III. HASIL DAN PEMBAHASAN
1. Depresi Penangkarandalam (Inbreeding Depression) Kelapa Dalam Mapanget No.32 (DMT-32) Hasil Penyerbukan Tertutup Selama Empat Generasi
Depresi Penangkarandalam Berdasarkan Penanda Morfologi
Kelapa Dalam Mapanget (DMT) adalah salah satu aksesi kelapa yang diunggulkan karena masa produktifnya sampai 60 tahun, produksi buah tinggi, kadar minyak tinggi dan kualitas protein baik (Polnaja 1999; Rompas et al. 1989). Karakterisasi terhadap sifat morfologi dan molekular pada tanaman kelapa Dalam Mapanget No.32 (DMT-32) hasil penyerbukan pohon kelapa menggunakan campuran polen selama tiga generasi (DMT-32 S1, DMT-32 S2, DMT-32 S3) dan pada generasi keempat dilakukan penyerbukan menggunakan polen dari masing-masing individu pohon yang diserbuki ke pohon yang sama (DMT-32 S4) telah dilakukan. Populasi pohon kelapa generasi pertama hasil penyerbukan menggunakan campuran polen dari pohon-pohon terpilih yaitu DMT-32 S1 tidak bisa diamati karena sudah ditebang beberapa tahun sebelum penelitian ini dilakukan. Hasil penyerbukan tertutup tiga generasi berikutnya memperlihatkan terjadinya depresi penangkarandalam baik pada penanda morfologi yaitu pada karakter vegetatif, generatif dan komponen buah maupun penanda molekular. Karakter Vegetatif
Karakter vegetatif pada kelapa Dalam Mapanget No.32 (DMT-32) hasil penyerbukan menggunakan campuran serbuksari beberapa pohon terpilih ini menunjukkan telah terjadi depresi penangkarandalam pada beberapa karakter. Generasi kedua hasil penyerbukan DMT-32 S2 mengalami depresi penangkaran-dalam pada karakter lingkar batang 150 cm di atas permukaan tanah, dan lebar anak daun. Pada generasi DMT-32 S3 karakter yang mengalami depresi penangkarandalam adalah lingkar batang 20 cm dan 150 cm di atas permukaan tanah, tinggi 11 bekas daun, dan lebar anak daun. Pada DMT-32 S4 ditemukan lebih banyak karakter vegetatif yang mengalami depresi penangkarandalam dengan nilai yang semakin besar. Karakter-karakter tersebut adalah lingkar batang
20 cm dan 150 cm di atas permukaan tanah, tinggi 11 bekas daun, lebar anak daun, dan jumlah daun (Tabel 3).
Tabel 3. Depresi penangkarandalam pada karakter vegetatif generasi Kedua, ktiga dan keempat kelapa Dalam Mapanget (DMT-32)
Karakter
Rata-rata PD (%)
OP S2 S3 S4 S2 S3 S4
pjg anak daun 124.80 131.11 127.90 127.32 -5.05 -2.48 -2.01
jum anak daun 102.80 105.67 106.68 112.18 -2.78 -3.76 -9.13
lbr tgkai daun 6.55 6.96 6.95 7.24 -6.19 -6.10 -10.52 tbl tgkai daun 2.61 2.74 2.74 3.26 -5.15 -4.78 -24.72 pjg lamina 339.70 403.33 426.15 470.16 -18.73 -25.44 -38.40 pjg tgk daun 100.70 108.33 126.08 152.71 -7.58 -25.19 -51.65 jumlah daun 25.50 25.67 25.53 22.03 -0.65 -0.09 13.62 lgkr btg 20 cm d.p.t 166.80 167.22 145.10 157.89 -0.25 13.01 5.39 tg 11 bks daun 113.90 115.56 100.45 94.08 -1.45 11.80 17.40 lbr anak daun 5.97 5.90 5.60 5.68 1.17 6.15 4.87 lgkr btg 150cm d.p.t 101.10 97.78 92.98 86.53 3.28 8.03 14.41 Keterangan : PD = Penangkarandalam
OP = Penyerbukan terbuka ditanam 1957
S2 = DMT-32 generasi kedua hasil penyerbukan tertutup ditanam 1969 S3 = DMT-32 generasi ketiga hasil penyerbukan tertutup ditanam 1979 S4 = DMT-32 generasi keempat hasil penyerbukan sendiri ditanam 1994 d.p.t = dari permukaan tanah
pjg anak daun = panjang anak daun (cm) jum anak daun = jumlah anak daun (cm) lbr tgkai daun = lebar tangkai daun (cm) tbl tgkai daun = tebal tangkai daun (cm) pjg lamina = panjang lamina (cm) pjg tgk daun = panjang tangkai dan (cm) lgkr btg = lingkar batang (cm)
tg 11 bks daun = tinggi 11 bekas daun (cm) lbr anak daun = lebar anak daun (cm)
Karakter vegetatif yang mengalami depresi penangkarandalam pada sifat lingkar batang 20 cm dari permukaan tanah terutama pada DMT-32 S4, mengakibatkan ukuran lingkar pangkal batang dekat permukaan tanah (bole) semakin kecil. Depresi penangkarandalam pada sifat lingkar batang 150 cm dari permukaan tanah, dan pada sifat tinggi 11 bekas daun yang semakin besar pada setiap generasi menyerbuk tertutup, mengakibatkan batang kelapa menjadi lebih
kecil dan tanaman lebih pendek sehingga cenderung mendekati penampilan kelapa Genjah.
Pada kelapa tipe Dalam diameter dasar batang yang disebut bole (20 cm di atas permukaan tanah) dapat mencapai 80 cm yang dengan cepat mengecil menjadi + 40 cm pada ketinggian 150 cm di atas permukaan tanah (Child 1974). Kelapa yang memiliki batang besar dan kekar diduga memiliki perakaran lebih banyak sehingga dapat mengabsorbsi air dan unsur hara lebih banyak untuk menunjang pertumbuhan lebih baik (Novarianto et al. 1999). Karakter tinggi batang 11 bekas daun berkaitan dengan kecepatan tumbuh pohon kelapa. Di tahap awal laju pertambahan tinggi batang terbesar mencapai 150 cm per tahun dan akan semakin menurun hingga mencapai 10-15 cm per tahun ketika tanaman kelapa berumur 40 tahun atau lebih (Santos et al. 1996).
Sifat pertumbuhan batang yang diinginkan adalah pohon yang tidak terlalu cepat tinggi artinya pertumbuhan daun pada batang lebih rapat sehingga lebih mudah dalam pemanenan (Novarianto et al. 1999). Depresi penangkarandalam yang terjadi pada sifat tinggi 11 bekas daun lebih menguntungkan karena lebih sesuai dengan pertumbuhan batang yang diinginkan pada tanaman kelapa. Umur tanaman kelapa dewasa berkorelasi positif dengan jumlah bekas daun. Jumlah seluruh bekas daun pada batang dibagi dengan 13 (rata-rata jumlah daun terbentuk dalam satu tahun) akan memberikan perkiraan umur pohon dalam tahun (Machindapala & Pinto 1991).
Secara umum tanaman kelapa dewasa memproduksi daun sebanyak 12-16 helai per tahun, dan setiap daun dapat bertahan selama tiga tahun lalu gugur dan meninggalkan bekas daun pada batang (Santos et al. 1996). Jumlah daun dalam satu mahkota berkorelasi positif dengan jumlah tandan yang keluar, umumnya setiap pelepah menghasilkan satu tandan. Sifat daun lainnya yang dinginkan pada tanaman kelapa adalah panjang petiole agak pendek, lebih lebar dan tebal sehingga lebih kuat menyangga tandan buah yang memiliki buah banyak dan berat (Tampake 1987; Novarianto et al. 1999). Tangkai daun yang panjang tidak mampu menopang tandan buah sehingga tandan keluar dari tangkai dan terkulai (Mahmud et al. 1990). Jumlah anak daun serta panjang dan lebarnya
mempengaruhi luas daun relatif sehingga lebih banyak cahaya matahari yang dapat diserap untuk proses fotosintesa (Awuy et al. 1999).
Hasil penelitian ini panjang tangkai daun, panjang lamina, tebal tangkai daun, lebar tangkai daun, jumlah anak daun, dan panjang anak daun yang tidak mengalami depresi penangkarandalam melainkan lebih tinggi dari pada DMT-32 menyerbuk terbuka (Tabel 3). Tangkai daun yang lebih panjang tidak diharapkan berkaitan dengan ketidakmampuan menyangga tandan dan buah kelapa yang berat dan banyak, sedangkan lamina yang panjang berpengaruh pada pemanfaatan lahan di bawah kelapa, dan jarak tanam kelapa yang lebih lebar. Karakter tebal tangkai daun, lebar tangkai daun, jumlah anak daun, dan panjang anak daun lebih sesuai dengan karakter daun yang diinginkan.
Fungsi daun antara lain untuk menopang tandan buah yang keluar dari ketiak daun. Jumlah daun dalam satu mahkota tanaman kelapa berkorelasi positif dengan jumlah tandan buah yang keluar. Umumnya tiap pelepah menghasilkan satu tandan.
Karakter Generatif
Depresi penangkarandalam pada karakter generatif memiliki kecenderungan yang sama dengan karakter vegetatif yaitu semakin banyak karakter yang mengalami depresi penangkarandalam dengan persentase yang semakin meningkat dari generasi ke generasi hasil penyerbukan tertutup pohon kelapa menggunakan campuran polen pada setiap generasi hingga generasi ketiga dan hasil penyerbukan menggunakan polen dari pohon yang sama pada generasi keempat (Tabel 4).
Pada generasi DMT-32 S2 hanya tiga karakter yang tidak mengalami depresi penangkarandalam, sedangkan pada DMT-32 S3 tinggal dua yang tidak mengalami depresi penangkarandalam. Pada DMT-32 S4 semua karakter generatif telah mengalami depresi penangkarandalam yang persentasenya meningkat (Tabel 4). Depresi penangkarandalam ditemukan pada karakter jumlah spikelet, jumlah tandan, dan jumlah buah kelapa Dalam Bali yang diserbuk tertutup (Wardiana 1996).
Karakter yang berhubungan langsung dengan produksi buah kelapa per pohon adalah jumlah tandan, dan jumlah bunga betina per tandan. Pada tanaman kelapa yang berproduksi tinggi rata-rata menghasilkan bunga betina 150 buah per pohon per tahun, berproduksi sedang rata-rata 120 bunga betina per pohon per tahun, dan berproduksi rendah rata-rata 41 bunga betina per pohon per tahun (Menon & Pandalai 1960). Produksi jumlah betina ini sangat dipengaruhi oleh kondisi lingkungan yaitu iklim dan tanah terutama pada tipe Genjah (Santos et al. 1996). Kelapa Dalam Mapanget adalah salah satu tipe kelapa Dalam yang yang memiliki jumlah bunga betina yang banyak yaitu rata-rata 24.89 buah per infloresensia (Rompas et al. 1989).
Tabel 4. Depresi penangkaran dalam pada karakter generatif generasi
Kedua, ketiga dan keempat kelapa Dalam Mapanget No 32 (DMT-32)
Karakter Rata-rata PD (%) OP S2 S3 S4 S2 S3 S4 bh / tdn (butir) 7.60 9.67 6.73 1.92 -27.19 11.51 74.72 tdn / phn (bh) 15.60 14.44 13.28 6.11 7.41 14.90 60.86 bunga betina (bh) 37.00 29.11 19.28 5.21 21.32 47.91 85.92 pjg tgkai tdn (cm) 65.50 43.11 40.50 28.24 34.18 38.17 56.89 pjg B. jantan (cm) 32.40 28.89 30.68 22.97 10.84 5.31 29.09 lbr tgkai tdn (cm) 3.23 3.39 3.59 2.16 -4.92 -11.07 33.11 tbl tgkai tdn (cm) 2.12 2.41 2.31 1.31 -13.73 -8.73 38.18 Keterangan : PD = Penangkarandalam
OP = Penyerbukan terbuka ditanam 1957
S2 = DMT-32 generasi kedua hasil penyerbukan tertutup ditanam 1969 S3 = DMT-32 generasi ketiga hasil penyerbukan tertutup ditanam 1979 S4 = DMT-32 generasi keempat hasil penyerbukan sendiri ditanam 1994 tdn / phn = jumlah tandan per pohon
bh / tdn = jumlah buah per tandan pjg tgkai tdn = panjang tangkai tandan
pjg B. jantan = panjang rangkaian bunga jantan lbr tgkai tdn = lebar tangkai tandan
tbl tgkai tdn = tebal tangkai tandan
Faktor utama penyebab rendahnya produksi buah pada tanaman kelapa adalah gugur buah akibat gagalnya polinasi, persaingan dalam mempertahankan fotosintat, kurangnya sarana penyerbuk seperti angin dan serangga, tercucinya serbuk sari akibat hujan, dan rendahnya kesuburan serbuk sari (Williams 1970).
Pada penyerbukan alami umumnya aborsi hasil pembuahan sebesar 50-70%, terutama pada musin kemarau (Santos et al. 1995).
Pembentukan dan jumlah bunga berperan banyak gen pada tanaman dikotil maupun monokotil (Irish 2000; Whiple et al. 2004; Zahn et al. 2005). Jumlah bunga betina, persentase jumlah bunga betina menjadi buah, dan jumlah buah pada tanaman kelapa dipengaruhi oleh gen-gen aditif (Nambiar & Nambiar 1970). Sedangkan jumlah tandan dan jumlah buah lebih banyak dipengaruhi oleh gen aditif dibanding gen dominan (Munier et al. 1984).
Sifat infloresensia penting lainnya adalah panjang tangkai tandan. Depresi penangkarandalam pada panjang tangkai tandan sebesar 56.89% dengan panjang tangkai tandan 28.2 cm, lebih menguntungkan karena tangkai tandan yang pendek dan tebal lebih bisa menahan buah yang banyak dan berat. Tandan yang secara keseluruhan lebih panjang, berbuah banyak, dan buah lebih berat akan lebih mudah patah (Mahmud et al. 1990).
Karakter-karakter vegetatif dan generatif yang berkorelasi positif maupun negatif secara nyata dengan jumlah buah per tandan adalah jumlah daun, panjang daun, jumlah anak daun, panjang rangkaian bunga jantan dan jumlah bunga betina per infloresensia (Miftahorrachman et al. 1992; Novarianto et al. 1999). Sedangkan produksi buah tinggi pada tanaman kelapa disebabkan oleh superioritas genetik yang terjadi karena adanya kesesuaian kombinasi gen dalam kondisi heterozigot dan adanya gen pengendali hasil (produksi buah) yang dominan (Harland 1957).
Kelapa Dalam Mapanget No.32 yang diseleksi berdasarkan produksi tandan dan buah banyak dan dibuat menyerbuk tertutup menggunakan polen dari pohon-pohon terseleksi pada setiap generasi selama tiga generasi dan penyerbukan individu menggunakan polen yang berasal dari pohon yang sama pada generasi keempat, telah menampakkan kecenderungan terjadinya depresi penangkarandalam secara fisik pada setiap generasi (Gambar 1, Gambar 2, dan Gambar 3).
Pada generasi kedua terlihat pada Gambar 1 (kiri) adalah pohon kelapa berbuah paling banyak yaitu 165 butir per pohon per tahun dari pohon DMT-32 S2 No.4, sedangkan Gambar 1 (kanan) adalah pohon kelapa yang berbuah paling
sedikit yaitu 104 butir per pohon per tahun dari pohon DMT-32 S2 No.5. Meskipun pohon No.5 DMT-32 S2 berbuah paling sedikit tetapi masih tergolong pohon kelapa berbuah banyak (>80 butir per pohon per tahun).
Gambar 1. Penampilan pohon dengan jumlah buah terbanyak dan tersedikit pada generasi DMT -32 S2
Pada generasi ketiga Gambar 2 (kiri) adalah pohon kelapa berbuah paling banyak yaitu 135 butir per pohon per tahun dari generasi DMT-32 S3 No.7, sedangkan Gambar 2 (kanan) adalah pohon kelapa yang berbuah paling sedikit yaitu 48 butir per pohon per tahun dari DMT-32 S3 No.38. Hasil penyerbukan tertutup DMT-32 generasi ketiga telah menampakkan kecenderungan penurunan produksi buah yang cukup besar sebagai akibat terjadinya depresi penangkarandalam.
Gambar 2. Penampilan pohon dengan jumlah buah terbanyak dan tersedikit pada generasi DMT -32 S3
Pada generasi keempat hasil penyerbukan menggunakan polen dari pohon yang sama, pohon kelapa berbuah paling banyak mendekati pohon kelapa berbuah paling sedikit pada generasi kedua. Pada Gambar 3 (kiri) adalah pohon kelapa berbuah paling banyak yaitu 105 butir per pohon per tahun dari generasi DMT-32 S4 No.36. Gambar 3 (kanan atas) adalah pohon kelapa yang berbuah sedang yaitu 60 butir per pohon per tahun dari DMT-32 S4 No.17, sedangkan Gambar 3 (kanan bawah) adalah pohon kelapa tidak berbuah atau sampai dengan umur 11 tahun belum berbuah dari DMT-32 S4 No.8. Pada DMT-32 S4 ditemukan sebanyak 16 pohon kelapa yang sampai dengan umur 11 tahun belum berbuah dari 38 pohon populasi tersebut.
Gambar 3. Penampilan pohon dengan jumlah buah terbanyak, buah sedang
dan tidak berbuah pada generasi DMT -32 S4 Komponen Buah
Penyerbukan tertutup pada populasi pohon kelapa DMT-32 tidak selalu menyebabkan depresi penangkarandalam pada komponen buah dari generasi OP, generasi kedua, dan generasi ketiga. Pada generasi kedua hanya satu komponen buah yang tidak mengalami depresi yaitu sabut buah. Sedangkan pada generasi ketiga air kelapa dan tempurung kelapa bertambah berat. Pada generasi keempat semua komponen buah mengalami depresi penangkarandalam (Tabel 5). Secara umum depresi penangkarandalam meningkat persentasenya dari DMT-32 S2 ke DMT-32 S4. Hasil ini sejalan dengan penelitian yang dilakukan oleh Akuba (2002).
Tabel 5. Depresi penangkarandalam pada karakter generatif generasi kedua, ketiga, dan keempat kelapa Dalam Mapanget No.32(DMT-32)
Karakter Rata-rata (g) PD (%) OP S2 S3 S4 S2 S3 S4 Buah utuh 1290.26 1173.98 1223.13 675.04 9.01 5.20 47.68 Sabut 320.30 323.61 318.44 173.40 -1.03 0.58 45.86 Air kelapa 310.20 260.36 324.15 297.78 16.07 -4.50 4.00 Tempurung 195.00 178.60 199.00 93.95 8.41 -2.05 51.82 Daging buah 425.55 393.43 398.44 195.59 7.55 6.37 54.04 Keterangan : PD = Penangkarandalam
OP = Penyerbukan terbuka ditanam 1957
S2 = DMT-32 generasi kedua hasil penyerbukan tertutup ditanam 1969 S3 = DMT-32 generasi ketiga hasil penyerbukan tertutup ditanam 1979 S4 = DMT-32 generasi keempat hasil penyerbukan sendiri ditanam 1994 Persentase depresi penangkarandalam pada karakter komponen buah DMT-32 S4 sudah sangat tinggi, umumnya diatas 45% kecuali air kelapa 4%. Ukuran dan berat buah pada DMT-32 S4 semakin kecil mendekati uk uran buah kelapa Genjah. Hasil penelitian ini juga memperlihatkan bahwa persentase depresi penangkarandalam pada DMT-32 S3 lebih rendah kecuali pada berat sabut dibandingkan dengan DMT-32 S2 dan meningkat lagi pada DMT-32 S4. Turunnya persentase depresi penangkarandalam pada DMT-32 S3 dan meningkat lagi pada DMT-32 S4 diduga disebabkan oleh (1) fenotipe yang terekspresi dikontrol oleh alel-alel resesif, ketika seleksi baik secara alami maupun buatan dilakukan alel-alel tersebut tidak terseleksi ke generasi berikutnya. Hilangnya alel-alel resesif yang menyebabkan terjadinya depresi penangkarandalam disebut penghilangan (Purging) (Ritland 1996); (2) penggunaan campuran serbuksari (polen) pada setiap melakukan penyerbukan buatan sampai pada generasi ketiga dan pada generasi keempat penyerbukan terhadap pohon terpilih menggunakan polen yang berasal dari pohon yang sama.
Berat buah tanpa sabut memiliki korelasi tinggi dan positif dengan berat kopra sehingga berat kopra dapat diduga dari berat buah tanpa sabut. Faktor yang menentukan tingginya hasil kopra per pohon adalah jumlah buah per tahun dan berat kopra per butir (Thampan 1981).
Pada tanaman kelapa pengaruh depresi penangkarandalam secara alami terekspresi pada kelapa tipe Genjah yang memiliki batang lebih pendek, lebih kecil dan tidak memiliki bole, buah kecil, kualitas kopra relatif rendah, dan kurang toleran tehadap variasi lingkungan dibandingkan dengan kelapa tipe Dalam (Akuba, 2002).
Depresi Penangkarandalam Berdasarkan Penanda Molekular
Dari 19 primer untuk lokus SSR yang digunakan dalam penelitian ini 15 primer memperlihatkan pita-pita polimorfis dengan jumlah alel antara 3 sampai 6 sehingga didapatkan seluruhnya 60 alel atau rata-rata 4 alel per lokus. Alel- lel yang telah ditransfer menjadi data genotipe setiap individu dianalisis menggunakan program POPGENE untuk mengetahui besarnya frekuensi heterozigot, indeks fiksasi dan depresi penangkarandalam DMT-32 S2, DMT-32 S3, dan DMT-32 S4.
Depresi penangkarandalam secara tidak langsung dapat dilihat pada tingkat heterozigositas dalam populasi. Depresi penangkarandalam direpresentasikan oleh koefisien inbreeding atau indeks fiksasi yang merupakan ukuran turun atau naiknya heterozigositas dalam populasi (Smouse 1986). Indeks fiksasi menggambarkan penurunan heterozigositas di dalam populasi relatif terhadap total populasi yang dikaitkan dengan adanya seleksi dan penghanyutan (drift). Indeks fiksasi positif mengindikasikan berkurangnya genotipe heterozigot, sedangkan indeks fiksasi negatif menunjukkan peningkatan genotipe heterozigot. Koefisien penangkarandalam menggambarkan perbedaan heterozigositas pengamatan dari heterozigositas harapannya di dalam populasi yang kawin tidak acak. Hasil analisis terhadap jumlah alel, frekuensi heterozigot, indeks fiksasi dan depresi penangkarandalam DMT-32 S2, DMT-32 S3 DAN DMT-32 S4 berdasarkan penanda SSR disajikan pada Tabel 6.
Nilai rata-rata heterozigot aktual dari kelapa DMT-32 S2, DMT-32 S3 dan DMT-32 S4 lebih kecil dari nilai heterozigot harapan, berarti bahwa pada setiap generasi hasil penyerbukan tertutup heterozigositas cenderung berkurang, sehingga struktur genotipe akan mengarah ke peningkatan homozigositas untuk lokus-lokus yang dideteksi (Tabel 6).
Tabel 6. Jumlah alel, Frekuensi heterozigot, Indeks fiksasi dan depresi penangkarandalam DMT-32 S2, DMT-32 S3 DAN DMT-32 S4 berdasarkan penanda SSR
Keterangan: PD = Penangkarandalam
Heterozigositas adalah karakteristik yang penting dari suatu lokus. Heterozigositas suatu lokus menentukan suatu individu bersifat heterozigot untuk lokus tertentu dalam suatu populasi. Untuk penanda molekular, suatu lokus dengan heterozigositas lebih dari 70% menunjukkan penanda tersebut memiliki polimorfisme tinggi (Ott 1992). Heterozigositas menurun dalam suatu populasi dapat terjadi karena adanya hambatan aliran gen dalam populasi, dan meningkatnya hubungan kekerabatan antar individu pohon (Gregorius & Namkoong 1983).
Depresi penangkarandalam ditemukan pula pada populasi kelapa DMT-32 S2, DMT-32 S3 dan DMT-32 S4 menggunakan penanda mikrosatelit, masing-masing sebesar 2.78%, 10.54%, 15.74%. Nilai depresi penangkarandalam menggunakan 15 lokus SSR dalam penelitian ini lebih rendah jika dibandingkan dengan yang diperoleh Akuba (2002) pada populasi kelapa DMT S3 dan DMT S4
Lokus Jumlah Alel Heterozigot Harapan (HE) Heterozigot Aktual (Ho) S2 S3 S4 S2 S3 S4 CNZ 05 4 0.905 0.805 0.707 0.567 0.725 0.500 CNZ 09 3 0.877 0.787 0.874 0.667 0.700 0.605 CNZ 18 6 0.961 0.941 0.891 0.888 0.850 0.763 CNZ 21 3 0.895 0.873 0.863 0.881 0.675 0.631 CNZ 51 3 0.864 0.794 0.764 0.778 0.579 0.425 CnCirA3 3 0.877 0.799 0.677 0.555 0.750 0.631 CnCirA9 4 0.891 0.859 0.791 0.555 0.500 0.605 CnCirC3’ 4 0.885 0.855 0.835 0.789 0.575 0.509 CnCirC7 3 0.797 0.758 0.658 0.556 0.525 0.447 CnCir E2 4 0.808 0.794 0.698 0.778 0.650 0.552 CnCirE10 5 0.929 0.878 0.729 0.889 0.825 0.821 CnCirE12 5 0.963 0.901 0.763 0.889 0.850 0.631 CnC ir F2 5 0.963 0.897 0.763 0.889 0.850 0.842 CnCirH4’ 4 0.834 0.794 0.734 0.778 0.700 0.737 CnCirH7 4 0.838 0.799 0.738 0.778 0.775 0.815 Rataan 4 0.886 0.836 0.766 0.749 0.689 0.649 Indeks Fiksasi - 0.074 0.049 0.041 PD - 0.028 0.105 0.157
masing- masing 94.08% dan 97.66%. Perbedaan dapat terjadi karena populasi yang digunakan Akuba adalah populasi kelapa DMT campuran dari No.10, No.32, dan No.55 dengan jumlah sampel yang lebih sedikit. Sedangkan dalam penelitian ini digunakan hanya zuriat dari DMT No.32, dan semua tanaman pada setiap generasi hasil penyerbukan tertutup digunakan dalam penelitian ini. Selain itu jenis dan jumlah lokus SSR yang digunakan juga berbeda.
Depresi penangkarandalam berdasarkan penanda mikrosatelit (SSR) ditemukan pula pada generasi ketiga dan keempat populasi kelapa DMT-32. Nilai depresi penangkarandalam berdasarkan penanda SSR ini lebih rendah dibandingkan dengan nilai depresi penangkarandalam berdasarkan penanda morfologi. Tetapi memiliki kecenderungan yang sama yaitu nilainya semakin besar pada populasi-populasi generasi berikutnya, yang menunjukkan bahwa semakin banyak lokus- lokus SSR yang homozigot pada setiap kali dilakukan penyerbukan tertutup pada setiap generasi populasi kelapa DMT-32. Tingkat heterozigositas merupakan ukuran kekekaran suatu individu artinya makin tinggi nilai heterozigositas makin kekar suatu individu (Smouse 1986). Menurunnya kekekaran tanaman terlihat dengan jelas pada populasi DMT-32 S4.
Adanya depresi penangkarandalam pada tanaman yang bersifat menyerbuk terbuka seperti kelapa dapat dijelaskan melalui dua hipotesis yaitu hipotesis overdominan dan hipotesis dominan. Hipotesis overdominan menerangkan bahwa depresi penangkarandalam adalah akibat dari peningkatan homozigositas pada karakter yang lebih menguntungkan dalam genotipe heterozigot. Hipotesis dominan menerangkan bahwa peningkatan homozigositas mengakibatkan peluang bertemunya alel-alel resesif yang merugikan menjadi lebih tinggi (Falconer & Mackay 1996). Alel-alel merugikan ini berpengaruh pada kekekaran tanaman secara umum. Pada awal pertumbuhan, banyak tanaman muda generasi keempat hasil penyerbukan sendiri kelapa Dalam Mapanget (DMT) mati. Kematian tanaman muda ini diduga karena bertemunya alel-alel resesif yang merugikan. Alel-alel resesif yang merugikan dan bukan alel overdominan adalah faktor utama penyebab depresi penangkarandalam (Ritland 1996). Berdasarkan hipotesis dominan, terjadinya depresi penangkarandalam pada populasi tanaman kelapa tipe Dalam dapat mengindikasikan adanya aksi gen dominan. Depresi
penangkarandalam terjadi hanya karena efek dominan, artinya pada karakter yang tidak ada dominansi tidak akan terdeteksi adanya depresi penangkarandalam (Filho 1997).
Dalam upaya merakit kelapa hibrida yang unggul diperlukan tetua-tetua homosigot. Persilangan antar dua tetua homozigot yang berbeda akan menghasilkan zuriat yang unggul akibat pengaruh heterosis (Allard 1960). Kelapa tipe Dalam pada umumnya menyerbuk silang sehingga zuriatnya memiliki penampilan beragam karena genotipe yang heterozigot (Menon & Pandalai 1960; Child 1974; Fremond et al. 1966; Foale 1992). Untuk mendapatkan tetua homozigot pada kelapa Dalam dapat diperoleh melalui teknik penyerbukan tertutup sampai beberapa generasi. Zuriat yang dihasilkan dari proses penyerbukan tertutup akan memiliki derajat homozigot yang meningkat dengan penurunan vigor sebagai akibat adanya peristiwa depresi penangkarandalam.
Depresi penangkarandalam dan heterosis adalah dua fenomena bertolak belakang yang banyak dipelajari pada tanaman dan binatang. Depresi penangkarandalam berkaitan dengan menurunnya kekekaran turunan hasil penyerbukan tertutup, sebaliknya heterosis berkaitan dengan keunggulan hibrida (F1) melebihi nilai atau rata-rata kedua tetuanya (Stebbins 1958).
Hasil penyerbukan pohon-pohon terpilih menggunakan polen campuran dari DMT-32 sampai generasi ketiga dan polen dari pohon yang menjadi tetua betina pada generasi keempat, mengakibatkan aliran gen tidak berlangsung secara bebas. Akibatnya beberapa lokus SSR yang digunakan tidak dalam kesetimbangan Hardy-Weinberg (HW) disajikan pada Tabel 7.
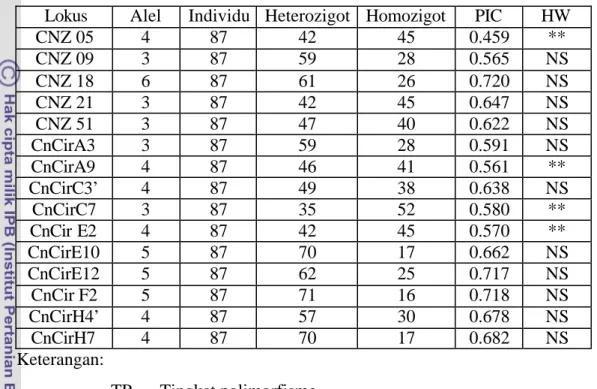
Dari Tabel 7 dapat dilihat ada empat lokus yang mengalami perubahan frekuensi gen yang besar sehingga lokus SSR CNZ05, CnCirA9, CnCirC7, dan CnCirE2 tidak pada kesetimbangan Hardy-Weinberg. Kejadian ini menunjukkan bahwa akibat seleksi yang dilakukan berdasarkan karakter jumlah buah lalu diikuti dengan persilangan di antara pohon-pohon yang berkerabat dekat, telah mengakibatkan beberapa lokus tidak lagi diwariskan secara bebas dari generasi ke generasi. Migrasi, mutasi dan seleksi adalah faktor- faktor penyebab terjadinya perubahan frekuensi gen dari generasi ke generasi dalam suatu populasi besar, sedangkan pada populasi kecil pemilihan sampel secara acak merupakan
faktor yang lebih berperan dalam perubahan frekuensi gen dibandingkan faktor lainnya (Liu 1998).
Tabel 7. Jumlah alel, individu heterozigot, individu homozigot, tingkat polimorfisme dan kesetimbangan Hardy-Weinberg
Lokus Alel Individu Heterozigot Homozigot PIC HW
CNZ 05 4 87 42 45 0.459 ** CNZ 09 3 87 59 28 0.565 NS CNZ 18 6 87 61 26 0.720 NS CNZ 21 3 87 42 45 0.647 NS CNZ 51 3 87 47 40 0.622 NS CnCirA3 3 87 59 28 0.591 NS CnCirA9 4 87 46 41 0.561 ** CnCirC3’ 4 87 49 38 0.638 NS CnCirC7 3 87 35 52 0.580 ** CnCir E2 4 87 42 45 0.570 ** CnCirE10 5 87 70 17 0.662 NS CnCirE12 5 87 62 25 0.717 NS CnCir F2 5 87 71 16 0.718 NS CnCirH4’ 4 87 57 30 0.678 NS CnCirH7 4 87 70 17 0.682 NS Keterangan: TP = Tingkat polimorfisme HW = Kesetimbangan Hardy-Weinberg
** = Tidak pada kesetimbangan Hardy-Weinberg NS = Pada kesetimbangan Hardy-Weinberg
2. Pelacakan Tetua Kelapa Dalam Mapanget No.32 Melalui Aliran Gen (Gene Flow) Berdasarkan Penanda Mikrosatelit (SSR)
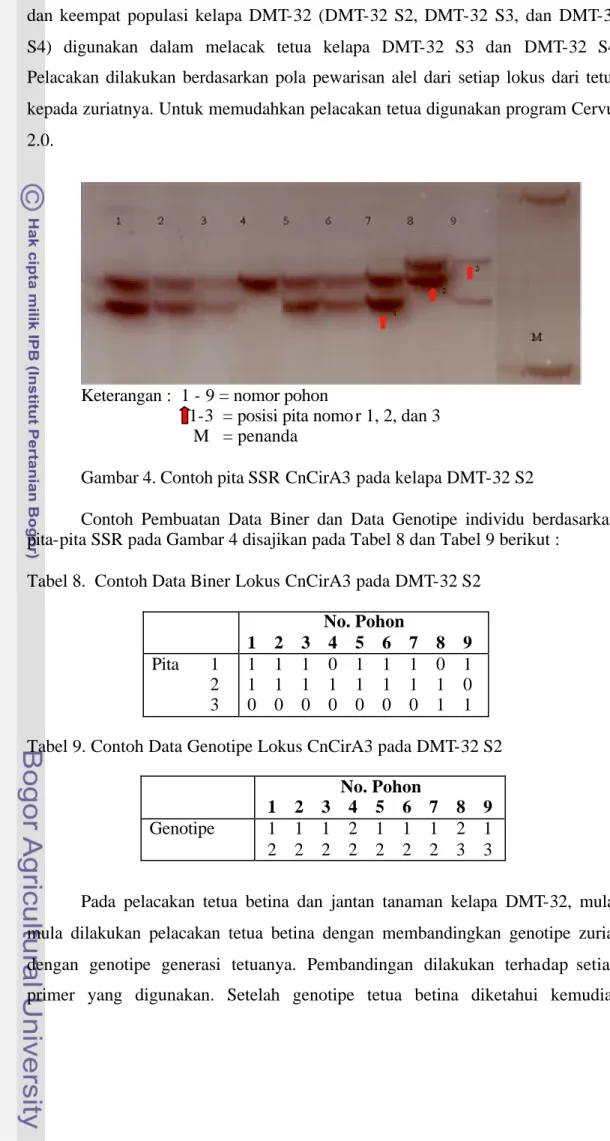
Hasil analisis SSR DNA kelapa DMT-32 S2, DMT-32 S3, dan DMT-32 S4 menggunakan 15 primer menunjukkan polimorfisme dengan ukuran alel berbeda, setiap primer terdiri atas 3-6 alel sehingga seluruhnya berjumlah 60 alel. Setiap individu memiliki 1-2 pita yang merupakan tipikal bagi penanda SSR yang bersifat kodominan bagi organisme diploid (Gambar 4). Mikrosatelit (SSR) adalah suatu jenis penanda DNA yang dapat digunakan untuk mempelajari genotipe individu (Mullis & Faloona 1987).
Pita-pita DNA dibuat dalam simbol genotipe menggunakan angka, dari generasi hasil penyerbukan tertutup kelapa 32 S2, 32 S3, dan DMT-32 S4. Data genotipe tersebut digunakan dalam melacak pohon kelapa tetua dari kelapa DMT-32 S3 dan DMT-32 S4. Semua genotipe dari generasi kedua, ketiga,
dan keempat populasi kelapa DMT-32 (DMT-32 S2, DMT-32 S3, dan DMT-32 S4) digunakan dalam melacak tetua kelapa DMT-32 S3 dan DMT-32 S4. Pelacakan dilakukan berdasarkan pola pewarisan alel dari setiap lokus dari tetua kepada zuriatnya. Untuk memudahkan pelacakan tetua digunakan program Cervus 2.0.
Keterangan : 1 - 9 = nomor pohon
1-3 = posisi pita nomo r 1, 2, dan 3 M = penanda
Gambar 4. Contoh pita SSR CnCirA3 pada kelapa DMT-32 S2
Contoh Pembuatan Data Biner dan Data Genotipe individu berdasarkan pita-pita SSR pada Gambar 4 disajikan pada Tabel 8 dan Tabel 9 berikut :
Tabel 8. Contoh Data Biner Lokus CnCirA3 pada DMT-32 S2
Tabel 9. Contoh Data Genotipe Lokus CnCirA3 pada DMT-32 S2
Pada pelacakan tetua betina dan jantan tanaman kelapa DMT-32, mula-mula dilakukan pelacakan tetua betina dengan membandingkan genotipe zuriat dengan genotipe generasi tetuanya. Pembandingan dilakukan terhadap setiap primer yang digunakan. Setelah genotipe tetua betina diketahui kemudian
No. Pohon 1 2 3 4 5 6 7 8 9 Pita 1 2 3 1 1 1 0 1 1 1 0 1 1 1 1 1 1 1 1 1 0 0 0 0 0 0 0 0 1 1 No. Pohon 1 2 3 4 5 6 7 8 9 Genotipe 1 1 1 2 1 1 1 2 1 2 2 2 2 2 2 2 3 3
dianalisis untuk mengetahui genotipe tetua jantan dari setiap individu generasi zuriat dengan generasi tetuanya. Pelacakan tetua jantan dilakukan dengan membandingkan genotipe zuriat dengan genotipe tetua betina yang telah diketahui dengan individu tetua yang memiliki genotipe paling sesuai sebagai pasangan tetua.
Mikrosatelit sebagai penanda kodominan juga telah digunakan untuk menganalisis tetua akasia (Prihatini et al. 2006), Cervus elaphus (Pemberton et al. 1992), dan ikan Lepomis macrochirus (Neff et al. 2000). Menurut Marshall et al (1998) beberapa faktor yang menyebabkan pelacakan tetua tidak berhasil adalah jumlah kandidat tetua yang besar, hubungan kekerabatan (relatedness) kandidat tetua yang tinggi, dan kesalahan dalam pelaksanaan penelitian.
Untuk menganalisis kandidat tetua jantan digunakan likelihood ratio. Individu dengan nilai likelihood ratio tertinggi (the most likely male parent) merupakan kandidat utama tetua jantan. Untuk memudahkan analisis digunakan program Cervus 2.0 (Meagher 1986; Marshall et al. 1998).
Hasil pelacakan tetua DMT-32 S3 dan DMT-32 S4 menggunakan program Cervus 2.0 disajikan pada Tabel 10 dan Tabel 11.
Hasil pelacakan tetua menggunakan program Cervus 2.0 menunjukkan bahwa sebanyak 21 zuriat DMT-32 S3 dapat dilacak tetuanya dengan tingkat kesesuaian 95% dan 19 zuriat lainnya pada tingkat kesesuaian 80% (Tabel 10). Sedangkan pada DMT-32 S4 sebanyak 23 zuriat dapat dilacak tetuanya pada tingkat kesesuaian 95% dan 15 zuriat lainnya pada tingkat kesesuaian 80% (Tabel 11).
Hasil pelacakan tetua DMT-32 S3 memperlihatkan bahwa ada tiga individu yang benar-benar hasil penyerbukan tertutup dari satu pohon yaitu dua pohon kelapa DMT-32 S3 No.25 dan No.39 merupakan zuriat dari DMT-32 S2 No.8, dan satu pohon yaitu DMT-32 S3 No.26 merupakan zuriat dari DMT-32 S2 No.3. Hasil pelacakan tetua dari DMT-32 S4 menunjukkan bahwa ada 5 individu yang benar-benar merupakan hasil penyerbukan dan zigot dengan polen yang berasal dari pohon tertentu. Tiga pohon kelapa yang merupakan zuriat dari pohon DMT-32 S3 No.28 adalah DMT-32 S4 No.8, No.10, dan No.22, satu pohon
adalah zuriat dari DMT-32 S3 No.32 yaitu DMT-32 S4 No.11, dan satu pohon adalah zuriat DMT-32 S3 No.35 yaitu DMT-32 S4 No.30 (Tabel 11).
Tabel 10. Pelacakan Tetua betina dan jantan Kelapa DMT-32 generasi S3 berdasarkan 15 lokus mikrosatelit (SSR)
Generasi S3 Tetua Betina Tetua jantan Ket. Generasi S3 Tetua Betina Tetua jantan Ket. S3-1 S2-7 S2-9 * S3-21 S2-9 S2-8 S3-2 S2-2 S2-6 * S3-22 S2-1 S2-4 S3-3 S2-7 S2-5 * S3-23 S2-1 S2-5 S3-4 S2-8 S2-3 * S3-24 S2-2 S2-9 S3-5 S2-5 S2-3 * S3-25 S2-8 S2-8 * S3-6 S2-1 S2-4 S3-26 S2-3 S2-3 * S3-7 S2-2 S2-6 * S3-27 S2-8 S2-4 * S3-8 S2-1 S2-9 S3-28 S2-8 S2-5 S3-9 S2-3 S2-5 S3-29 S2-8 S2-7 * S3-10 S2-5 S2-8 S3-30 S2-7 S2-9 S3-11 S2-3 S2-1 S3-31 S2-2 S2-8 S3-12 S2-7 S2-1 * S3-32 S2-4 S2-7 S3-13 S2-8 S2-5 S3-33 S2-5 S2-7 * S3-14 S2-3 S2-9 * S3-34 S2-3 S2-2 * S3-15 S2-3 S2-7 * S3-35 S2-2 S2-6 S3-16 S2-6 S2-3 * S3-36 S2-2 S2-7 * S3-17 S2-7 S2-1 S3-37 S2-7 S2-3 * S3-18 S2-2 S2-9 * S3-38 S2-5 S2-3 S3-19 S2-2 S2-8 * S3-39 S2-8 S2-8 S3-20 S2-8 S2-5 S3-40 S2-1 S2-4 *
Keterangan : * sesuai pada taraf 95% tanpa * sesuai pada taraf 80%
S2 dan S3 = hasil penyerbukan generasi kedua dan ketiga Angka dibelakang S2 dan S3= nomor pohon masing- masing generasi
Individu- individu pohon kelapa yang mempunyai yang mempunyai pohon kelapa tetua betina dan jantan yang sama pada populasi DMT-32 S3 adalah No.6, No.22, dan No.40 zuriat dari persilangan pohon 32 S2 No.1 x No.4: DMT-32 S3 No.2, No.7, dan No.35 zuriat dari persilangan pohon DMT-DMT-32 S2 No.2 x No.6; DMT-32 S3 No.18 dan No.24 zuriat dari persilangan pohon DMT-32 S2 No.2 x No.9; 32 S3 No.19 dan No.31 zuriat dari persilangan pohon DMT-32 S2 No.2 x No.8; DMT-DMT-32 S3 No.13, No.20, dan No.28 zuriat dari persilangan pohon DMT-32 S2 No.8 x No.5; DMT-32 S3 No.1 dan No.30 zuriat dari persilangan pohon DMT-32 S2 No.7 x No.9; DMT-32 S3 No.12 dan No.17 zuriat
dari persilangan pohon DMT-32 S2 No.7 x No.1; DMT-32 S3 No.5 dan No.38 zuriat dari persilangan pohon DMT-32 S2 No.5 x No.3 (Tabel 10).
Tabel 11. Pelacakan Tetua betina dan jantan Kelapa DMT-32 generasi S4 berdasarkan 15 lokus mikrosatelit (SSR)
Generasi S4 Tetua Betina Tetua Jantan Ket Generasi S4 Tetua Betina Tetua Jantan Ket S4-1 S3-28 S3-12 S4-20 S3-34 S3-8 S4-2 S3-28 S3-26 S4-21 S3-33 S3-14 S4-3 S3-8 S3-32 * S4-22 S3-28 S3-28 * S4-4 S3-34 S3-33 * S4-23 S3-12 S3-35 S4-5 S3-1 S3-18 * S4-24 S3-1 S3-3 S4-6 S3-4 S3-28 * S4-25 S3-8 S3-32 * S4-7 S3-32 S3-10 * S4-26 S3-28 S3-12 S4-8 S3-28 S3-28 * S4-27 S3-5 S3-36 * S4-9 S3-28 S3-12 * S4-28 S3-11 S3-32 * S4-10 S3-28 S3-28 S4-29 S3-10 S3-6 * S4-11 S3-32 S3-32 S4-30 S3-35 S3-35 * S4-12 S3-4 S3-8 * S4-31 S3-28 S3-4 S4-13 S3-3 S3-14 * S4-32 S3-33 S3-36 S4-14 S3-33 S3-1 * S4-33 S3-34 S3-5 * S4-15 S3-23 S3-35 S4-34 S3-35 S3-26 S4-16 S3-23 S3-6 * S4-35 S3-4 S3-34 S4-17 S3-18 S3-1 * S4-36 S3-1 S3-14 * S4-18 S3-18 S3-5 * S4-37 S3-3 S3-14 * S4-19 S3-34 S3-1 * S4-38 S3-3 S3-21 Keterangan : * sesuai pada taraf 95%
tanpa * sesuai pada taraf 80%
S3 dan S4 = DMT-32 generasi ketiga dan keempat
Angka dibelakang S3 dan S4= nomor pohon masing- masing generasi
Dari Tabel 11 dapat dilihat individu-individu pohon kelapa pada DMT-32 S4 yang merupakan zuriat dari persilangan pohon DMT-32 S3 No.28 x No.12 adalah No.1, No.9, dan No.26. Sedangkan zuriat dari persilangan pohon DMT-32 S3 No.8 x No.32 adalah DMT-32 S4 No.3 dan No.25.
Hasil penelusuran menunjukkan pula bahwa semua individu pada generasi DMT-32 S2 menjadi tetua dari DMT-32 S3, sedangkan pada populasi DMT-32 S3 beberapa nomor pohon tidak menjadi tetua dari zuriat DMT-32 S4. Hasil ini dapat dipahami karena sebelum dilakukan penyerbukan pada setiap generasi, diseleksi
terlebih dahulu pohon-pohon yang berproduksi tinggi. Nomor- nomor pohon yang berproduksi rendah tidak digunakan sebagai tetua. Hasil penelusuran ini jika dikaitkan dengan data produksi buah per pohon per tahun yang diamati selama penelitian, menunjukkan bahwa pada populasi DMT-32 S2 semua pohon masih berpotensi produksi lebih dari 100 butir per pohon per tahun sehingga semuanya digunakan sebagai tetua untuk DMT-32 S3. Sedangkan pada DMT-32 S3 beberapa pohon di antaranya berproduksi rendah yaitu berpotensi produksi kurang dari 80 butir per pohon per tahun sehingga tidak digunakan sebagai tetua untuk generasi DMT-32 S4 (Tabel 12).
Berdasarkan hasil pelacakan menggunakan program Cervus 2.0 pada Tabel 10 dan Tabel 11, maka dapat dibuat silsilah dari setiap nomor pohon pada populasi kelapa DMT-32 generasi selfing kedua seperti pada Gambar 5 sampai dengan Gambar 13.
Dari Gambar 5 dapat dilihat bahwa DMT-32 S2 No.1 merupakan tetua betina dan jantan dari masing- masing empat individu DMT-32 S3. Lima dari zuriatnya adalah tetua betina dan jantan dari 13 individu DMT-32 S4. Zuriat DMT-32 S2 No.1 merupakan tetua betina dari pohon kelapa DMT-32 S3 No.6 dan No.8 artinya benih dari pohon-pohon kelapa tersebut dipanen dari DMT-32 S2 No.1, sedangkan pohon kelapa DMT-32 S2 No.1 sebagai tetua jantan dari
DMT-32 S3 No.11, No.12 dan No.23. Tiga nomor pohon kelapa DMT-32 S3 yang lain yaitu No.17, No.22, dan No.40 tidak menjadi tetua dari DMT-32 S4. Sebanyak tujuh pohon kelapa DMT-32 S4 dipanen dari zuriat DMT-32 S2 No.1, sedangkan enam nomor pohon kelapa DMT-32 S4 lainnya menerima polen dari DMT-32 S2 No.1. Dengan demikian 13 individu pohon kelapa generasi DMT-32 S4 merupakan zuriat dari DMT-32 S2 No.1.
Dari Gambar 6 dapat dilihat bahwa DMT-32 S2 No. 2 adalah tetua betina dari delapan individu DMT-32 S3 dan tetua jantan dari satu individu DMT-32 S3, artinya delapan zuriat tersebut dipanen dari pohon kelapa DMT-32 S2 No.2 dan satu pohon lainnya DMT-32 S2 No.2 menyumbang polen pada pohon kelapa yang lain. Empat nomor DMT-32 S3 yaitu pohon No.18, No.34, No.35, dan No.36 adalah tetua dari DMT-32 S4 sedangkan 5 nomor pohon yang lain tidak menjadi tetua dari DMT-32 S4. Dengan demikian 14 individu pohon kelapa generasi DMT-32 S4 merupakan zuriat dari DMT-32 S2 No.2. Pada DMT-32 S4 ditemukan satu nomor pohon yaitu DMT-32 S4 No.30 merupakan hasil penyerbukan dan zigot dengan polen yang berasal dari pohon yang sama yaitu DMT-32 S3 No.35.
Dari Gambar 7, menunjukkan bahwa DMT-32 S2 No.3 merupakan tetua betina dari dua individu DMT-32 S3, tetua jantan dari delapan individu DMT-32 S3, dan menjadi tetua betina dan sekaligus tetua jantan dari satu individu DMT-32 S3 yaitu DMT-32 S3 No.26. Enam nomor pohon kelapa zuriat dari DMT-32 S2 No.3 menjadi tetua betina dan jantan dari 18 individu DMT-32 S4, dan 5 nomor pohon tidak menjadi tetua dari DMT-32 S4. Dari Gambar 7 juga dapat dilihat ada delapan pohon dari populasi DMT-32 S4 yang dipanen dari zuriat DMT-32 S2 No.3. Pohon-pohon kelapa DMT-32 S4 pohon No.6, No.12, dan No.35 dipanen dari DMT-32 S3 No.4; DMT-32 S4 pohon No.27 dipanen dari DMT-32 S3 No.5; DMT-32 S4 pohon No.28 dipanen dari DMT-32 S3 No.11; dan DMT-32 S4 pohon No.4, No.19, dan No.29 dipanen dari DMT-32 S3 No.34. Sedangkan 10 nomor pohon kelapa DMT-32 S4 lainnya dipanen dari pohon kelapa DMT-32 S3 yang lain, zuriat DMT-32 S2 No.3 berfungsi sebagai tetua jantan.
Gambar 8, memperlihatkan bahwa DMT-32 S2 No.4 memiliki 5 zuriat yaitu sebagai tetua betina dari satu individu dan tetua jantan dari empat individu DMT-32 S3, artinya hanya DMT-32 S3 No.32 yang benihnya dipanen dari pohon DMT-32 S2 No.4. Dua dari lima zuriat DMT-32 S2 No.4 yaitu DMT-32 S3 No.6 dan No.32 menjadi tetua betina dan jantan dari enam individu DMT-32 S4. DMT-32 S3 No.6 berperan sebagai tetua jantan, sedangkan DMT-32 S3 No.32 berperan sebagai tetua betina DMT-32 S4 dari No.7 dan No.11 artinya benih dari
kedua pohon tersebut dipanen dari pohon kelapa DMT-32 S3 No.32, dan sebagai tetua jantan dari DMT-32 S4 pohon No.3 dan No.25.
Dari Gambar 9 dapat dilihat bahwa DMT-32 S2 No.5 merupakan tetua betina dari 4 individu DMT-32 S3 dan tetua jantan dari 6 individu DMT-32 S3, artinya empat nomor pohon kelapa DMT-32 S3 dipanen dari individu DMT-32 S2 No.5, sedangkan lainnya dipanen dari pohon lain yang diserbuki dengan polen individu DMT-32 S2 No.5. Enam nomor pohon diantaranya menjadi tetua betina
dan jantan dari 24 individu individu DMT-32 S4, dan empat lainnya tidak menjadi tetua dari DMT-32 S4 yaitu pohon No.9, No.13, No.20, No.38. DMT-32 S3 No.3 adalah tetua dari DMT-32 S4 yaitu pohon No.13, No.24, No.37, dan No.38. Pohon kelapa DMT-32 S4 No.8 dan No.22 merupakan hasil zigot dengan polen yang berasal dari pohon yang sama yaitu DMT-32 S3 No.28.
Dari Gambar 10, dapat dilihat bahwa dari DMT-32 S2 No.6 memiliki 4 zuriat yaitu DMT-32 S3 pohon No.2, No.7, No.16 dan No.35. Hanya DMT-32 S3 No.35 yang menjadi tetua dari 4 individu DMT-32 S4 yaitu pohon No.15, No.23, No.30 dan No.34. Pohon kelapa DMT-32 S4 No.30 dan No.34 dipanen dari individu DMT-32 S3 No.35, sedangkan individu DMT-32 S4 No.30 adalah hasil penyerbukan dan zigot dengan polen yang berasal dari pohon kelapa yang sama yaitu DMT-32 S3 No.35.
Dari Gambar 11, terlihat bahwa DMT-32 S2 No.7 merupakan tetua betina dan jantan dari masing- masing lima individu DMT-32 S3. Lima individu pohon kelapa DMT-32 S3 merupakan tetua dari 20 individu pohon kelapa DMT-32 S4, sedangkan lima individu pohon lainnya tidak menjadi tetua dari DMT-32 S4. Pada populasi kelapa DMT-32 S3 ada lima nomor pohon yang benihnya dipanen dari DMT-32 S3 No.7. Pada DMT-32 S4 masing- masing ada tiga nomor pohon yang benihnya dipanen dari DMT-32 S3 No.1 dan No.33, satu pohon dari DMT-32 S3 No.12, dan dua pohon dari DMT-32 S3 No.32 merupakan hasil zigot dengan
polen yang berasal dari pohon itu sendiri. Sedangkan 11 pohon DMT-32 S4 lainnya dipanen dari pohon-pohon kelapa 32 S3 yang bukan zuriat DMT-32 S2 No.7.
Gambar 12, dapat dilihat bahwa DMT-32 S2 No.8 memiliki 12 zuriat pada DMT-32 S3. Empat individu diantaranya adala h tetua dari 15 individu DMT-32 S4, dan delapan individu lainnya tidak menjadi tetua dari DMT-32 S4. Lima dari zuriat tersebut benihnya dipanen dari DMT-32 S2 No.8 artinya pohon kelapa No.8 dari generasi kedua berperan sebagai tetua betina, sedangkan tujuh nomor pohon lainnya dipanen dari pohon lainnya yang diserbuki dengan polen DMT-32 S2 No.8 (tetua jantan). Dapat dilihat pula pada populasi kelapa DMT-32 generasi ketiga ada dua pohon yaitu DMT-32 S3 No.25 dan No.39 merupakan zigot hasil penyerbukan denga n polen yang berasal dari pohon yang sama yaitu DMT-32 S2 No.8. Sedangkan pada populasi kelapa DMT-32 generasi keempat ada tiga pohon yaitu DMT-32 S4 No.8, No.10, dan No.22 merupakan zigot hasil penyerbukan dengan polen yang berasal dari pohon itu sendiri yaitu pohon kelapa DMT-32 S3 No.28.
Dari Gambar 13, dapat dilihat bahwa DMT-32 S2 nomor 9 merupakan tetua tetua betina dari satu individu, dan tetua jantan dari 6 individu DMT-32 S3. Hanya satu pohon yang benihnya dipanen dari pohon DMT-32 S2 No.9. Enam pohon yang lainnya merupakan hasil penyerbukan yang polennya berasal dari DMT-32 S2 No.9. Empat dari zuriat DMT-32 S2 No.9 merupakan tetua betina dan jantan dari 17 individu DMT-32 S4, tiga lainnya tidak menjadi tetua dari
32 S4. Pada 32 S4 ada tiga pohon yang benihnya dipanen dari DMT-32 S3 No.1, dua pohon benihnya dipanen dari pohon DMT-DMT-32 S3 No.8, dan dua pohon individu kelapa populasi DMT-32 S3 No.18, sedangkan DMT-32 S3 No. 14 berperan sebagai tetua jantan artinya tidak ada benih yang dipanen dari pohon tersebut.
Dengan diketahuinya silsilah masing- masing individu kelapa DMT-32 hasil penyerbukan sendiri generasi ketiga (DMT-32 S3) dan keempat (DMT-32 S4) secara jelas, maka peneliti kelapa yang akan mempelajari sifat-sifat tertentu yang berkaitan dengan produksi, ketahanan terhadap hama dan penyakit, kekeringan, protein, asam lemak dan lain- lain dapat memanfaatkan informasi ini. 3. Identikasi Pita Spesifik Terpaut Dengan Karakter Produksi Buah Pada
Tanaman Kelapa Berdasarkan Penanda Mikrosatelit (SSR)
Hasil pengamatan pada setiap generasi hasil penyerbukan tertutup dari pohon-pohon terseleksi pada setiap generasi kelapa 32 (32 S2, DMT-32 S3, dan DMT-DMT-32 S4), terhadap produksi buah per pohon per tahun dan jumlah tandan per pohon per tahun telah menunjukkan telah terjadi penurunan rata-rata produksi buah per pohon per tahun. Pada DMT-32 S2, pohon yang memiliki produksi buah per pohon per tahun paling rendah adalah DMT-32 S2 No.5 yaitu 104 butir. Pada DMT-32 S3, pohon yang memiliki produksi buah paling rendah adalah DMT-32 S3 No.38 yaitu 48 butir. Sedangkan pada DMT-32 S4 ditemukan 16 pohon yang sampai dengan umur 11 tahun setelah tanam tidak/belum berbuah. Data produksi buah per tandan per pohon dan jumlah tandan per pohon per tahun untuk generasi DMT-32 S2, DMT-32 S3, dan DMT-32 S4 disajikan pada Tabel 12.
Untuk mengetahui ada atau tidaknya pita spesifik yang berkaitan dengan produksi buah kelapa, maka populasi pohon kelapa tiap generasi DMT-32 dibuat pengelompokan berdasarkan produksi buah per pohon per tahun. Untuk DMT-32 S2 hanya mengelompok dalam satu kelompok karena semua pohon memiliki produksi di atas 100 butir. Pada DMT-32 S3 dalam empat kelompok yaitu > 100 butir/pohon, 80-99 butir/pohon, 60-79 butir/pohon, 40-59 butir/pohon. Pada
DMT-32 S4 mengelompok menjadi empat kelompok yaitu : > 100 butir/pohon, 8-60 butir/pohon, bertandan tanpa buah, dan tanpa tandan.
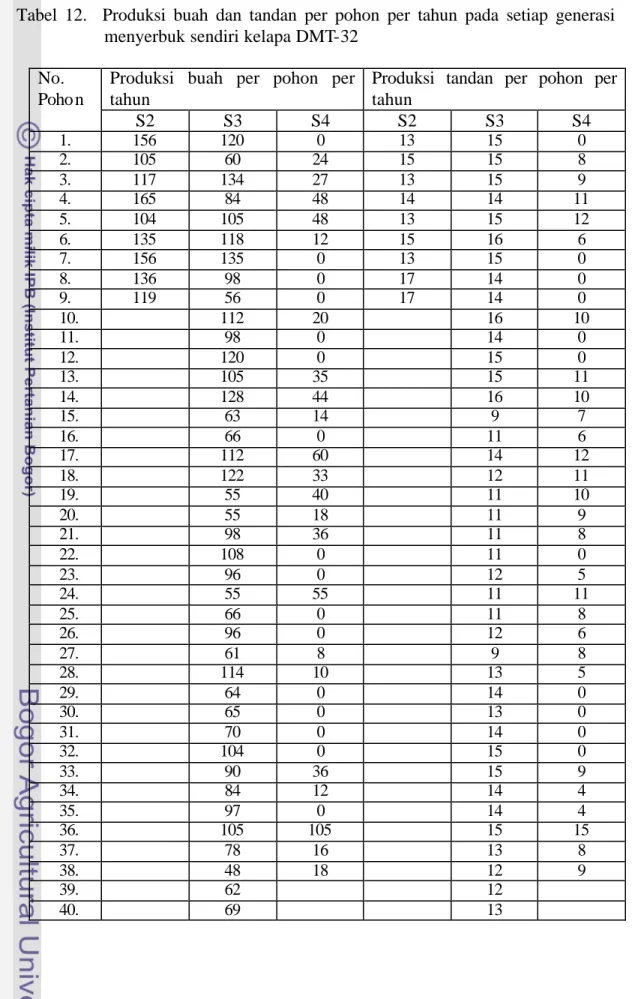
Tabel 12. Produksi buah dan tandan per pohon per tahun pada setiap generasi menyerbuk sendiri kelapa DMT-32
No. Poho n
Produksi buah per pohon per tahun
Produksi tandan per pohon per tahun S2 S3 S4 S2 S3 S4 1. 156 120 0 13 15 0 2. 105 60 24 15 15 8 3. 117 134 27 13 15 9 4. 165 84 48 14 14 11 5. 104 105 48 13 15 12 6. 135 118 12 15 16 6 7. 156 135 0 13 15 0 8. 136 98 0 17 14 0 9. 119 56 0 17 14 0 10. 112 20 16 10 11. 98 0 14 0 12. 120 0 15 0 13. 105 35 15 11 14. 128 44 16 10 15. 63 14 9 7 16. 66 0 11 6 17. 112 60 14 12 18. 122 33 12 11 19. 55 40 11 10 20. 55 18 11 9 21. 98 36 11 8 22. 108 0 11 0 23. 96 0 12 5 24. 55 55 11 11 25. 66 0 11 8 26. 96 0 12 6 27. 61 8 9 8 28. 114 10 13 5 29. 64 0 14 0 30. 65 0 13 0 31. 70 0 14 0 32. 104 0 15 0 33. 90 36 15 9 34. 84 12 14 4 35. 97 0 14 4 36. 105 105 15 15 37. 78 16 13 8 38. 48 18 12 9 39. 62 12 40. 69 13
Campuran DNA dari pohon-poho n kelapa yang produksinya ada dalam satu kelompok yang sama dicampur dan diPCR. Pita DNA yang spesifik adalah pita yang ditemukan selalu ada atau tidak ada pada pohon-pohon yang berbuah banyak atau tidak atau belum berbuah selama penelitian. Kalau ditemukan pita yang selalu ada pada pohon-pohon yang berbuah banyak atau tidak berbuah akan sangat membantu dalam melakukan seleksi bibit-bibit yang berpotensi produksi tinggi sejak dini.
Sekuen DNA pengapit SSR dirakit menjadi primer untuk digunakan mengamplifikasi lokus- lokus SSR tertentu menggunakan PCR. Setiap pita DNA mewakili satu alel dari suatu lokus. Perbedaan panjang pita DNA terjadi karena perbedaan jumlah unit pengulangan pada lokus-lokus mikrosatelit (SSR) tertentu (Gupta et al. 1996; Karp et al. 1997; Liu 1998).
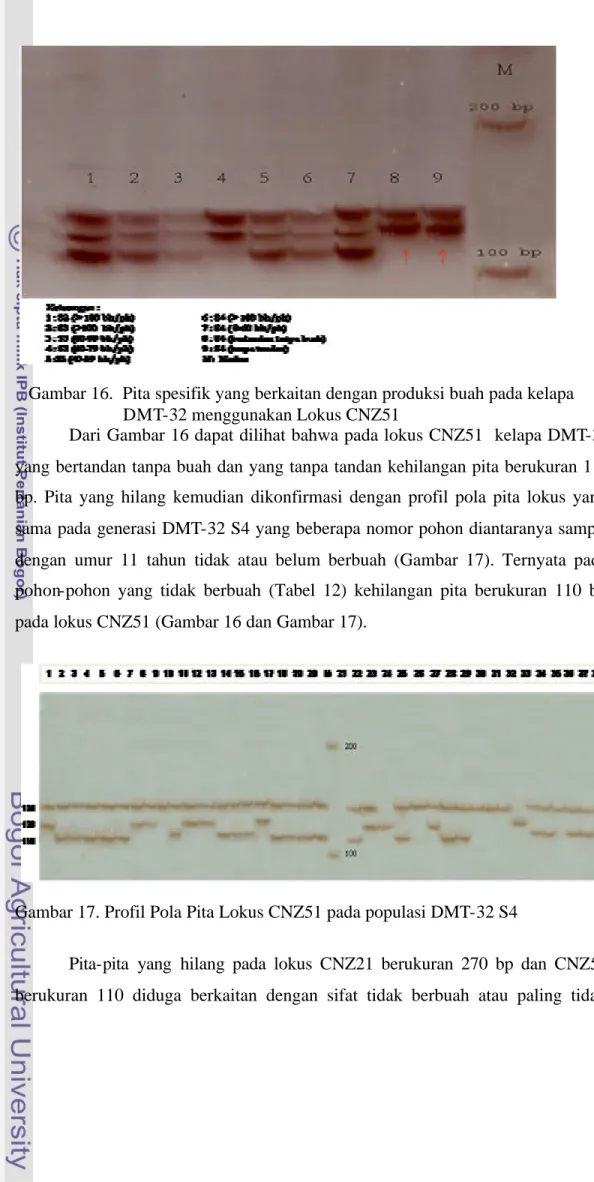
Hasil PCR-SSR yang diperoleh divisualisasikan menggunakan PAGE (Polyacrylamide Gel Electrophoresis) dengan pewarnaan perak (Creste et al 2001). Primer yang memberi informasi adanya pita yang khas pada hasil PCR campuran DNA berdasarkan penge lompokan yang telah dibuat, dibandingkan dengan hasil PCR pada DNA masing- masing individu pohon kelapa DMT-32 populasi DMT-32 S2, DMT-32 S3, dan DMT-32 S4. Hasil visualisasi 15 primer (lokus) SSR menggunakan PAGE pada pengelompokan DNA, ditemukan ada dua primer dari 15 primer yang digunakan memiliki pita khas yaitu CNZ21 dan CNZ51. Visualisasi SSR primer (lokus) CNZ21 pada campuran DNA berdasarkan pengelompokan (Gambar 14) dan pada generasi DMT-32 S4 (Gambar 15), sedangkan primer (lokus) CNZ51 pada campuran DNA (Gambar 16) dan pada generasi DMT-32 S4 (Gambar 17).
Gambar 14. Pita spesifik yang berkaitan dengan produksi buah pada kelapa DMT-32 menggunakan primer CNZ-21
Dari Gambar 14 dapat dilihat pola pita lokus CNZ21 pada DNA dari pohon kelapa yang dikelompokkan berdasarkan produksi buah. Hasil penelitian menunjukkan bahwa pada lokus CNZ-21 kelapa DMT-32 yang bertandan tanpa buah dan yang tanpa tandan kehilangan pita berukuran 270 bp. Hasil ini kemudian dikonfirmasi dengan profil pola pita lokus yang sama pada generasi DMT-32 S4 (Gambar 15). Hasil penelitian menunjukkan bahwa nomor-nomor pohon yang tidak berbuah sampai dengan umur 11 tahun setelah tanam kehilangan pita tersebut.
Keterangan : 1-38 = nomor pohon DMT-32 S4 M = Marker 100 bp DNA ladder
Gambar 16. Pita spesifik yang berkaitan dengan produksi buah pada kelapa DMT-32 menggunakan Lokus CNZ51
Dari Gambar 16 dapat dilihat bahwa pada lokus CNZ51 kelapa DMT-32 yang bertandan tanpa buah dan yang tanpa tandan kehilangan pita berukuran 110 bp. Pita yang hilang kemudian dikonfirmasi dengan profil pola pita lokus yang sama pada generasi DMT-32 S4 yang beberapa nomor pohon diantaranya sampai dengan umur 11 tahun tidak atau belum berbuah (Gambar 17). Ternyata pada pohon-pohon yang tidak berbuah (Tabel 12) kehilangan pita berukuran 110 bp pada lokus CNZ51 (Gambar 16 dan Gambar 17).
Gambar 17. Profil Pola Pita Lokus CNZ51 pada populasi DMT-32 S4
Pita-pita yang hilang pada lokus CNZ21 berukuran 270 bp dan CNZ51 berukuran 110 diduga berkaitan dengan sifat tidak berbuah atau paling tidak
sampai dengan umur 11 tahun setelah tanam nomor- nomor pohon kelapa tersebut belum berbuah.
Untuk mengetahui korelasi antar lokus yang dipakai dalam penelitian ini dan korelasi antara sifat morfologi dengan penanda molekular dilakukan analisis korelasi Pearson dengan Program komputer Minitab 14. Berdasarkan hasil analisis korelasi antar alel dari 15 lokus yang digunakan ternyata tidak saling berkaitan, tetapi antara alel (pita) dalam lokus yang sama saling berkorelasi pada taraf nyata >95% (Tabel 13).
Penanda mirosatelit (SSR) telah digunakan untuk pemetaan kromosom berbagai tanaman seperti peach (Dirlewangger et al. 1998), gandum (Kojima et al. 1998), barley (Davila et al. 1999), dan tomat (Saliba-Colombani et al. 2000). Lebrun at al (2001) membuat peta keterpautan (linkage map) pada kelapa Dalam RIT (Rennell Island Tall) dan menemukan 16 kelompok keterpautan, hasil ini sesuai dengan genom haploid kelapa. Tanaman kelapa merupakan tanaman diploid dengan jumlah kromosom 2 x = 32. Kelima belas lokus SSR yang digunakan dalam penelitian ini dibuat peta keterpautan (linkage map) menggunakan MAPMAKER dengan nilai LOD 3. Hasil pemetaan menunjukkan bahwa 15 lokus SSR terbagi ke dalam 15 kelompok artinya tidak ada lokus yang terpaut dengan yang lain dalam satu kelompok. Jika setiap kelompok dianggap sebagai kromosom, maka hasil pengelompokan (group) 15 lokus yang digunakan tersebar di lima belas kromosom berbeda pada tanaman kelapa DMT-32. Hasil pengelompokan ini tidak menggambarkan keseluruhan jumlah kromosom kelapa karena lokus SSR yang digunakan tidak cukup banyak. Menurut Herran et al (2000) dan Lebrun et al (2001) untuk membuat peta keterpautan maka diperlukan jumlah lokus SSR dan zuriat yang cukup banyak. Rendahnya korelasi antar lokus diduga karena lokus- lokus tersebut tersebar pada kromosom yang berbeda.
Tabel 13 Korelasi antar alel (pita) dari lokus CNZ21 dan CNZ51 Korelasi CNZ21-1 CNZ21-2 CNZ21-3 CNZ51-1 CNZ51-2 CNZ21-2 0.726 0.002 CNZ21-3 0.721 0.166 0.849 0.020 CNZ51-1 -0.174 0.107 0.197 0.067 0.213 0.047 CNZ51-2 0.095 0.379 -0.077 0.479 -0.201 0.062 0.691 0.000 CNZ51-3 0.028 0.495 -0.063 0.564 -0.026 0.509 0.578 0.099 0.760 0.015 Keterangan:
Nilai pada bagian atas setiap kolom adalah nilai korelasi antar alel dari lokus CNZ21 dan CNZ51
Nilai pada bagian atas bawah kolom adalah tingkat kepercayaan (Pearson)
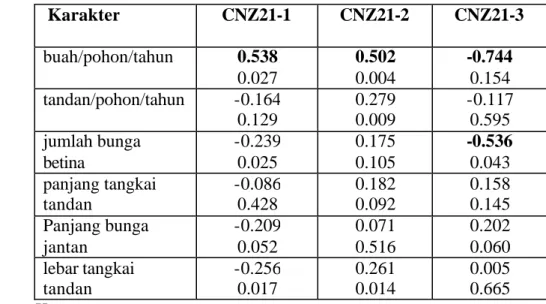
Hasil korelasi antara sifat morfologi dengan penanda molekular lokus CNZ21 menunjukkan bahwa pita yang hilang pada lokus tersebut yaitu pita CNZ21-3 yang berukuran 270 bp berkorelasi negatif dengan jumlah buah, sedangkan pita berukuran 250 bp dan 238 bp berkorelasi positif dengan jumlah buah pada tingkat kepercayaan 99%. Artinya jika pita CNZ21-270bp tidak ada maka pohon kelapa DMT-32 sampai dengan umur 11 tahun setelah tanam belum berbuah, sedangkan jika pita CNZ21-250bp dan CNZ21-238bp ada maka pohon-pohon kelapa DMT-32 sampai umur 11 tahun setelah tanam memiliki buah.
Tabel 14. Korelasi antara sifat generatif dengan Lokus CNZ21 Karakter CNZ21-1 CNZ21-2 CNZ21-3 buah/pohon/tahun 0.538 0.027 0.502 0.004 -0.744 0.154 tandan/pohon/tahun -0.164 0.129 0.279 0.009 -0.117 0.595 jumlah bunga betina -0.239 0.025 0.175 0.105 -0.536 0.043 panjang tangkai tandan -0.086 0.428 0.182 0.092 0.158 0.145 Panjang bunga jantan -0.209 0.052 0.071 0.516 0.202 0.060 lebar tangkai tandan -0.256 0.017 0.261 0.014 0.005 0.665 Keteranga n:
Nilai pada bagian atas setiap kolom adalah nilai korelasi antar alel dan karakter generatif
Nilai pada bagian atas bawah kolom adalah tingkat kepercayaan (Pearson)
Hasil korelasi antara sifat morfologi dengan penanda molecular lokus CNZ51, menunjukkan bahwa pita yang hilang pada lokus CNZ51-1 berkorelasi dengan sifat tidak atau belum berbuah sampai dengan umur 11 tahun setelah tanam. Lokus CNZ51-1 berukuran 110 bp berkorelasi dengan jumlah buah per pohon per tahun, jumlah tandan per pohon, dan jumlah bunga betina pada tingkat kepercayaan >99% (Tabel 14 dan Tabel 15). Hasil penelitian ini memperlihatkan pula bahwa jika pita CNZ51-110bp tidak ada maka jumlah tandan per pohon per tahun dan jumlah bunga betina per tandan semakin sedikit.
Penanda molekular dapat digunakan untuk mengidentifikasi karakter fenotipik dalam studi genetik populasi (Virk et al. 1996 ; Pillen et al. 2000). Identifikasi penanda molekular yang terpaut dengan karakter dan variasi dalam populasi sangat bergantung pada hubungan kekerabatan antara tetua yang digunakan untuk pemetaan populasi (Ford-Lloydet al. 2001).
Tabel 15. Korelasi antara sifat generatif dengan Lokus CNZ51 Karakter CNZ51-1 CNZ51-2 CNZ51-3 buah/pohon/tahun -0.692 0.002 -0.258 0.016 0.125 0.249 tandann/pohon/tahun -0.590 0.000 -0.330 0.002 0.178 0.100 jumlah bunga betina -0.426
0.036 -0.141 0.193 0.097 0.370 panjang tangkai tandan 0.350 0.001 -0.230 0.032 0.185 0.086 Panjang bunga jantan 0.418
0.000
-0.403 0.000
0.085 0.433 lebar tangkai tandan 0.463
0.000 -0.310 0.003 0.061 0.572 Keterangan:
Nilai pada bagian atas setiap kolom adalah nilai korelasi antar alel dan karakter generatif
Nilai pada bagian atas bawah kolom adalah tingkat kepercayaan (Pearson)
Dengan teridentifikasinya fragmen DNA spesifik yang berkorelasi dan berkaitan dengan karakter pohon yang tidak berbuah sampai dengan umur 11 tahun akan sangat membantu melakukan seleksi dini terhadap pohon-pohon kelapa yang kemungkinan tidak berbuah atau berbuah sangat lambat.