6), merupakan sel utama penghasil hormon α-MSH, yaitu hormon yang bertanggung jawab dalam proses pigmentasi kulit dan sebagai regulator yang bekerja pada melanosit kulit dalam memproduksi pigmen melanin. Pada vertebrata tingkat rendah, hormon ini berperan dalam proses perubahan warna kulit saat beradaptasi dengan lingkungan. Pada manusia dewasa, fungsi PI tidak diketahui secara pasti, hal ini disebabkan PI mengalami rudimenter setelah fetus dilahirkan. Namun demikian, PI diduga berperan penting pada masa fetal, yaitu
berhubungan erat dengan pertumbuhan tubuh fetus. Walaupun PI yang merupakan sumber penghasil MSH tidak ditemukan pada manusia dewasa, namun peran MSH sebagai regulator pigmentasi kulit pada melanosit tetap berlangsung. Pars distalis merupakan sumber MSH pada manusia, yang bekerja sama dengan ACTH yang dikenal dengan melanokortin. Hormon inilah yang berperan penting sebagai faktor regulasi terhadap melanosit kulit.
Sekresi α-MSH oleh melanotrop PI dirangsang oleh regulator dari hipotalamus, yaitu melanocyte stimulating hormon releasing factor (MRF) dan dihambat oleh melanocyte stimulating hormone inhibiting factor (MIF). Defisiensi α-MSH menyebabkan kepucatan pada kulit, sedangkan kelebihan hormon
tersebut akan meningkatkan pigmentasi (kehitaman) pada kulit
(Carolla et al. 1990).
Proopiomelanocortin (POMC)
Pada pertengahan tahun 1970, molekul POMC ditemukan di jaringan hipofise dan jaringan syaraf lainnya. Prohormon POMC merupakan molekul peptida berukuran besar, sebagai prekursor dari beberapa hormon peptida penting, yaitu ACTH, MSH dan endorphin (END). Proses perubahan molekul POMC menjadi hormon dengan peptida yang lebih kecil merupakan suatu fenomena spesifik (Gambar 7). Pembentukan ACTH dari POMC berlangsung di PD, sedangkan pembentukan MSH dan END berlangsung di melanotrop PI. Perubahan POMC tidak hanya menghasilkan α-MSH, β-MSH dan variasi γ-MSH, ACTH serta α, β, dan γ-END, tetapi juga menghasilkan corticotropin like
intermediate lobe peptide (CLIP) dan fragmen-fragmen lainnya. α, β dan γ-MSH
secara struktural dibentuk dari ACTH pada melanotrop PI (Hadley 1992; Greidanus et al. 2000).
Secara garis besar, proses perubahan POMC berlangsung dalam dua tahap. Pertama, ACTH dan β-lipotropin (β-LPH) pada PD dan PI yang berasal dari prekursor POMC berhenti membelah. Kedua, seluruh ACTH yang terdapat di PI dikonversikan menjadi α-MSH dan CLIP, sedangkan β-lipotropin dikonversikan menjadi β-endorfin dan γ-lipotropin (Gambar 7) (Brown 1994).
Gambar 7 Proses pemecahan prohormon POMC. α-MSH berasal dari pemecahan rantai asam amino ACTH di PI (Sumber: KUL 2000).
Menurut Gantz dan Fong (2003), pembentukan prohormon POMC berawal dari ekspresi gen POMC yang sebagian besar terdapat di susunan syaraf pusat, nuklei arkuatus di hipotalamus dan nuklei traktus solitarius di batang otak. Selain itu, gen POMC diekspresikan pula oleh melanosit dan keratinosit kulit. Berdasarkan penelitian yang dilakukan pada ikan barfin flounder, ditemukan tiga jenis gen POMC, yaitu POMC-A, B dan C di PD dan PI. Perbedaan jumlah peptida yang berasal dari ketiga gen POMC di PD dan PI hipofise, telah berhasil diidentifikasi. Peptida-peptida yang berasal dari gen POMC-A lebih banyak ditemukan di PD dibandingkan di PI, sedangkan POMC-C ditemukan dalam jumlah sedikit di PD. Adapun jenis peptida yang ditemukan di PD adalah: ACTH-A, des-acetyl (Ac)-A/B (susunan asam amino α-MSH-A dan α-MSH-B identik), β-MSH-α-MSH-A, CLIP-α-MSH-A dan N-terminal peptide (N-POMC)-α-MSH-A. Sedangkan di PI ditemukan peptida-peptida des-Ac-α-MSH A/B, α-MSH A/B, β-MSH A/B, N-β-lipotopin-A, CLIP-A, N-Ac-β-END-A dan N-POMC-A. Secara spesifik, ACTH-A dihasilkan di PD, tetapi proses perubahan des-Ac-α-MSH, CLIP-A dan β-MSH A menjadi peptida yang lebih kecil terjadi di PI (Takahashi et al. 2006).
γ-MSH
1 2 3 4 5 6 7 8 9 10 Try – Val - Met – Glu – His – Phe – Arg – Trp – Asp – Arg
12 11 Gly – Phe
β -MSH
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 Asp – Glu – Gly – Pro – Try – Lys – Met – Glu – His – Phe – Arg – Trp – Gly – Ser –Pro
18 17 16 Asp – Lys – Pro
α-MSH
1 2 3 4 5 6 7 8 9 10 11 12 13
Ac – Ser – Try – Ser - Met – Glu – His – Phe – Arg – Trp – Gly – Lys – Pro – Val – NH2
ACTH
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Ser – Try – Ser - Met – Glu – His – Phe – Arg – Trp – Gly – Lys – Pro – Val – Gly – Lys
32 31 30 29 28 27 26 25 24 23 22 21 20 19 18 17 16
Ala – Ser – Glu – Asp – Glu – Ala – Gly – Asn – Pro – Try – Val – Lys – Val – Pro – Arg – Arg – Lys 33 34 35 36 37 38 39
Glu – Ala – Phe – Pro – Leu – Glu – Phe
CLIP
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 Arg – Pro – Val – Lys – Val – Try – Pro – Asn – Gly – Ala – Glu – Asp – Glu – Ser – Ala – Glu 22 21 20 19 18 17 Phe – Glu – Leu – Pro – Phe – Ala
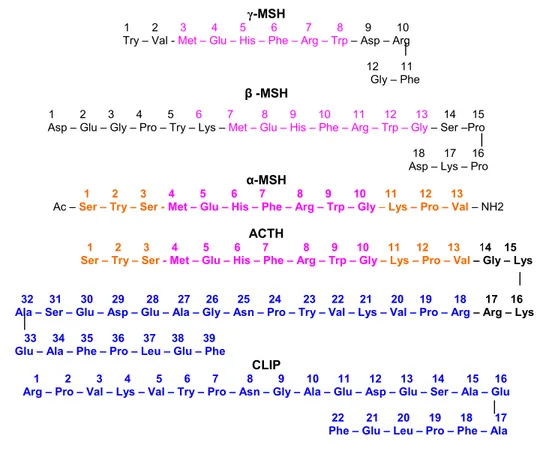
Gambar 8 Susunan rantai asam amino hormon-hormon derivat POMC. γ-MSH, β-MSH, α-MSH, ACTH dan CLIP. Rantai asam amino 1-13 α-MSH dan ACTH identik (Sumber: Hadley 1992).
Melanosit Stimulating Hormon (MSH)
Terdapat tiga jenis MSH yang berhasil diisolasi dari ekstrak jaringan hipofise, yaitu α-MSH (13 asam amino), β-MSH (18 asam amino) dan γ-MSH (12 asam amino). PI hipofise terutama menghasilkan α-MSH, sedangkan β-MSH dan γ-MSH dihasilkan di PD dan di beberapa lokasi jaringan tubuh lainnya. Ketiga jenis hormon tersebut berasal dari pemecahan prohormon POMC yang memiliki fungsi spesifik (Bicknel 2002).
Carola et al. (1990) menyatakan bahwa, pada manusia kehadiran MSH secara tidak langsung meningkatkan aktivitas melanosit, yaitu sel yang menentukan warna kulit, mata dan rambut. α-MSH yang sebagian besar berasal dari PI, dihasilkan melalui stimulasi ACTH. Secara normal, jumlah α-MSH yang disekresikan tidak cukup untuk merangsang melanosit. Oleh karena itu diperlukan kombinasi ACTH dan α-MSH dalam mengatur fungsi melanosit pada
proses pigmentasi kulit. Menurut Tsatmali et al. (2002), walaupun PI pada manusia menghilang setelah dilahirkan, namun α-MSH masih dihasilkan oleh jaringan tubuh lainnya, terutama kulit. Proses sintesis hormon α-MSH di kulit, tetap dimulai dari pemecahan POMC yang terjadi pada melanosit dan keratinosit kulit. Selain itu, sel-sel Langerhans kulit juga menghasilkan α-MSH dalam jumlah yang lebih rendah.
Pada hewan-hewan mamalia, karakteristik kimiawi α-MSH memiliki struktur identik pada susunan asam aminonya, tetapi sedikit berbeda pada hewan poikiloterem. Perbedaan α-MSH antara ikan hiu (Squallus acanthias) dan ikan salmon (Onchorhychus keta) terletak pada rantai asam amino, yaitu pada N-terminal serin yang tidak terasetilasi. Sedangkan pada hiu kodok, asam amino valin seperti yang terdapat pada α-MSH manusia, posisinya diganti dengan metionin. Jenis MSH lainnya yaitu β-MSH, pada manusia hormon ini memiliki rantai asam amino berjumlah 22, lebih panjang dibandingkan α-MSH. Sedangkan pada mamalia lainnya, β-MSH memiliki rantai asam amino yang lebih pendek, yaitu 18 buah (Gambar 8) (Hadley 1992). Fungsi β-MSH pada manusia, berhubungan erat dengan fungsi ACTH, yaitu pada proses fisiologis tubuh (Greidanus et al. 2000).
Menurut Donohue dan Jacobowitz (1980), melalui teknik imunofluorosen, diketahui bahwa jaringan otak tikus juga menghasilkan α-MSH. Hormon ini ditemukan pada nuklei arkuatus dan varikositas serabut syaraf yang menyebar di bagian batang otak. Selain itu, serabut syaraf yang mengandung α-MSH juga terdapat pada beberapa nuklei di hipotalamus, area preoptik, septum, amigdala, korpus mamilaris dan substansi abu-abu. Namun demikian, konsentrasi α-MSH yang lebih tinggi ditemukan di median eminens, medial preoptik, hipotalamus anterior, nuklei arkuatus, nuklei periventrikular dan nuklei paraventrikular. Penyebaran α-MSH di otak, diduga lebih berperan sebagai neurotransmiter dari pada neuromodulator. Hal ini menunjukkan adanya interaksi antara α-MSH dengan sistem neuronal di otak.
Pada manusia, α-MSH dihasilkan oleh melanosit kulit disamping produk
lokal lainnya. Proses pembentukan hormon ini tidak terlepas dari pemecahan prohormon POMC yang dihasilkan melanosit pada lapisan epidermis kulit. Proses pemecahan prohormon POMC sebagian besar berlangsung di melanosit,
dan sebagian kecil di keratinosit dan sel-sel Langerhans kulit
Pengaturan Sintesis dan Sekresi MSH
Dopamin merupakan senyawa yang mirip dengan katekolamin yang berperan dalam mengontrol sekresi MSH. Senyawa ini telah berhasil diisolasi dari lobus neurointermedia (PI dan neurohipofise) bersama-sama dengan norepinefrin dan epinefrin. Dopamin dianggap sebagai senyawa yang mampu menghambat sekresi MSH oleh hipofise. Senyawa ini dilepaskan dari neuron dopaminergik yang berasal dari nukleus arkuatus menuju PI melalui tangkai hipofise (Fink 2000). Selanjutnya dopamin berinteraksi dengan reseptor dopaminergik yang terletak di membran melanotrop PI, konsentrasi dopamin yang tinggi mengakibatkan melanotrop berada pada posisi hiperpolar. Keadaan ini akhirnya menghambat sekresi MSH oleh melanotrop. Selain mengontrol sekresi MSH, dopamin juga berperan pada proses sekresi hormon derivat POMC lainnya, yaitu ACTH, γ-LPH, β dan γ-endorfin (Hadley 1992).
Pada vertebrata tingkat rendah, kontrol dopamin terhadap sekresi MSH berhubungan dengan fungsi mata dalam menerima rangsangan cahaya dari lingkungan. Rangsangan ini diteruskan ke hipotalamus oleh sistem syaraf, dan selanjutnya akan ditentukan apakah terjadi penghambatan atau peningkatan sekresi MSH oleh sel-melanotrop PI (Turner dan Bagnara 1976).
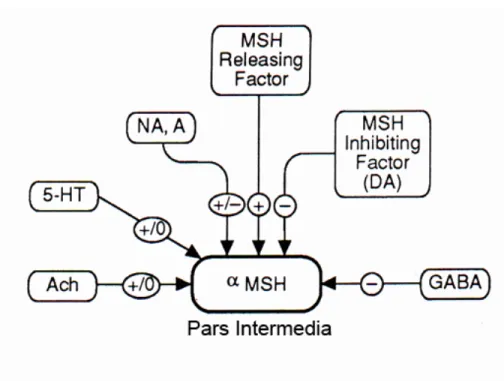
Menurut Brown (1994), beberapa neurotransmiter lain juga berperan sebagai faktor penghambat dan pelepas MSH. Senyawa tersebut adalah GABA sebagai faktor penghambat; noradrenalin (NA) dan adrenalin (A) sebagai faktor penghambat dan pelepas; sedangkan asetilkolin dan serotonin (5 HT) diduga berperan sebagai faktor pelepas, namun perannya terbatas. Dopamin dan beberapa neurotransmiter tersebut bekerja langsung pada melanotrop PI (Gambar 9).
Pada bagian lain, PI juga mendapat inervasi langsung dari neuron sekretori (neuron peptidergik) yang berasal dari nukleus paraventrikularis (PVN) hipotalamus. Aktivitas hambatan dan peningkatan sekresi MSH dilakukan oleh MIF dan MRF. Kedua faktor ini diduga merupakan derivat oksitosin (c-terminal tripeptida dari oksitosin), yaitu Pro-Leu-Gly-NH2 yang menunjukkan aktivitas MIF, sedangkan H-Cys-Tyr-Ile-Gly-Asn-OH berperan sebagai MRF (Bennet dan Whitehead 1983).
Gambar 9 Pengaturan neurotransmiter pada sintesis dan sekresi MSH. Faktor pelepas terdiri atas: MRF, NA, 5 HT, Ach; factor penghambat terdiri atas: MIF, A dan GABA. NA= noradrenalin A= adrenalin, 5 HT= 5-hidroksitriptamin (serotonin), Ach= asetilkolin, GABA= gamma amino butyric acid, sekresi dirangsang (+), sekresi dihambat (-), peran terbatas (0) (Sumber: Brown 1994).
Beberapa faktor lingkungan yang berpengaruh terhadap peningkatan
dan penghambatan sekresi MSH telah ditemukan pada vertebrata tingkat rendah dan mamalia. Pada amfibi dan ikan, kondisi stres akan merangsang dan meningkatkan sekresi MSH dari PI, sedangkan intensitas cahaya yang tinggi akan menghambat sekresi MSH. Pada peristiwa ini, terdapat faktor lain yang ikut berperan, yaitu melatonin, hormon yang dihasilkan oleh kelenjar pineal (epifise). Senyawa ini memiliki peran antagonis terhadap MSH (Bennet dan Whitehead
1976; Fujita et al. 1988).
Faktor penghambat sekresi MSH lainnya adalah neuropeptida Y (NPY) yang disintesis oleh nuklei arkuatus dan median eminens yang berfungsi sebagai molekul neurotransmiter. Selanjutnya senyawa ini disekresikan oleh akson terminal nuklei arkuatus ke nuklei paraventrikular (PVN) dan bagian otak lainnya (Greidanus et al. 2000). Fungsi NPY sebagai faktor penghambat sekresi MSH telah ditemukan pada katak bercakar Afrika (Xenopus leavis). Senyawa ini terdapat pada nuklei suprasiasmatik di hipotalamus yang dikenal dengan
suprachiasmatic melanotrope inhibiting neurons (SMINs). Selain menghasilkan
NPY, SMINs juga menghasilkan dopamin dan GABA (Kramer et al. 2002). Berbeda dengan aksi neurotransmiter lainnya (dopamin, GABA) yang bekerja langsung pada melanotrop PI, NPY bekerja secara tidak langsung. Selain melanotrop, PI juga memiliki sel folliculo-stellate yang berhungan erat dengan fungsi melanotrop. Sel folliculo-stellate ini memiliki reseptor khusus terhadap NPY. Melalui reseptor inilah NPY bekerja secara tidak langsung untuk menghambat sekresi MSH dari melanotrop (Hadley 1992).
Fungsi Fisiologis MSH
Proses pigmentasi kulit pada sebagian besar vertebrata merupakan fungsi utama MSH disamping proses fisiologis lainnya. Pada kulit mamalia dan vertebrata lainnya, sel penghasil pigmen (melanosit, melanofor) terletak di
stratum basale epidermis (Turner dan Bagnara 1976). Produk utama melanosit
adalah melanin, yaitu pigmen endogenous yang didistribusikan pada jaringan tubuh hewan dengan konsentrasi berbeda. Komponen utama penyusun pigmen melanin adalah dihydroxyindoxylic acid. Secara kimia, melanin tidak identik dengan neurotransmiter dan hormon, tetapi memiliki prekursor yang sama dengan prekursor katekolamin, yaitu dopamin (Fujita et al. 1988).
Proses pembentukan pigmen melanin (melanogenesis), diawali dengan bergabungnya melanin dengan struktur subseluler yang disebut premelanosom. Apabila struktur ini telah dipenuhi oleh melanin, maka premelanosom berubah menjadi melanosom (granul melanin) (Hadley 1992). Melanin terdiri atas
eumelanin yang membawa warna coklat dan hitam dan phaeomelanin yang
membawa warna merah. Hewan yang memiliki variasi warna kulit menghasilkan satu atau beberapa jenis melanin pada lokasi kulit yang berbeda (Turner dan Bagnara 1976).
Pada rambut dan bulu hewan, melanosit terletak di dalam folikel rambut untuk mensintesis melanin dan bertanggung jawab pada proses pigmentasi rambut dan bulu. MSH dan hormon steroid berperan dalam produksi eumelanin dan phaeomelanin pada melanosit. Walaupun peran MSH dalam poses pigmentasi melanin pada kulit dan rambut pada sebagian mamalia telah terbukti, namun perannya dalam pigmentasi bulu burung (aves) belum diketahui dengan jelas. Hal ini diduga berkaitan dengan absennya PI hipofise pada aves (Kent dan Carr 2001).
Sintesis hormon-hormon yang berasal dari POMC terjadi pada tahap
yang berbeda. MSH dan CLIP ditemukan dalam konsentrasi yang tinggi pada periode fetus manusia dan beberapa spesies primata. Konsentrasi ini menurun pada periode postnatal dan akhirnya menghilang pada waktu dewasa. Kehadiran hormon-hormon tersebut berhubungan erat dengan aspek fisiologis tubuh fetus, seperti perkembangan hubungan kelenjar hipofisa dan kelenjar adrenal (Hadley 1992).
Kortek adrenal manusia memiliki zona fetus yang luas selama masa prenatal dan menghilang pada masa postnatal, sebaliknya zona kortek mengalami hipertropi dan berkembang sebagai zona kortek definitif. Hal ini membuktikan bahwa zona fetus adrenal lebih responsif terhadap α-MSH dibandingkan dengan ACTH, sedangkan zona kortek adrenal lebih responsif terhadap ACTH. Selain itu, α-MSH dan ACTH dibawah pengaruh corticotropin
releasing hormone (CRH) diduga berperan pada respon fisiologis fetus terhadap
stres (Hadley 1992).
Pada proses pertumbuhan dan perkembangan fetus, α-MSH lebih
berperan dibandingkan hormon koriogonadotropin dan oksitosin. Berdasarkan penelitian pada anak yang mengalami craniofaringioma, yaitu suatu gangguan dimana somatotropin hormone (STH)/growth hormone (GH) gagal disekresikan, konsentrasi STH meningkat secara signifikan setelah pemberian α-MSH. Hal ini disebabkan STH dan α-MSH berasal dari nukleus arkuatus di hipotalamus, sehingga dalam kondisi ini α-MSH bertindak sebagai STH releasing hormon. Dengan fungsi tersebut, α-MSH disebut juga sebagai growth promoting pada periode kehidupan fetus (Hadley 1992).
Dari beberapa penelitian yang berhubungan dengan sekresi aldosteron,
ditemukan adanya korelasi antara melanotropin (MSH) dan aldosteron. Jenis melanotropin tersebut adalah β-MSH yang secara langsung merangsang sekresi aldosteron dan bekerja pada reseptor adrenocortical. Hormon MSH lainnya, yaitu γ-MSH dan α-MSH memiliki kemampuan yang sama dengan β-MSH dalam mengontrol zona glomerulosa adrenal dalam menghasilkan hormon aldosteron (Hadley 1992; Bicknell 2002).
Melanotropin mempengaruhi aktivitas susunan syaraf pusat (SSP) pada
manusia dan hewan laboratorium. Efek tersebut berupa sikap bangkit (bangun), peningkatan motivasi, konsentrasi, memori dan learning (Greidanus et al. 2000). Hal ini dibuktikan melalui pemeriksaan imunositokimia yang memperlihatkan
aktivitas α-MSH pada neuron spesifik di otak. Potensi aksi melanotropin terhadap SSP semakin meningkat bila hormon ini diinjeksikan langsung ke dalam ruang ventrikel otak. Dapat disimpulkan, α-MSH dan melanotropin lainnya juga bertindak sebagai neurotransmiter dan neuromodulator di otak (Hadley 1992; Brown 1994).
Peran melanotropin sebagai termoregulator ditemukan pada kadal
Arizona (Urosaurus ornatus) yang mampu mengubah warna kulit menjadi gelap akibat peningkatan pencahayaan dan kenaikan temperatur lingkungan. Hal ini berkaitan dengan kemampuan melanosit kulit beradaptasi pada kondisi tersebut dan kemampuan dalam menyerap energi. Proses ini berlangsung dibawah kontrol α-MSH. Pada penelitian lebih lanjut, secara imunositokimia, α-MSH dan ACTH ditemukan pada area preoptik hipotalamus anterior yang merupakan pusat regulasi suhu tubuh (Samson et al 1980; Chang 1997).
Struktur dan Fungsi Kulit
Kulit merupakan organ tubuh terbesar yang berfungsi sebagai barrier biologis dalam mempertahankan kondisi homeostasis internal dari pengaruh lingkungan eksternal. Beberapa kondisi lingkungan yang mempengaruhi homeostasis kulit adalah radiasi sinar ultra violet (UV), temperatur, energi mekanis, agen biologis dan kimia serta kelembaban udara (Slominski dan Wortsman 2000).
Kulit terdiri atas dua bagian utama, yaitu epidermis yang tersusun atas struktur epitelial dan dermis tanpa struktur epitelial (Gambar 10). Lapisan subkutan terletak di profundal lapisan dermis. Pada tubuh hewan, struktur dan ketebalan epidermis dan dermis bervariasi, sesuai dengan lokasi anatomis tubuh. Ketebalan lapisan epidermis berkisar antara 0.1-1.6 mm (Slominski dan Wortsman 2000). Sel-sel penyusun epidermis terdiri atas keratinosit, melanosit yang disebut unit epidermis kulit dan sel Langerhans (Gambar 11), sedangkan dermis terdiri atas jaringan ikat kolagen dan elastin, fibroblast, makrofag, jaringan adipose, syaraf, folikel rambut, pembuluh darah dan kelenjar eksokrin kulit. Kelenjar tersebut adalah kelenjar sebaseus, kelenjar apokrin (keringat) dan kelenjar serominus. Rambut yang merupakan bagian dari kulit, terdiri atas kolum (bagian yang mengandung keratin), batang rambut yang menjulur dari kulit dan akar rambut yang terdapat di bagian dermis (Anonim 2006).
Gambar 10 Struktur umum kulit. Secara umum terdiri atas lapisan epidermis, dermis dan hipodermis (Sumber: Anonim 2000).
Melanosit
Melanosit pada vertebrata berasal dari neural crest lapisan ektoderm umum. Selanjutnya melanosit yang masih berbentuk melanoblas bermigrasi dan berdiferensiasi menjadi melanosit pada lapisan epidermis kulit dan rambut serta jaringan tubuh lainnya. Pada manusia, melanosit tersebar di stratum basale epidermis, rambut, retina, iris, pleksus koroideus mata dan leptomeningen yang menyelubungi susunan syaraf pusat, membrana labirin telinga, substansia nigra dan lokus seruleus batang otak. Melanosit pada hewan poikiloterem diinervasi oleh syaraf otonom, yaitu melanin aggregating nervus dan melanin dispersing
nervus. Sedangkan melanosit pada epidermis manusia tidak diinervasi oleh
syaraf otonom, sehingga sel tersebut tidak sensitif terhadap hormon spesifik (Fujita et al. 1980).
Sebagai organ terbesar dan multifungsi, kulit manusia menghasilkan pigmen melanin dan substansi lainnya yang berhubungan dengan fungsi hipotalamus, hipofise dan adrenal. Substansi tersebut terdiri atas CRH, urokortin dan POMC. Pemecahan POMC di melanosit kulit menghasilkan ACTH, α-MSH dan β-endorfin. Selain hormon, kulit juga menghasilkan vitamin D, katekolamin
dan asetilkolin yang berfungsi sebagai respon kulit terhadap stres yang berasal dari lingkungan. Sistem neuroendokrin kulit saling berhubungan melalui jalur humoral dan neuronal untuk merangsang sistem vaskular, imun atau perubahan pigmen untuk menetralisir agen-agen berbahaya, sehingga sistem neuroendokrin kulit berperan penting dalam memelihara struktur dan integritas fungsional kulit (Slominski dan Wortsman 2000; Kauser et al. 2005).
Hubungan PI dan Melanosit Kulit
Secara fungsional, hubungan PI dengan melanosit kulit beragam pada berbagai spesies hewan dan manusia, yang disebabkan adanya variasi perkembangan dan fungsi PI hipofise. Pada vertebrata tingkat rendah (reptil, amfibi dan ikan), hubungan PI dan melanosit terlihat jelas. Aktivitas melanosit kulit berupa perubahan warna kulit dapat berlangsung dengan peran α-MSH yang disekresikan PI (Fujita et al. 1988). Sedangkan pada manusia dewasa, hubungan tersebut tidak erat, namun pada masa prenatal, PI fetus berkembang dengan baik dalam menghasilkan α-MSH yang berperan sebagai growth
promoting yang berhubungan erat dengan sintesis growth hormone dalam
proses pertumbuhan tubuh fetus (Hadley 1992). Menurut Carola et al. (1990), dengan menghilangnya PI hipofise pada manusia dewasa, sekresi α-MSH yang berasal dari hipofise tidak cukup untuk merangsang aktivitas melanosit kulit, oleh karena itu diperlukan adanya kombinasi antara ACTH dan α-MSH dalam mengatur fungsi melanosit kulit.
Proses sintesis α-MSH dan ACTH di melanosit dan keratinosit kulit menurut Gantz dan Fong (2003), diawali dari radiasi sinar UV yang diterima oleh reseptor kedua hormon tersebut yang terdapat di keratinosit. Pada kondisi ini, keratinosit yang mensintesis ACTH dan α-MSH bersifat parakrin, yaitu merangsang melanosit untuk menghasilkan pigmen melanin. Sedangkan melanosit yang juga mensintesis ACTH dan α-MSH bersifat otokrin dalam menghasilkan pigmen melanin serta bersifat parakrin dalam melindungi sel tersebut dari gangguan sistem imunitas kulit. Sejauh mana kontribusi α-MSH yang berasal dari PI hipofise pada proses pigmentasi kulit manusia, secara pasti belum diketahui. Hal ini disebabkan rendahnya konsentrasi α-MSH di dalam aliran darah dan serum ACTH.
Menurut Slominski dan Wortsman (2000), kemampuan kulit melalui melanosit, keratinosit dan Langerhans dalam menghasilkan produk neuroendokrin yang sama dengan hipofise dan adrenal, disebabkan ketiga jaringan tersebut berasal dari sumber yang sama pada masa embrional. Jaringan hipofise, adrenal dan kulit berasal dari sel-sel epitel luar ektoderm. Proses fisiologis yang melibatkan peran hipofise, kelenjar adrenal dan jaringan kulit pada proses pigmentasi merupakan proses yang kompleks. Proses tersebut diawali dengan adanya stimulus dari lingkungan yang diterima lapisan epidermis kulit. Rangsangan diteruskan ke susunan syaraf pusat berupa sinyal neuronal dan sinyal humoral untuk menghasilkan produk neuroendokrin yang mengontrol fungsi kulit.
Gambar 11 Lapisan epidermis kulit dan bagian-bagiannya. Melanosit dengan granul melanosom terletak di membran basal (stratum basale), keratinosit di lapisan superfisial berhubungan erat dengan melanosit (Sumber: Anonim 2006).
Pembentukan Pigmen Melanin
Pigmen melanin merupakan produk akhir dari proses melanogenesis, yaitu suatu proses yang sangat kompleks, melibatkan berbagai komponen yang memiliki fungsi regulator, reseptor dan sekresi produk hormonal, dan neurotransmitter. Proses tersebut berlangsung di melanosit yang terletak pada
epidermis, yaitu di bagian stratum basale. Pigmen melanin berperan penting pada proses kamuflase, mimikri, komunikasi sosial dan perlindungan kulit dari radiasi sinar UV yang mempengaruhi homeostasis kulit (Slominski et al. 2003).
Melanogenesis diawali dengan terjadinya ikatan antara α-MSH dan ACTH (melanokortin) dengan reseptor MC-1R pada membran melanosit. Ikatan yang diperantarai oleh protein G tersebut mengaktifkan adenilat siklase (AC), senyawa ini selanjutnya meningkatkan c-AMP intrasel yang berhubungan dengan enzim tirosinase. Tirosinase mengubah tirosin menjadi L-dihidroksifenilalanin (L-DOPA). Tahap selanjutnya adalah oksidasi L-DOPA menjadi dopakuinon, yang merupakan prekursor melanin, baik eumelanin maupun phaeomelanin (Gambar 12) (Slominski et al. 2003). Pigmen melanin yang dihasilkan melanosit, selanjutnya disekresikan ke keratinosit melalui granul-granul melanin yang disebut melanosom. Formasi, maturasi dan perjalanan melanosom menuju keratinosit merupakan tahapan penting dalam proses pigmentasi kulit. Gangguan pada sebagian atau keseluruhan dari tahapan tersebut akan mengakibatkan kegagalan proses pigmentasi, seperti pada Hermansky-Pudlak Syndrome (HPS) yang mempelihatkan penurunan derajat pigmentasi pada kulit, rambut dan mata,
nystagmus dan fotofobia (Lin dan Fisher 2007).
Gambar 12 Proses melanogenesis di melanosit. Reseptor MSH di membran melanosit, berikatan dengan MSH dan AC yang diperantarai oleh Gs, menghasilkan cAMP dan PKA. PKA meningkatkan sintesis dan aktivitas tirosinase yang berperan dalam melanogenesis di granul melanosom. Granul melanosom ditransfer ke keratinosit melalui dendrit melanosit. Gs= protein G, AC= adenilat siklase, cAMP= adenosin monofosfat siklik, PKA= protein kinase (Sumber: Chang 2001).
Perjalanan melanosom yang mengandung melanin dari melanosit ke keratinosit tidak berlangsung secara eksositosis, tetapi menyerupai aliran akson pada sel syaraf yang disebut cytocrinia. Pada manusia dengan ras kulit berwarna hitam, pigmen melanin tersebar di ruang ekstra seluler keratinosit, namun demikian proses sintesis pigmen tersebut tidak berlangsung di keratinosit (Fujita et al. 1980). Penyebaran melanosom di keratinosit terjadi apabila kulit menerima rangsangan sinar UV, sehingga melanosom menyebar mengelilingi inti keratinosit (Lin dan Fisher 2007). Menurut Kauser et al. (2005), proses sintesis pigmen melanin tidak hanya berlangsung di melanosit, namun juga berlangsung di folikel rambut. Proses tersebut diregulasi oleh derivat POMC, yaitu α-MSH, ACTH dan β-endorfin.