http://dx.doi.org/10.17503/Agrivita-2012-34-1-p028-035
GENETIC CONTROL OF SOYBEAN RESISTANCE TO SOYBEAN POD SUCKER
(
Riptortus linearis
L.)
Asadi1), Aziz Purwantoro2) and Sahiral Yakub3)
1)
Indonesian Center for Agricultural Biotechnology and Genetic Resources Research and Development. Jl. Tentara Pelajar 3 A, Bogor 16111 West Java Indonesia
2)
Faculty of Agriculture, Gadjah Mada University Bulaksumur Yogyakarta 55281 Indonesia
3)
Faculty of Agriculture Sultan Ageng Tirtayasa University Jl. Raya Jakarta Km 4 Pakupatan Serang Banten Indonesia
*) Correspond author Phone: +62-251-8337975 E-mail: [email protected]
Received: February 22, 2011/ Accepted: December 2, 2011
ABSTRACT
Riptortus linearis represents the most common pod sucking pest on soybean. Genetic studies on inheritance of resistance to the pest are needed. The study used B4400 and B3802 genotypes as resistant parents, Tambora as susceptible parent. Crosses were made to form F2 population of Tambora x B4400, and Tambora x B3802. A total of 10-20 seeds each of resistant and susceptible parents, 220 F2 seeds of Tambora x B4400 crosses, and 232 F2 seeds of Tambora x B3802 crosses were grown in pots, one plant/pot. The plants were infested with adult R. linearis (riptortus), at the R2 stage. The results showed that the population of riptortus in each of the F2 soybean population at 7 and 15 days after infestation were >2 insects/25 plants, which enables the study of genetic resistance to soybean pod sucker. Heritability values indicated that resistance to pod sucker was controlled by genetic factors. The resistance to riptortus in B4400 and B3802 genotypes was controlled by two recessive genes located at different loci, and interacted with each other with epistatic dominant reaction. With an assumption that B and C genes were dominant, hence the resistance genes found in both soybean genotypes were bbcc.
Keywords: genetic resistance, soybean pod sucker Riptortus linearis L.
INTRODUCTION
Currently, soybean to which the govern-ment gives priority is one of the most important food commodities after rice and maize. Soybean
consumption in Indonesia continues to increase annually, but the national production of soybean in the past 10 years (1988-2005) are still relatively low, about 1,22 million ton/year with an average productivity of about 1.18 t/ha (Sudaryanto and Swastika, 2007) although the soybean productivity based on research results reaches 3:07 t/ha (Taufiq and Wijanarko, 2007). One reason for the low productivity is the high yield losses by pests and diseases.
The soybean pod sucker has a similar morphology to the stinky bug of rice pest, with brownish yellow color, but it has yellowish white lines along the side of the body. This young and adult Coccinellidae attack soybean plants by sucking the liquid and the seed pod. The reproductive developmental stages of the plant defined as R3, R4 and R5 were sensitive to the pod suckers (Acle and Rolim 1994; Marwoto and Sri Hardaningsih, 2007). This insect jabbed its mouth into the pod skin and penetrated into the seed to suck the seed liquid. The pest attacks during the pod and seed development growth stage causing the pod and seed to turn flat, then dry and fall. When the attack occurs during the pod ripening stage, it generates brown spot symptoms on the pod skin and seed, resulting in losses of seed quantity and quality (Marwoto and Sri Hardaningsih, 2007).
In general, efforts to control soybean pod sucker still rely on the use of chemical insecticides. Unwise use of insecticides can cause resistance and resurgence to the insects, kill useful insects, and pollute the environments. The use of resistant varieties is an alternative to control the soybean pod sucker and they are environmentally friendly. Currently, no soybean varieties were reported resistant the soybean pod suckers. Of the 55 soybean varieties that conjunction with this, knowledge about the genetics of host resistance against the pest is very important as a breeding ground in selecting the appropriate method for resistance evaluation. Currently, information on genetic inheritance of resistance of soybean to the pod sucker is still very limited.
To study the genetics of soybean resistance to the soybean pod sucker, the genetic materials needed are resistant parents, susceptible parents, as well as F1 and F2 generations from crosses between the resistant and susceptible parents. From the previous selection of soybean germplasm for resistant to the soybean pod sucker 4 resistant and 5 susceptible parents were obtained. Cross-breeding between the two resistant parents (B4400, B3802) and a susceptible one (Tambora) have also been carried out to obtain seeds of F1
and F2 generations (Purwantoro et al., 2008; Purwantoro et al., 2009).
harvest based on the percentage of soybean pod sucker attack, which is determined by the percentage of seed damages by riptortus.
Phenotypic variability in resistance to soybean pod sucker was calculated using the formula:
where; = standard deviation of parent; Xi = resistance score of each plant to pod sucker at observation-i, μ = mean value of resistance score, N = Number of plant observed
Heritability in a broader sense was calculated using the following formula of Chahal and Gosal (2002):
where H2 = Heritability value; = genetic
variance, and = environmental variance.
While the genetic variability was calculated using the formula:
The environment variability was estimated from the environmental variability of the parent based on the formula of Suharsono and Yusuf calculated based on the ratio of the resistant and the susceptible plants. Model of inheritance and the number of genes controlling the resistance to the soybean pod sucker was predicted on the basis of genetic ratio fit according to Fehr (1987) and Wagner et al. (1980).
The frequency of observation values (scores) of resistance in the F2 plant population was determined based on the percentage of seed damages using a scoring of 0-4 (Table 3), which were grouped into two, three, or four score 2 = susceptible group 2, and scores 3-4 = susceptible group 3.
Suitability of genetic ratio was tested by Chi-square formula according to Gomez and Gomez (1984): Based on the genetic ratio, the number and characteristics of the controlling resistance to the soybean pod sucker can be predicted.
RESULTS AND DISCUSSION
Population of Soybean Pod Sucker
The population growth of riptortus in each of the F2 plant populations from the two-cross increased sharply in each week after infestation. Pest population reached more than two insects per 25 plants, which are considered above the economic threshold. The populations of reptortus from 7 to 15 DAI was high enough, thus met the requirement as a basis for resistance evaluation to soybean pod sucker. According to Ewete and Joda (1996) the severity of seed damage of soybean is closely related to riptortus density. A population of two to eight insects per plant caused significant damage on two soybean varieties TGX 536-02D and TGX 849-294D, when compared to the non-infested check.
Heritability
means the amount or portion of a character is controlled by its genetical factor (Allard, 1966). The heritability value can be used to determine a selection method, and whether the selection needs to be conducted at early generation using pedigree or at late generation using bulk or SSD selection method.
In this study, the heritability value of the two cross combinations between the susceptible parent and the resistance parents, was 0.99, which is very high (Table 2). The heritability value >0.5 is considered high, which means that the effect of environment on the variability of score of resistance is very low. It indicated that resistance of the soybean genotypes to the soybean pod sucker was controlled by genetic factors (resistance genes). The high heritability values indicated that the evaluation of the soybean breeding materials for resistance to the soybean pod sucker might have been done at early generation (F2 generation) by using pedigree method.
F2 Populations of Tambora x B4400 and Tambora x B3802
Scoring for resistance of the individual plant of the F2 soybean generation to the soybean pod
sucker was done after harvest. The frequency distribution of the F2 soybean plant populations of Tambora x B 4400 and Tambora x B 3802 which was scored based on the percentages of the soybean pod sucker was shown in Table 3, while the morphological performances of the F2 soybean plants were shown in Figure 1 and 2.
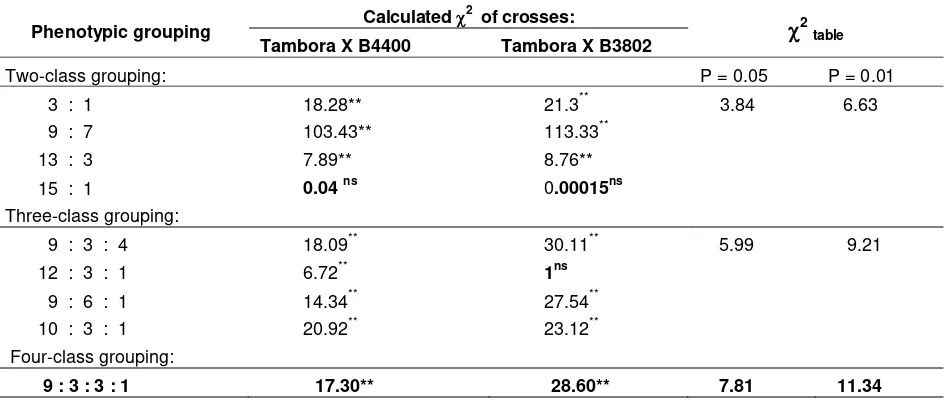
Frequency values of both F2 soybean populations based on Chi-square analysis fit to the two class grouping with ratio of 15 susceptible : 1 resistant (Table 4). Genetic ratio of 15 : 1 in the F2 soybean populations indicated that the resistance of both B4400 and B3802 soybean genotypes was controlled by an interaction of two genes which were located at different loci with a duplicate epistatic dominance. The dominant allele at both two loci masked the expression of recessive alleles at the other two loci. If the dominant resistant genes to the pest were B and C, while the recessive genes were b and c, then the interaction was gene B epistatic to genes C and c, and gene C epistatic to genes B and b. Hence, the probabilities of the combination of resistance genes in both soybean genotypes are: B_C_ = 9 (susceptible), B_cc = 3 (susceptible), bbC_ = 3 (susceptible), bbcc = 1 (resistance).
Table 1. Development of riptortus populations in two F2 populations of soybean crosses in the screen house at 1, 7, and 15 DAI
F2 Plant Population of: Number of Riptortus linearis (insects/25 plant) at:
1 DAI 7 DAI 15 DAI
Tambora x B-4400 8 21 20
Tambora x B-3802 7 20 47
Remarks: DAI: days after infestation.
Table 2. Variance of parents, environment, phenotype, genetic, and heritability value in two F2 populations of soybean crosses
F2 Populations σ2p1 σ2p2 σ2E σ2ph σ2G H2
Tambora x B4400 29.155 18.880 4.697 1186.901 1182.204 0.996 Tambora x B3802 29.155 11.191 4.372 477.645 473.273 0.991
Remarks: σ2p1 = variance of parent 1; σ2p2 = variance of parent 2; σ2E =variance of environment;σ2ph = variance of
Table 3. Frequency distribution of F2 populations of Tambora x B 4400 and Tambora x B 3802 based on scores of resistance to the soybean pod sucker.
Score Plant reaction Pest infestation (%)
Individual number of F2 population of:
Tambora x B 4400 Tambora x B 3802
0 Resistant 0 – 20 16 14
1 Moderately resistant 21 – 40 19 16 2 Moderately susceptible 41 – 60 36 33
3 Susceptible 61 – 80 70 78
4 Highly susceptible >80 78 85
Total 219 226
Table 4. Frequency ratio of F2 population of Tambora x B4400 and Tambora x B3802 cross combinations
Phenotypic grouping Calculated
2 of crosses:
2 table Tambora X B4400 Tambora X B3802Two-class grouping: P = 0.05 P = 0.01
3 : 1 18.28** 21.3** 3.84 6.63 9 : 7 103.43** 113.33**
13 : 3 7.89** 8.76**
15 : 1 0.04 ns 0.00015ns
Three-class grouping:
9 : 3 : 4 18.09** 30.11** 5.99 9.21 12 : 3 : 1 6.72** 1ns
9 : 6 : 1 14.34** 27.54** 10 : 3 : 1 20.92** 23.12** Four-class grouping:
9 : 3 : 3 : 1 17.30** 28.60** 7.81 11.34
Remarks : * : significant; ** : highly significant; tn : not significant, at 5% level of significance
In addition that the F2 frequency values of the F2 populations of Tambora x B 4400 and Tambora x B 3802 were suitable for the two-class grouping (15 : 1), Tambora x B 3802 crossed was also suitable for the three-class grouping (12:3:1). Based on the probability values from the calculated Chi-square (2), however, the 2 values
F2: Tambora x BB4400
F2: Tambora x BB3802
CONCLUSIONS
Populations of Riptortus linearis in each of the F2 population of Tambora x B4400 and Tambora x B3802 cross combinations were considered sufficient to separate resistance and susceptible plant segregation.
Heritability value of the two soybean cross combination (Tambora x B4400 and Tambora x B3802) was high (0.99), meaning that the resistance to the soybean pod sucker was controlled by the genetic factors. Selection for soybean resistance to pod sucker could be done at early generation of soybean crosses.
The resistance of soybean genotypes B4400 and B3802 to pod sucker was controlled by the interaction of two epistatic dominant genes located at different loci. With an assumption that B and C genes were dominant causing susceptible to pod sucker, hence the resistance genes found in both soybean genotypes were bbcc.
ACKNOWLEDGEMENTS
The authors wish to express their grati-tude to the Indonesian Agency for Agriculture Research and Development for providing the research budget, to Mr. Harnoto, MS, an entomologist, for his deliberate comments during the research and paper writing, as well as to sucking bugs. International Journal Tropical Insect Science 15: 337-341. Allard. R.W. 1966. Principle of plant breeding.
John Wiley an Sons, New York. pp. 485 Asadi, Soemartono, Woerjono, M., dan
Jumanto, H. 2003. Kendali genetik ketahanan kedelai terhadap penyakit virus kerdil (Sybean Stunt Virus). Zuriat. J.Pemuliaan Indonesia 14(2): 1-11, Cardoso de Codoi C.R. and J.B. Pinheiro. 2009.
Genetic parameters and selection strategies for soybean genotypes resistant to the stink bug-complex. Genet. Mol.Biol. 32(2). Sao Paulo. pp. 22
Chahal, G.S., and S.S. Gosal (2002): Principles and procedures of plant breeding: Biotechnological and conventional approaches. Third reprint 2006. Alpha Science International Ltd, Pangbourne RG8 8UT, UK. pp. 604
Ewete F.K. and O.A. Joda. 1996. The development of Riptortus dentipes Fabricius (hemiptera: alydidae) and damage caused on soybean. African Crop Science Journal 4(3): 345-350. Ferh, W.R. 1987. Principle of cultivar
develop-ment. Theory and technique. Vol 1. Mac Millan Publ., Cohen Stuart, New York. pp.536.
Gomez, K.A., and A.A. Gomez. 1984. Statistical procedures for agricultural research. John Wiley and Sons. New York, pp. 680.
Koswanudin D, dan T. Djuwarso. 1997. Per-kembangan pengisap polong kedelai R.linearis pada beberapa jenis tanaman inang. Tantangan Entomologi pada Abad XXI. Prosiding Seminar Nasional PEI, Bogor, 8 Januari 1997.
Marwoto dan Sri Hardaningsih. 2007. Pe-ngendalian hama terpadu pada tanaman kedelai. Dalam Kedelai, teknik produksi dan pengembangan. (Penyunting: Sumarno, Suyamto, AdiWijono, Hermanto dan Husni Kasim) Badan Litbang Pertanian. Puslitbang Tanaman Pangan Bogor. p. 296-318.
Petr F.C. and K.J. Frey. !966. Genotypic correlation dominance, and heritability of quantitative characters in oats. Crop Sci. 6:259-262.
Purwantoro, A., Asadi, M. Woerjono and Harnoto. 2008. Pendayagunaan sumberdaya gene-tik kedelai untuk perbaikan ketahanan terhadap hama pengisap polong. Laporan akhir kegiatan penelitian KKP3T Tahun 2008. Lembaga Penelitian dan Peng-abdian Kepada Masyarakat, Universitas Gajahmada (UGM), Yogyakarta pp.25. Purwanto. 2009. Laporan akhir kegiatan
peneli-tian KKP3T Tahun 2009. Lembaga Penelitian dan Pengabdian Kepada Masyarakat UGM, Yogyakarta.
teknik produksi dan pengembangan. Badan Litbang Pertanian. Puslitbang Tanaman Pangan Bogor. p. 1-27. Suharsono dan M. Yusuf. 2009. Analisis
generasi F2 seleksi pertama dari persilangan kedelai antara kultivar Slamet dan Wase. J. Agron. Indonesia 37(1): 21-27.
Taufik, S.A. dan A. Wijanarko. 2007. Peluang peningkatan produksi kedelai di Indonesia. (Penyunting: Sumarno, Suyamto, AdiWijono, Hermanto dan Husni Kasim) Dalam Kedelai, teknik produksi dan pengembangan. Badan Litbang Pertanian. Puslitbang Tanaman Pangan Bogor, p. 130-167.
Tengkano W, Sugito, A.M. Tohir and T. Okada. 1988. Pengujian ketahanan varietas kedelai terhadap serangan pengisap polong (Reptortus linearis F., Nezara viridula L. dan Piezodorus
rubrofascia-tus F. Seminar Balittan Bogor, 6 Desember 1988, Bogor.
Panizzi, A.R. 1987. Nutritional ecology of seed sucking insect of soybean and their management. Mem. Inst. Oewaldo Cruz. Rio de Janeiro, Vol 82, Sppl. II:161-175. Proyogo Y., T. Santoso dan Widodo. 2005.
Keefektifan cendawan entomopatogen dalam mengendalikan hama pengisap polong kedelai Riptortus linearis L., dan dampaknya terhadap predator Oxyopes javanus. Penelitian Pertanian Tanaman Pangan. 24 (2): 26-34
Tod J.W., 1982. Effect of stink bug damage on soybean quality in soybean seed quality and stand establishment. Insoy Series. 22: 46-51.