AKUMULASI ENZIM ANTIOKSIDAN DAN PROLIN PADA
BEBERAPA VARIETAS KEDELAI TOLERAN DAN PEKA
CEKAMAN KEKERINGAN
KHAIRUL ASHRI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2006
ABSTRAK
KHAIRUL ASHRI.
Akumulasi Enzim Antioksidan dan Prolin Pada
Beberapa Varietas Kedelai Toleran dan Peka Cekaman Kekeringan.
Dibawah bimbingan MUNIF GHULAMAHDI dan HAMIM.
Penelitian dengan judul Akumulasi Enzim Antioksidan dan Prolin Pada Beberapa Varietas Kedelai Toleran dan Peka Cekaman Kekeringan telah dilaksanakan sejak akhir bulan Mei 2005 hingga bulan Maret 2006, di laboratorium Fisiologi Tumbuhan, Departemen Biologi, dan laboratorium Biologi terpadu Fakultas Matematika dan Ilmu Pengetahuan Alam IPB Baranang Siang Bogor, serta laboratorium PSPT Departemen Agronomi dan Hortikultura Fakultas Pertanian - IPB Darmaga Bogor. Secara umum penelitian ini bertujuan untuk mempelajari akumulasi enzim antioksidan dan prolin serta responnya terhadap pertumbuhan serta produksi tanaman pada beberapa varietas kedelai toleran dan peka cekaman kekeringan.
Dalam penelitian ini diamati tingkat aktifitas enzim antioksidan yaitu enzim Glutation Peroksidase (GPX), Glutation reduktase (GR), dan Superoxida Dismutase (SOD), tingkat akumulasi prolin daun, pertumbuhan dan produksi tanaman pada 5 varietas kedelai budidaya (Glycine max L.) dan kedelai liar (Glycine tomentella) yang diberi perlakuan cekaman kekeringan. Perlakuan diberikan dengan menunda pengairan selama 14 hari pada kedelai budidaya dan 24 hari pada kedelai liar. Varietas yang digunakan adalah V1 (Tidar) dan V2 (Pangrango) yang merupakan varietas toleran cekaman kekeringan, V3 (Krakatau) dan V4 (Burangrang) yang merupakan varietas moderat cekaman kekeringan, V5 (Panderman) sebagai varietas peka cekaman kekeringan, dan V6 (Kedelai Liar).
Dari hasil penelitian diketahui bahwa cekaman kekeringan yang diberikan ketika tanaman memasuki awal fase generatif tidak menyebabkan terjadi interaksi antara faktor varietas yang digunakan dan faktor cekaman kekeringan yang diberikan pada semua pengamatan vegetatif. Untuk faktor tunggal, faktor varietas menunjukkan berbeda nyata pada semua peubah vegetatif, yaitu tinggi tanaman, jumlah cabang, jumlah daun trifoliat, bobot kering tajuk, serta panjang dan bobot
kering akar. Adapun pada faktor cekaman kekeringan, perbedaan hanya terjadi pada peubah bobot kering tajuk dan bobot kering akar.
Cekaman kekeringan selama 14 hari pengamatan akan menurunkan nilai RWC menjadi 43-33% pada varietas kedelai budidaya, sedangkan pada kedelai liar setelah 24 hari pengamatan terjadi penurunan nilai RWC menjadi sebesar 30%. Panderman (V5) sebagai varietas peka, memiliki nilai RWC terendah (33%) dibandingkan varietas moderat dan toleran. Hal ini menunjukkan bahwa RWC dapat dijadikan ukuran ketahanan tanaman terhadap cekaman kekeringan.
Tidak terjadi peningkatan aktifitas enzim antioksidan GPX, GR dan SOD secara signifikan pada tiap varietas. Peningkatan yang konsisten hanya terlihat pada kedelai liar (V6).
Pada pengamatan akumulasi prolin, terjadi peningkatan secara nyata pada semua tanaman setelah mengalami cekaman kekeringan. Peningkatan akumulasi prolin tertinggi ditunjukkan oleh varietas peka (V5), Data ini mendukung pernyataan bahwa prolin merupakan indikator beratnya cekaman yang dialami tanaman.
Surat Pernyataan
Dengan ini saya menyatakan bahwa tesis dengan judul :
“
Akumulasi Enzim Antioksidan dan Prolin Pada Beberapa Varietas
Kedelai Toleran dan Peka Cekaman Kekeringan
”adalah gagasan atau hasil penelitian saya sendiri di bawah bimbingan Komisi Pembimbing. Tesis ini belum pernah diajukan untuk memperoleh gelar pada program sejenis di Perguruan Tinggi lain.
Bogor, September 2006 Yang menyatakan,
PENDAHULUAN
Latar Belakang
Produk pertanian adalah sumber utama penghasil bahan pangan bagi kehidupan manusia. Kebutuhan terhadap bahan pangan akan terus meningkat seiring dengan semakin bertambahnya jumlah penduduk, oleh karena itu perlu dilakukan berbagai upaya untuk memenuhi kebutuhan tersebut diantaranya dengan cara meningkatkan produksi hasil pertanian.
Kedelai (Glycine max L.) termasuk komoditi tanaman pangan yang penting di Indonesia. Setiap tahun Indonesia masih harus mengimpor biji kedelai untuk memenuhi kebutuhan dalam negeri tak kurang dari 1,1 juta ton. Hal ini antara lain disebabkan terjadinya penyusutan lahan pertanian dan rendahnya produksi. Data yang dikeluarkan Badan Statistik Nasional (Februari 2003) menyebutkan bahwa kebutuhan kedelai nasional mencapai 2,2 juta ton per tahun dan akan terus meningkat. Akan tetapi kebutuhan kedelai baru dapat terpenuhi oleh produksi kedelai nasional pada tahun 2002 sebesar 0,65 juta ton, bahkan mengalami penurunan sekitar 21,06% dari tahun 2001 yang mencapai 0,83 juta ton biji kering. Menurut data BPS penurunan ini disebabkan oleh penurunan luas panen dan produktifitas yang baru mencapai 11,95 kwintal per hektar biji kering. Hal ini diperparah oleh semakin sedikitnya daerah yang mendapat pengairan akibat rusaknya jaringan irigasi teknis, sehingga semakin banyak daerah yang mengalami kekeringan (BPS 2003).
Sebagai solusi untuk peningkatan produksi kedelai nasional, maka pemerintah telah melakukan upaya intensifikasi dan ekstensifikasi. Intensifikasi dilakukan dengan mengatasi kendala yang terdapat pada areal penanaman yang sudah ada sehingga produksi dapat meningkat. Sebagai contoh pada lahan kering tadah hujan tanpa pengairan teknis, dilakukan upaya pemilihan dan penggunaan varietas yang tahan melalui proses seleksi benih yang dilakukan oleh para pemulia. Ekstensifikasi diperlukan dengan melakukan penanaman pada lahan-lahan kritis yang bermasalah akibat semakin berkurangnya lahan-lahan potensial yang dapat ditanami, diantaranya dengan menanam pada lahan marginal seperti pada lahan kering. Oleh sebab itu diperlukan juga upaya peningkatan adaptasi
2
ekofisiologis tanaman terhadap kondisi sub optimal lahan dan iklim agar keterbatasan tersebut tidak menjadi masalah dan produksi dapat berjalan optimal.
Kekeringan merupakan salah satu faktor penghambat utama produksi tanaman di seluruh dunia (Nemeth 2002), Hal ini sangat berpengaruh terhadap penurunan produksi kedelai, terutama pada awal dan pertengahan fase pengisian biji yang dapat menyebabkan polong hampa akibat terhambatnya proses fisiologis dan metabolisme seperti terhambatnya penyerapan unsur hara, berkurangnya hasil fotosintesis dan terhambatnya transportasi bahan hasil fotosintesis (Levitt 1980; Salisbury and Cleon 1991) .
Salah satu penyebab kerusakan tanaman pada kondisi kekeringan adalah terjadinya cekaman oksidatif yang disebabkan terakumulasinya senyawa Reactive
Oxygen Species (ROS) seperti singlet oksigen ( O2), hidrogen peroksida (H2O2),
superoksida (O2¯ ) dan radikal hidroksil ( OH), akibat terhambatnya proses
fotosintesis dengan menutupnya stomata sehingga menyebabkan kerusakan oksidatif pada organel fotosintesis (Prochazkova et al. 2001). Radikal ini dapat menyebabkan kerusakan melalui beberapa cara yaitu memutus ikatan rantai protein, merusak membran lemak dan bereaksi dengan DNA sehingga menyebabkan mutasi sel (Sgherri dan Navari-Izzo 1995).
Untuk mengatasi terjadinya cekaman oksidatif, tanaman memiliki mekanisme untuk meningkatkan ketahanannya diantaranya dengan meningkatkan pembentukan dan aktifitas enzim antioksidan seperti Glutation Peroksidase (GPX), Glutation reduktase (GR), Superoxida Dismutase (SOD), dan senyawa antioksidan lainnya yang dapat menyelamatkan tanaman dari ROS (Rhodes dan Samaras 1994). Kerusakan cekaman oksidatif terjadi apabila terdapat ketidakseimbangan antara kemampuan enzim antioksidan dan toksifikasi ROS (Rodriguez et al. 2002).
Selain enzim antioksidan tanaman juga meningkatkan mekanisme untuk menghadapi kekeringan dan cekaman oksidatif dengan mengakumulasi senyawa osmoprotektan dan larutan yang sesuai seperti prolin. Akumulasi prolin sudah sangat umum ditemukan pada tanaman yang mengalami cekaman osmotik seperti kekeringan, salinitas, serta suhu tinggi dan suhu rendah, sebagai upaya tanaman untuk melindungi enzim dari proses denaturasi. Selain itu prolin juga dapat
3
berinteraksi dengan sistem membran, mengatur keseimbangan kemasaman sitosol dengan perbandingan NADH/NAD+ berfungsi sebagai sumber energi dan membantu sel untuk menghadapi cekaman oksidatif, oleh karena itu prolin disebut sebagai osmoprotektan. (Konstantinova et al. 2002).
Tiap varietas tanaman mungkin memiliki reaksi yang sangat kompleks menghadapi cekaman kekeringan, yang ditunjukkan oleh bentuk morfologi, anatomi dan metabolisme tanaman yang berbeda. Oleh karena itu perlu dipelajari berbagai mekanisme yang dilakukan tanaman agar tetap dapat tumbuh dan berproduksi dengan baik walau dalam kondisi cekaman kekeringan.
Berdasarkan masalah ini maka dilakukan penelitian mengenai Akumulasi Enzim Antioksidan dan Prolin Pada Beberapa Varietas Kedelai Toleran dan Peka Cekaman Kekeringan.
Tujuan
Penelitian ini bertujuan untuk mengamati tingkat aktivitas enzim Glutation Peroksidase (GPX), Glutation Reduktase (GR), dan Superoxida Dismutase (SOD), dan tingkat akumulasi prolin daun selama cekaman kekeringan serta responnya terhadap pertumbuhan serta produksi tanaman.
Hipotesis
Tiap varietas kedelai kekeringan yang terkena cekaman oksidatif akibat cekaman kekeringan, mensintesis enzim Glutation Peroksidase (GPX), Glutation Reduktase (GR), dan Superoxida Dismutase (SOD), kadar prolin, Kandungan Air relatif daun (RWC), pertumbuhan dan produksi hasil tanaman yang berbeda
Varietas kedelai toleran kekeringan yang terkena cekaman oksidatif akibat cekaman kekeringan, mensintesis sejumlah enzim Glutation Peroksidase (GPX), Glutation Reduktase (GR), dan Superoxida Dismutase (SOD), kadar prolin, dan Kandungan Air relatif daun (RWC), serta memiliki pertumbuhan dan produksi hasil tanaman yang lebih besar dibandingkan varietas peka kekeringan setelah mengalami fase kekeringan
4
TINJAUAN PUSTAKA
Morfologi Tanaman Kedelai
Kedelai merupakan salah satu tanaman penting di Indonesia karena memiliki kandungan gizi yang cukup tinggi dibandingkan jenis tanaman serealia lainnya. Kedelai memiliki kandungan protein sebesar 35%, lebih tinggi dibandingkan padi yang hanya sebesar 7%. Selain itu kedelai juga mengandung asam amino seperti metionin, tripsin dan lisin yang cukup tinggi sehingga dapat diandalkan untuk memenuhi kebutuhan gizi dan bahan pangan bagi manusia (Suprapto 1997).
Tanaman kedelai termasuk dalam divisi Spermatophyta, sub divisi Angiospermae, kelas Dicotyledoneae, ordo Rosales, famili Leguminosae, genus Glycine dan spesies Glycine max (L.) Merril. Tanaman kedelai umumnya tumbuh tegak, berbentuk semak dan merupakan tanaman semusim. Morfologi tanaman kedelai didukung oleh komponen utamanya yaitu akar, daun, batang, polong, dan biji sehingga pertumbuhannya bisa optimal.
Kedelai dapat tumbuh disetiap jenis tanah, akan tetapi untuk mencapai tingkat pertumbuhan dan produksi yang optimal, maka kedelai harus ditanam pada jenis tanah berstruktur lempung berpasir atau liat berpasir, sebab hal ini berhubungan dengan ketersediaan air untuk mendukung pertumbuhan. Kedelai cukup toleran terhadap cekaman kekeringan, karena masih bertahan dan berproduksi pada kondisi cekaman kekeringan maksimal 50 persen dari kapasitas lapang atau kondisi tanah yang optimal, akan tetapi kedelai memiliki fase kritis dalam pertumbuhannya yaitu pada saat perkecambahan, masa berbunga da pengisian polong yang nantinya akan mempengaruhi jumlah produksi (Adisarwanto 2005).
Cekaman kekeringan Bagi Tanaman
Air memiliki peranan yang sangat penting dalam kehidupan tanaman. Setiap gram pembentukan bahan organik penyusun tanaman rata-rata membutuhkan 500 g air yang diabsorbsi oleh akar ditransportasikan ke seluruh bahagian tanaman, dan selanjutnya air akan hilang ke atmosfir.
5
Setiap tanaman harus dapat menyeimbangkan antara proses kehilangan air dan proses penyerapannya, bila proses kehilangan air tidak diimbangi dengan penyerapan melalui akar maka akan terjadi kekurangan air didalam sel tanaman yang dapat menyebabkan berbagai kerusakan pada banyak proses dalam sel tanaman (Taiz dan Zeiger 2002).
Menurut Adisarwanto (2005) jumlah air yang dibutuhkan tanaman untuk pertumbuhannya berbeda-beda pada tiap jenis tanaman. Pada tanaman kedelai jumlah air yang dibutuhkan selama hidupnya sebesar 300 – 650 mm, bila kebutuhan ini tidak terpenuhi maka proses pertumbuhan dan perkembangan tanaman akan terganggu.
Cekaman lingkungan merupakan tantangan utama dalam memproduksi tanaman secara berkelanjutan. Dalam menghadapai kondisi stress lingkungan, tanaman memberikan respon secara fisiologi dan biokimia, sebagai upaya untuk menerima, menghindari dan menetralisir pengaruh dari cekaman. Sifat peka dan toleran suatu tanaman bergantung kepada sifat genetik dan respon biokimia yang dimiliki suatu spesies, sehingga selama bertahun-tahun para peneliti memfokuskan diri untuk membangun adaptabilitas tanaman secara genetik dan biokimia menghadapi berbagai kondisi cekaman lingkungan (Dubey 1995).
Cekaman kekeringan merupakan pengaruh faktor lingkungan yang menyebabkan air tidak tersedia bagi tanaman, yang dapat disebabkan antara lain oleh tidak tersedianya air di daerah perakaran tanaman dan permintaan air yang besar di daerah daun dimana laju evaporasi melebihi laju absorbsi air oleh akar tanaman (Hamim 2004).
Ketika jumlah absorbsi air mulai terbatas, maka tanaman memiliki mekanisme untuk mencegah kehilangan air dengan melakukan penutupan stomata. Perubahan pada ketahanan mekanisme stomata sangat diperlukan untuk mengatur kehilangan air oleh tanaman dan untuk mengatur pengambilan karbon dioksida (CO2) yang penting untuk ketersediaan fiksasi CO2 selama proses
fotosintesis (Taiz dan Zeiger 2002).
Relative Water Content (RWC) yang menggambarkan kadar relatif air
daun merupakan parameter ketahanan tanaman menghadapi cekaman kekeringan. Proses fotosintesis pada sebagian besar tanaman akan mulai tertekan bila nilai
6
RWC tanaman lebih rendah dari 70 persen, sehingga tanaman memerlukan pengaturan dalam tubuhnya diantaranya dengan melakukan penutupan stomata (Quilambo 2004).
Mekanisme penutupan stomata selain dipengaruhi oleh ABA juga dipengaruhi oleh adanya penurunan konduktansi hidrolik daun (leaf hidraulic
conductance) sehingga terjadi pengurangan tekanan turgor daun sebagai upaya
penyelamatan proses fotosintesis agar tetap dapat berjalan (Broodribb dan Holbrook 2003).
Tanaman memiliki reaksi yang sangat kompleks menghadapi cekaman kekeringan. Bentuk morfologi, anatomi dan metabolisme tanaman yang berbeda menyebabkan tanaman memiliki respon yang beragam. Ketika kekeringan semakin meningkat maka tanaman menyesuaikan diri melalui proses fisiologi yang kemudian diikuti perubahan struktur morfologi tanaman seperti layu, meningkatkan pertumbuhan akar dan menghambat pertumbuhan pucuk. Penurunan pertumbuhan vegetatif menyebabkan penurunan proses fotosintesis dan pertumbuhan, sehingga tanaman juga mengalami penurunan produksi seperti berkurangnya hasil panen secara kualitas maupun kuantitas (Taiz dan Zeiger 2002).
Pada kondisi cekaman, tanaman tidak dapat mengekspresikan potensial genetiknya secara penuh. Berbagai kondisi cekaman lingkungan menyebabkan perubahan penting pada ekspresi gen tanaman, terjadinya peningkatan akumulasi ion anorganik, terjadinya perubahan pada sintesis protein dengan mengeluarkan protein baru yang spesifik pada kondisi cekaman tertentu, serta perubahan prilaku banyak enzim (Patakas et al. 2002).
Cekaman Oksidatif dan Enzim Antioksidan
Reactive Oxygen Species (ROS) merupakan radikal bebas yang sangat
berbahaya bagi makhluk hidup. Pada tanaman ROS terbentuk dalam sel melalui beberapa cara yaitu : 1. Produksi fotokimia di atmosfer akibat pencemaran udara, 2. Penyumbangan elektron langsung ke oksigen ketika terjadi fotosintesis terutama pada kondisi cahaya yang tinggi dan konsentrasi CO2 pada kloroplas
7
yang rendah. 3. Respon terhadap kondisi cekaman seperti kekeringan, suhu tinggi, salinitas, ozon dan serangan mikroba (Pritchard et al. 2000).
ROS yang terbentuk akan berbahaya bagi sel tanaman karena ia dapat mengoksidasi membran lipid dan aparatus fotosintesis. Melalui berbagai reaksi metabolisme tanaman oksigen dapat diubah menjadi bentuk molekul yang sangat reaktif seperti singlet oksigen ( O2), hidrogen peroksida (H2O2), superoksida
(O2¯ ) dan radikal hidroksil ( OH). Superoksida dapat berubah bentuk menjadi
hidrogen peroksida (H2O2), radikal ini dapat menyebabkan kerusakan melalui
beberapa cara yaitu memutus ikatan rantai protein, merusak membran lemak dan bereaksi dengan DNA sehingga menyebabkan mutasi sel (Sgherri dan Navari-Izzo 1995).
Anion superoksida juga dapat terbentuk di dalam tanaman dari proses autooksidasi dengan adanya oksigen , transport elektron didalam fotosintesis dan bereaksi dengan NAD(P)H dalam membran glioksisom dan peroksisomal, yang dapat merusak jaringan tanaman (Borsani et al. 2001).
Terbentuknya H2O2 di kloroplas terjadi karena terhambatnya proses
fotosintesis menyebabkan tanaman kelebihan energi elektron yang ditangkap oleh pusat reaksi tidak dapat dilepas atau dipantulkan secara aman sehingga merusak perangkat Fotosistem II pada kloroplas dan membran lipid (Kader 2001).
Dari hasil penelitian Pedregosa et al. (2002) pada akar tanaman bawang (Allium cepa) hidrogen peroksida ditemukan pada lokasi yang berbeda, akan tetapi yang utama ia banyak ditemukan pada dinding sel dari epidermal dan sel meristematik, serta pada sel yang mengalami lignifikasi. Dari hasil penelitian Rodriguez et al. (2002) pada tanaman jagung, diketahui bahwa ROS diproduksi dan banyak terdapat pada bagian yang aktif masih membelah.
Prochazkova et al. (2001) menerangkan bahwa cekaman kekeringan diketahui dapat menyebabkan kerusakan oksidatif pada tanaman dengan menginduksi pembentukan senyawa ROS. Hal ini karena tanaman yang terkena cekaman kekeringan akan menginduksi ABA sehingga ia dapat menutup stomatanya untuk mengurangi laju transpirasi. Stomata yang tetutup menyebabkan rendahnya kandungan CO2 pada kloroplas tidak dapat melepaskan
8
superoxide anion (O2–) yang menyebabkan kerusakan oksidatif pada organel
fotosintesis.
Sebagai upaya untuk mengurangi dampak kerusakan sel akibat cekaman oksidatif maka tanaman membentuk enzim senyawa antioksidan (lihat Gambar 1) yang dapat mengkatalisis reaksi untuk mengurangi tingkat detoksifikasi ROS secara langsung maupun tidak langsung di dalam sel tanaman (Pritchard et al. 2000). Telah banyak penelitian yang membuktikan terjadinya peningkatan aktivitas enzim antioksidan pada berbagai tanaman (Borsani et al. 2001)
Gambar 1. Ringkasan sistem enzim dapat bekerja dalam menetralisir racun dari ROS pada mitokondria tanaman Ket: ETC; electron transport chain; ICDH; isocitrate dehydrogenase; MnSOD; Mn-superoksida dismutase; Trx; thioredoxin (Pritchard et al. 2000).
Enzim antioksidan yang dibentuk oleh tanaman diantaranya adalah Glutation Peroksidase (GPX), Glutation reduktase (GR), Superoxida Dismutase (SOD), Ascorbat Peroksidase (APOD), selain itu terdapat juga enzim Katalase,
Kerusakan ETC O2 + H2O H2O H2O H2O Sistem GPX
Katalase Mn SOD Enzim transhidrogenase Sistem TRX reduktase H2O Sistem Askorbat/ Glutation
9
Peroxidase, Dehydroascorbate Reduktase (DAR), Monodehydroascorbate Reductase (MDAR) (Pritchard et al. 2000).
Sistem antioksidan didalam sel tumbuhan menyediakan perlindungan melawan pengaruh racun dari oksigen spesies yang aktif. Komponen penting dari sistem pelindungan itu adalah pertahanan secara enzimatis, seperti SOD dan katalase yang dapat menghindari O2 - dan H2O2 selain metabolit seperti askorbat,
glutation dan tokoperol yang berfungsi untuk mengatur tingkat keaktifan oksigen pada jaringan tanaman (Bosch dan Alegre 2002).
Pada tanaman tahunan diketahui terdapat 3 bentuk SOD. Ketiga bentuk SOD ini berbeda dalam ko-faktor logam dan letaknya pada sub seluler. SOD yang terletak pada sitosol dan kloroplas yaitu CuZn SOD, pada mitokondria yaitu Mn SOD dan pada kloroplas yaitu Fe SOD (Aroca et al. 2001).
Untuk melindungi aparatus fotosintesis dari kerusakan oksidatif, tanaman harus melepaskan kelebihan energi cahaya yang diterima. Perlindungan dapat terjadi dengan penurunan efisiensi fotokimia lewat siklus xanthophyl atau dengan pengaturan aliran elektron yang melibatkan lintasan alternatif seperti fotorespirasi dan reaksi peroksidasi mehler. Lintasan fotorespirasi dan siklus xanthophyl dapat menyebabkan peningkatan jumlah produksi spesies oksigen yang dapat meracuni tanaman berupa O2- dan H2O2 (Bosch dan Alegre 2002).
Selain itu pada beberapa spesies tanaman juga dilaporkan terjadi peningkatan sintesis sukrosa dan prolin pada kondisi cekaman air yang disebabkan meningkatnya aktifitas enzim yang mensintesis metabolit tersebut. (Prochazkova 2001).
Akumulasi asam amino prolin
Prolin merupakan asam amino bebas yang disintesis tanaman dalam jaringan floem, akar dan biji (Simpson 2001). Prolin merupakan asam amino yang paling stabil dan paling sedikit menghambat pertumbuhan tanaman dibandingkan asam amino lainnya (Levitt 1980).
Pada kondisi cekaman kekeringan dan berbagai cekaman osmotik lainnya, beberapa tanaman memiliki mekanisme adaptasi berupa kemampuan untuk mensintesis senyawa osmoprotektan atau larutan yang sesuai (Ronde et al. 2000).
10
Osmoprotektan merupakan larutan yang tidak beracun sehingga dapat diakumulasi sampai batas tertentu tanpa mengganggu metabolisme tanaman, biasanya terdiri dari beberapa grup asam amino (Rhodes dan Samaras 1994).
Akumulasi prolin sebagai respon terhadap cekaman osmotik telah umum diketahui (Konstantinova et al. 2002). Telah banyak peneliti yang menemukan bahwa tanaman yang terkena cekaman kekeringan akan mengakumulasi asam amino prolin dalam jumlah tertentu dan bervariasi tergantung pada jenis tanaman, umur, dan varietas tanaman yang digunakan (Hamim 2004).
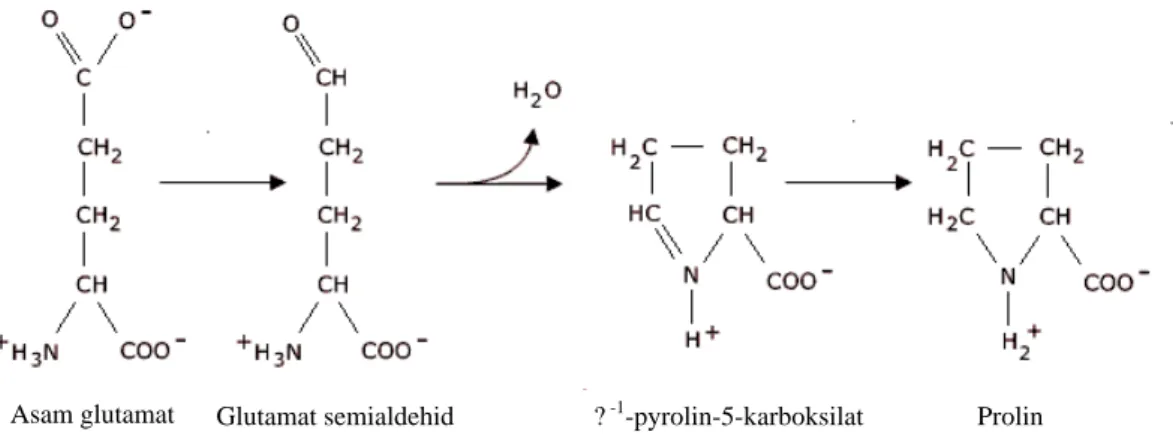
Gambar 2. Skema biosintesis asam amino prolin
Seperti pada Gambar 2 di atas, prolin disintesis oleh enzim ?-1 -pyrroline-5-carboxylate synthetase (P5CS) dari asam glutamat. Pada awalnya, gugus karboksil direduksi menjadi aldehid menghasilkan glutamat semialdehid, lalu gugus aldehid bereaksi dengan grup a-amino, menghilangkan gugus air. Akhirnya pada tahap reduksi kedua barulah terbentuk prolin
Dari beberapa hasil penelitian pada tanaman kedelai, prolin lebih banyak dijumpai pada varietas yang toleran kekeringan sebagai upaya tanaman untuk meningkatkan keseimbangan osmotik pada kondisi kekeringan. Akan tetapi beberapa penelitian juga menyebutkan bahwa prolin diakumulasi sebagai respon tanaman terhadap berbagai kondisi cekaman osmotik yang sangat umum terjadi pada tanaman (Delauney dan Verma 1993).
Berbagai penelitian membuktikan bahwa akumulasi prolin memiliki berbagai keuntungan bagi sel. Prolin dapat berfungsi sebagai sumber energi, nitrogen dan karbon, dan sebagai osmolit sebagai respon dari kekeringan, selain itu prolin juga dapat mengurangi radikal bebas di dalam sel sehingga dapat
Glutamat semialdehid
11
mencegah kerusakan akibat cekaman oksidatif (Hong et al. 2000). Prolin juga terlibat dalam perlindungan enzim dari denaturasi, dapat berinteraksi dengan sistem membran, mengatur kemasaman (pH) sitosol, mengatur keseimbangan antara NADH/NAD+ (Konstantinova et al. 2002).
Berbagai penelitian telah banyak dilakukan sebagai upaya untuk membentuk varietas tanaman toleran terhadap berbagai kondisi cekaman dengan memasukkan gen P5CS sehingga tanaman akan mensintesis kandungan prolin yang lebih baik dari tanaman tetuanya. Hasil penelitian yang dilakukan oleh Konstantinova et al. (2002) pada tanaman tembakau dengan membentuk tanaman transgenik yang diinduksi gen P5CS dapat meningkat sintesis kandungan prolin hingga 5 kali dari tanaman kontrol pada kondisi cekaman kekeringan.
Cekaman lingkungan dapat menginduksi kerusakan cekaman oksidatif, oleh karena itu diperlukan berbagai penelitian untuk menentukan respon tanaman pada kondisi cekaman sebab tanaman akan selalu dipengaruhi oleh berbagai faktor cekaman lingkungan (Keles et al. 2002)
12
BAHAN DAN METODE
Waktu dan Tempat
Penelitian ini dilakukan pada akhir bulan Mei 2005 hingga bulan Maret 2006, di laboratorium Fisiologi Tumbuhan, Departemen Biologi, dan laboratorium Biologi Terpadu Fakultas Matematika dan Ilmu Pengetahuan Alam IPB Baranang Siang Bogor, serta laboratorium PSPT Departemen Agronomi dan Hortikultura Fakultas Pertanian - IPB Darmaga Bogor
Metode Penelitian
Rancangan yang digunakan dalam penelitian ini adalah Rancangan Acak Lengkap (RAL) dalam faktorial, terdiri atas 2 faktor dan 5 ulangan (Layout penelitian dapat dilihat pada Lampiran 1). Faktor utama adalah 6 varietas kedelai yang terdiri dari varietas toleran, moderat dan peka cekaman kekeringan (Deskripsi varietas dapat dilihat pada Lampiran 2). 6 varietas yang digunakan adalah:
V1 : Varietas kedelai Tidar (toleran cekaman kekeringan) V2 : Varietas kedelai Pangrango (toleran cekaman kekeringan) V3 : Varietas kedelai Krakatau (moderat cekaman kekeringan) V4 : Varietas kedelai Burangrang (moderat cekaman kekeringan) V5 : Varietas kedelai Panderman (peka cekaman kekeringan) V6 : Kedelai liar (toleran cekaman kekeringan)
Faktor kedua adalah cekaman kekeringan yang terdiri dari 2 taraf yaitu : A1 : Tanpa diberi perlakuan cekaman (kontrol)
A2 : Diberi perlakuan cekaman kekeringan sampai fase kritis tanaman, yaitu 14 hari pada varietas kedelai budidaya dan 14 hari pada kedelai liar.
Model linear yang digunakan adalah sebagai berikut : Y ijk = + i + j + ( )ij + ijk
dimana :
13
tingkat cekaman taraf ke j
i = 1, 2, 3 . . . 6 (faktor varietas kedelai) j = 1, 2 (faktor kekeringan)
= Nilai rata-rata umum
i = Pengaruh varietas tanaman ke-i
j = Pengaruh faktor tingkat cekaman kekeringan taraf ke-j
( )ij = Pengaruh varietas tanaman ke i dan perlakuan tingkat cekaman kekeringan ke-j
ijk = Galat percobaan
Selanjutnya dilakukan analisis ragam (Anova), apabila berpengaruh nyata maka akan dilakukan analisis lanjutan dengan menggunakan uji Duncan Multiple Range Test (DMRT).
Pelaksanaan Penelitian Bahan Tanaman
Bahan tanaman dari 6 varietas kedelai ditanam pada polibag berdiameter 20 cm yang diisi campuran pasir dan tanah yang dihaluskan dengan perbandingan 2:1 sebanyak 8 kg yang telah diberikan pupuk NPK sebanyak 3,5 g per polibag, dimana 1,5 g diberikan sebagai pupuk dasar dan 2 g ketika tanaman telah berumur 3 minggu. Sebelum ditanamai terlebih dahulu dilakukan analisis fisik dan kimia tanah untuk mengetahui kadar air tanah pada keadaan kapasitas lapang, serta kandungan hara dan pH tanah.
Sebelum diberikan perlakuan semua polibag diberikan air sampai kapasitas lapang setiap hari hingga tanaman memasuki awal fase reproduktif (1 bulan setelah tanam) agar diyakini bahwa tanaman sudah memiliki pertumbuhan yang stabil. Selanjutnya dilakukan dua kelompok perlakuan pemberian air, dimana kelompok pertama merupakan tanaman kontrol yang mendapat penyiraman setiap hari sehingga tidak mengalami cekaman kekeringan, sedangkan kelompok kedua adalah tanaman yang diberikan cekaman kekeringan dengan menunda pemberian air hingga tanaman sudah mulai menunjukkan gejala layu berat (14 hari pada tanaman budidaya dan 24 hari pada tanaman kedelai liar).
14
Ketika tanaman sudah menunjukkan gejala mulai memasuki fase kritis (layu berat), tanaman diberi air kembali untuk mengamati pemulihannya dari kondisi cekaman kekeringan, dan mengamati produksi tanaman. Selama perlakuan berlangsung, permukaan tanah polibag perlakuan kekeringan ditutupi mulsa plastik untuk mencegah terjadinya kehilangan air melalui evaporasi.
Pengambilan Bahan Analisis
Pada varietas tanaman kedelai budidaya, pengambilan bahan sampel perlakuan cekaman dan kontrol untuk analisis enzimatis dan kandungan prolin daun dilakukan pada hari ke 0, 4, 8, 12, dan 14, setelah perlakuan cekaman kekeringan (HSP). Pada tanaman kedelai liar sampel yang diambil adalah pada hari ke 0, 4, 8, 10, 14, 16, 20, dan 24 setelah perlakuan cekaman kekeringan (HSP). Sampel yang diambil adalah daun ketiga dari pucuk tanaman yang telah berkembang sempurna (fully expanded leaf).
Sampel daun diambil dan ditimbang menggunakan timbangan analitik seberat 0,2 g untuk analisis enzim GPX, GR dan SOD, dan 0,5 g untuk analisis prolin. Sampel yang telah diambil dibungkus dengan alumunium foil dan dibekukan dalam nitrogen cair (N2) untuk menghentikan proses enzimatis dan kerusakan metabolisme yang terjadi, kemudian sampel bahan dipindahkan kedalam freezer pada suhu -30 0C sampai bahan siap dianalisis.
Dekstruksi tanaman dan media dilakukan untuk mengamati bobot basah dan kering akar, bobot basah dan kering tajuk, RWC, dan analisis kandungan air tanah (KAT).
Pengamatan
Pengamatan yang dilakukan meliputi pengamatan terhadap pertumbuhan tajuk tanaman, pertumbuhan akar, Relatif Water Content (RWC), Kadar Air Tanah (KAT), analisis kandungan enzim antioksidan, analisis prolin, analisis klorofil dan produksi tanaman.
15
1. Pertumbuhan tajuk tanaman
Pengamatan pertumbuhan tanaman dilakukan ketika tanaman telah mendapat perlakuan cekaman kekeringan pada umur 65 hari. Pengamatan yang dilakukan meliputi:
a. Tinggi tanaman b. Jumlah daun trifoliat c. Jumlah cabang d. Bobot kering tajuk 2. Pertumbuhan akar
Pertumbuhan akar diamati dengan melakukan destruksi pada tanah didalam polibag yang telah diambil tajuk dan daunnya dengan hati-hati, agar tidak merusak sistem perakaran tanaman. Pengamatan akar yang dilakukan adalah :
a. Panjang akar b. Bobot kering akar.
3. Relative Water Content (RWC)
Pengukuran RWC dilakukan dengan mengambil bagian daun tanaman yang telah mengembang sempurna (fully expanded leaf) menggunakan kock bor sehingga terbentuk lingkaran daun berdiameter 1 cm sebanyak 10 lembar. Bagian daun ini ditimbang berat basahnya, lalu dilakukan perendaman dalam air selama 24 jam dan selanjutnya ditimbang berat jenuhnya, untuk mengetahui besarnya air maksimum yang dapat diserap jaringan. Selanjutnya daun dikering oven pada suhu 80 0C selama 48 jam dan diukur bobot keringnya. Untuk mengetahui persentase nilai RWC digunakan persamaan Prochazkova et al. (2001) yaitu: RWC = berat basah – berat kering x 100
berat jenuh – berat kering 4. Pengukuran kadar air tanah (KAT)
Kadar air tanah diukur selama perlakuan cekaman kekeringan, yang diamati ketika dilakukan destruksi sampel yaitu pada hari ke 0, 4, 8, 10, 12, dan 14 pada varietas tanaman budidaya dan pada hari ke 0, 4, 8, 10, 14,16, 20, dan 24 pada kedelai liar setelah perlakuan kekeringan. Pengamatan dilakukan dengan
16
mengambil sampel masing-masing sebanyak 50 g tanah pada bagian atas, tengah dan bawah dari tiap-tiap polibag. Tanah dibungkus dengan kertas dan dioven pada suhu 70 0C selama 72 jam, lalu ditimbang bobot keringnya dan diambil angka rata-ratanya.
5. Aktifitas senyawa dan enzim antioksidan
Daun tanaman seberat 0,2 g ditambahkan 50 mM Bufer Fosfat (pH 7.0), 1% (w/v) polyvinyl polypyrrolydone dan 2 mM asam askorbat (ASA). Sampel dihaluskan dalam mortar sampai diperoleh homogenat sebanyak 4 ml. Homogenat disentrifuse pada 15.000 g selama 30 detik pada suhu 4 0C. Selanjutnya supernatan dipisahkan dan siap dianalisis.
a. Enzim Glutation Peroxidase (GPX)
Pengamatan aktifitas enzim GPX dilakukan berdasarkan metode yang dilakukan oleh Jiang dan Huang (2001). Untuk mengukur aktivitas enzim GPX, maka 0,5 ml supernatan hasil ekstraksi ditambahkan kedalam tabung reaksi yang telah diisi 0,2 M Bufer fosfat (pH 7,0), 0.5 mM EDTA, 10 mM NaNO3 0,1 ml, 10 mM GSH 0,1 ml, 1.6 mM NADPH 0,1 ml, dan 2,5 unit GR
0,1 ml. Tabung diletakkan pada alat spektrofotometer, selanjutnya pada tabung ditambahkan 4 mM H2O2 0,1 ml dan diukur tingkat oksidasi
NADPH.
Aktivitas GPX ditentukan melalui pengukuran menggunakan spektrofotometer berdasarkan kepada oksidasi NADPH pada panjang gelombang 340 nm. Aktivitas enzim diekspresikan sebagai nmol dari NADPH yang dioksidasi per mg protein per menit
b. Enzim Glutation Reduktase (GR)
Pengamatan aktifitas enzim GPX dilakukan berdasarkan metode yang dilakukan oleh Cakmak dan Marschner (1993). Untuk mengukur aktivitas enzim GR, 0,15 ml supernatan ditambahkan kedalam tabung reaksi yang telah berisi 1 mM EDTA, 0,5 mM Glutation disulfida (GSSG), 0,15 mM NADPH dan 100 mM buffer Sodium Fosfat (pH 7,8). Selanjutnya diukur tingkat oksidasi NADPH yang terjadi menggunakan spektrofotometer pada panjang gelombang 340 nm
17
Aktivitas GR ditentukan melalui pengukuran menggunakan spektrofotometer berdasarkan kepada oksidasi NADPH pada panjang gelombang 340 nm. Aktivitas enzim diekspresikan sebagai nmol dari NADPH yang dioksidasi per mg protein per menit.
c. Enzim Superoxida Dismutase (SOD)
Pengamatan aktifitas enzim SOD dilakukan berdasarkan metode yang dilakukan oleh Jiang dan Huang (2001). Untuk mengukur aktivitas enzim SOD, 0,05 ml supernatan ditambahkan kedalam tabung reaksi yang telah berisi 0,2 mL EDTA 0,1 M, 0,1 mL Nitroblue tetrazolium (NBT), serta buffer fosfat 0,0067 M pada pH 7,8. hingga diperoleh campuran sebanyak 3 ml.
Setelah itu campuran tersebut diinkubasikan kedalam kotak cahaya dengan diberi penerangan lampu Neon 20 watt selama 3 menit, dimana sebelumnya kotak telah dipanaskan selama 15 menit. Setelah itu campuran ditambahkan 0,05 ml riboflavin 0,12 mM dan diinkubasikan kembali. Pengukuran dilakukan setiap 1 menit dengan mengukur tingkat absorbansinya pada panjang gelombang 560 nm.
6. Analisis prolin
Analisis prolin dilakukan menggunakan metode modifikasi Bates (1973), dengan menggunakan spektrofotometer dengan prolin murni sebagai standar. Asam ninhydrin disiapkan sebagai pereaksi dengan melarutkan 1 g ninhidrin dalam 30 ml asam asetat glasial dan 20 ml 6 mol asam asetat. Larutan didinginkan dan disimpan selama 24 jam hingga pereaksi ini siap digunakan. Daun sampel tanaman sebanyak 0,05 g, diekstraksi dalam 10 ml asam sulfosalisilik 3% menggunakan mortar, lalu disentrifuse dengan kecepatan 6000 rpm selama 5 menit dan diambil supernatannya.
Setelah dipisahkan, supernatan diterra sebanyak 10 ml, 2 ml cairan sampel diambil dan direaksikan dengan 2 ml asam ninhidrin dan 2 ml asam asetat glasial. Setelah itu tabung reaksi dipanaskan selama 1 jam pada suhu 100 0C, kemudian proses reaksi diakhiri dalam ice-bath. Campuran ini selanjutnya diekstraksi dengan 4 ml toluen, dikocok menggunakan test tube stirrer selama 15 – 20 detik. Larutan toluen dipisahkan dari endapan yang terbentuk, lalu diukur absorbansinya
18
= ( µg prolin/ml x ml toluen) / 115,5 µg/ µmol (g sampel) / 5
= µ mol prolin / g bobot basah
pada panjang gelombang 520 nm, untuk blanko digunakan toluen. Konsentrasi prolin ditentukan dari kurva standar dan dihitung berdasarkan berat segar yaitu :
[
7. Analisis klorofil
Analisis klorofil dilakukan ketika tanaman yang diberi perlakuan cekaman sudah mencapai tahap kritis yaitu pada hari ke 14 sebelum dilakukan recovery. Daun tanaman diambil dan ditimbang menggunakan timbangan analitik seberat 0,05 g, lalu daun dihaluskan menggunakan mortar setelah ditambahkan 2 ml aseton 80%. Selanjutnya homogenat diambil sebanyak 2 ml dan dimasukkan kedalam mikrofilter dan di sentrifuse pada 5.410 g selama 20 detik untuk memisahkan supernatan dari bahan tanaman (daun). Ekstraksi dilakukan beberapa kali sampai bahan tidak berwarna lagi. Supernatan ditera sebanyak 10 ml dan diamati menggunakan spektrofotometer pada panjang gelombang 645 nm untuk mengamati klorofil A dan 663 nm untuk klorofil B.
Kandungan klorofil A dan B diketahui dengan menggunakan rumus : klorofil A (mg/mg sampel) = {(12,7 x A 663) – (2,69 x A 645)} x Fp
Bobot sampel (mg)
klorofil B (mg/mg sampel) = {(22,9 x A 645) – (4,68 x A 645)} x Fp Bobot sampel (mg)
dimana Fp (faktor pengencer) = 10 ml x 1 liter
1000 ml
8. Produksi
Nilai produksi hasil tanaman diamati setelah dilakukan panen ketika tanaman sudah menunjukkan gejala matang fisiologis. Pengamatan yang dilakukan adalah :
a. Jumlah polong isi pertanaman b. Jumlah polong hampa
c. Jumlah biji pertanaman d. Bobot total biji
19
HASIL DAN PEMBAHASAN
Respon Pertumbuhan Tanaman
Dari enam varietas yang diamati, terlihat perbedaan pertumbuhan antara kedelai varietas budidaya dengan kedelai liar. Kedelai liar memiliki pertumbuhan yang lebih lambat, waktu berbunga yang lebih panjang, serta memiliki morfologi batang dan daun yang lebih kecil dibandingkan varietas budidaya. Akan tetapi kedelai liar memiliki ketahanan yang lebih baik menghadapi cekaman kekeringan. Hal ini ditunjukkan dari lamanya tanaman mencapai fase kritis setelah diberi perlakuan cekaman kekeringan yang mencapai 24 hari setelah perlakuan, berbeda dengan tanaman budidaya yang sudah memasuki fase kritis setelah 14 hari perlakuan. Oleh sebab itu terjadi waktu pengamatan yang berbeda antara varietas kedelai budidaya dan kedelai liar.
Perlakuan cekaman kekeringan yang diberikan kepada tanaman menyebabkan terjadinya perubahan morfologi yang berbeda pada tiap varietas. Perubahan morfologi akibat cekaman kekeringan biasanya sangat tergantung pada faktor waktu terjadinya cekaman dan besarnya perlakuan cekaman (Keles dan Oncel 2002).
Dari data pertumbuhan vegetatif tanaman pada umur 65 hari setelah tanam pada penelitian ini, secara umum antar tidak terjadi interaksi antara faktor varietas yang digunakan dan faktor cekaman kekeringan yang diberikan pada semua pengamatan. Dari pengamatan, varietas menunjukkan berbeda nyata pada pengamatan tinggi tanaman, jumlah cabang, jumlah daun trifoliat, bobot kering tajuk, serta panjang dan bobot kering akar. Adapun faktor cekaman kekeringan, perbedaan hanya terjadi pada bobot kering tajuk dan bobot kering akar.
Tidak terjadinya interaksi antara varietas dan faktor cekaman kekeringan pada penelitian ini, diduga karena cekaman diberikan pada saat tanaman sudah mulai memasuki pertumbuhan generatif (7 minggu setelah tanam), sehingga pertumbuhan vegetatif tanaman hampir mencapai maksimal, sehingga perlakuan cekaman yang diberikan tidak berpengaruh nyata terhadap peubah pengamatan.
Cekaman kekeringan dapat mengurangi pertumbuhan tanaman. Besarnya penurunan pertumbuhan sangat tergantung pada besarnya cekaman yang diterima
20
dan waktu terjadinya cekaman pada tanaman. Tanaman memiliki berbagai mekanisme untuk menghindari diri dari kondisi cekaman yang dihadapi, seperti mengurangi kehilangan air melalui transpirasi dengan penutupan stomata, dan memperbesar penyerapan air dengan meningkatkan pertumbuhan akar. Cekaman kekeringan menyebabkan penutupan stomata yang menyebabkan terhambatnya proses fotosintesis dan secara langsung akan berpengaruh terhadap pertumbuhan dan produksi tanaman. (Taiz dan Zeiger 2002).
Sebagian penelitian telah membuktikan bahwa cekaman kekeringan berpengaruh negatif terhadap pertumbuhan, diantaranya penelitian Blanco et al. (2002) pada tanaman cistus albidus yang membuktikan bahwa perlakuan cekaman kekeringan menyebabkan pengurangan tinggi tanaman, jumlah daun dan bobot kering akar.
Tinggi tanaman (cm)
Secara umum perlakuan cekaman kekeringan akan menyebabkan terhambatnya pertumbuhan tanaman. Hambatan pertumbuhan disebabkan oleh berkurangnya tekanan turgor sel akibat menurunnya potensial air sehingga proses pembesaran dan pemanjangan sel akan terhambat (Levitt 1980).
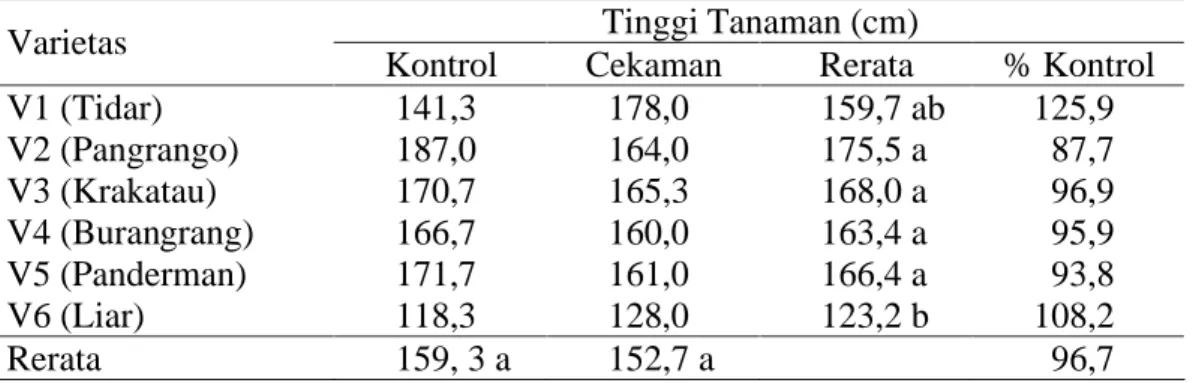
Sidik ragam terhadap pengamatan tinggi tanaman menunjukkan hasil yang tidak berbeda nyata pada perlakuan kekeringan dan interaksi antara faktor perlakuan cekaman kekeringan dan varietas yang diamati. Sidik ragam hanya berbeda nyata pada faktor varietas yang digunakan (tabel sidik ragam pada Lampiran 3).
Untuk perlakuan varietas yang digunakan, diketahui bahwa varietas yang memiliki pertumbuhan tertinggi ditunjukkan oleh varietas V2 dengan 175,5 cm, tidak berbeda nyata dengan seluruh varietas budidaya lainnya seperti varietas V1 yang memiliki tinggi 159,7 cm. Perbedaan tinggi tanaman hanya terjadi pada kedelai liar yang memiliki tinggi tanaman 123,2 cm, seperti terlihat pada Tabel 1.
21
Tabel 1. Data pertumbuhan tinggi tanaman (cm) umur 65 hari pada perlakuan kontrol dan cekaman kekeringan
Tinggi Tanaman (cm) Varietas
Kontrol Cekaman Rerata % Kontrol
V1 (Tidar) 141,3 178,0 159,7 ab 125,9 V2 (Pangrango) 187,0 164,0 175,5 a 87,7 V3 (Krakatau) 170,7 165,3 168,0 a 96,9 V4 (Burangrang) 166,7 160,0 163,4 a 95,9 V5 (Panderman) 171,7 161,0 166,4 a 93,8 V6 (Liar) 118,3 128,0 123,2 b 108,2 Rerata 159, 3 a 152,7 a 96,7
Keterangan : angka-angka yang diikuti huruf yang sama pada kolom dan baris yang sama tidak berbeda nyata menurut uji DMRT pada taraf a=5%.
Perbedaaan tinggi tanaman antar tiap varietas yang digunakan diduga lebih banyak disebabkan oleh faktor genetis tanaman, dimana varietas kedelai budidaya memiliki pertumbuhan yang lebih besar dibandingkan kedelai liar. Adapun perlakuan cekaman kekeringan yang diberikan tidak berbeda nyata dari tanaman kontrol diduga disebabkan perlakuan yang diberikan selama 14 hari dilakukan ketika tanaman sudah memasuki fase pertumbuhan generatif sehingga sel meristem sudah tidak aktif membelah dan energi tanaman lebih banyak terpakai untuk pembentukan bunga dan polong yang telah menjadi sink dari fotosintat.
Perlakuan cekaman kekeringan diduga akan sangat berpengaruh bila tanaman masih dalam tahap pertumbuhan vegetatif, seperti hasil penelitian Keles dan Oncel (2002) pada tanaman gandum dimana cekaman kekeringan dan salinitas akan sangat berpengaruh terhadap terhambatnya pemanjangan pucuk (tinggi tanaman).
Jumlah cabang tanaman
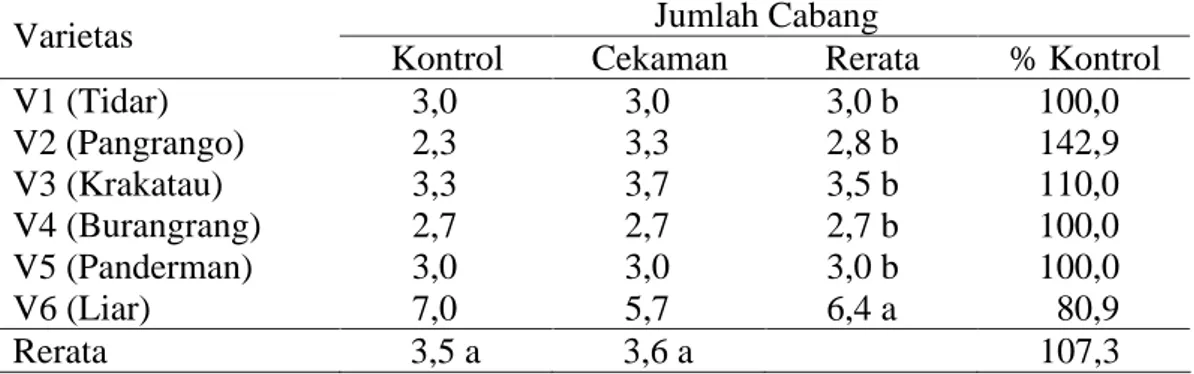
Sidik ragam terhadap pengamatan jumlah cabang menunjukkan hasil yang tidak berbeda nyata pada perlakuan kekeringan dan interaksi antara faktor perlakuan cekaman kekeringan dan varietas yang diamati. Sidik ragam hanya berbeda nyata pada faktor varietas yang digunakan (tabel sidik ragam pada Lampiran 3).
Pengamatan pertumbuhan vegetatif untuk jumlah cabang sebagaimana terlihat pada Tabel 2, diketahui bahwa rata-rata jumlah cabang terbanyak dimiliki oleh kedelai V6 sebanyak 6,4 cabang, sedangkan varietas yang memiliki jumlah
22
cabang terkecil adalah V2 dengan 2,8 cabang dan tidak berbeda nyata dibandingkan dengan semua varietas budidaya.
Tabel 2. Data pertumbuhan jumlah cabang tanaman umur 65 hari pada perlakuan kontrol dan cekaman kekeringan
Jumlah Cabang Varietas
Kontrol Cekaman Rerata % Kontrol
V1 (Tidar) 3,0 3,0 3,0 b 100,0 V2 (Pangrango) 2,3 3,3 2,8 b 142,9 V3 (Krakatau) 3,3 3,7 3,5 b 110,0 V4 (Burangrang) 2,7 2,7 2,7 b 100,0 V5 (Panderman) 3,0 3,0 3,0 b 100,0 V6 (Liar) 7,0 5,7 6,4 a 80,9 Rerata 3,5 a 3,6 a 107,3
Keterangan : angka-angka yang diikuti huruf yang sama pada kolom dan baris yang sama tidak berbeda nyata menurut uji DMRT pada taraf a=5%.
Perbedaan banyaknya jumlah cabang antara kedelai liar dan kedelai budidaya diduga lebih banyak disebabkan oleh faktor genetis tanaman, dimana dari hasil pengamatan jumlah cabang varietas budidaya hanya berjumlah 2 - 4 cabang, berbeda nyata dengan kedelai liar. Adapun perlakuan cekaman kekeringan yang diberikan tidak mempengaruhi pembentukan cabang tanaman.
Menurut Adisarwanto (2005), cabang tanaman kedelai akan muncul di batang tanaman dan jumlah cabang tergantung dari varietas dan kondisi tanah, tetapi ada beberapa varietas kedelai yang tidak bercabang. Jumlah cabang tidak mempunyai hubungan yang signifikan dengan jumlah biji yang diproduksi, artinya walaupun tanaman memiliki jumlah cabang yang banyak belum tentu menghasilkan produksi yang tinggi.
Jumlah daun trifoliat
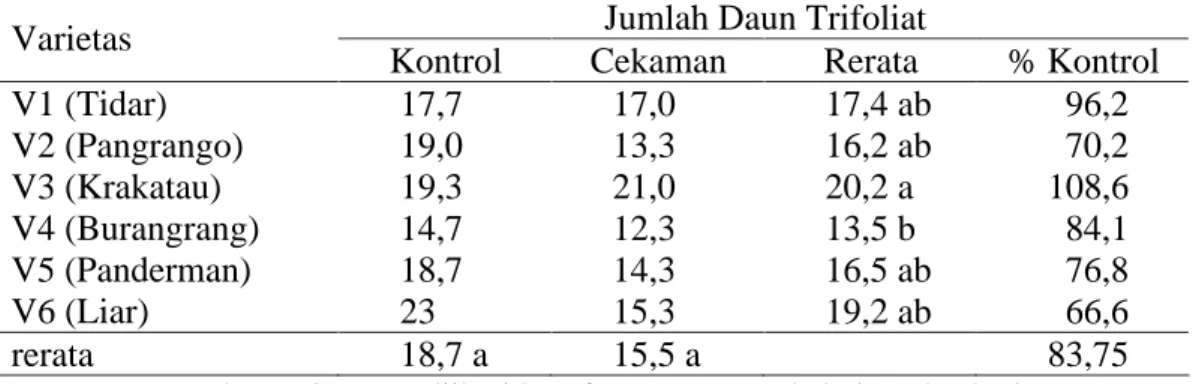
Untuk pengamatan terhadap jumlah daun trifoliat, sidik ragamnya menunjukkan hasil yang tidak berbeda nyata pada perlakuan kekeringan dan interaksi antara faktor perlakuan cekaman kekeringan dan varietas yang diamati. Sidik ragam hanya berbeda nyata pada faktor varietas yang digunakan (tabel sidik ragam pada Lampiran 3).
Dari hasil pengamatan jumlah daun trifoliat, diketahui jumlah rerata daun terbanyak ditunjukkan oleh V3 sebagai varietas moderat cekaman kekeringan sebanyak 20,2 helai pada kondisi cekaman, jumlah ini berbeda nyata dengan varietas (V4) yang memiliki rata-rata 13,5 helai, seperti terlihat pada Tabel 3.
23
Tabel 3. Data pertumbuhan jumlah daun trifoliat tanaman umur 65 hari pada perlakuan kontrol dan cekaman kekeringan.
Jumlah Daun Trifoliat Varietas
Kontrol Cekaman Rerata % Kontrol
V1 (Tidar) 17,7 17,0 17,4 ab 96,2 V2 (Pangrango) 19,0 13,3 16,2 ab 70,2 V3 (Krakatau) 19,3 21,0 20,2 a 108,6 V4 (Burangrang) 14,7 12,3 13,5 b 84,1 V5 (Panderman) 18,7 14,3 16,5 ab 76,8 V6 (Liar) 23 15,3 19,2 ab 66,6 rerata 18,7 a 15,5 a 83,75
Keterangan : angka-angka yang diikuti huruf yang sama pada kolom dan baris yang sama tidak berbeda nyata menurut uji DMRT pada taraf a=5%.
Pertumbuhan sel tanaman akan menurun sebagai respon tanaman menghadapi cekaman kekeringan karena perkembangan sel berhubungan dengan ketersediaan air di dalam sel dan akan berhubungan dengan tekanan turgor sel. Sel turgor dapat menurun akibat kekeringan sehingga menghambat pembentukan dan pembesaran daun. Sel daun dapat berkembang kembali dengan menggunakan 50 – 60% air dari total air batang (Levitt 1980).
Kedelai merupakan tanaman indeterminan, sehingga ketika terjadi cekaman kekeringan akan menyebabkan tanaman mengurangi jumlah daun, sehingga secara tidak langsung menyebabkan terjadinya penurunan jumlah dan pertumbuhan rata-rata tanaman. Berkurangnya pembentukan daun tanaman pada kondisi kekeringan merupakan mekanisme tanaman untuk menghindari kehilangan air yang akan terjadi bila tanaman memiliki daun dalam jumlah yang besar (Taiz dan Zeiger 2002). Perbandingan antara kedelai budidaya dan kedelai liar yang terkena cekaman kekeringan dapat dilihat pada Gambar 3.
24
Gambar 3. Perbedaan respon tanaman kedelai varietas budidaya (Glycine max (L.) merr ) dan kedelai liar (Glycine tomentella) menghadapi perlakuan cekaman kekeringan pada hari ke 10.
Perbedaan jumlah daun antar varietas ini menunjukkan bahwa sebagian besar varietas budidaya adalah tipe “drought avoidance” sebagaimana ciri yang dikemukakan oleh Levitt (1980) bahwa tanaman golongan “avoidance water
spender” akan mengurangi berat dan jumlah daun yang dimilikinya untuk
mengurangi besarnya kehilangan air dari tanaman. Bobot kering tajuk tanaman (g)
Secara umum cekaman kekeringan akan menghambat pertumbuhan tanaman sehingga akan mengurangi jumlah biomas yang dihasilkan. Hal ini disebabkan cekaman kekeringan akan menghambat pertumbuhan dan perkembangan tanaman akibat terhambatnya proses pembelahan sel
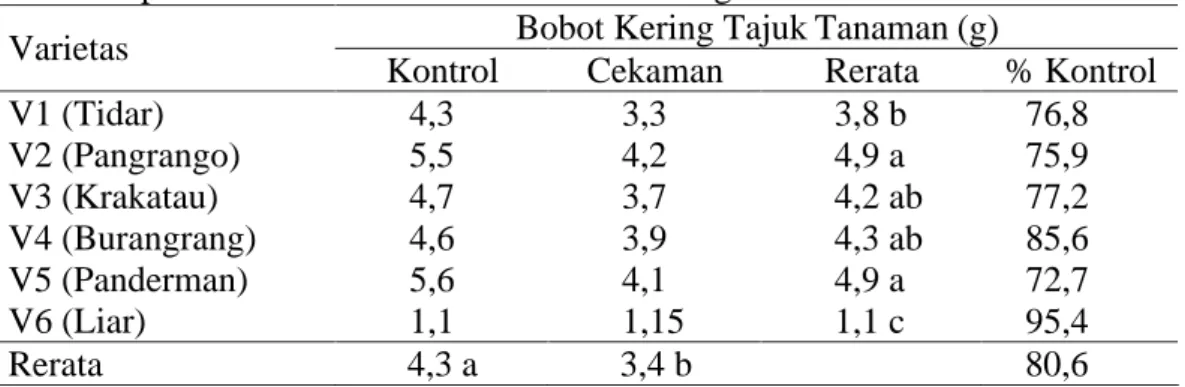
Untuk pengamatan terhadap bobot kering tajuk, sidik ragamnya menunjukkan hasil yang berbeda nyata pada perlakuan kekeringan dan varietas yang diamati, akan tetapi tidak terjadi interaksi antara faktor perlakuan cekaman kekeringan dan varietas (tabel sidik ragam pada Lampiran 3). Untuk hasil pengamatan terhadap bobot kering tajuk tanaman dapat dilihat pada Tabel 4.
25
Tabel 4. Data pertumbuhan bobot kering tajuk tanaman (g) umur 65 hari pada perlakuan kontrol dan cekaman kekeringan
Bobot Kering Tajuk Tanaman (g) Varietas
Kontrol Cekaman Rerata % Kontrol
V1 (Tidar) 4,3 3,3 3,8 b 76,8 V2 (Pangrango) 5,5 4,2 4,9 a 75,9 V3 (Krakatau) 4,7 3,7 4,2 ab 77,2 V4 (Burangrang) 4,6 3,9 4,3 ab 85,6 V5 (Panderman) 5,6 4,1 4,9 a 72,7 V6 (Liar) 1,1 1,15 1,1 c 95,4 Rerata 4,3 a 3,4 b 80,6
Keterangan : angka-angka yang diikuti huruf yang sama pada kolom dan baris yang sama tidak berbeda nyata menurut uji DMRT pada taraf a=5%.
Data pada Tabel 4 menunjukkan bahwa tanaman kedelai yang memiliki nilai bobot kering tajuk terbesar dimiliki oleh varietas V2 dan V5 yaitu masing-masing seberat 4,9 g, tidak berbeda nyata dengan varietas budidaya yang ada, kecuali varietas V1 yang memiliki bobot kering tajuk sebesar 3,8 g. Adapun tanaman yang memiliki bobot kering tajuk terendah adalah kedelai liar (V6) sebesar 1,1 g.
Secara umum perlakuan cekaman menyebabkan terjadinya penurunan bobot tajuk tanaman yang mengalami cekaman kekeringan dibandingkan tanaman kontrol sebesar 19,4%. Penurunan tertinggi terjadi pada varietas V5 sebesar 27,3%, sedangkan varietas yang memiliki penurunan terendah ditunjukkan oleh kedelai liar (V6) sebesar 4,6%.
Terjadinya perbedaan nilai bobot kering tajuk selain disebabkan oleh terhambatnya laju pertumbuhan dan perkembangan tanaman, juga disebabkan faktor genetis tanaman. Kedelai liar yang memiliki nilai bobot kering tajuk terkecil, disebabkan karena secara genetis kedelai liar memiliki morfologi yang lebih kecil dibandingkan varietas budidaya. Akan tetapi kedelai liar yang termasuk toleran cekaman kekeringan, dapat mempertahankan pertumbuhan dan perkembangannya, sehingga tidak terjadi penurunan yang besar ketika mengalami cekaman kekeringan.
Pembesaran sel sangat sensitif terhadap cekaman kekeringan, hal ini tidak terlepas dari terhambatnya proses biokimia dan molekuler yang terjadi akibat berkurangnya kandungan air didalam tanaman. Cekaman kekeringan menyebabkan terjadinya penurunan nilai pertumbuhan rata-rata tanaman.
26
Terhambatnya pertumbuhan antara lain disebabkan terhambatnya berbagai proses penting seperti biosintesis dinding sel dan membran, pembelahan sel dan sintesis protein. Setiap gram pembentukan bahan organik penyusun tanaman rata-rata membutuhkan 500 g air yang diabsorbsi oleh akar ditransportasikan ke seluruh bahagian tanaman (Taiz dan Zeiger 2002).
Terjadinya penurunan bobot kering tajuk pada tanaman juga disebabkan oleh berkurangnya aktifitas fotosintesis akibat menutupnya stomata. Fotosintat hasil fotosintesis merupakan sumber energi untuk proses pertumbuhan dan pembesaran sel tanaman. Transpor fotosintat melalui floem juga membutuhkan air, sehingga cekaman kekeringan menyebabkan pasokan fotosintat didalam tanaman menjadi terhambat (Taiz dan Zeiger 2002)
Penutupan stomata menyebabkan berkurangnya aktifitas fotosintesis tidak lepas dari aktifitas kerja Asam Absisik (ABA) yang merupakan hormon tanaman yang sangat berperan ketika tanaman mengalami cekaman kekeringan dengan menginduksi penutupan stomata (Sharp 2002).
Rendahnya potensial air tanah ketika terjadinya cekaman kekeringan menyebabkan berkurangnya serapan hara akibat menurunnya kandungan hara yang tersedia di dalam tanah yang dapat diserap akar tanaman. Unsur hara dan mineral tanah merupakan bahan penyusun utama dari bahan organik tanaman, sehingga berkurangnya suplai hara yang larut bersama air yang diserap akar secara langsung akan mengurangi bobot kering tanaman.
Levitt (1980) menyatakan bahwa air berfungsi sebagai pelarut gas-gas, garam-garam dan zat-zat terlarut yang bergerak ke dalam sel tanaman. Ketersediaan air tanah -0,1 sampai -10 bar esensial bagi setiap proses peningkatan ketersediaan hara.
Hara yang paling nyata dipengaruhi oleh kandungan air tanah adalah nitrat, sulfat, yang merupakan dekomposisi bahan organik tanaman. Kandungan air tanah mempengaruhi transpor hara ke permukaan akar dengan cara mempengaruhi laju difusi dan aliran massa air ke akar. Berbagai percobaan dengan unsur hara makro (N, P, dan K) serta sekunder (Ca, Mg, dan S) memperlihatkan bahwa penurunan suplai air mengakibatkan peningkatan yang
27
nyata pada konsentrasi N, penurunan yang nyata pada konsentrasi K dan pengaruh yang bervariasi untuk konsentrasi P, Ca, dan Mg dalam tanaman.
Pertumbuhan dan respon akar
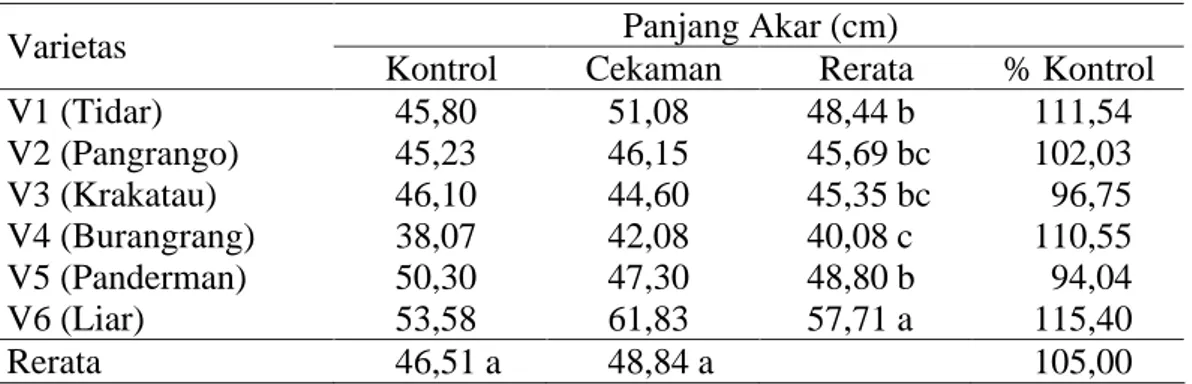
Panjang akar menunjukkan hasil yang berbeda nyata, dimana tiap varietas kedelai memiliki respon panjang akar yang beragam ketika menghadapi perlakuan cekaman kekeringan, sebagaimana terlihat pada Tabel 5. Pengamatan terhadap panjang akar, sidik ragamnya menunjukkan hasil yang tidak berbeda nyata pada perlakuan kekeringan dan interaksi antara faktor perlakuan cekaman kekeringan dan varietas yang diamati. Sidik ragam hanya berbeda nyata pada faktor varietas yang digunakan (tabel sidik ragam pada Lampiran 3).
Pada pengamatan ini, perlakuan cekaman kekeringan tidak menyebabkan perbedaan yang signifikan antara tanaman perlakuan dan tanaman kontrol, walau secara umum perlakuan cekaman kekeringan cendrung menyebabkan terjadinya peningkatan panjang akar.
Tabel 5. Panjang akar pada perlakuan kontrol dan cekaman kekeringan Panjang Akar (cm)
Varietas
Kontrol Cekaman Rerata % Kontrol
V1 (Tidar) 45,80 51,08 48,44 b 111,54 V2 (Pangrango) 45,23 46,15 45,69 bc 102,03 V3 (Krakatau) 46,10 44,60 45,35 bc 96,75 V4 (Burangrang) 38,07 42,08 40,08 c 110,55 V5 (Panderman) 50,30 47,30 48,80 b 94,04 V6 (Liar) 53,58 61,83 57,71 a 115,40 Rerata 46,51 a 48,84 a 105,00
Keterangan : angka-angka yang diikuti huruf yang sama pada kolom dan baris yang sama tidak berbeda nyata menurut uji DMRT pada taraf a=5%.
Dari Tabel 5 terlihat bahwa pada kondisi cekaman kekeringan varietas V6 sebagai varietas toleran cekaman kekeringan memiliki sistem perakaran yang terpanjang (57,71 cm). Nilai panjang akar kedelai liar berbeda nyata dengan varietas budidaya yang ada. Pada varietas budidaya, akar terpanjang ditunjukkan oleh varietas V5 sebesar 48,80 cm walau tidak berbeda nyata dengan varietas lainnya seperti V1, V2 dan V3 masing-masing memiliki panjang 48,44 cm, 45,69 cm dan 45,35 cm, akan tetapi berbeda nyata dengan varietas V4 yang memiliki nilai panjang akar terpendek yaitu 40,08 cm.
28
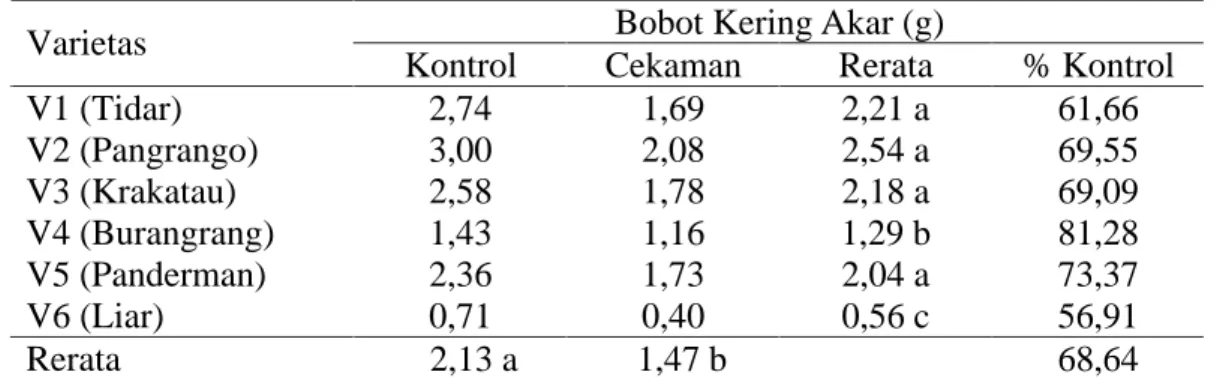
Pemanjangan akar pada tanaman yang mengalami cekaman kekeringan akan mempengaruhi bobot kering akar yang dimiliki tanaman. Pengamatan terhadap bobot kering akar, sidik ragamnya menunjukkan hasil yang berbeda nyata pada perlakuan kekeringan dan varietas yang diamati. Sidik ragam menunjukkan tidak berbeda nyata pada interaksi antara faktor perlakuan cekaman kekeringan dan faktor varietas yang digunakan (tabel sidik ragam pada Lampiran 3).
Dari data bobot kering akar pada Tabel 6 diketahui bahwa bobot kering akar terbesar ditunjukkan oleh varietas V2 sebesar 2,54 g, walau tidak berbeda nyata dengan pengamatan pada varietas V1, V3, dan V5 yang masing masing memiliki nilai 2,21 g, 2,18 g, dan 2,04 g, akan tetapi berbeda nyata pada varietas V4 yang memiliki bobot kering akar 1,29 g. Adapun bobot kering akar terkecil dimiliki oleh kedelai liar sebesar 0,56g. .
Tabel 6. Bobot kering akar pada perlakuan kontrol dan cekaman kekeringan Bobot Kering Akar (g)
Varietas
Kontrol Cekaman Rerata % Kontrol
V1 (Tidar) 2,74 1,69 2,21 a 61,66 V2 (Pangrango) 3,00 2,08 2,54 a 69,55 V3 (Krakatau) 2,58 1,78 2,18 a 69,09 V4 (Burangrang) 1,43 1,16 1,29 b 81,28 V5 (Panderman) 2,36 1,73 2,04 a 73,37 V6 (Liar) 0,71 0,40 0,56 c 56,91 Rerata 2,13 a 1,47 b 68,64
Keterangan : angka-angka yang diikuti huruf yang sama pada kolom dan baris yang sama tidak berbeda nyata menurut uji DMRT pada taraf a=5%.
Perbedaan panjang dan bobot akar ini menunjukkan bahwa tiap varietas memiliki respon akar yang berbeda menghadapi cekaman kekeringan dan dapat menggambarkan sistem perakaran yang dimiliki tanaman. Kedelai liar walau memiliki panjang akar terpanjang, akan tetapi ia justru memiliki bobot akar terkecil. Hal ini menunjukkan bahwa kedelai liar memiliki sistem perakaran yang sangat kecil dan lebih banyak memiliki rambut akar yang halus. Hal ini berbeda dengan varietas lainnya seperti V2 yang memiliki panjang akar yang relatif pendek tetapi memiliki bobot akar terbesar, yang menunjukkan bahwa varietas ini memiliki sistem perakaran yang banyak dan besar.
Perbandingan pertumbuhan dan respon akar tanaman yang diberi perlakuan cekaman kekeringan dan kontrol dapat dilihat pada Gambar 4.
29
Gambar 4. Perbedaan perakaran tanaman kontrol dan perlakuan cekaman kekeringan pada varietas budidaya V1
Kemampuan tanaman untuk merespon kondisi cekaman kekeringan diantaranya dengan melakukan perpanjangan akar sangat diperlukan sebagai upaya untuk memperbesar penyerapan air dari partikel tanah sampai lapisan yang paling bawah yang masih menyimpan air.
Pemanjangan akar pada kondisi cekaman kekeringan dimungkinkan karena tanaman memiliki mekanisme pengaturan perbandingan pertumbuhan tajuk akar (root and shoot ratio). Pada kondisi cekaman kekeringan tanaman akan menahan laju pertumbuhan tajuk dengan mensintesis hormon retardan pertumbuhan yang menghambat pertumbuhan tajuk, sehingga memperbesar laju pertumbuhan akar. Mekanisme ini dilakukan tanaman untuk mencegah besarnya kehilangan air dari tanaman, sebab untuk perpanjangan akar diperlukan lebih sedikit air dibandingkan pemanjangan pucuk yang akan memperbesar proses respirasi dengan pembentukan daun. Proses pemanjang akar juga akan dapat menjangkau volume tanah yang lebih besar sehingga lebih banyak menyerap air (Levitt 1980).
30
Terhambatnya pembesaran daun akan mengurangi konsumsi karbon dan energi tanaman, sehingga sebagian besar porsi asimilat tanaman akan didistribusikan menuju sistem perakaran untuk menunjang pertumbuhan akar (Taiz dan Zeiger 2002).
Perbedaan panjang akar pada penelitian ini diduga juga dipengaruhi oleh suhu rumah kaca yang mencapai 30 – 40 oC, seperti hasil penelitian Keles dan Oncel (2002) bahwa cekaman kekeringan pada suhu 30–40oC akan menyebabkan pemanjangan akar lebih besar dibandingkan pada temperatur normal.
Dari struktur akar pada Gambar 4 terlihat bahwa akar tanaman kontrol terlihat lebih banyak dan besar, sedangkan pada tanaman yang diberi perlakuan akar cenderung lebih kecil, tetapi memiliki ukuran yang lebih panjang. Hal ini karena pemanjangan akar juga tergantung kepada aktifitas fotosintesis, dimana pengurangan laju fotosintesis menyebabkan suplai fotosintat ke akar menjadi berkurang, sementara akar dituntut untuk menyerap air sebanyak-banyaknya, sehingga menyebabkan tanaman memberhentikan pertumbuhan tajuk dan untuk mengalirkan fotosintat menuju akar membentuk akar yang lebih panjang walau memiliki diameter yang lebih kecil.
Untuk membentuk jaringan perakaran, tanaman harus menyediakan karbohidrat dalam jumlah yang besar dialirkan melalui aliran floem sebagai sumber energi untuk perkembangan zona apikal sebagai pendukung pembesaran dan perpanjangan sel (Taiz dan Zeiger 2002).
Akar pada tanaman terkena cekaman kekeringan memiliki ukuran yang lebih panjang tetapi memiliki bobot kering yang lebih kecil akibat terbatasnya tekanan turgor, bila tanaman kembali dilakukan recovery maka nilai bobot kering akar cenderung sama dengan tanaman kontrol (Blanco et al. 2002).
31
Produksi Tanaman Jumlah dan persentasi polong isi
Untuk data jumlah polong, sidik ragamnya menunjukkan hasil yang berbeda nyata pada perlakuan kekeringan dan varietas yang diamati. Sidik ragam menunjukkan tidak berbeda nyata pada interaksi antara faktor perlakuan cekaman kekeringan dan faktor varietas yang digunakan (tabel sidik ragam pada Lampiran 3). Data jumlah polong tanaman dapat dilihat pada Tabel 7.
Tabel 7. Jumlah polong pada perlakuan kontrol dan cekaman kekeringan Jumlah Polong
Varietas
Kontrol Cekaman Rerata % Kontrol
V1 (Tidar) 24,0 17,3 20,65 a 72,2 V2 (Pangrango) 23,0 15,3 19,15 ab 66,6 V3 (Krakatau) 19,3 14,0 16,65 abc 72,4 V4 (Burangrang) 15,3 11,3 13,30 bc 73,9 V5 (Panderman) 13,3 8, 7 11,00 c 65,0 V6 (Liar) 12,3 10,3 11,30 c 83, 8 rerata 17,9 a 12, 9 b 72,4
Keterangan : angka-angka yang diikuti huruf yang sama pada kolom dan baris yang sama tidak berbeda nyata menurut uji DMRT pada taraf a=5%.
Hasil panen tanaman menunjukkan bahwa varietas V1 menghasilkan jumlah polong terbanyak pada kondisi cekaman kekeringan (20,65) tidak berbeda dengan jumlah polong dari beberapa varietas toleran dan moderat cekaman kekeringan lainnya yaitu V2 (19,15), dan V3 (16,65). Jumlah polong terendah dihasilkan oleh varietas V5 sebanyak 11 polong, tidak berbeda dengan varietas V4 (13,30) dan kedelai liar (11,30).
Untuk data persentase polong isi, sidik ragam tidak menunjukkan berbeda nyata pada semua faktor tunggal dan interaksi yang diamati (tabel sidik ragam pada Lampiran 3). Walaupun tidak berbeda nyata, secara umum terjadi penurunan persentase polong isi pada tanaman yang diberi perlakuan cekaman kekeringan. Penurunan jumlah polong terbesar ditunjukkan oleh varietas V5 dan kedelai liar (V6) sebesar 75% dan 74,2%, sedangkan pada varietas V1, tanaman masih mengalami peningkatan jumlah polong sebesar 101,5% ketika mengalami cekaman kekeringan.
32
Tabel 8. Persentase polong isi pada perlakuan kontrol dan cekaman kekeringan Persentase Polong Isi
Varietas
Kontrol Cekaman Rerata % Kontrol
V1 (Tidar) 95,8 97,2 96,50 a 101,5 V2 (Pangrango) 92,7 87,0 89,85 a 93,8 V3 (Krakatau) 93,1 92,9 93,00 a 99,8 V4 (Burangrang) 93,5 92,5 93,00 a 99,0 V5 (Panderman) 97,5 73,1 85,30 a 75,0 V6 (Liar) 100,0 74,2 87,10 a 74,2 Rerata 95,5 a 86,2 a 90,5
Keterangan : angka-angka yang diikuti huruf yang sama pada kolom dan baris yang sama tidak berbeda nyata menurut uji DMRT pada taraf a=5%.
Tingginya jumlah polong yang dihasilkan varietas V1 pada kondisi cekaman menunjukkan bahwa ia temasuk varietas yang toleran cekaman kekeringan. Hal ini ditunjukkan dari produksi hasilnya yang masih mendekati potensi hasil yang dimiliki, walau tanaman telah mengalami cekaman kekeringan. Hal ini dimungkinkan karena varietas toleran kedelai memiliki berbagai mekanisme untuk dapat mempertahankan proses fotosintesis tetap tinggi pada kondisi cekaman kekeringan.
Polong yang dihasilkan tanaman merupakan sink hasil dari jumlah fotosintat yang dihasilkan tanaman selama proses pembentukan polong. Banyaknya jumlah polong yang dihasilkan tanaman merupakan tanda bahwa jumlah fotosintat yang dihasilkan tanaman sebanding dengan jumlah polong yang dihasilkan. Bila jumlah fotosintat yang tersedia tidak dapat memenuhi perkembangan polong, maka tanaman akan menggugurkan beberapa polongnya (palmer et al. 1995)
Jumlah biji dan bobot total biji
Sidik ragam pada jumlah biji menunjukkan hasil yang tidak berbeda nyata pada perlakuan kekeringan dan interaksi yang diamati, sidik ragam hanya menunjukkan berbeda nyata pada faktor varietas yang digunakan (tabel sidik ragam pada Lampiran 3).
Data rata-rata jumlah biji yang dihasilkan tanaman, diketahui bahwa jumlah biji terbanyak saat tanaman mendapat perlakuan cekaman kekeringan dihasilkan oleh kedelai liar (V6) sebanyak 73,8 butir. Hasil ini tidak berbeda
33
dengan jumlah biji yang dihasilkan oleh V1 sebanyak 70,2 butir. Sebaliknya, jumlah biji terendah dihasilkan oleh varietas V4 dan V5 yang hanya menghasilkan sebanyak 31,2 dan 34,5 butir.
Tingginya jumlah biji yang dihasilkan, menunjukkan bahwa kedelai liar (V6) dan V1 yang merupakan varietas toleran cekaman kekeringan dapat mempertahankan proses fisiologis dan metabolisme yang terjadi didalam tubuhnya lebih baik dibandingkan varietas peka ketika terkena cekaman.
Tabel 9 Jumlah Biji pada perlakuan kontrol dan cekaman kekeringan Jumlah Biji
Varietas
Kontrol Cekaman Rerata % Kontrol
V1 (Tidar) 89,00 51,33 70,20 b 57,67 V2 (Pangrango) 66,67 39,00 52,80 ab 58,50 V3 (Krakatau) 72,33 44,33 58,30 ab 61,29 V4 (Burangrang) 40,33 22,00 31,20 a 54,55 V5 (Panderman) 44,00 25,00 34,50 a 56,82 V6 (Liar) 80,00 67,50 73,80 b 84,38 Rerata 65,40 a 41,50 a 62,20
Keterangan : angka-angka yang diikuti huruf yang sama pada kolom dan baris yang sama tidak berbeda nyata menurut uji DMRT pada taraf a=5%.
Untuk data bobot total biji, sidik ragam menunjukkan hasil yang tidak berbeda nyata pada perlakuan kekeringan dan interaksi yang diamati. Sidik ragam menunjukkan tidak berbeda nyata pada faktor varietas yang digunakan (tabel sidik ragam pada Lampiran 3).
Untuk data faktor varietas yang diamati sebagaimana terlihat pada Tabel 10, diperoleh bahwa varietas V5 memiliki bobot biji terbesar yaitu 2,79 g, tidak berbeda nyata dengan seluruh varietas budidaya lainnya, dimana V1 yang memiliki bobot biji terkecil sebesar 2,08 g dan V2 yang menghasilkan 2,59 g. Adapun bobot biji terendah ditunjukkan oleh kedelai liar (V6) yaitu sebesar 0,29 g.
Perbedaan berat biji dari varietas budidaya dan kedelai liar lebih banyak disebabkan faktor genetis, dimana varietas budidaya memiliki ukuran biji yang besar dan V6 memiliki ukuran biji yang sangat kecil. Selain itu waktu terjadinya cekaman terjadi ketika tanaman masih berada pada awal fase generatif, sehingga cekaman kekeringan belum banyak berpengaruh terhadap jumlah dan berat biji yang dihasilkan.