TINJAUAN PUSTAKA
Klasifikasi, Morfologi, dan Anatomi Scylla olivacea
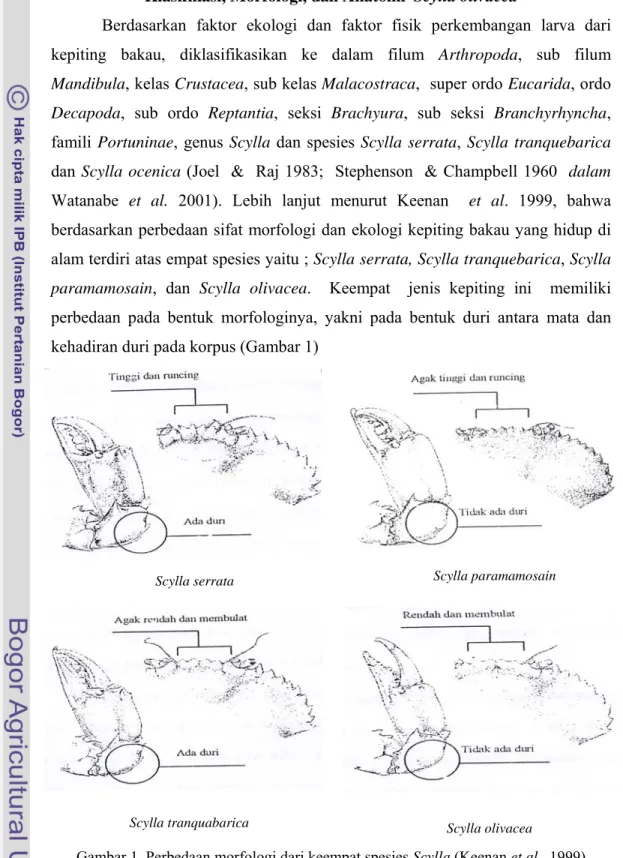
Berdasarkan faktor ekologi dan faktor fisik perkembangan larva dari kepiting bakau, diklasifikasikan ke dalam filum Arthropoda, sub filum Mandibula, kelas Crustacea, sub kelas Malacostraca, super ordo Eucarida, ordo Decapoda, sub ordo Reptantia, seksi Brachyura, sub seksi Branchyrhyncha, famili Portuninae, genus Scylla dan spesies Scylla serrata, Scylla tranquebarica dan Scylla ocenica (Joel & Raj 1983; Stephenson & Champbell 1960 dalam Watanabe et al. 2001). Lebih lanjut menurut Keenan et al. 1999, bahwa berdasarkan perbedaan sifat morfologi dan ekologi kepiting bakau yang hidup di alam terdiri atas empat spesies yaitu ; Scylla serrata, Scylla tranquebarica, Scylla paramamosain, dan Scylla olivacea. Keempat jenis kepiting ini memiliki perbedaan pada bentuk morfologinya, yakni pada bentuk duri antara mata dan kehadiran duri pada korpus (Gambar 1)
Gambar 1 Perbedaan morfologi dari keempat spesies Scylla (Keenan et al. 1999)
Scylla tranquabarica Scylla olivacea Scylla serrata Scylla paramamosain
6 Bentuk badan kepiting secara umum adalah badan yang pendek dengan abdomen yang tereduksi. Badan yang pendek diakibatkan oleh fusi antara kepala dan torak membentuk cefalotorak dan ditutupi oleh karapas. Sedangkan abdomen tereduksi menjadi tipis, rata dan terlipat dibawah cefalotorak. Oleh sebab itu, kepiting dinamakan brachyuna atau ekor pendek (Garth & Abbott 1980). Kepiting bakau ditutupi oleh karapas yaitu kulit yang terdiri atas kitin bercampur bahan kapur yang telah mengeras. Karapas berbentuk bulat pipih, dilengkapi dengan sembilan duri pada sisi kiri dan kanan. Empat duri yang lain terdapat diantara kedua matanya. Mempunyai sepasang kaki jalan yang bentuknya besar disebut capit yang berfungsi untuk memegang, tiga pasang kaki jalan dan sepasang kaki renang berbentuk bulat telur dan pipih seperti alat pendayung (Karim 1998 ).
Penyebaran dan Habitat
Scylla olivacea hidup pada berbagai habitat dan sebagian besar hidup di laut, sebagian hidup di perairan bakau atau di estuari. Pada masa juvenil sampai menjelang dewasa atau dewasa, kepiting hidup di pantai, muara-muara sungai dan hutan bakau dengan cara membuat lubang (Kasry 1986). Sebagian besar siklus hidup kepiting bakau dilalui di sekitar muara sungai dan hutan bakau (Sulaeman & Hanafi 1992). Kemampuannya yang tinggi untuk beradaptasi pada perubahan kualitas air seperti salinitas, menyebabkan kepiting sering dijumpai di sungai yang jauh dari laut dengan salinitas rendah (sekitar 5 ppt). Kepiting bakau melangsungkan perkawinan di perairan hutan bakau, selanjutnya kepiting betina beruaya ke laut untuk memijah dan kepiting jantan tetap di perairan hutan bakau atau muara sungai (Hill 1979).
Siklus Hidup
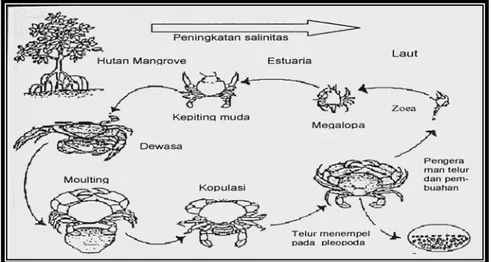
Kepiting bakau betina yang matang kelamin akan beruaya dari perairan pantai ke perairan laut untuk memijah (Brick 1974). Kemudian kepiting bakau betina dan anak-anaknya tersebut akan berusaha kembali ke perairan pantai, muara sungai atau perairan hutan bakau untuk berlindung, mencari makanan atau membesarkan anaknya. Kepiting jantan yang telah melakukan perkawinan atau
7 telah dewasa berada di perairan bakau, di tambak atau disela-sela akar bakau atau paling jauh sekitar perairan pantai yaitu pada bagian-bagian yang berlumpur yang makanannya melimpah. Mosa et al. (1985), menyatakan bahwa kepiting bakau mengalami beberapa tingkatan perkembangan mulai telur sampai dewasa.
Tingkat perkembangan tersebut dimulai dari telur, zoea, megalopa, kepiting muda dan kepiting dewasa. Telur yang telah dibuahi menetas dalam waktu 2-4 minggu menjadi larva dengan lima tingkatan zoea dan satu megalopa. Larva tersebut akan terbawa arus ke pantai. Hill (1974) menemukan bahwa larva kepiting bakau tahap zoea I tidak dapat mentolerin salinitas dibawah 14 ppt. Setelah tahap zoea, perkembangan selanjutnya adalah megalopa. Megalopa akan berenang dan masuk kembali ke muara sungai atau mangrove untuk mencari makanan dan perlindungan. Selanjutnya megalopa akan berkembang menjadi kepiting bakau muda yang bersifat bentik kemudian menjadi dewasa. Siklus hidup kepiting bakau dapat dilihat pada Gambar 2.
Gambar 2 Siklus Hidup Kepiting Bakau (Scylla spp) Modifikasi Kanna (2000) Sistem Reproduksi Kepiting Bakau Jantan
Kepiting merupakan organisme dioecious, artinya mempunyai jenis kelamin jantan dan betina pada individu yang berbeda. Berdasarkan struktur organ reproduksinya kepiting bakau tergolong kepiting yang melakukan internal fertilization (pembuahan di dalam). Sistem reproduksi pada jantan terdiri atas testis, saluran spermatozoa, dan alat ejakulasi. Sedangkan pada reproduksi kepiting betina terdiri atas ovarium, saluran telur, dan spermateka. Perbedaan
8 kepiting jantan dan betina adalah pada ruas-ruas abdomennya. Untuk jantan memiliki ruas-ruas abdomen yang lebih sempit dibandingkan dengan betina yang memiliki ruas-ruas abdomen yang lebar dan membulat (Mossa et al. 1985). Kepiting betina memiliki empat pasang pleopod yang digunakan untuk membawa telur selama musim reproduksi sedangkan jantan hanya memiliki dua pasang pleopod yang digunakan sebagai organ kopulasi.
Gambar 3 Skema dari Sistem Reproduksi Kepiting Jantan Secara Umum. Anterior Testis (AT) bergabung dengan Commissure (C),Posterior Testis (PT), Ekspansi (EX) dan Posterior Vas Deferens (PV)
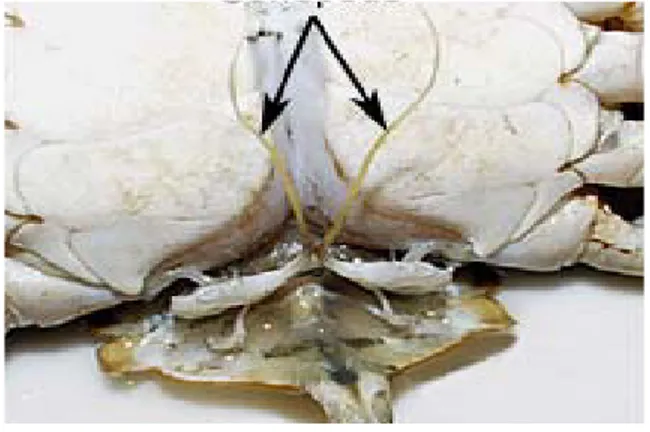
Gambar 4 Anatomi Organ Kelamin Jantan Kepiting Bakau (Scylla sp)
Sistem reproduksi kepiting jantan secara umum terdiri atas testis, saluran spermatozoa, dan alat ejakulasi (Gambar 3). Kepiting bakau jantan memiliki ruas-ruas abdomen yang lebih sempit dibandingkan dengan betina, dua pasang pleopod yang digunakan sebagai organ kopulasi (Gambar 4).
9 Testes
Susunan sistem reproduksi kepiting bakau jantan dari Scylla spp, terdapat dibawah cefalotorak. Terdiri dari sepasang testes dan vas deferen sama halnya dengan sistem reproduksi yang telah digambarkan untuk ordo Decapoda secara umum. Organ reproduksi jantan pada sub kelas Malacostraca, ditemukan dalam cefalotorak atau torak (Krol et al. 1992).
Saluran reproduksi jantan pada beberapa Decapoda, terdiri dari sepasang testis dan saluran genital (Johnson, 1980 pada C. Sapidus & Leite, 2002 pada U. Cordatus). Tiap saluran genital terdiri dari suatu kumpulan pipa, bagian vas deferen, dan suatu saluran ejakulator yang berakhir dalam seminal vesicle atau suatu ampoule terminal, bergantung pada masing-masing spesies (Krol et al. 1992).
Vas Deferen
Bagian vas deferen dari Scylla olivacea terdiri dari dua bagian utama secara umum seperti halnya pada Decapoda lainnya, namun tidak selalu sama bergantung spesies. Pada kasus lainnya, vas deferen pada L. emarginata (Hinsch & Walker 1974), C. sapidus (Johnson 1980), dan T. orientalis (Burton 1995) membagi vas deferen menjadi tiga bagian, sedangkan pada S. chacei (Hinsch & Mcknight 1988), vas deferen terbagi menjadi empat bagian dan pada D. pugilator (Manjon-Cabeza & Evenness 2000) vas deferen terbagi menjadi delapan bagian. Kriteria perbedaannya ditentukan secara mikroskopis dan makroskopis bergantung spesies.
Binford (1913), menggambarkan struktur dari vas deferen berupa garis lurus dengan epithelium kolumnar yang mensekresikan bahan kimia yang diperlukan untuk menghasilkan dinding dari spermatofor. Cronin (1947) meneliti bahwa kebanyakan bagian anterior dari vas deferen berperan dalam pengeluaran bahan-bahan kimia yang diperlukan untuk membungkus dan penyimpanan spermatofor. Hinsch dan Mcknight (1988) juga memberikan pernyataan pada pembentukan awal dari spermatofor dan produksi cairan seminal terjadi di bagian anterior dari vas deferen. Namun menurut Marie (1999) bahwa sel sperma meninggalkan testes dan memasuki vas deferen untuk berkembang sepenuhnya menjadi spermatozoa, tetapi diduga beberapa sel spermatozoa
10 menyelesaikan pematangan mereka di dalam spermateka. Sedangkan penelitian pada cairan seminal plasma dari crustacea menunjukkan bahwa seminal plasma beberapa spesies antara lain; Cirripede (Barnes 1962) dan kepiting (Jeyalectumie & Subramoniam 1987) memiliki kesamaan dengan semen dari mamalia.
Spermatofor
Kepiting atau Brachyuran menghasilkan spermatofor tunggal yang berada dalam medium cair dari seminal plasma (Jeyalectumie & Subramoniam 1997). Spermatofor crustacea memiliki bentuk yang bervariasi (Gambar 5), namun pengetahuan mengenai komposisi kimiawi dari spermatofor sangat sedikit. Beberapa peneliti memberikan pernyataan bahwa dinding dari spermatofor tersusun dari kitin (Spalding 1942; King 1948).
Uma dan Subramoniam (1979) tidak menemukan kitin pada Brachyuran, kepiting bakau Scylla serrata. Sedangkan spermatofor yang terdapat dalam individu betina pada crustacea laut, pada membran spermatozoanya terdapat kantung yang melekat, diduga mengandung gelatin kental (Berry & Heydorn 1970). Walaupun begitu ada beberapa substrat pada selaput membran dari spermatozoa dalam transport spermatozoa, namun komposisi kimianya belum diketahui (Subramoniam 1984).
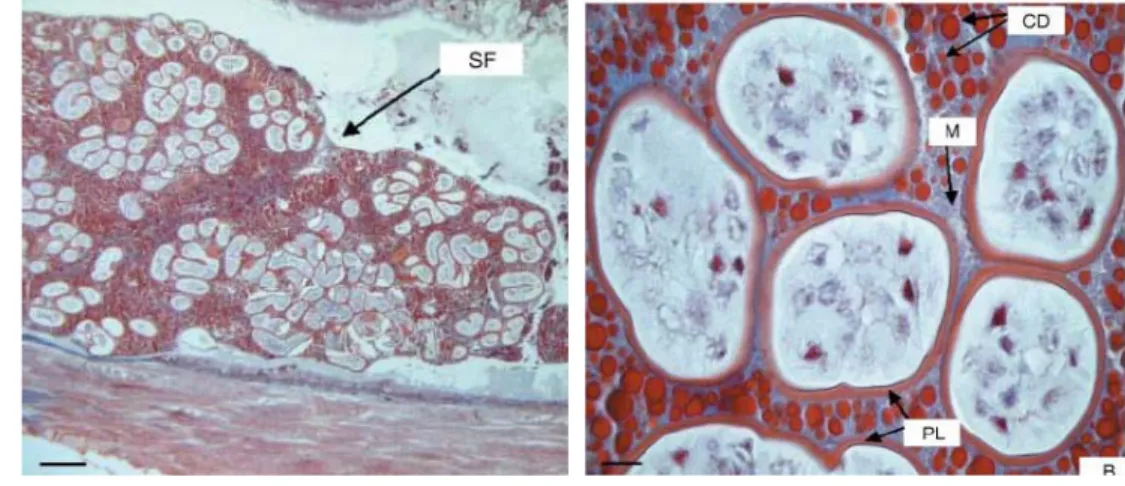
Gambar 5 Spermatofor dari C. quadricarinatus A: Gambar secara umum dari spermatofor (stained with Masson-Trichrome; bar: 286 μm). B: Gambaran detail dari spermatofor (stained with Masson-Trichrome; bar: 26 μm).CD: cytoplasmic droplets; M: matrix; PL: primary layer dari spermatofor; SF: spermatofor (Greco & Nostro 2007)
11 Spermatofor yang baik ditunjukkan dengan jumlah spermatozoa yang tinggi di dalamnya. Spermatofor memelihara kemampuan fertilisasi spermatozoa yang sudah pasti mengarah pada tingginya angka pembuahan yang tinggi, berdampak, pada kesempatan yang tinggi bagi embrio untuk bertahan hidup, menetas dan hidup sebagai larva (Akarasanon et al. 2004). Spermatozoa
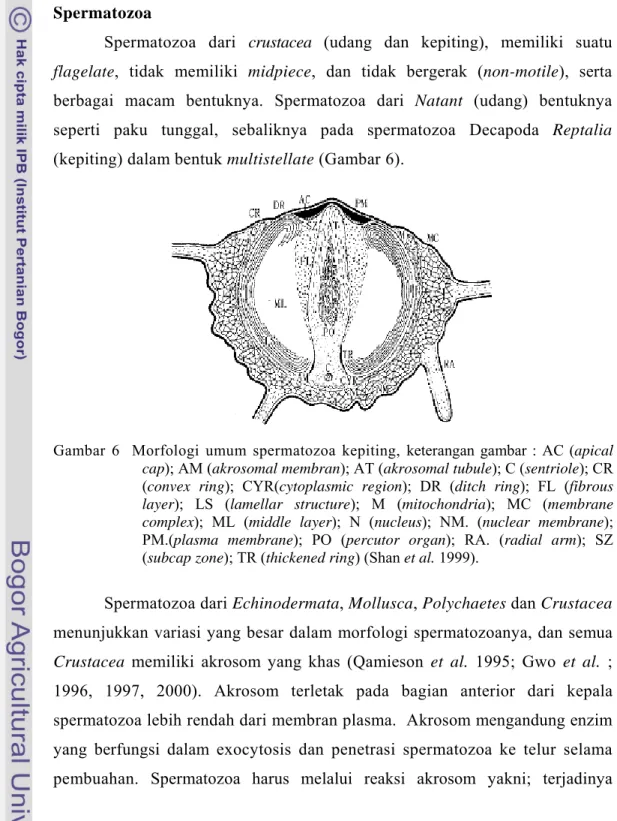
Spermatozoa dari crustacea (udang dan kepiting), memiliki suatu flagelate, tidak memiliki midpiece, dan tidak bergerak (non-motile), serta berbagai macam bentuknya. Spermatozoa dari Natant (udang) bentuknya seperti paku tunggal, sebaliknya pada spermatozoa Decapoda Reptalia (kepiting) dalam bentuk multistellate (Gambar 6).
Gambar 6 Morfologi umum spermatozoa kepiting, keterangan gambar : AC (apical cap); AM (akrosomal membran); AT (akrosomal tubule); C (sentriole); CR (convex ring); CYR(cytoplasmic region); DR (ditch ring); FL (fibrous layer); LS (lamellar structure); M (mitochondria); MC (membrane complex); ML (middle layer); N (nucleus); NM. (nuclear membrane); PM.(plasma membrane); PO (percutor organ); RA. (radial arm); SZ (subcap zone); TR (thickened ring) (Shan et al. 1999).
Spermatozoa dari Echinodermata, Mollusca, Polychaetes dan Crustacea menunjukkan variasi yang besar dalam morfologi spermatozoanya, dan semua Crustacea memiliki akrosom yang khas (Qamieson et al. 1995; Gwo et al. ; 1996, 1997, 2000). Akrosom terletak pada bagian anterior dari kepala spermatozoa lebih rendah dari membran plasma. Akrosom mengandung enzim yang berfungsi dalam exocytosis dan penetrasi spermatozoa ke telur selama pembuahan. Spermatozoa harus melalui reaksi akrosom yakni; terjadinya
12 perubahan dalam bagian akrosom sebelum menuju telur dan penetrasi secara ekstraseluler yang membutuhkan waktu (Clark et al. 1981; Jamieson et al. 1995; Gwo et al. 2000). Kelangsungan hidup spermatozoa secara umum dapat diperkirakan dari motilitas spermatozoa tetapi metode evaluasi ini tidak dapat diaplikasikan untuk spermatozoa dari crustacea, Decapoda. Spermatozoa crustacea, Decapoda tidak bergerak (Clark et al. 1981). Untuk menentukan kelangsungan hidup dari spermatozoa crustacea menggunakan metode pewarnaan eosin-nigrosin (Jeyalectumie & Subramoniam 1989) atau pewarnaan trypan blue (Bhavanishankar & Subramoniam 1997). Spermatozoa hidup tidak terwarnai, sedangkan sperma mati terwarnai dengan baik. Untuk kelangsungan hidup viabilitas dari spermatozoa Scylla serrata, bervariasi antara 95% - 67% dari suhu -196°C sampai -4°C (Jeyalectumie & Subramoniam 1997). Produksi, kualitas spermatozoa, dan pembentukan spermatofor merupakan variabel penting dalam reproduksi jantan (Leung-Trujillo & Lawrence 1987; Díaz et al. 2001). Pembentukan karbohidrat sebagai substrat utama yang digunakan dalam pemeliharaan spermatozoa sampai didalam spermateka kepiting betina matang gonad (Jeyalectumie 1989).
Fisiologi dan Pertumbuhan Gonad
Seluruh Decapoda termasuk kepiting bakau pada ukuran kepiting muda (juvenil) memiliki gonad yang belum berdiferensiasi, artinya terdapat saluran sperma (vas deferen) dan saluran telur (oviduct) bersama-sama. Perkembangan ke arah jenis kelamin jantan atau betina ditentukan oleh berkembangnya atau tidak berkembangnya kelenjar androgen. Testis dan karakteristik seksual jantan dirangsang oleh hormon androgenik yang disekresikan oleh kelenjar androgen. Sedangkan ovarium akan berkembang apabila tidak adanya kelenjar androgen (Meussy & Payen 1988; Lockwood 1996).
Mekanisme Hormonal pada Individu Jantan
Informasi mengenai peranan hormon dalam pengontrolan proses spermatogenesis pada kepiting bakau masih sedikit, tetapi untuk golongan crustacea secara umum digambarkan peranan dan pengontrolan dari kelenjar androgen terhadap proses spermatogenesis pada individu jantan (Gambar 7).
13 Gambar 7 Diagram Sistematik dari kontrol endokrin pada reproduksi jantan dalam udang. Keterangan : AH: hormone kelenjar androgenic, MF: methyl
farnesoate, MOIH: mandibular organ-inhibiting hormone (Okumura 2004) Methyl farnesoate merupakan substansi yang memiliki fungsi dalam reproduksi jantan disertai beberapa faktor yang ada dalam system saraf pusat, otak, dan ganglion torak juga berperan dalam memelihara spermatogonia dan perkembangan dari testes. Untuk reproduksi jantan pada udang, hormon kelenjar androgen berperan dalam kontrol differensiasi seksual (Okumura 2004).
Ada dua neurohormon yang berperan dalam siklus kematangan gonad, Gonad Inhibiting Hormone (GIH), yang dilepaskan dari kelenjar sinus, dan Gonad Stimulating Hormone (GSH) yang ditemukan pada otak dan ganglion torak kepiting (Sarojini et al. 1995).
Fase-Fase Reproduksi
Fase – fase reproduksi kepiting dimulai dari kopulasi (transfer spermatofor), perkembangan testes, kopulasi, pembuahan dan penetasan. Aktivitas reproduksi kepiting bakau dapat dilihat pada Tabel 1.
14 Tabel 1 Aktivitas Reproduksi Kepiting Bakau
Aktivitas Reproduksi Waktu (hari) Kopulasi
Proses Perkembangan Testes 30
Telur keluar sampai menetas 17
Kopulasi
Kopulasi hanya terjadi pada kepiting betina dan jantan yang telah dewasa kelamin. Kegiatan kopulasi dilakukan setelah betina berganti kulit sebelum pemijahan. Fermon yang dikeluarkan oleh kepiting betina pada saat ganti kulit mampu menarik kepiting jantan untuk mendekatinya (Primavera 1985; Soim 1994). Setelah karapak betina mulai mengeras, kepiting jantan membalikkan tubuh betina sehingga perut dan alat kelaminnya saling berhadapan dan selanjutnya kepiting jantan akan mentransfer spermatofor kedalam saluran reproduksi betina, yang disebut dengan spermateka dengan bantuan pleopod jantan yang berfungsi sebagai alat kopulasi (Barnes 1987).
Perkembangan Gonad
Tahap perkembangan lobul testikular pada kepiting Telmessus cheiragonus, dilaporkan oleh Nagoa dan Munehara (2003), yang dibagi menjadi 6 tahapan berdasarkan studi histologi, yaitu :
Tahap I: Tahap Proliferasi Spermatogonia
Tahap ini ditunjukkan dalam Gambar 8a, lobul mengandung spermatogonia dengan sitoplasma tipis dan nukleus berbentuk bulat panjang. Terdapat kromatin sekitar membran nuklear. Spermatogonia dalam lobul testikular memperlihatkan mitosis asynchronous. Diameter nuklei dari spermatogonia adalah 9.9 ± 1.3 x 8.0 ± 0.8mm.
Tahap II: Tahap Spermatosit Pertama
Tahap ini ditunjukkan pada Gambar 8b, sel-sel germ dalam lobul testikular selama tahap ini tidak mencapai fase zygoten dari meiosis. Sitoplasma dari spermatosit adalah kental sebagai mana yang terdapat pada spermatogonia, dan
15 nuklei menunjukkan berbagai bentuk dengan aglutinasi dari kromatin. Diameter dari nuklei sebelum hilangnya membran nuklear adalah 9.4 ± 1.1 x 8.1 ± 0.7 mm. Terjadinya meiosis secara sikronisasi pada tiap lobul individu. Tahap III: Tahap Pembelahan Reduksi (Reduction division stage)
Tahap ini ditunjukkan pada Gambar 8c dan 8d, lobul testikular selama tahap ini termasuk dalam spermatosit pertama setelah fase diploten dari meiosis atau spermatosit sekunder sebelum telofase dari bagian kedua. Pada tahap ini, spermatosit menunjukkan bentuk irregular. Nukleus dari spermatid terlihat seperti bola dengan rata-rata diameter 4.2 ± 0.6 mm.
Tahap VI : Tahap Spermatogenesis
Tahap ini ditunjukkan pada Gambar 8e, spermiogenesis terjadi di dalam lobul testikular. Penampakan kantung akrosomal sekitar nuklei saat spermiogenesis dimulai. Nukleus secara bertahap memanjang pada kantung akrosomal dan berada di sekitar area kantung saat proses spermigenesis sempurna.
Tahap V : Tahap Spermatozoa Bebas
Tahap ini ditunjukkan pada Gambar 8f, spermatozoa terpisah dari sel nutrisi di lapisan lumen dari lobul testikular. Spermatozoa memiliki suatu kantung akrosomal dan nukleus dengan tiga lengan nuklear yang memanjang membentuk jari-jari lingkaran. Sel nutrisi menyerupai sel-sel epitelia dari saluran seminiferus, spermatozoa yang diobservasi dalam lumen serta yang di dalam saluran seminiferus dikelilingi oleh sel nutrisi. Saluran seminiferus dianggap sebagai suatu lobul.
Pada brachyuran, spermatosit berkembang secara sikronisasi pada tiap lobul testikular. Lobul yang paling berkembang adalah pada tahap VI, dimana terdapat banyak spermatozoa bebas (Nagoa dan Munehara 2003).
16 Gambar 8 Gambaran Histologi dari lobul testikular pada saat tahap perkembangan. (a) Tahap I (spermatogonia proliferation stage). (b) Tahap II (first spermatocyte stage). (c) Tahap III (reduction division stage). (d) Tahap IV (spermatid stage). (e) Tahap V (spermiogenesis stage). (f) Tahap VI (free sperm stage). Bar 50 mm (Nagoa & Munehara 2003).
Proses Spermatogenesis pada Kepiting
Studi histologi pada sistem reproduksi kepiting jantan Callinectes sapidus telah dilaporkan, bahwa testis Brachyuran mengandung sel kelamin selama fase spermatogenesis yang berbeda-beda dari spermatogonia ke spermatozoa. Fluktuasi presentase kehadiran dari sel germ selama fase spermatogenesis ditemukan pada beberapa Brachyuran dan perubahan tersebut berhubungan dengan aktivitas mating (Nagoa dan Munehara 2003).
17 Spermatogenesis pada brachyuran telah dilaporkan secara mendetail, pada kepiting Cancer magister oleh Fasten (1918) dapat dilihat pada Gambar 9, 10, 11, dan 12, yaitu :
Tahap Spermatogonia
Sel spermatogonia merupakan sel yang berukuran besar, dengan garis sitoplasmik yang jelas serta nuklei menonjol. Tahap spermatogonia primer dan sekunder bisa dibedakan. Spermatogonia primer ukurannya lebih besar dibandingkan dengan spermatogonia sekunder.
Spermatogonia primer mengandung sejumlah besar kromatin dengan nukleus. Ketika sel terbagi menjadi kumpulan besar kromatin pada nukleus, dimulai proses pembelahan. Proses ini berlanjut sampai kromatin tersusun ke dalam sejumlah besar sel berbentuk bulat panjang (elliptical) yang tebal, terdistribusi di seluruh nukleus. Jumlah dari kumpulan kromatin yang diamati berbeda-beda, dengan jumlah ber kisaran 40 - 65. Binford (1913) melaporkan pada Menippe mercenaria dan Cambarus virilis, menemukan sequence yang sama pada proses tersebut. Beberapa kumpulan kromatin sebagai kromosom yang masuk kedalam piringan equator dari tahap metafase spermatogonia. Jumlah kromosom akhir ditentukan pada tahap metafase. Penampakan kumpulan kromatin menunjukkan lebih banyak kromosom tunggal.
Keadaan sekitar dinding nuklear dikelilingi oleh kumpulan kromatin yang masuk kedalam sel pada tahap metafase. Kromosom berada dalam garis equator dengan bentuk dumb-bell shaped dan munculnya kumparan serabut halus. Sentromer secara mudah dipisahkan pada kutub yang berlawanan. Penyebaran kromosom pada seluruh bidang equator. Penampakan kromosom kecil, bulat dan jumlahnya banyak tidak bisa dihitung. Tahap anafase, kemudian diikuti oleh tahap telofase, kemudian sel membagi kedalam spermatogonia sekunder.
Pembelahan spermatogonia sekunder serupa dengan spermatogonia primer, pembelahan berlanjut pada pola yang sama. Pembelahan pokok dari spermatogonia sekunder menghasilkan spermatosit primer dorman. Pada bidang spermatogonia dari tubular, beberapa sel ditemukan mengalami reduksi. Perubahan struktur nuklei ke bentuk irregular yang lebih besar dengan penonjolan (pseudopodia-like).
18 Beberapa sel yang terdapat di tubuli disebut dengan sel nutrisi "nutritive cells", telah dipelajari pada spermatozoa matang Cambarus virilis. Sel-sel nutrisi, memiliki struktur yang unik. Nuklei dari sel nutrisi mengandung masa kromatin, sementara pada sitoplasmanya memiliki banyak butiran lemak.
Umumnya penelitian awal pada Decapoda seperti : Grobben 1978; Gilson 1986; Hermann 1990, yang melaporkan bahwa spermatogonia berasal dari sel nutrisi. Sebaliknya, Keppen (1906) dan George (1992) melaporkan hal yang berlawanan bahwa sel nutrisi berasal dari perubahan spermatogonia. Hal ini sama yang didapatkan pada Cancer magister. Saat potongan sel nutrisi diuji coba, terjadi amitosis. Fakta bahwa terjadi pembelahan dalam sel-sel nutrisi. Studi nuklei yang menunjukkan bahwa sel-sel nutrisi menjadi ambceoid dalam penampakannya.
Tahap Spermatosit Primer
Tahap ini menunjukkan pembelahan spermatogonia. Setelah periode waktu pertumbuhan dan sinapsi, terjadi reduksi. Selama dua periode pertumbuhan terbentuk body spherical, dikelilingi oleh ruang kosong yang membuat spermatosit primer terlihat penampakannya dalam sitoplasma.
a. Periode Pertumbuhan
Periode ini meliputi: tahap persiapan, sinapsi dan pembentukan tetra. Awal dari fase profase, kromatin di dalam nukleus dari spermatosit yang dorman terdiri dari kumpulan kromatin yang sedikit. Beberapa spermatosit yang jumlahnya sedikit terlihat benang-benang berupa garis tipis dan adanya penampakan granular. Kromatin pada nukleus membelah ke dalam struktur yang lebih kecil dan kemudian mulai masuk ke dalam. Pada waktu yang bersamaan sel mengalami peningkatan dalam ukuran. Tahap ini merupakan awal dimulainya periode pertumbuhan dan peningkatan ukuran sel dengan jelas. Spireme terbentuk, benang-benang leptoten terpisah dan bisa dibedakan antara satu dengan yang lainnya.
Benang leptotene berpindah ke salah satu kutub dari nukleus, disebut pula dengan "synaptic pole" dalam garis lintang sejajar yang berpasangan. Wilson (1913), menggambarkan badan polar yang sama pada Pentatoma, disebut
19 dengan "chromatoid body." Pada Cambarus virilis, ditemukan sepasang badan kromatoid yang terlihat juga pada tahap yang sama dalam perkembangan spermatosit Cancer magister.
Benang-benang yang terdapat pada garis lintang menjadi lebih dekat dan berpasangan pada kutub synaptic dan sel masuk pada tahap pachyten, dimana sepasang dari benang leptoten menyatu ke dalam gemini thick. Setelah penyatuan yang konstan untuk beberapa lama, komponen dari tiap geminus mulai terurai. Tiap geminus membelah membujur, dan secara serempak pembukaan dari benang-benang yang dempet pada akhirnya, menghasilkan penampakan seperti 8, V, atau kurang lebih seperti U. Batang yang lain membagi secepatnya seluruh lengan dari tiap geminus. Dimulai dari potongan dua bidang yang membujur dan mengkerut secara jelas terlihat dalam dua lengan dari 8 yang terletak ditengah pada nukleus.
Dua bidang pembelahan yang membujur, empat benang kromatid terbentuk. Dua pasang dari beberapa benang melanjutkan kembali untuk bercabang saling berlawanan sampai akhirnya mereka berbentuk X. Sepasang benang yang berada pada posisi berlawanan dari titik penyatuan pusat. Tiap X kemudian berpindah mendekat satu sama lain, sampai pada garis lintang sejajar. Titik penyatuan pusat kemudian hilang, dan empat benang tipis tersusun dalam dua garis lintang sejajar yang dihasilkan.
Tiap benang menjadi tipis dan bertambah tipis setelah masuk ke dalam masa kromatid yang berbentuk kumparan bola dan segera tiap geminus mengalami perubahan kedalam bentuk empat spherical kromosom, menggambarkan empat serangkai. Tiap tetra mengandung empat univalen kromosom. Selanjutnya sepasang dari beberapa penyatuan univalen kromosom menghasilkan dua bivalen yang besar yang berdempetan antara satu dengan yang lainnya. Kondensasi dari bivalen berlanjut sampai mereka berubah ke dalam struktur dumb-bell shaped. Proses pertumbuhan ini telah selesai dan sel siap untuk mengalami pengurangan.
b. Pembelahan Reduksi (reduction division)
Tahap akhir periode pertumbuhan, dinding nuklear mulai membelah. Dua sentromer yang telah terbentuk dari pembelahan sentomer asli, berpindah
20 ke kutub yang berlawanan, dan terlihat kumparan serabut tipis antara sentromer dan kromosom. Kromosom segera tertarik ke bidang equator, mengalami reduksi. Bivalen yang berbentuk dumb-bell shaped terlihat tersusun. Terlihat badan kromatid pada kutub yang berlawanan. Tiap badan dikelilingi dan dikarakteristik oleh ruang kosong dan lebih mudah dikenali.
Periode metafase, semua kromosom berkelompok pada bidang equator dan penampakannya seperti lonceng tumpul (large dumb-bells). Pada tahap metafase terdapat enam puluh kromosom yang telah dibedakan. Secara umum berbentuk oval, ada beberapa ukuranya lebih besar dibandingkan yang lain dan terdistribusi pada bidang equator. Badan kromatid selalu berpindah ke kutub yang berlawanan dari sel. Kedudukan badan kromatid berada pada kumparan serabut, sementara pada kasus lainnya badan kromatid terlihat dalam sitoplasma.
Anafase adalah tahap setelah tahap metafase. Bivalen berbentuk lonceng dumb-bells terpisah dan tertarik pada kutub yang berlawanan. Badan kromatid berpindah dalam beberapa arah. Proses pembelahan berlanjut dan setahap demi setahap spermatosit primer masuk pada tahap telofase. Pada akhir telofase, kromosom yang lengkap berpindah ke kutub yang berlawanan kemudian dikelilingi oleh dinding nuklear yang tipis. Sitoplasma tertarik kedalam dua bagian dan selama proses itu, kumparan serabut juga ikut tertarik pada pusatnya dan terbentuk "zwischenkorper". Ketika tahap ini selesai dua spermatosit sekunder dihasilkan.
Tahap Spermatosit Sekunder
Spermatosit sekunder terbentuk pada tahap telofase, terjadinya pembelahan reduksi. Tidak ada periode istirahat, sel diasumsikan pada tahap metafase, dan kromosom pada garis atas equator berbentuk lonceng (dumb-bells). Tahap anafase dan telofase dengan urutan sistematik, menghasilkan pembelahan spermatosit sekunder ke bentuk spermatid. Dua tipe dari spermatid yang terbentuk, satu tipe yang mengandung badan kromatid tunggal dalam sitoplasma sedangkan tipe kedua tanpa badan kromatid.
21 Transformasi dari Spermatids menjadi Spermatozoa
Produksi spermatid pada pertama kali adalah kecil dan nukleinya mengandung masa yang sebagian besar material kromatid. Sitoplasma homogen dan sentromer yang menonjol ditemukan.
Pada tipe kedua dari perkembangan spermatid, sitoplasma mengandung sentrosomer juga memiliki badan kromatid. Perubahan yang dialami oleh spermatid dalam pergantian bentuk ke spermatozoa terjadi dalam nukleus. Masa kromatin dari nukleus berkurang secara bertahap. Pertama pada kumpulan besar dari khromatin yang membelah ke dalam masa granular dan beberapa bagian diperlengkap sampai tahap akhir, tiga badan kromatin yang bundar. Struktur kromatin tetap spherical. Spermatid menampakan struktur yang jelas, yaitu; 1) nukleus yang homogen dengan suatu karyosome sebagai tubuh pusat, 2) sitoplasma ditemukan, 3) sentromer, dan 4) masa mitokondria.
Nukleus mengembara pada satu kutub spermatid, sementara pada kutub yang berlawanan terdapat vakuola. Pada saat bersamaan masa mitokondria berkelilingi di antara nukleus dan vakuola dan akhirnya mengisi seluruh ruangan. Peningkatan ukuran sentrosomer dan mengambil bagian dalam pusat massa mitokondria. Masa mitokondria kemudian ditransformasi ke dalam kumparan, dan sentrosomer menempati pusat pada bagian dalam. Bagian atas dari nukleus terletak pada ruang ini. Bersamaan badan karyosome terletak pada pusat nukeus pindah ke atas bagian tengah dari nukleus secara langsung di bawah sentrosomer. Perubahan terakhir pada vakuola dalam anterior. Pada vakuola terlihat seperti gelembung kecil yang berupa cairan.
Selama sentrosomer dan badan karyosome berada pada nukleus, dalam bentuk memanjang menyerupai batang yang disebut dengan badan sentral. Pada awal, badan sentral berbentuk seperti lonceng (a dumb-bell), panjangnya tidak diketahui. Tahap ini terjadi pembukaan pada bagian tengah dari terluar atau akhir distal dari kantung kedua dan bersamaan dengan badan sentral lebih memanjang dan bagian luar berbentuk pipa tipis yang terhubung dengan pembukaan distal dalam kantung kedua.
22 Kumparan kromatid terlihat lebih jelas sebagai akhir distal pada badan sentral. Penyatuan dua elemen kedalam struktur tunggal, nuklear-mitokhondrial menjadi pendek. Perubahan berlanjut kantung kedua tersusun rapat ke dalam kantung pertama. Bagian terluar dari badan sentral secara lengkap berubah ke bentuk tabung yang berlubang dengan dinding yang tipis. Menunjukkan beberapa struktur, mulai dari jari-jari yang berbentuk lingkaran pada penampilan spermatozoa. Setelah transformasi spermatid secara lengkap ke bentuk spermatozoa yang matang, dan pada tahap ini spermatid terlihat berbentuk oval atau bulat dengan lengan yang berbentuk lingkaran sangat rapat mengelilingi nuklear-mitokhondria.
Spermatozoa matang, menunjukkan badan sentral terletak pada bagian tengah, dikelilingi kantung kedua, kantung pertama, dan nuklear-mitokhondria. Struktur spermatozoa berupa badan sentral terdiri dari akhir distal dan akhir proksimal. Khromatin-ring, terletak pada bagian atas dari kantung kedua. Spermatozoa lepas pada saat matang, kemudian spermatozoa masuk ke dalam vas deferen, dan dikelilingi oleh kantung membran atau spermatofor.
Pengeluaran dari Spermatozoa Matang
Tahap pertama dalam pengeluaran spermatozoa adalah ektrusi dari kantung kedua. Kantung ini secara normal mengelilingi badan sentral dan melekat dalam kantung pertama. Ketika kantung kedua mulai dilepaskan pada ukuran dan waktu yang sama. Kantung ini dibawa kebagian atas dari kantung pertama dan terlihat seperti lingkaran yang tipis. Bersamaan dengan akhir distal dari badan sentral yang juga extruded, mendesak kantung tersebut pada bagian padat proximal, perubahan ke bentuk seperti duri. Ketika kantung kedua telah kompleks untuk bebas, kemudian kantung pertama memulai untuk evert dan melanjutkan proses ini sampai lengkap pada bagian luar. Selama eversi, pada akhir proximal dari badan sentral mendorong ke atas pada bagian tubular distal sampai yang kedua akhirnya bebas ke luar.
Pelepasan sempurna spermatozoa dapat terlihat pada Gambar 10, bagian atas terdiri dari kantung kedua, bagian dalam mengandung kantung pertama, dengan bentuk tubuh yang cenderung keatas seperti duri. Bagian bawah terdiri dari nuklear-mitokhondria, yang berubah ke bentuk kurang lebih seperti bola.
23 Pada beberapa kasus, tahap ini ditunjukkan pada Gambar terakhir dari proses spermatogenesis pada kepiting Cancer magister.
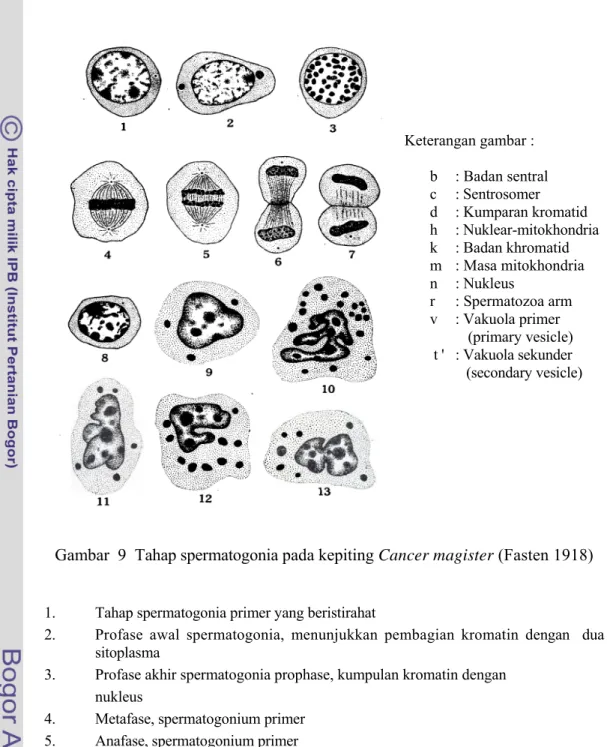
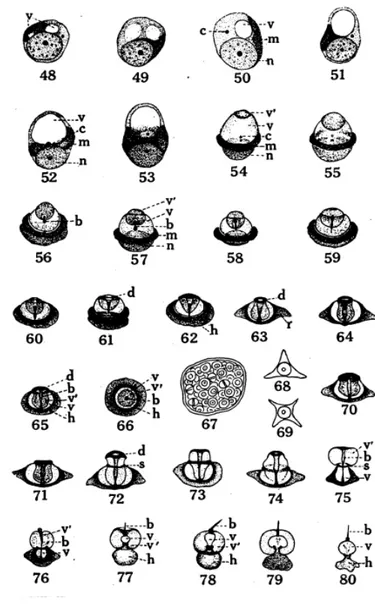
Gambar 9 Tahap spermatogonia pada kepiting Cancer magister (Fasten 1918)
1. Tahap spermatogonia primer yang beristirahat
2. Profase awal spermatogonia, menunjukkan pembagian kromatin dengan dua
sitoplasma
3. Profase akhir spermatogonia prophase, kumpulan kromatin dengan
nukleus
4. Metafase, spermatogonium primer
5. Anafase, spermatogonium primer
6-7. Telofase, spermatogonium primer
8. Tahap spermatogonia sekunder yang beristirahat
9-12. Sel nutrisi, nuklei irregular dan globul lemak dengan sitoplasma
13. Sel nutrisi, nukleus menunjukkan keadaan terdesak ke tengah terlihat peristiwa
amitosis. Keterangan gambar : b : Badan sentral c : Sentrosomer d : Kumparan kromatid h : Nuklear-mitokhondria k : Badan khromatid m : Masa mitokhondria n : Nukleus r : Spermatozoa arm v : Vakuola primer (primary vesicle) t ' : Vakuola sekunder (secondary vesicle)
24
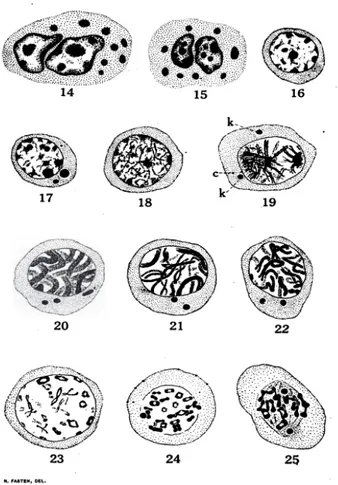
Gambar 10 Tahap Spermatosit Primer pada kepiting Cancer magister
(Fasten 1918)
14-15. Sel nutrisi, dengan dua nuklear diduga amitosis.
16-17. Awal profase, tahap spermatosit primer pada gambar 17. Dua masa kromatid dengan sitoplasma terwarnai dengan gelap.
18. Tahap leptoten
19. Tahap sintesis dan sinapsis. Tahap leptoten ini, berupa benang-benang yang
berpasangan, tersusun pada equator yang terbagi pada kutub synaptic dari sel. Keterangan : badan khromatid (k), dan sentrosome (c),terlihat dalam sitoplasma.
20. Tahap pachyten dan leptoten, sepasang benang membentuk geminius ditengah.
21-22. Tahap diploten.
23. Tahap post diploten
24. Pembentukan tetra, pada tahap ini dinding inti (nuklear) mulai membagi.
25. Perubahan tetra ke bentuk dumb-bells, sel masuk ke fase metafase
dan badan kromatid dikelilingi oleh ruang kosong pada kutub yang berlawanan.
25
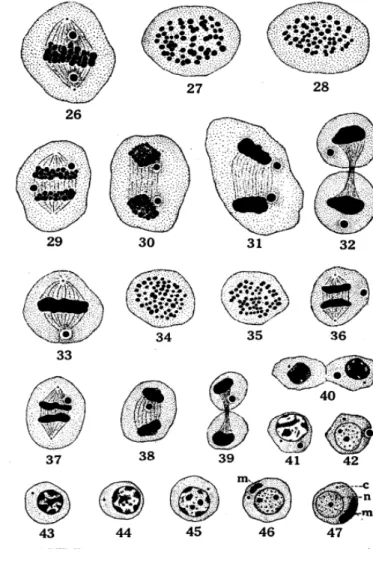
Gambar 11 Tahap Spermatosit Sekunder pada kepiting Cancer magister
(Fasten 1918)
26. Metafase, spermatosit primer menunjukkan badan khromatid pada kumparan
serabut pada kutub yang berlawanan.
27-28. Kutub spermatosit primer, menunjukkan 60 kromosom.
29-32. Tahap anafase dan telofase dari spermatosit primer menunjukkan badan kromatid pada kutub yang berlawanan. Gambar 32, badan kromatid yang diamati tetap berada pada sitoplasma.
33. Metafase spermatosit sekunder, menunjukkan badan kromatid tunggal
pada satu kutub.
34-35. Kutub spermatosit sekunder, menunjukkan 60 kromosom.
36-37. Tahap anafase dari spermatosit sekunder, menunjukkan kedudukan yang
berbeda, dimana badan kromatid mungkin menempati bagian dalam dinding sel.
38-39. Tahap telofase spermatosit sekunder, badan kromatid tunggal pada
26
40. Dua tipe spermatid yang terbentuk, satu tanpa badan kromatid dan yang
lainnya mengandung badan kromatid. Sentromer berwarna gelap ditemukan di sitoplasma pada kedua tipe spermatid.
41. Tahap awal dalam perubahan spermatid yang berisi badan kromatid
42. Pada tahap ini badan kromatid dikeluarkan dari spermatid
43-47. Tahap perubahan awal dari spermatid yang minus akan badan khromatid, reduksi
dari khromatin dan penampakan dari masa mitokhrondria (m) dalam sitoplasma. Pada gambar 47, karyosom tunggal menempati pusat dari nukleus.
Gambar 12 Transformasi Spermatid menjadi Spermatozoa pada kepiting Cancer magister (Fasten 1918)
48-53. Perubahan spermatid secara bertahap, dihasilkan dalam vakuola primer. terbentuk pada satu kutub, sementara nukleus (n) menempati kutub yang berlawanan, dan masa mitokhrondria (m) dan sentrosomer (c) juga menempati
27
54. Vakuola sekunder (t/) terlihat penampakan akhir distal pada vakuola primer.
55-58. Tahap transformasi dari spermatid dalam vakuola primer dan sekunder berubah ke dalam bentuk kantung, dan central body (b) terlihat seperti lonceng.
59-62. Perubahan spermatid menunjukkan lubang terluar akhir distal dari central body. pembentukan ring kromatin (d), dan penyatuan inti dan masa mitokondria kedalam bentuk nuclear-mitochondria (a).
63-64. Pembentukan arm dari spermatozoa.
65-66. Bagian sisi samping dan bawah menunjukkan secara detail struktur spermatozoa matang
67. Spermatofor, berisi spermatozoa matang.
68-69. Tipe spermatozoa, yang terdiri dari tiga dan empat bagian yang terlihat menggantung pada cancer magister.
70-78. Secara bertahap mengeluarkan spermatozoa matang ketika dipenuhi tekanan osmotik dengan larutan garam dari air laut.
79. Spermatozoa terlepas.
80. Keluarnya spermatozoa dari kantung sekunder dengan sempurna.
Pengaruh Lingkungan Terhadap Siklus Reproduksi
Faktor lingkungan seperti suhu, cahaya, dan ketersediaan pakan adalah faktor luar yang umum mempengaruhi reproduksi (Sastry 1983). Namun faktor-faktor ini mempunyai pengaruh yang berbeda terhadap rangkaian tahap-tahap gametogenesis maupun fase lain dalam siklus reproduksi. Primavera (1985) mengemukakan bahwa pakan berpengaruh terhadap pematangan gonad Penaeid, tetapi tidak demikian terhadap perkawinan dan pemijahan. Sedangkan suhu, salinitas, dan cahaya tidak saja berpengaruh terhadap pematangan gonad, tetapi juga terhadap kopulasi, pemijahan, pembuahan dan penetasan.