Oseana, Volume XX, Nomor 2, 1995 : 13 – 23 ISSN 0216 – 1877
STRUKTUR DAN DINAMIKA ORGAN REPRODUKSI HOLOTHURIA
FORSKALI (ECHINODERMATA : ASPIDOCHIROTA) oleh
Ambo Tuwo *)
ABSTRACT
STRUCTURES AND DYNAMICS OF THE REPRODUCTIVE ORGAN
OF Holothuria forskali (ECHINODERMATA : ASPIDOCHIROTA). The reproduce-
tive organ of H. forskali consists of several size classes of tubules. The smallest contains the oogony or spermatogony, and the largest contains the oocyte or spermatozoa. All tubules are composed of three layers, namely peritoneum, connective tissue and inner epithelium. Gametogenesis of H. forskali is a long process. It begins in small tubules in April. In October, the large tubules contain
oocytes with a modal diameter of 140 - 150 um, and the large testicular tubules
contain abundance of spermatozoas and little spermatogony. Just before spawning, the ovarian tubules contain only oocytes and only spermatozoas in the testicular tubules. During spawning, in April, the large tubules are transformed into spent tubules. Following spawning, the predominant activity within the spent tubules is phagocytosis of the residual gametes.
PENDAHULUAN
Teripang ditangkap sejak ribuan tahun yang lalu oleh nelayan cina. Penangkapan secara besar-besaran saat itu menyebabkan penurunan stok, sehingga mereka terpaksa pergi ke India, Filipina dan Indonesia untuk mencari teripang.
Harga teripang yang tinggi mendorong nelayan kita untuk menangkapnya dalam jumlah besar. Hal ini menempatkan Indonesia pada posisi teratas dari negara-negara
penghasil teripang di dunia (TUWO & CONAND 1992a). Posisi ini sulit dipertahankan pada masa yang akan datang sebab sejak tahun 1989 sudah terlihat adanya kecenderungan penurunan produksi (TUWO & CONAND, 1992a; TUWO & NESSA 1991). Hal ini wajar saja karena alam mempunyai keterbatasan.
Budidaya teripang merupakan salah satu cara yang dapat ditempuh untuk meningkatkan produksi teripang, namun masih banyak masalah yang harus dihadapi,
diantaranya adalah kekurangan benih. Benih alam jauh dari cukup untuk dapat memenuhi kebutuhan benih pada usaha budidaya teripang. Penggadaan benih pada panti-panti pembenihan juga masih pada tahap uji coba. Masih banyak masalah yang harus diselesaikan sebelum sampai pada produksi masal benih teripang.
Salah satu aspek penting yang harus diperhatikan di dalam usaha pembenihan ada-lah biologi reproduksinya. Pemahaman yang baik akan aspek ini dapat membantu kita da-lam hal pengadaan dan perlakuan induk di balai benih. Pengadaan dan perlakuan induk yang kurang tepat dapat menyebabkan rendahnya kualitas telur dan larva sehingga tidak dapat melewati fase metamorfosis yang sangat kritis bagi hewan bentik seperti teripang.
Akan diuraikan struktur dan dinamika organ reproduksi jenis teripang dari bangsa Aspidochirota, yaitu H. forskali. Meskipun ia merupakan jenis teripang yang hidup di daerah subtropis, namun aspek biologi reproduksi-nya, terutama struktur dan dinamikareproduksi-nya, sama dengan jenis teripang dari bangsa Aspidochirota yang ada di Indonesia. Telah diketahui bahwa teripang yang bernilai ekonomi tinggi yang terdapat di Indonesia umumnya berasal dari bangsa Aspidochirota (TUWO & NESSA 1991).
Penelitian ini diharapkan dapat memberikan sumbangan yang berarti bagi para peneliti yang sedang bekerja keras mengembangkan usaha pembenihan teripang.
BAHAN DAN METODA
Pengambilan contoh sebanyak 25 ekor dilakukan setiap bulan selama dua tahun atau dua siklus reproduksi di perairan pulau Panfret, Finister, Perancis.
Setelah hewan dibelah, sebagian go-nad diambil dan difiksasi dengan menggunakan larutan Bouin alkohol selama kita-kira seminggu; setelah itu gonad dicuci dan didehidrasi dengan menggunakan alkohol yang persentasinya dinaikkan secara bertahap, yaitu 50%, 70%, 95% dan 100%, masing-masing sebanyak tiga kali. Setelah selesai maka gonad di pak dengan menggunakan amil asetat sebagai larutan perantara untuk mempermudah penetrasi lilin. Setelah pengepakan selesai, maka gonad selanjutnya dipotong setebal 6 um diwarnai dengan pewarna Schiff dan Trikrom Masson.
HASIL
STRUKTUR ORGAN REPRODUKSI
Berdasarkan organ reproduksinya, H. forskali dapat dibedakan antara hewan jantan dan betina. Organ reproduksinya terdiri dari sebuah dasar gonad dimana semua jenis tabung gonad melekat. Tabung ini dapat dibedakan atas : tabung primer (Tl). tabung sekunder (T2), tabung pematangan (T3) tabung matang (T4) dan tabung terabsorpsi (T5). Tabung terabsorpsi adalah tabung matang yang telah mengecil karena kehilangan isinya. Tabung ini segera diserap setelah pemijahan selesai (Gambar 1).
Struktur Dasar Gonad
Dasar gonad H. forskali berukuran sekitar 5 mm (Gambar 1). Semua leher tabung (Gambar 2.1 dan 2.2) mengarah ke suatu saluran pada dasar gonad yang disebut gonoduk atau saluran telur yang berfungsi menyalurkan telur pada saat pemijahan. Dinding dasar gonad dapat dibedakan atas tiga lapis, yaitu : peritoneum, konektif dan epitelium dalam.
Struktur Tabung Gonad
Seperti halnya dengan dasar gonad, tabung dinding gonad juga terdiri dari tiga lapis, yaitu : peritoneum, konektif dan epitelium dalam.
Peritoneum (EP) yang merupakan lapisan teriuar dari dinding tabung tersusun dari sel kulit berkadar air rendah. Ada bagian yang menyerap pewarna lebih banyak dan membentuk bulatan kecil (Gambar 3.1 dan 3.2). Epitelium dalam (El) terdiri dari sel parietal, sel folikuler dan telur atau sperma. Sel parietal dan folikuler merupakan lapisan lendir. Konektif (C) terletak di antara perito-neum dan epitelium dalam. Konektif pada
daerah lipatan longitudinal (RL) sebagian besar dipenuhi oleh hemal sinus (Gambar 3.1 dan 3.2).
EVOLUSI ORGAN REPRODUKSI
Pada H. forskali, sebuah tabung akan mengalami evolusi yang berawal dari tabung primer dan berakhir pada tabung terabsorpsi.
Tabung Primer (Tl)
Tabung primer dibentuk pada bagian depan dasar gonad (Gambar 1). Pembentukan tabung primer berlangsung beberapa bulan sebelum pemijahan berlangsung. Pada bulan
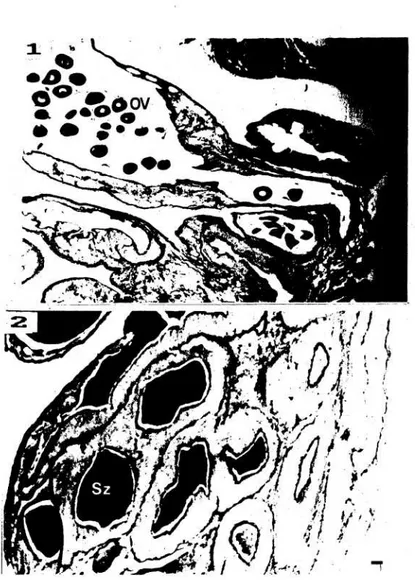
Gambar 2. Struktur dasar gonad Holothuria forskali.
1 dan 2 : dasar tabung hewan jantan dan betina; OV : telur matang; Sz : spermatozoa. Panjang garis : 50 um. Januari, tabung primer berukuran panjang
sekitar 10 mm dan berat kurang dari 0,01 gram. Tabung ini belum memperlihatkan adanya percabangan.
Tabung Sekunder (T2).
Pada musim pemijahan, awal bulan mei, tabung primer telah berkembang menjadi
tabung sekunder. Tabung sekunder ini panjangnya sekitar 2 cm dan berdiameter kurang dari 0,3 mm. Tabung ini sudah bercabang dua sampai tiga kali (Gambar 1). Berdasarkan tahap perkembangannya, tabung ini dapat dibedakan atas dua, yaitu : tabung sekunder awal (3.1 dan 3.2) dan lanjut (3.3 dan 3.4).
Tabung Sekunder Awal Tabung Sekunder Lanjut Pada hewan betina dijumpai telur yang
berukuran 10 sampai 20 um (Gambar 3.1). Bintik hitam yang dijumpai pada telur menunjukkan awal dari proses pembentukan kuning telur atau pravitelogenesis (Opv) (Gambar 3.1). Sedangkan pada hewan jantan, dijumpai spermatosit yang berukuran 5 sampai 10 um (Gambar 3.2). Lipatan longitudinal (RL) terlihat dengan jelas (3.1 dan 3.2).
Pada hewan betina, telur sudah masuk ke fase aktif pembentukan kuning telur atau fase vitelogensis. Telur vitelogenesis (OV) pada tabung ini berukuran sekitar 30 um (Gambar 3.3). Pada hewan jantan, ada bebe-rapa spermatosit (Sc) yang sudah berkembang menjadi spermatozoa (Sz) (Gambar 3.4).
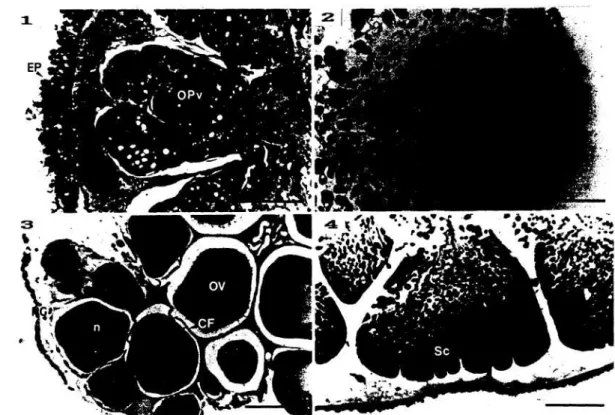
Gambar 3. Struktur tabung sekunder (T2) Holothuria forskali.
1 dan 2 : tabung sekunder awal hewan betina dan jantan; 3 dan 4 : tabung sekunder akhir pada hewan betina dan jantan; C : konektif; CF : sel folokelur; EI: epitelium dalam; EP : pertoneum; MP : otot; n : nukleolus; N : nukleus; OPV : telur pravitelogenesis; OV : telur vitelogenesis; PG : dinding tabung; RL : lipatan longitudinal; Sc : Spermatosit; Sz : spermatozoa. Panjanggaris : 50 um.
Tabung Pematangan (T3)
Tabung ini panjangnya dapat mencapai 25 mm dan berdiameter 2 mm. Sebuah ta-bung dapat bercabang sampai enam kali (Gambar 1). Berdasarkan tahap perkem-bangannya, jenis tabung ini dapat dibedakan atas dua yaitu awal dan akhir pematangan.
Tabung pada Awal Pematangan
Pada hewan betina, pembentukan kuning telur berlangsung sangat cepat. Diam-eter telur sudah ada yang mencapai 100 um
(Gambar 4.1). Pada hewan jantan, spermato-zoa sudah mulai melimpah (Gambar 4.2)
Tabung pada Akhir Pematangan
Pada hewan betina, pembentukan kuning telur terus berlangsung. Diameter telur sudah ada yang mencapai 150 um (Gambar 4.3). Di dekat dinding tabung masih dijumpai beberapa telur pra-vitelogenesis. Pada hewan jantan, spermatozoa seolah-olah membentuk gerakan melingkar di dalam tabung. Spermatosit masih dijumpai di sekitar dinding tabung (Gambar 4.4).
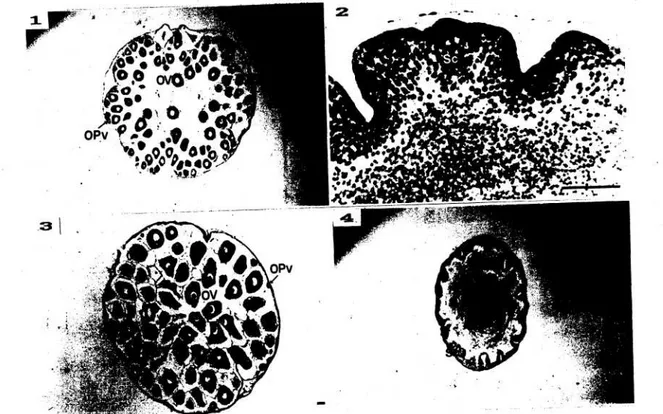
Gambar 4. Struktur tabung pematangan (T3) Holothuria forskali.
1 dan 2: tabung pematangan awal pada hewan betina dan jantan; 3 dan 4: tabung pematangan akhir pada hewan betina dan jantan; OPv: telur pra-vitelogenesis; OV : telur vitelogenesis; Sc : spermatosit; Sz : spermatozoa.
Tabung Matang (T4)
Pada hewan betina, proses vitelogenesis telah selesai dan beberapa telur mulai lepas dari sel folikulernya (Gambar 5.1). Pada he-wan jantan, spermatogenesis juga telah selesai. Spermatozoa memenuhi tabung (Gambar 5.2).
Tabung terabsorpsi (T5)
Setelah pemijahan selesai, telur (OA) dan spermatozoa (SA) yang tidak dipijahkan segera diabsorpsi atau diserap (Gambar 5.3 dan 5.4). Setelah itu, tabung juga akan terserap hingga pada akhirnya hilang.
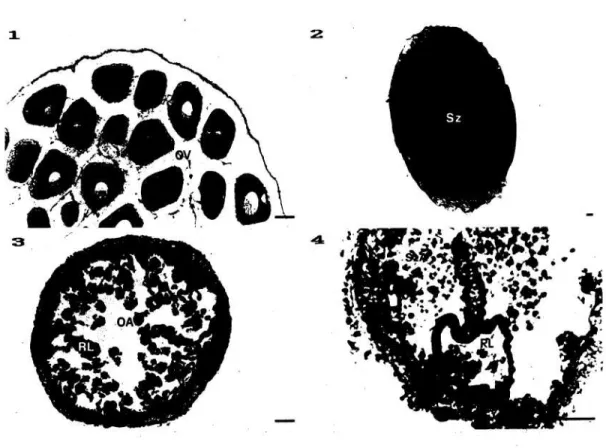
Gambar 5. Struktur matang (T4) dan tabung terserap (T5) Holothuria forskali. 1 dan 2 : tabung matang pada hewan betina dan jantan; 3 dan 4 : tabung terserap pada hewan betina dan jantan; OA : telur terabsorpsi; OV : telur vitelogenesis; PG : dinding tabung; RL: lipatan longitudinal; Sz: Sperma-tozoid; SzA : spermatozoid teransorpsi.
Panjang garis : 50 um..
DISKUSI Struktur Organ Reproduksi
Dasar gonad teripang dibungkus oleh peritoneum dari mesenter dorsal (SMILEY 1988) yang merupakan hasil pembelahan dari somatosoel pada waktu metamorfosis (SMILEY 1986). Strukturnya sana dengan peritoneum tabung gonad (SMILEY & CLONEY 1985). Pada bagian ventral-dorsal dari dari dasar gonad terdapat saluran yang menghubungkan saluran darah gonad dengan saluran darah usus (SMILEY 1988). Jaringan pengikatnya sangat kompleks, terdapat kumpulan sel berukuran sekitar 50 um (ATWOOD 1973, SMILEY 1988; SMILEY et al. 1991). Gonoduknya tertancap pada bagian anterior-dorsal dan menjalar di dalam konektif mesenter dorsal menuju ke gonopor yang terletak pada interambulakral CD (HYMAN 1955).
Peritoneum dari dinding tabung gonad selain berfungsi sebagai pelindung terluar, juga berfungsi menyerap nutrien dari air yang terdapat di dalam rongga tubuh atau soelom melalui mikrovili dan flagela yang terdapat pada permukaannya (KRISHNAN & DALE 1975).
Sel-sel syaraf yang terdapat pada peri-toneum kemungkinan berfungsi untuk mengontrol proses pematangan, ovulasi dan pemijahan. Sel-sel ototnya selain berperan pada saat pemijahan, juga berfungsi untuk menghomogenkan kandungan nutrien cairan darah, yaitu antara cairan yang terdapat pada dinding tabung yang kaya dengan nutrien dengan cairan darah yang terdapat jauh di dalam lipatan longitudinal yang kadar nutriennya lebih rendah (SMILEY & CLONEY 1985).
Problem hidrodinamika pada teripang menyebabkan transfer cairan darah dari saluran darah dorsal ke saluran darah genital yang terdapat pada ujung tabung sangat sulit. Karena itu penyerapan nutrien berlangsung juga pada peritoneum. Nutrien yang diserap oleh peritoneum kemudian disintesa dan hasilnya berupa butiran yang kaya dengan lemak, protein dan karbohidrat. Butiran ini kemudian ditransfer ke dalam cairan darah.
Telur pada H. forskali hanya mempunyai satu nukleolus. sedangkan pada Stichopus californicus (SMILEY & CLONEY 1985), Ypsilothuria talismani (TYLER & GAGE 1983), Aslia lefevrei dan Pawsonia
saxicola (TUWO 1993, TUWO & TRESNATI
1994b) dapat dijumpai lebih dari tiga nukleolus pada setiap telur. Semua jenis teripang mempunyai bentuk spermatozoa yang sangat primitif. Spermanya mempu-nyai nukleus yang lonjong, satu sentriol fossa yang kurang berkembang dan bebe-rapa mitokondria yang sederhana (PLADELLORENS & SUBIRANA 1974).
Evolusi Organ Reproduksi
Empat jenis tabung (Tl, T2, T3 dan T4) dari H. forskali dapat dibedakan dengan mudah berdasarkan struktur makroskopik dan mikroskopiknya. Proses perubahan dari tabung primer (Tl) ke tabung matang (T4) merupakan proses yang panjang.
Sebuah tabung primer (Tl) yang terbentuk pada bulan Januari tahun t akan berkembang menjadi tabung sekunder (T2) pada bulan April tahun yang sama. Setelah pemijahan berlangsung, tabung T2 mulai mengalami pra-vitelogenesis yang menandai awal perubahan T2 menjadi tabung pematangan (T3). Pada tabung T3 proses vitelogenesis atau pembentukan kuning telur
berlangsung sangat pesat. Proses ini berlangsung dari bulan April sampai bulan Oktober tahun t. Pada tabung matang (T4), sejak bulan November tahun t sudah terdapat telur matang yang layak untuk dipijahkan. Namun telur ini belum dipijahkan karena kondisi lingkungan yang buruk pada musim dingin sehingga hewan ini menahan telurnya hingga musim semi atau bulan Mei tahun t+1 (TUWO & TRESNATI 1994a), dimana pada saat yang sama terjadi blooming fitoplankton (RIAUX 1977). Peranan fitoplankton terhadap pemijahan invertebrata laut telah diuraikan oleh HIMMELMAN (1975).
Selama pemijahan, tabung T4 mengalami pengecilan karena kehilangan isinya. Telur dan spermatozoa yang tersisa selanjutnya diserap. Pada fase ini tabung T4 telah berubah menjadi tabung terserap (T5). Pada saat yang bersamaan dengan penyerapan tabung T5 ini, proses pematangan segera dimulai lagi pada tabung T2 lainnya yang terbentuk pada bulan Januari tahun t+1 yang lalu. Hal ini memungkinkan proses gameto-genesis secara berkelanjutan dari tahun ke tahun. Setelah semua telur dan spermatozoa terserap, tiba giliran dinding tabung itu sendiri yang terserap. Penyerapan dinding tabung T5 berlangsung sampai bulan September tahun t+1. Hasil penyerapan tabung ini digunakan kembali untuk pertumbuhan go-nad (SMILEY 1988).
Dari uraian di atas dapat diketahui bahwa setiap tabung pada H. forskali hanya mengalami satu kali masa gametogenesis. Setelah itu hilang tanpa bekas. Dari masa awal pembentukan tabung Tl pada bulan Januari tahun t sampai September tahun t+1 dapat diperkirakan bahwa umur sebuah tabung pada H. forskali ini adalah sekitar 21 bulan.
DAFTAR PUSTAKA
ATWOOD, D.G. 1973. Ultrastruktur of the gonadal wall of the sea cucumber, Leptosynapta clarki (Echinodermata : Holothuroidea). Z Zellforsch. 141 : 319 – 330.
HIMMELMAN, J.H. 1975. Phytoplankton as a stimulus for spawning in three ma-rine invertebrates. J. Exp. Mar. Biol. Ecol. 20 : 199 – 214.
HYMAN, L.H. 1955. The
invertabrates-Echinodermata. The coelomate
bilateria. Mc. Graw. Hill Comp. Lon-don : 1 – 763.
KRISHNAN S. and T. DALE 1975. Ultrastruktural studies on the testis of Cucumaria frondosa (Holothuroidea : Echinodermata). Norw. J. Zool. 23 : 1 – 15.
PLADELLORENS, M. and J.A. SUBIRANA, 1975. Spermiogenesis in the sea cu-cumber Holothuria tubulosa. J. Ultrastruct. Res. 52 : 235 – 242. RIAUX, C. 1977. Facteurs determinant
revolution de la biomasse phytoplanctonique et microphy-tobenthique dans lestuaire de la Penze (Nord-Finistere). J. Rech. Oceanogr. 2 (4) : 23 – 29.
SMILEY, S. 1986. Metamorphosis of the Stichopus californicus and its phylo-genetic implication. Biol. Bull. 171 : 611 – 631.
SMILEY, S. 1988. The dynamics of oogen-esis and the annual ovarian cycle of Stichopus californicus (Echinodermata : Holothuroidea). Biol. Bull. 175 : 79 – 93.
SMILEY, S. and R.A. CLONEY 1985. Ovu-lation and the fine structure of the Stichopus californicus (Echinodermata : Holothuroidea) Fecund ovarian tu-bules. Biol. Bull. 169 : 342 – 364. 21
SMILEY, S., MCEUEN, C. CHAFFEE and S. KRISHNAN 1991. Reproduction of marine invertebrates. Vol. VI. Echino-derms and Lophophorates. The box-wood press. California : 663 – 750. TANAKA, Y. 1958. Seasonal changes
occur-ring in the gonad of Stichppus japonicus. Bull. Fac. Fish. Hokkaido Univ. : 29 – 36.
TUWO, A. 1993. Biologie et ecologie de trois especes d'holothuries, Holothuria forskali, Aslia lefevrei et Pawsonia saxicola, en Bretagne Occidentale. These du Doctorat Nouveau Regime. UBO-Brest. 147 pp.
TUWO, A. and C. CONAND 1992a. Devel-opments in beche-de-mer production in Indonesia during the last decade. Beche-de-mer Information Bulletin 4 : 2 – 3.
TUWO, A. and C. CONAND 1992b. Repro-ductive biology of the holothurian Holothuria forskali (Echinodermata). J. Mar. Biol. Ass. U.K. 72 : 745 – 758. TUWO, A. dan M.N. NESSA 1991. Beberapa
aspek biologi teripang ekonomis. Torani 1 (1) : 1 – 20.
TUWO, A dan J. TRESNATI 1994a. Pengaruh variasi suhu dan spasial terhadap biologi reproduksi makrobentos Holothuria forskali. Torani 4 (4) : 22 – 30.
TUWO, A. dan J. TRESNATI 1994b. Studi fekunditas dengan pendekatan strategi reproduksi (Aplikasi pada teripang). Torani 4 (4) : 31 – 36.
TYLER, P.A. and J.D. GAGE 1983. The reproductive biology of ypsolothuria
talismani (Holothuridea :
Dendro-chirota) from the N.E. Atlantic. J. Mar. Biol. Ass. U.K. 63 : 609 – 616.