Journal of Experimental Marine Biology and Ecology 253 (2000) 115–127
www.elsevier.nl / locate / jembe
Effects of sediment type and intra- and interspecific
competition on growth rate of the marine snails Hydrobia
ulvae and Hydrobia ventrosa
*
Johan Grudemo , Torgny Bohlin
¨ ¨
Department of Zoology, Goteborg University, Box 463, SE-405 30 Goteborg, Sweden Received 21 January 2000; received in revised form 22 May 2000; accepted 30 June 2000
Abstract
The combined effects of sediment grain size and intra- and interspecific competition on growth rate of Hydrobia ulvae and H. ventrosa were investigated in a 2-month laboratory experiment. Individuals of H. ulvae grew faster than H. ventrosa at all competition intensities. Growth rate did not differ between fine- and coarse-grained sediments in any of the species. This suggests that other factors than the sediment quality cause the often observed size differences between Hydrobia snails from these sediment types in nature. Competition from 1.6 to 1.9 times larger competitors affected growth of the smaller target snails. Competition between the species was asymmetrical and, in both species, per capita effect of competition was stronger from H. ulvae than from H. ventrosa, regardless of sediment type. From growth, Lotka-Volterra competition coefficients were estimated to 0.2 from H. ventrosa on H. ulvae and 1.8 from H. ulvae on H. ventrosa. Since competition between the species seems to be independent of sediment grain size, the shift in numerical dominance in nature, from H. ulvae on coarse-grained sediment to H. ventrosa on fine-grained, could not be explained with different competitive ability in different environments. 2000 Elsevier Science B.V. All rights reserved.
Keywords: Competition; Hydrobia; Sediment; Growth rate; Character displacement; Experimental design
1. Introduction
The relation between body size and competition among closely related species has been a recurrent subject during the last decades, ever since the influential papers by Brown and Wilson (1956) and Hutchinson (1959). The size in sympatry and allopatry of
*Corresponding author. Fax:146-31-416-729.
E-mail address: [email protected] (J. Grudemo).
the two marine mud-snails Hydrobia ulvae (Pennant) and H. ventrosa (Montagu), which are common on littoral muddy sediments, and which partly coexist in this seemingly homogeneous habitat, is one example. Fenchel (1975b) found that the two species were of similar size when living apart from each other (in allopatric localities), but that H. ulvae was larger than H. ventrosa when together (in sympatric localities). A similar size segregation was also found in Sweden (Grudemo and Johannesson, 1999), although H. ventrosa tended to be larger than H. ulvae in allopatry. In contrast, other studies from Denmark (Hylleberg, 1986) and England (Cherril and James, 1987a) showed no size difference between allopatry and sympatry.
Fenchel (1975b) proposed that the reason for the observed size pattern was character displacement, caused by body size-dependent food size preference, with the same relation in both species. Hence, the two species were proposed to be affected by limiting similarity for body size, which has led to genetically based divergence in size when coexisting. These size relations in Hydrobia are among the most widely cited to support or illustrate character displacement.
Other studies, however, have stressed the possibility of non-genetic environmental causes (salinity, sediment quality, water movements, etc.) to this pattern (Hylleberg, 1986; Cherril and James, 1987a; Saloniemi, 1993). H. ulvae is often larger on fine-grained sediment than on coarse-grained (Chatfield, 1972; Fish and Fish, 1974; Morrisey, 1990; Grudemo and Johannesson, 1999; but see Barnes (1988) for a divergent observation). H. ventrosa is less well studied; at the Swedish West coast they are, however, like H. ulvae, generally larger on fine-grained than on coarse-grained sediment (Grudemo and Johannesson, 1999). It is thus possible that size differences might be caused by differences in sediment type, or by conditions correlated to this factor, such as water movements (Barnes, 1988). In addition, the distribution of the species is correlated to sediment type, at least at the Swedish West coast, where H. ulvae dominates on coarse-grained sediments and H. ventrosa on fine-grained (Grudemo and Johannesson, 1999). It can therefore not be excluded that the combined effects of the environment on the distribution and body size may be an alternative to the character displacement explanation.
In this study we attempted to explore this possibility further. Since growth has a role in determining body size, a first question was if differences in sediment type (including differences in grain size, water content, organic content and other differences) lead to differences in growth rate. This might explain why both species generally are larger on silty sediments. Such differences may occur either if food production or the efficiency to utilise this depends on sediment type. For example, food production may be higher on fine-grained sediments due to larger particle surface (Newell, 1965). Previous studies on the effect of sediment type on the growth of Hydrobia species give no solid information. For H. totteni, Levinton (1982) found no difference in growth between one fine-grained and one coarse-grained sediment. Unfortunately, the experiment was unreplicated and his results thereby circumstantial. Morrisey (1990) studied growth of H. ulvae on sediments of different particle size in the field with conflicting results, partly because of poor replication. In addition, his study was conducted in the field, where sediment structure is correlated with other factors, and the sediment effect thereby not possible to estimate.
J. Grudemo, T. Bohlin / J. Exp. Mar. Biol. Ecol. 253 (2000) 115 –127 117
therefore how competition within and between the species affects growth and thereby possibly body size. Indeed, competition over food within and among Hydrobia species seems to be important in nature (Fenchel, 1975b; Fenchel and Kofoed, 1976; Hylleberg, 1986; Levinton, 1985; Cherril and James, 1987b; Morrisey, 1987; Gorbushin, 1996). Morrisey (1987) found that growth of H. ulvae decreased rapidly due to intraspecific
22
competition up to a population density of 10–20 000 m , and then more slowly.
22
Growth stopped entirely at 50 000 m . Fenchel and Kofoed (1976) suggested from
experiments that intra- and interspecific competition were of the same strength in H. ulvae and H. ventrosa, whereas Cherril and James (1987b) concluded, from a field experiment, that intraspecific competition was stronger than interspecific. Gorbushin (1996) measured growth in the field and found that competition between H. ulvae and H. ventrosa was asymmetrical with H. ulvae as the stronger competitor. Thus, although these three studies indicate that the two species do compete, the conclusions concerning the effect of this are conflicting. However, none of them satisfied the demands for replication and unconfounded design of the competition levels (Underwood, 1986), so the conclusions should be interpreted cautiously. Our second objective was therefore estimate the effect of intra- and interspecific competition on growth in these species using an experimental set-up specifically designed for this purpose.
The observation that abundance and body size are correlated to sediment quality, together with the evidence for competition under natural conditions, leads to three questions of interest, not previously investigated experimentally: does sediment type affect the outcome of competition between the species? Is the predominance of H. ventrosa on finer substrates caused by a greater competitive ability on such sediments. And, is H. ulvae the better competitor on coarser substrates?
To investigate these questions further we conducted an experiment where the main purpose was to test the following hypotheses:
1. both species grow faster on fine-grained sediment than on coarse-grained (Chatfield, 1972; Fish and Fish, 1974; Morrisey, 1990);
2. intra- and interspecific competition is of the same strength (Fenchel, 1975b); 3. intraspecific competition is stronger than interspecific in both species (Cherril and
James, 1987b)
4. competition is asymmetric, with H. ulvae as the stronger competitor (Gorbushin, 1996);
5. the effect of interspecific competition is dependent of sediment type.
In the experiment, we used shell growth of young individuals as the response variable. We thereby assumed that growth is a predictor of fitness of non-reproductive individuals.
2. Materials and methods
2.1. Experimental set-up
¨ ¨ ¨
experiment was performed at Tjarno Marine Biological Laboratory, nearby Stromstad at the Swedish West coast. Snails were placed on a 1-cm thick sediment layer in a plastic
2
cup with a bottom area of 13 cm . The cups were placed in a climate chamber with a light regime (16:8 h light / dark cycle) and in running sea water (30‰ salinity) that could reach the inside of the cups through a net that served to keep the snails within the cups
2
(aperture size, 0.8 cm ; mesh size, 0.5 mm). To make sure that water was changed, all water in the cups was drained and refilled once every second day. The water was taken from 30 m depth in the nearby Koster fjord. The temperature of the water was ambient
and decreased gradually from 158C in the beginning of the experiment and 108C in the
end. The cups were lighted by fluorescent tubes with appropriate light for algae growth.
2.2. Sediments and snails
¨
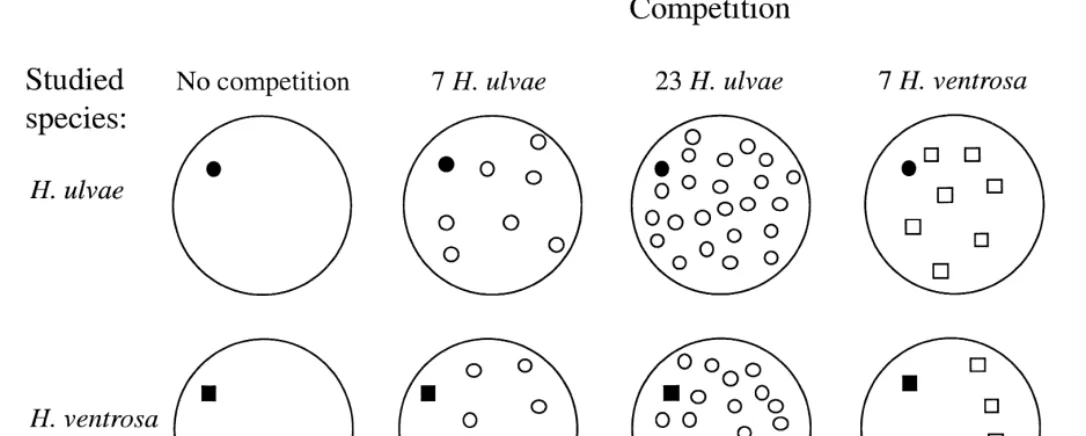
All snails and sediments were collected in the almost atidal area around Stromstad at the Swedish West Coast (Grudemo and Johannesson, 1999). In each plastic cup, we put one randomly chosen marked snail from one of the species. This snail was used for growth measurement and will further be called the target snail. Most cups also contained other snails, used as competitors (Fig. 1). Before and after growth, we measured the length of the target snails from the apex to the anterior part of the margin to the nearest 0.04 mm with an ocular micrometer. At the start of the experiment, the size of the target H. ulvae individuals were 1.5860.09 mm (n5240, mean6S.D.). The size target H. ventrosa individuals were 1.5960.11 mm (n5240). As a measure of the growth rate of these, we used the absolute length increment, which is a relevant growth estimate when, as in this experiment, the target individuals are of equal initial size. The size of the competitors of both species was 2.6–3.0 mm, thus 1.6 to 1.9 times larger than the target snails.
The properties of the sediments used were determined with the method in Buchanan
(1984). The weight of the silt-clay fraction (,64mm) as a proportion of the weight of
all fractions smaller than 1 mm was used to characterise the sediment. On the basis of ¨
this, we divided the 16 sediments collected in the area around Stromstad at the Swedish West coast, into two sediment types, one fine-grained (with eight sediments) with
45.6621.9% (mean6S.D.) silt and clay (fractions ,64 mm), and one coarse-grained
(eight sediments) with 4.461.1% (mean6S.D.) silt and clay. Before the start of the
experiment, we picked out most of the snails from each of the 16 sediments and froze them for 3 days to kill undiscovered individuals. After melting, an unfrozen, well-picked sample of each sediment was added to restore the native microalgal flora.
2.3. Competition treatments
We used five levels of competition in the experiment (Fig. 1): level 1, no competition
22
(1 target snail of H. ulvae or H. ventrosa: population density, 770 m ); level 2,
medium competition from H. ulvae (one target snail1seven H. ulvae competitor snails:
22
total population density 6160 m ); level 3, high competition from H. ulvae (one target
22
J.
Grudemo
,
T
.
Bohlin
/
J.
Exp
.
Mar
.
Biol
.
Ecol
.
253
(2000
)
115
–
127
119
medium competition from H. ventrosa (one target snail1seven H. ventrosa competitor
22
snails: density 6160 m ); and level 5, high competition from H. ventrosa (one target
22
snail123 H.ventrosa competitor snails: density 18 480 m ). These figures were
chosen to represent low, intermediate and high population density and competition (Fenchel, 1975a; Morrisey, 1987).
2.4. Statistical analyses
Growth of the target snails was analysed with a four-factor analysis of variance (ANOVA). In this, there were three orthogonal factors, Species (SP), a fixed factor with two levels (H. ulvae and H. ventrosa); Competition (C), a fixed factor with five levels (see above); and Sediment type (ST), a fixed factor with two levels (sandy or silty sediment, see above). The fourth factor was a random Bay (B) factor with eight levels, nested under Sediment type (ST). That is, eight sediments came from bays with coarse-grained sediment and eight from bays with fine-grained sediments.
This experimental design resulted in an ANOVA with the following linear model:
xijklm5m 1SPi1Cj1STk1B(ST)l(k )1SP3Cij
1SP3STik1SP3B(ST)il(k )1C3STjk
1C3B(ST)jl(k )1SP3C3STijk
1SP3C3B(ST)ijl(k )1eijklm (1)
where xijklmis the growth of a specific target individual, m is the growth averaged over
all treatments, and eijklm is the residual.
We used three replications of each combination of treatments, resulting in a total number of 480 target snails. To obtain appropriate interactions (Hurlbert and White, 1993), and to reduce the heterogeneity in variances, we used the logarithm of growth
(ln(growth11)) in the ANOVA. Degrees of freedom and expected mean sum of squares
were calculated with the method in Underwood (1997, pp. 364–369). To obtain a balanced statistical analysis, we replaced dead snails with the average growth of the survivors within the same treatment, and we accordingly reduced the degrees of freedom in the residual (Underwood, 1997). Student–Newman–Keul’s (SNK) test was used as post-hoc test to unveil differences among levels of the significant factors and interac-tions. If the relationship between growth and level of competition is similar between
species, then the interaction term (SP3C) will not be significant. If, on the other hand,
intra- or interspecific competition is more important, the interaction effect will become large.
3. Results
3.1. General pattern
J. Grudemo, T. Bohlin / J. Exp. Mar. Biol. Ecol. 253 (2000) 115 –127 121
lost the paint or died. These 14 snails came from all competition treatments. The high survival at all competition intensities suggests that a difference in survival is no good measure of differences in fitness within the timespan of 2 months.
In contrast, there was large and significant variation in growth rate among snails from different treatments. The ANOVA (Table 1) showed that a major part of this variation could be attributed to the main factors Species and Competition. All other factors and interactions were of minor magnitude, although some of them were significant.
3.2. Effect of sediment grain size on growth
The two sediment-related main factors, Sediment type and Bay, were neither significant nor important. Thus, if there is a difference in snail growth between these sediment types in nature, this experiment suggests that it is not caused by the sediment type itself, but with other biotic or abiotic factors that may be correlated to sediment type.
The interaction Species3Sediment type was weak (low SS) and just significant
(P50.050), and the SNK-test did not (due to lower power) discover any pairwise
differences. It appears that this interaction was caused by H. ventrosa growing slightly better on coarse-grained sediments than on fine-grained, whereas H. ulvae grew similarly
well on both sediment types. Finally, the interactions C3ST and S3C3ST were
non-significant, indicating that different competition intensities had similar impact on growth on individuals of both species regardless of sediment type.
3.3. Effects of intra- and interspecific competition
The interaction Species3Competition and the main factors Species and Competition
were all significant. The interaction Species3Competition is of interest since it is
directly related to the different hypotheses about the strength of intra- and interspecific competition. Although the interaction was significant, it was rather weak. The SNK-test
Table 1
Result of the ANOVA based on data transformed with ln(x11)
Factor SS d.f. MS F-ratio F P
versus
Species, SP 17.662 1 17.662 SP3B(ST) 669.71 ,0.0001
Competition, C 32.500 4 8.125 C3B(ST) 206.48 ,0.0001
Sediment type, ST 0.010 1 0.010 B(ST) 0.48 0.51
Bay, B(ST) 0.289 14 0.021 Residual 0.77 0.69
SP3C 0.898 4 0.224 SP3C3B(ST) 7.93 ,0.0001
SP3ST 0.122 1 0.122 SP3B(ST) 4.61 0.05
SP3B(ST) 0.369 14 0.026 Residual 0.99 0.46
C3ST 0.183 4 0.046 C3B(ST) 1.16 0.33
C3B(ST) 2.204 56 0.039 Residual 1.48 0.02
SP3C3ST 0.083 4 0.021 SP3C3B(ST) 0.73 0.58
SP3C3B(ST) 1.584 56 0.028 Residual 1.06 0.37
Residual 8.051 302 0.027
of the interaction showed that H. ulvae grew significantly faster than H. ventrosa at all competition intensities (level 1, 69%; level 2, 68%; level 3, 329%; level 4, 74%; level 5, 167%). However, the SNK-test showed different grouping of growth of the two species
at different competition levels. In H. ulvae, competition levels grouped as 154.5.2.
3, indicating that growth was similar without competition and with medium competition from H. ventrosa, and that high competition from H. ventrosa as well as medium and high competition from H. ulvae reduced growth. Thus, for H. ulvae, intraspecific competition was stronger than interspecific. In contrast, for H. ventrosa the levels
grouped as 154.2.5.3, showing that H. ventrosa was more influenced by
competition from H. ulvae than from its own species. The significant interaction in the ANOVA was caused by H. ulvae being less affected by high population density of H. ventrosa, than H. ventrosa itself (Fig. 2).
3.4. Calculation of competition coefficients
Although not primarily designed for the purpose, we also used the results to estimate
Fig. 2. Interaction term Species3Competition. Average growth (ln(growth11) of target snails, with standard error bars indicated, is shown for different treatments. (j) H. ulvae target snails without competition and with competition from medium and high densities of H. ulvae. (h) H. ulvae target snails with competition from H.
ventrosa. (d) H. ventrosa target snails without competition and with competition from H. ulvae. (s) H.
J. Grudemo, T. Bohlin / J. Exp. Mar. Biol. Ecol. 253 (2000) 115 –127 123
the quantitative effect of competition on growth by regression, assuming a linear relation (Fig. 3) between population density and growth. In this analysis we used growth in mm (not on the logarithmic scale). For each species we first tested the slopes of the regression lines, with growth explained by the factor competition, the regressor population density, and the interaction between them (ANCOVA). The interactions were
significant (P,0.0001) for both species, suggesting that the strength of intra- and
interspecific competition were different in both species. We therefore used linear regressions separately for intra- and interspecific competition in each species to test and estimate the slopes (Fig. 3).
The results were:
2
• H. ulvae: intraspecific competition: n5135, r 50.695, P,0.0001, slope b(uu)5
25 2
29.25310 ; interspecific competition: n5140, r 50.066, P50.0013, slope
25
b(uv)5 21.89310 ;
2
• H. ventrosa: intraspecific competition: n5143, r 50.403, P,0.0001, slope b(vv)5
Fig. 3. Linear regressions of growth of H. ulvae and H. ventrosa target snails at different population densities. Filled symbols show growth with intraspecific competition and open symbols growth with interspecific competition. Each symbol is the average growth of 43–48 independent individuals, with standard error bars
2
25 2
23.85310 ; interspecific competition: n5138, r 50.727, P,0.0001, slope
25
b(vu)5 27.10310 .
For H. ulvae, interspecific competition had thus less effect on growth than intra-specific, whereas H. ventrosa suffered more from interspecific than intraspecific competition.
As the relations above between population density and growth were approximately linear (Fig. 3), we used the slopes to calculate Lotka-Volterra (LV) competition coefficients, for each species, obtained as the ratio (slope under interspecific competi-tion) /(slope under intraspecific competicompeti-tion). The LV-coefficients were thus calculated to
a 50.2 [5b(uv) /b(uu)] from H. ventrosa on H. ulvae, andb 51.8 [5b(vu) /b(vv)] from
H. ulvae on H. ventrosa.
4. Discussion
During the 2 months of the experiment, the growth in all treatments was positive and
the survival rate high (.97%). This indicates that the experimental conditions, including
the algal production on the sediments, corresponded to the requirements of the snails, and thus that their responses to the treatments may reflect those under natural conditions. As always, however, results from the laboratory should be evaluated with care, since all conditions are not exactly as in the field.
A first conclusion is that there was no difference in growth between fine- and coarse-grained sediments in any species. This indicates that the answer to our first question, whether both species grow better on finer sediment, is no. The hypothesis that sediment-dependent growth would explain why both species in nature usually are larger on fine-grained sediments (Chatfield, 1972; Fish and Fish, 1974; Morrisey, 1990) was therefore not supported. This suggests that other environmental conditions, correlated to sediment composition, may affect growth; Barnes (1988) argued that water movements may affect body size in H. ulvae, but did not test this hypothesis. A second possibility is that average age, and thereby body size, is larger on fine-grained substrates. A crucial question is thus whether the larger size in this type of locality is related to growth or age. Fenchel’s (1975b) suggestion that the size segregation in sympatry in H. ulvae and H. ventrosa may be caused by differential recruitment time remains untested, so compara-tive field studies on growth and time of recruitment would be most valuable.
J. Grudemo, T. Bohlin / J. Exp. Mar. Biol. Ecol. 253 (2000) 115 –127 125
simply be a result of differential growth rates. Thus, the question why they are of similar size in size in allopatry may be more useful than why they are dissimilar in sympatry. The second main conclusion is that the growth rate of both species was affected by competition, and specifically that H. ulvae affected growth of both species more than H. ventrosa did. Our results confirmed the findings by Morrisey (1987) and Gorbushin (1996) that competition reduces growth within the two species even at medium population densities, suggesting that growth in the field is limited by competition, and that the species do compete under natural conditions. Our experiment indicated that the competition coefficients were largely independent of population densities within the range of densities used. This is not in concordance with Gorbushin’s (1996) conclusion of density-dependent competition coefficients. However, his experiment, like the present, was not especially designed for this purpose, so further studies are required to resolve this matter.
We found that competition between the species was asymmetrical, with competition
coefficients of a¯0.2, from H. ventrosa on H. ulvae, and b¯2 from H. ulvae on H.
ventrosa. This asymmetry may be surprisingly large, since the species are similar-sized and related. However, from studies of other animals and plants, it seems that asymmetry is the rule rather than the exception (Schoener, 1983). The three previous experimental studies of competition between H. ulvae and H. ventrosa published so far give little solid information on this matter. In the first, Fenchel and Kofoed (1976) found interspecific competition coefficients close to 1 for both species. Unfortunately, the growth of H. ventrosa in their study was around zero or lower in all treatments except one, which makes the estimates of competition coefficients less accurate. A second study was that of Cherril and James (1987b), who used survival as the response variable, and found interspecific competition coefficients considerably lower than 1 in both species. They seem, however, to have misinterpreted their data; mortality (loss rate) was not density-dependent, as they concluded, but density independent.
In the third study, Gorbushin (1996) found, as in the present study, asymmetric competition with H. ulvae as the stronger competitor. He used, however, the problematic replacement design (Underwood, 1986) and a pseudoreplicated set-up. From the previous studies, it is therefore not possible for us to judge the generality of our finding that competition asymmetry between these species is large. Indeed, since such a result is not expected given the type of species and environment, it would be of value to conduct similar experiments over a wider range of environmental factors.
may have had to disperse. This is well described from lagoons in England (Barnes and Gandolfi, 1998; Barnes, 1999), but what prevents H. ulvae from dominating sheltered bays at the Swedish West coast is not known. Anyhow, as the two species are similar in most respects except in their mode of reproduction, it appears that the difference in dispersal, and thereby possibly also juvenile survival, is a factor of importance for the distribution and abundance of the species. In addition, the dominance of H. ventrosa in sheltered areas may be caused by a lower adult mortality; Hylleberg and Siegismund (1987) found that H. ventrosa survived freezing better than H. ulvae, which might be an advantage in sheltered and thereby brackish localities.
Is the hypothesis of non-genetic, environmentally determined body sizes and distributions a strong alternative to Fenchel’s character displacement theory based on limiting similarity? The results of the present study did not support that sediment-dependent growth in combination with sediment-sediment-dependent distribution would explain the size similarity in allopatry; other and so far unknown mechanisms must then be assumed. However, a crucial experiment to assess the character displacement theory would be to test if individuals affect each other more when of equal than when of unequal size. In addition, the character displacement theory is based on local adaptation to interspecific competition, requiring low dispersal between allopatric and sympatric population. In H. ventrosa, which has viviparous reproduction, this seems more likely than in H. ulvae with its planktonic larvae. In order to discriminate among these alternative explanations, a second major task would therefore be to compare the genetic structure of the two species.
Acknowledgements
We thank Kerstin Johannesson and Thomas Dahlgren for help with the study, and two anonymous referees for valuable comments on the manuscript. The study was performed
¨ ¨
at Tjarno Marine Biological Laboratory, and the staff at the laboratory is thanked for
˚ ¨
much help. The study was financed by Radman och Fru Ernst Collianders stiftelse for
¨ ˚ ¨
vetenskapliga andamal, Kungliga och Hvitfeldtska stipendieinrattningen and Wilhelm
och Martina Lundgrens vetenskapsfond to J.G. [AU]
References
Barnes, R.S.K., 1988. On reproductive strategies in adjacent lagoonal and intertidal marine populations of the gastropod Hydrobia ulvae. J. Mar. Biol. Assoc. UK 68, 365–375.
Barnes, R.S.K., 1999. What determines the distribution of coastal hydrobiid mudsnails within North-Western Europe? P.S.Z.N. Mar. Ecol. 20, 97–110.
Barnes, R.S.K., Gandolfi, S.M., 1998. Is the lagoonal mudsnail Hydrobia neglecta rare because of competitively induced reproductive depression and, if so, what are the implications for its conservation? Aquatic Conserv: Mar. Freshwat. Ecosyst. 8, 737–744.
Brown, W.L., Wilson, E.O., 1956. Character displacement. Syst. Zool. 5, 49–64.
J. Grudemo, T. Bohlin / J. Exp. Mar. Biol. Ecol. 253 (2000) 115 –127 127 Chatfield, J.E., 1972. Studies on variation and life history in the prosobranch Hydrobia ulvae (Pennant). J.
Conch. 27, 463–473.
Cherril, A.J., James, R., 1987a. Character displacement in Hydrobia. Oecologia 71, 618–623.
Cherril, A.J., James, R., 1987b. Evidence for competition between mudsnails (Hydrobiidae): a field experiment. Hydrobiologia 150, 25–31.
Fenchel, T., 1975a. Factors determining the distribution patterns of mud snails (Hydrobiidae). Oecologia 20, 1–17.
Fenchel, T., 1975b. Character displacement and coexistence in mud snails (Hydrobiidae). Oecologia 20, 19–32.
Fenchel, T., Kofoed, L.H., 1976. Evidence for exploitative interspecific competition in mud snails (Hydro-biidae). Oikos 27, 367–376.
Fish, J.D., Fish, S., 1974. The breeding cycle and growth of Hydrobia ulvae in the Dovey Estuary. J. Mar. Biol. Assoc. UK 54, 685–698.
Gorbushin, A.M., 1996. The enigma of mud snail shell growth: asymmetrical competition or character displacement? Oikos 77, 85–92.
Grudemo, J., Johannesson, K., 1999. Size of mudsnails, Hydrobia ulvae (Pennant) and H. ventrosa (Montagu), in allopatry and sympatry: conclusions from field distributions and laboratory growth experiments. J. Exp. Mar. Biol. Ecol. 239, 167–181.
Hutchinson, G.E., 1959. Homage to Santa Rosalia, or Why are there so many kinds of animals? Am. Nat. 93, 145–159.
Hurlbert, S.H., White, M.D., 1993. Experiments with freshwater invertebrate zooplanctivores: quality of statistical analyses. Bull. Mar. Sci. 53, 128–153.
Hylleberg, J., 1986. Distribution of Hydrobiid snails in relation to salinity, with emphasis on shell size and co-existence of the species. Ophelia 4, 85–100.
Hylleberg, J., Siegismund, H.R., 1987. Niche overlap in mud snails (Hydrobiidae): freezing tolerance. Mar. Biol. 94, 403–407.
Levinton, J.S., 1982. The body size-prey size hypothesis: the adequacy of body size as a vehicle for character displacement. Ecology 63 (3), 869–872.
Levinton, J.S., 1985. Complex interactions of a deposit feeder with its resources: roles of density, a competitor, and detrital addition in the growth and survival of the mudsnail Hydrobia totteni. Mar. Ecol. Prog. Ser. 22, 31–40.
Morrisey, D.J., 1987. Effect of population density and presence of a potential competitor on the growth rate of the mud snail Hydrobia ulvae (Pennant). J. Exp. Mar. Biol. Ecol. 108, 275–295.
Morrisey, D.J., 1990. Factors affecting individual body weight in field populations of the mudsnail Hydrobia
ulvae. J. Mar. Biol. Assoc. UK 70, 99–106.
Newell, R., 1965. The role of detritus in the nutrition of two marine deposit feeders, the prosobranch Hydrobia
ulvae and the bivalve Macoma balthica. Proc. Zool. Soc. London 144, 25–45.
Saloniemi, I., 1993. An environmental explanation for the character displacement pattern in Hydrobia snails. Oikos 67, 75–80.
Schoener, T.W., 1983. Field experiments on interspecific competition. Am. Nat. 122 (2), 240–285. Underwood, A.J., 1986. The analysis of competition by field experiments. In: Kikkawa, J., Anderson, D.J.
(Eds.), Community Ecology: Pattern and Process. Blackwell, Melbourne, pp. 240–268.