BAB V
ANALISIS SEKUEN LENGKAP GEN YANG
TERKAIT ADAPTASI KEDELAI TERHADAP
INTENSITAS CAHAYA RENDAH
Abstrak
Penelitian ini bertujuan untuk melakukan karakterisasi sekuen lengkap JJ3 yang merupakan salah satu kandidat gen yang telah berhasil diisolasi dari tanaman kedelai yang terkait adaptasi kedelai terhadap intensitas cahaya rendah. Karakterisasi sekuen lengkap mencakup analisis struktur sekuen lengkap, analisis kerangka translasi atau sekuen pengkode, analisis tingkat homologi, dan analisis domain protein yang dikode JJ3. Kegiatan karakterisasi dilakukan menggunakan jasa bioinformatika dan beberapa program pendukung. Hasil penelitian menunjukkan bahwa: sekuen lengkap cDNA JJ3 yang terkait dengan mekanisme adaptasi kedelai terhadap intensitas cahaya rendah memiliki coding sequence (CDS) 633 bp dengan 210 asam amino deduksi tersusun atas 136 asam amino terkonservasi dan 77 asam amino beragam. cDNA JJ3 yang diperoleh pada tanaman kedelai mempunyai fungsi yang sama dengan gen psaD yang terdapat pada tanaman tembakau kayu (Nicotiana sylvestris), padi (Oryza sativa), tomat (Lycopersicon esculentum), barley (Hordeum vulgare), bayam (Spinacia oleracea), Arabidopsis (Arabidopsis thaliana), dan kentang (Solanum tuberosum). Kata kunci: sekuen lengkap, gen, sekuen pengkodean (CDS), homolog, basa
ANALYSIS OF FULL LENGTH SEQUENCE OF GENE
RELATED TO ADAPTATION OF SOYBEAN
TO LOW LIGHT INTENSITY
Abstract
The aim of this study was to characterise full length sequence of JJ3, a cDNA corresponding to adaptation of soybean to low light intensity, obtain from soybean leaves under low light intensity. Characterization of the full length sequence of JJ3 was carried out using bioinformatics tools, in the aspects of analyses of prediction of sequence structure, open reading frame (ORF) or coding sequence, homology, and domain analysis of protein JJ3. The results of this study showed that: full length sequence of JJ3, a gene related to mechanism of adaptation of soybean to low light intensity, consisted of 633 bp nucleotides coding sequence was constituted of conserved domain (136 amino acids) and 77 varied domains (77 amino acids). JJ3 isolated from low irradiance tolerant soybean leaves have the same function with psaD from wood tobacco (Nicotiana sylvestris), rice (Oryza sativa), Spinach (Spinacia oleracea), tomato (Lycopersicon esculentum), barley (Hordeum vulgare), Arabidoposis (Arabidopsis thaliana), and potato (Solanum tuberosum).
__________
Key words: full length sequence, gene, coding sequence (CDS), homolog, nucleotide.
PENDAHULUAN
Latar Belakang
Organisme fotosintetik, termasuk tanaman kedelai, yang dikembangkan pada kondisi lingkungan intensitas cahaya rendah seperti di bawah tegakan perkebunan karet atau tumpang sari dengan tanaman lain yang berpotensi menaungi, hendaknya mampu mengembangkan berbagai perubahan atau mekanisme baik pada tingkat morfologi, fisiologi sampai tingkat molekuler sehingga mampu beraklimatisasi dalam kondisi lingkungan cahaya terbatas. Pada tingkat molekuler, kemampuan tanaman untuk beradaptasi pada lingkungan intensitas cahaya rendah tergantung dari ada tidaknya gen yang mengendalikan karakter-karakter yang terkait dengan kemampuan tanaman untuk memanfaatkan intensitas cahaya yang terbatas secara efisien untuk melangsungkan proses fotosintesis maksimum.
Hasil penelitian sebelumnya tentang ‘analisis genetik adaptasi kedelai terhadap intensitas cahaya rendah’ (Percobaan 2) dilaporkan bahwa adaptasi kedelai terhadap intensitas cahaya rendah yang terkait dengan proses fotosintesis dikendalikan sekurang-kurangnya oleh dua gen mayor untuk karakter kualitatif seperti kandungan klorofil dan oleh banyak gen untuk karakter kuantitatif seperti hasil dan morfologi daun. Handayani (2003) juga melaporkan bahwa terdapat banyak gen yang mengendalikan adaptasi kedelai terhadap intensitas cahaya rendah.
Menurut Allen dan Forsberg (2001); Pfannschmidt (2003); Fey et al. (2005), gen-gen fotosintesis yang berperan kunci dalam proses adaptasi tanaman terhadap intensitas cahaya rendah berasal dari inti (gen fotosintetik inti) dan ada juga yang berasal dari kloroplas (gen fotosintetik kloroplas). Gen-gen tersebut sangat menentukan kelangsungan proses fotosintesis termasuk transfer elektron di dalam pusat reaksi fotosistem I dan II. Terdapat 10 gen-gen fotosintetik inti yang mengkode protein komponen PSI, salah satunya adalah gen psaD (Hiyama 1997). Gen fotosintetik kloroplas terutama menyandi komponen protein dari 4 kelompok kompleks protein yang terdapat pada membran tilakoid yaitu: 6 gen untuk protein PSI (psaA-C,I,J,M), 12 gen untuk protein PSII (psbA-F,H-N), 6 gen untuk cyt
b6/f (petA-B,D,G), dan 6 gen untuk ATPase (atpA-B, atpE-I), serta gen yang mengkode subunit besar Rubisco (rbcL) (Hachtel 1997; Joshi 1997; Tyagi et al. 2000).
Sistem kerja gen-gen fotosintetik baik yang berasal dari inti maupun kloroplas umumnya dikontrol oleh status redoks yang tergantung dari aliran elektron fotosintetik di dalam fotosistem. Komponen dari rantai transpor elektron fotosintetik tersebut akan mengalami perubahan yang signifikan apabila mendapat stres lingkungan seperti intensitas cahaya rendah. Signal redoks seperti ini berfungsi sebagai parameter signaling yang mengontrol ekspresi gen fotosintetik kloroplas dan inti dan berperan penting di dalam koordinasi dari kedua kompartmen genetik tersebut (Fey et al. 2005). Perubahan intensitas cahaya yang diterima tanaman dapat mempengaruhi transport elektron fotosintetik dan dapat mengurangi efisiensi fotosintesis dengan cepat. Pada kondisi stres seperti ini, respon aklimatisasi dapat mempertahankan aliran elektron fotosintetik, dengan demikian fiksasi energi netto sebanyak mungkin dapat berlangsung terus.
Di dalam PSI, terminal kofaktor transfer elektron bekerja berangkai seperti kabel elektron yang menghubungkan elektron dari tempat awal pemisahan muatan yang terletak di dalam membran ke ferredoksin yang merupakan akseptor elektron solubel yang terletak di dalam subunit PsaD pada permukaan stroma membran tilakoid. Dengan demikian keberadaan subunit PsaD, protein ekstrinsik yang terletak periferal di sisi stroma, menjadi sangat penting. Menurut Lagoutte et al. (2001) subunit PsaD PSI yang dikode gen psaD berperan kunci dalam stabilitas transpor elektron fotosintetik di dalam kompleks PSI karena merupakan tempat docking ferredoxin (Fd).
Beberapa kajian tentang PsaD subunit PSI sudah dilaporkan sebelumnya antara lain analisis in vivo PsaD pada tanaman jagung (Heck et al. 1999), bentuk isoformis PsaD sub unit pada tanaman tembakau (Obokata et al. 1993), tingkat homologi dan struktur gen psaD dari beberapa tanaman seperti mentimun (Cucumis sativus), barley (Hordeum vulgare), tomat (Lycopersicum esculentum), tembakau kayu (Nicotiana sylvestris), dan bayam (Spinacia oleracea) (Hiyama 1997). Pada tanaman kedelai, penelitian tentang gen psaD dan gen-gen fotosintetik lain masih sangat terbatas.
Khumaida (2002) telah memulai kajian molekuler adaptasi kedelai terhadap cekaman naungan dengan melaporkan bahwa terdapat tiga fragmen cDNA yaitu JJ3, EE2, dan E3 yang merupakan kandidat gen fotosintetik yang positif terkait erat dengan gen yang mengkode protein kompleks membran tilakoid berturut-turut fotosistem I (PS-I), sitokrom, dan fotosistem II (PS-II). Kandidat gen JJ3 yang terkait dengan fotosistem I tersebut menjadi menarik untuk dikaji lebih lanjut karena homolog dengan psaD, gen yang mengkode protein PsaD pada subunit PSI. Selanjutnya, fragmen cDNA JJ3 yang berukuran 261 bp basa nukleotida tersebut berhasil dibuat sekuen lengkapnya (full length sequence) menjadi 841 bp (Sopandie et al. 2005).
Sejauh ini karakterisasi, konfirmasi fungsi dan analisis pola ekspresi sekuen lengkap cDNA JJ3 tersebut terkait dengan mekanisme molekuler adaptasi kedelai terhadap intensitas cahaya rendah belum dilakukan. Diharapkan sekuen lengkap cDNA JJ3 tersebut juga dapat didaftarkan di GenBank melalui situs NCBI (www.ncbi.nlm.nih.gov) untuk dapat diakses oleh pengguna di seluruh dunia. Dengan semakin banyak gen-gen yang diperoleh yang terkait dengan kemampuan tanaman beradaptasi pada lingkungan cekaman intensitas cahaya rendah, maka semakin banyak tersedia materi genetika dan molekuler yang dapat digunakan, dan akan semakin mempermudah dan mempercepat perbaikan adaptasi tanaman terhadap lingkungan cekaman tersebut.
Tujuan
Penelitian ini bertujuan untuk melakukan karakterisasi sekuen lengkap cDNA JJ3 yang diisolasi dari tanaman kedelai toleran naungan, dan selanjutnya dilakukan pendaftaran di database GenBank sebagai coding sequence lengkap (complete CDS) baru yang diisolasi dari tanaman kedelai toleran naungan.
BAHAN DAN METODE
BahanBahan yang digunakan adalah sekuen lengkap (full length) cDNA JJ3 yang telah diperoleh pada penelitian sebelumnya (Sopandie et al. 2005).
Analisis Sekuen Lengkap cDNA JJ3
Analisis sekuen lengkap (full length) cDNA JJ3 dilakukan melalui beberapa tahapan sebagai berikut. Deduksi asam amino sekuen lengkap cDNA JJ3 menggunakan program translasi (www.expasy.ch/tools/dna.html). Analisis tingkat homologi urutan basa nukleotida dan deduksi asam amino sekuen lengkap cDNA JJ3 dengan database gen di GenBank menggunakan program BLAST (Basic Local Aligment Search Tool) (www.ncbi.nlm.nih.gov/BLAST) dan program progam clustalW (http://www.ebi.ac.uk/clustalW/index.html). Analisis kerangka translasi (ORF) atau sekuen pengkodean (CDS) dan domain terkonservasi (conserved region) menggunakan program ORF Finder (www.ncbi.nlm.nih.gov/gorf/gorf.html) dan program GeneScanW (http://genes.mit.edu/GENSCANS.html). Analisis kedekatan hubungan dan kekerabatan JJ3 dengan gen homolog dari berbagai tanaman menggunakan program phylodraw. Domain sekuen lengkap cDNA JJ3 ditentukan dengan program motif scan (www.expasy.ch/prosite). Analisis hidrofobisitas dilakukan menggunakan program BioEdit versi 7.0.0 (Hall 1999).
HASIL DAN PEMBAHASAN
Sekuen Lengkap (Full Length) JJ3
Sekuen lengkap JJ3 yang berukuran 841 bp terdiri atas 232 adenin (a), 234 sitosin (c), 192 guanin (g) dan 183 timin (t) seperti yang disajikan pada Gambar 24. Sekuen lengkap ini merupakan hasil penggabungan sekuen-sekuen parsial hasil pemanjangan 5’-RACE dan 3’-RACE. Dengan membandingkan sekuen lengkap tersebutt dengan data fragmen sebelum pemanjangan dengan RACE, maka terdapat penambahan sejumlah 485 basa nukleotida ke arah ujung 5’ dan sekitar 73 bp ke arah ujung 3’. Hasil translasi sekuen lengkap cDNA JJ3 menggunakan program translasi (www.expasy.ch/tools/dna.html) diperoleh asam amino sejumlah 284 asam amino dengan empat asam amino start codon (Met) dan dua stop codon (*). Start codon tersebut tidak berada persis di awal sekuen begitu juga stop codon tidak di akhir sekuen.
Gambar 24 (A). Sekuen lengkap basa nukleotida JJ3 hasil pemanjangan menggunakan metode PCR-RACE. (B). Asam amino hasil translasi dari sekuen lengkap basa nukleotida JJ3.
Analisis Struktur Sekuen Lengkap JJ3
Estimasi struktur sekuen lengkap JJ3 dilakukan dengan menggunakan program GeneScanW (http://genes.mit.edu/GENSCANS.html) dengan hasil seperti pada Gambar 25. Tampak bahwa sekuen lengkap JJ3 terdiri atas sekuen pengkode (coding sequence, CDS) lengkap yang merupakan ekson tunggal (single exon) dengan panjang 633 bp dimulai dari nukleotida nomor 68 sampai 700 dengan nilai probabilitas cukup tinggi (85%). Panjang poly-Adenin (polyA) adalah 6 bp dimulai dari nukleotida nomor 813 sampai 818. Sekuen full length ini tidak memiliki intron karena merupakan sekuen cDNA. Peptida yang dihasilkan CDS JJ3 tersusun atas 210 asam amino. Pada coding sequence tersebut terdapat domain terkonservasi (asam amino 78 sampai 210) dan domain tidak terkonservasi beragam yang terletak pada asam amino 1 sampai 77 seperti pada Gambar 25. A. BASE COUNT 232 a 234 c 192 g 183 t 1 gcaagcagtggtatcaacgcagagtggccattacggccggggaacacttgtattatctca 61 agcaaccatggcaatggcaacccaagcctctctcttaaccccacccctctccggtctcaa 121 agccagcgaccgcgcctccgtgccatggaagcaaaactccagcctctccttctccagccc 181 gaagcccctcaagttctccagaacaatcagagcagcagccgccgacgagaccacagaggc 241 accagcaaaagtagaggctgcaccggtcgggttcaccccaccagaacttgacccaaacac 301 cccttccccgatcttcgggggcagcaccggcgggctcctgcgcaaggcacaggtggagga 361 gttttatgtcattacgtgggactcacccaaagaacagatctttgaaatgcccactggcgg 421 cgccgctatcatgagggagggtcctaaccttctcaagttggccaggaaggagcagtgctt 481 ggctcttgggactaggctcaggtccaagtacaagatcaagtaccagttctacagggtctt 541 ccctaatggggaggttcagtatttgcaccctaaggatggtgtttaccctgagaaggtcaa 601 cgccggacgccaaggggtgggtcaaaacttcaggtctattggtaagaatgttagtcctat 661 tgaggtcaagttcactggcaagcagccctatgatttgtgagcacacaactctatcttcat 721 catcatcatcccccgtgcttcctttatatgctatatattctcatgtgatatcatgtacct 781 attgtcaattttattatgccacaaatattgctaaaaaaaaaaaaaaaaaaaaaaaaaaaa 841 a B. 284 asam amino X X X X Q A V V S T Q S G H Y G R G T L V L S Q A T M A M A T Q A S L L T P P L S G L K A S D R A S V P W K Q N S S L S F S S P K P L K F S R T I R A A A A D E T T E A P A K V E A A P V G F T P P E L D P N T P S P I F G G S T G G L L R K A Q V E E F Y V I T W D S P K E Q I F E M P T G G A A I M R E G P N L L K L A R K E Q C L A L G T R L R S K Y K I K Y Q F Y R V F P N G E V Q Y L H P K D G V Y P E K V N A G R Q G V G Q N F R S I G K N V S P I E V K F T G K Q P Y D L * A H N S I F I I I I P R A S F I C Y I F S C D I M Y L L S I L L C H K Y C * K K K K K K K K K
Gambar 25 (A) Estimasi struktur sekuen lengkap JJ3, (B) Prediksi sekuen pengkodean, CDS lengkap, dan (C) prediksi sekuen peptida, (D) domain terkonservasi (conserved domain) berwarna merah dan yang tidak terkonservasi (domain beragam) berwarna hitam.
Untuk mengetahui fungsi dari sekuen lengkap JJ3 yang diperoleh maka perlu dilakukan analisis bagian yang bisa dibaca dalam proses translasi (open reading frame, ORF) atau sekuen pengkodean (Coding sequence, CDS) yang merupakan bagian yang ditranslasikan membentuk protein fungsional. Hasil analisis menggunakan program ORF Finder (www.ncbi.nlm.nih.gov/gorf/gorf. html) menunjukkan bahwa terdapat sekuen pengkodean (coding sequence, CDS)
Hasil GENSCANW sekuen cDNA JJ3 A. Prediksi struktur gen/exon:
Gn.Ex Type S .Begin ...End .Len Fr Ph I/Ac Do/T CodRg P.... Tscr.. --- ---- - --- --- ---- -- -- ---- ---- --- --- --- 1.01 Sngl + 68 700 633 1 0 106 53 397 0.848 33.89 1.02 PlyA + 813 818 6 -0.45 B. Prediksi coding sequence, CDS: 633 bp
atggcaatggcaacccaagcctctctcttaaccccacccctctccggtctcaaagccagcga ccgcgcctccgtgccatggaagcaaaactccagcctctccttctccagcccgaagcccctca agttctccagaacaatcagagcagcagccgccgacgagaccacagaggcaccagcaaaagta gaggctgcaccggtcgggttcaccccaccagaacttgacccaaacaccccttccccgatctt cgggggcagcaccggcgggctcctgcgcaaggcacaggtggaggagttttatgtcattacgt gggactcacccaaagaacagatctttgaaatgcccactggcggcgccgctatcatgagggag ggtcctaaccttctcaagttggccaggaaggagcagtgcttggctcttgggactaggctcag gtccaagtacaagatcaagtaccagttctacagggtcttccctaatggggaggttcagtatt tgcaccctaaggatggtgtttaccctgagaaggtcaacgccggacgccaaggggtgggtcaa aacttcaggtctattggtaagaatgttagtcctattgaggtcaagttcactggcaagcagcc ctatgatttgtga
C. Prediksi sekuen peptida: 210 asam amino
MAMATQASLLTPPLSGLKASDRASVPWKQNSSLSFSSPKPLKFSRTIRAAAADETTEAPAKV EAAPVGFTPPELDPNTPSPIFGGSTGGLLRKAQVEEFYVITWDSPKEQIFEMPTGGAAIMRE GPNLLKLARKEQCLALGTRLRSKYKIKYQFYRVFPNGEVQYLHPKDGVYPEKVNAGRQGVGQ NFRSIGKNVSPIEVKFTGKQPYDL
atau open reading frame (ORF) dan domain terkonservasi (putative conserved domain) sekuen lengkap JJ3 seperti pada Gambar 26.
Gambar 26 Open reading frame (ORF) atau coding sequence dari sekuen lengkap JJ3. Sekuen yang ditulis dengan huruf kecil merupakan CDS JJ3 dan huruf kapital merupakan deduksi asam amino.
Hasil analisis menunjukkan bahwa salah satu segmen dari sekuen lengkap JJ3 memiliki salah satu daerah pengkodean (coding region) lengkap yang merupakan kerangka translasi atau open reading frame (ORF) lengkap dengan coding sequence (CDS) yang menghasilkan protein PsaD photosystem I (PS-I) subunit, protein ekstrinsik yang terletak di bagian sisi stroma pada membran
Gcaagcagtggtatcaacgcagagtggccattacggccggggaacacttgtattatctcaa gcaacc 68 atggcaatggcaacccaagcctctctcttaaccccacccctctcc M A M A T Q A S L L T P P L S 113 ggtctcaaagccagcgaccgcgcctccgtgccatggaagcaaaac G L K A S D R A S V P W K Q N 158 tccagcctctccttctccagcccgaagcccctcaagttctccaga S S L S F S S P K P L K F S R 203 acaatcagagcagcagccgccgacgagaccacagaggcaccagca T I R A A A A D E T T E A P A 248 aaagtagaggctgcaccggtcgggttcaccccaccagaacttgac K V E A A P V G F T P P E L D 293 ccaaacaccccttccccgatcttcgggggcagcaccggcgggctc P N T P S P I F G G S T G G L 338 ctgcgcaaggcacaggtggaggagttttatgtcattacgtgggac L R K A Q V E E F Y V I T W D 383 tcacccaaagaacagatctttgaaatgcccactggcggcgccgct S P K E Q I F E M P T G G A A 428 atcatgagggagggtcctaaccttctcaagttggccaggaaggag I M R E G P N L L K L A R K E 473 cagtgcttggctcttgggactaggctcaggtccaagtacaagatc Q C L A L G T R L R S K Y K I 518 aagtaccagttctacagggtcttccctaatggggaggttcagtat K Y Q F Y R V F P N G E V Q Y 563 ttgcaccctaaggatggtgtttaccctgagaaggtcaacgccgga L H P K D G V Y P E K V N A G 608 cgccaaggggtgggtcaaaacttcaggtctattggtaagaatgtt R Q G V G Q N F R S I G K N V 653 agtcctattgaggtcaagttcactggcaagcagccctatgatttg S P I E V K F T G K Q P Y D L 698 tga 700 * gcacacaactctatcttcatcatcatcatcccccgtgcttcctttatatgctatatattct catgtgatatcatgtacctattgtcaattttattatgccacaaatattgctaaaaaaaaaa aaaaaaaaaaaaaaaaaaa
tilakoid kloroplas. Protein PsaD tersebut tersusun atas 210 asam amino yang diawali dengan start codon atg (Met) dan diakhiri dengan stop codon tga (Gambar 26). PsaD merupakan protein ekstrinsik PSI subunit yang terletak pada sisi stromal pada membran tilakoid (Xia et al. 1998), fungsinya terlibat dalam transpor elektron dalam transduksi signal cahaya. Analisis fungsi dari protein PsaD menunjukkan bahwa protein berberat molekul rendah tersebut terkait dengan transport elektron dalam fotosintesis.
Protein PsaD yang dikode JJ3 tersebut terletak di bagian luar (periferal) sisi stroma membran tilakoid pada fotosistem I (PSI). Hasil kajian bioinformatika tentang struktur tiga dimensi protein PsaD menunjukkan bahwa PsaD terkait dengan dengan tiga subunit lain (PsaC dan PsaE) dalam menstabilkan docking feredoksin yang berperan penting dalam transport elektron selama proses fotosintesis berlangsung (Klukas et al. 1999). Lagoutte et al (2001) dalam penelitiannya menggunakan cyanobacterium Synechocystis sp PCC 6803, melaporkan bahwa subunit PsaD fotosistem I (PSI) mempunyai peranan penting di dalam stabilitas kompleks PSI dan merupakan bagian dari tempat docking ferredoxin (Fd). Fotosistem I (PSI) merupakan kompleks multisubunit yang menempel pada membran tilakoid yang mampu memfoto-induksikan transfer elektron menjadi akseptor elektron yaitu ferredoxin (Fd) yang bersifat dapat larut. PSI terdiri atas 11 subunit, baik yang bersifat integral seperti yang mengikat sentral kofaktor menjadi transfer elektron, maupun yang bersifat periferal seperti PsaD subunits.
Lagoutte et al. (2001) menduga bahwa subunit PsaD periferal mempunyai beberapa fungsi yang berbeda pada kompleks PSI. yang pertama berperan penting secara struktural dalam melengkapi kompleks PSI menjadi subunit yang dikode inti pertama dari kompleks yang disintesa dengan iluminasi atau pencahayaan. Polipeptida ini berinteraksi dengan banyak subunit berbeda. Fungsi secara struktural dari PsaD adalah mendukung sekitar 70% dari pemanjangan dan pelebaran rantai polipeptida. Juga terdapat sekuen dasar yang kuat pada bagian tengah dari PsaD yang menjaga stabilitas hubungan PsaD dengan core PSI. Fungsi yang lain dari PsaD adalah memberikan sekuen spesifik untuk mengarahkan dan menempatkan (guiding and docking) elektrostatik feredoksin (Fd) pada PSI.
Menurut Jiao et al. (2004), protein fotosistem tipe I (PSI) seperti PsaD juga berfungsi sebagai oksidoreduktase dari plastocyanin : ferredoxin yang mengkatalisis transfer elektron dari plastocyanin terreduksi ke ferredoxin teroksidasi di dalam rantai transport elektron pada membran tilakoid. PSI menghasilkan ATP dan NADPH dan juga berfungsi untuk meregulasi metabolisme karbon dan nitrogen di dalam kloroplas. Selanjutnya ekspresi gen-gen fotosintesis di dalam inti maupun kloroplas berikut prosesing subunit protein untuk pembentukan kompleks protein (PSI, PSII) diaktivasi oleh cahaya. (Biswal 1997; Tyagi et al. 2000).
Analisis Homologi Sekuen Lengkap JJ3
Analisis homologi basa nukleotida atau asam amino sekuen lengkap cDNA JJ3 dengan gen psaD tanaman lain yang tersimpan di database DNA di GenBank dilakukan dengan menggunakan program BLAST (Basic Local Aligment Search Tool) melalui situs (www.ncbi.nlm.nih. gov/BLAST2). Hasil program BLAST ditampilkan dalam bentuk skoring dari yang paling tinggi sampai yang paling rendah, disertai dengan persentase identitas JJ3 dengan gen psaD tanaman lain. Semakin tinggi skor dan persentase identitas maka tingkat homologi sekuen lengkap JJ3 dengan gen psaD semakin tinggi. Dengan menggunakan Local aligment dapat diketahui daerah terkonservasi (conserved region) yang merupakan domain utama suatu gen. Program BLAST merupakan alat bioinformatika yang cukup penting untuk menentukan identitas suatu sekuen yang belum diketahui dengan menganalisis tingkat homologinya dengan gen lain di GenBank yang telah dikarakterisasi (Mount 2001).
Seperti yang dilaporkan sebelumnya (Khumaida 2002) bahwa fragmen JJ3 homolog dengan gen psaD pada fotosistem I subunit II. Oleh karena itu pensejajaran sekuen lengkap dilakukan terhadap sekuen DNA yang terkait dengan gen psaD dan memiliki skor BLAST di atas skor 100 sampai tertinggi. Hasil analisis pensejajaran menggunakan program ClustalW (http://www.ebi.ac.uk/ clustalW/index.html) (Tabel 16) mencerminkan tingkat kesamaan seluruh basa nukleotida atau asam amino dari sekuen lengkap JJ3 yang diperoleh termasuk bagian yang berada diluar sekuen pengkodean (coding sequence) atau daerah
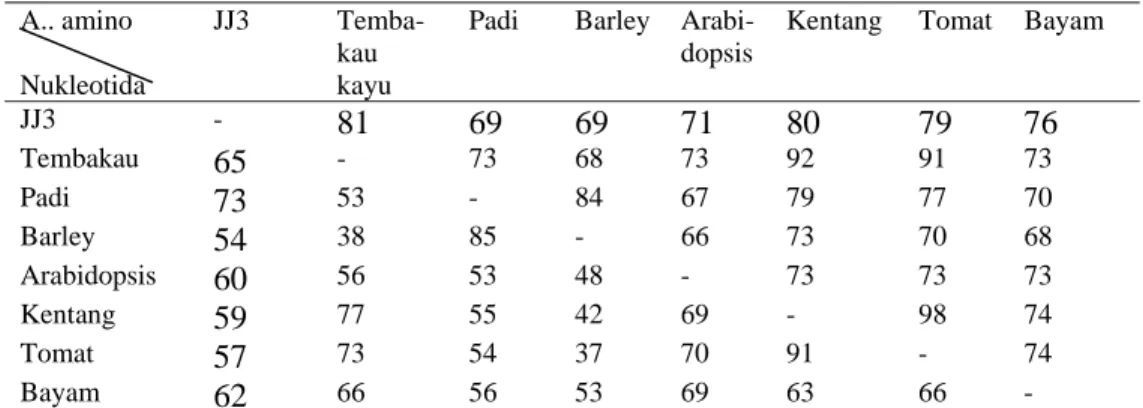
terkonservasi (conserved region). Diperoleh bahwa sekuen lengkap JJ3 memiliki tingkat homologi yang cukup tinggi baik berdasarkan homologi sekuen basa nukleotida maupun homologi sekuen asam amino. Tingkat homologi tertinggi berdasarkan sekuen lengkap basa nukleotida ditunjukkan pada tanaman padi (Oryza sativa) (XM48378) (73) kemudian beberapa tanaman lain seperti pada tembakau kayu (Nicotiana sylvestris) (Q70PN9) (65%), bayam dan Arabidopsis. Tingkat homologi sekuen lengkap asam amino JJ3 tertinggi (81%) dengan tembakau kayu (Nicotiana sylvestris) (P29302) dan kentang (Solanum tuberosum) (Q70PN9) kemudian tomat dan bayam.
Tabel 16 Matriks tingkat homologi (%) sekuen lengkap basa nukleotida dan asam amino JJ3 dengan tanaman lain
A.. amino
Nukleotida
JJ3 Temba- kau
kayu
Padi Barley Arabi-dopsis
Kentang Tomat Bayam
JJ3 - 81 69 69 71 80 79 76 Tembakau 65 - 73 68 73 92 91 73 Padi 73 53 - 84 67 79 77 70 Barley 54 38 85 - 66 73 70 68 Arabidopsis 60 56 53 48 - 73 73 73 Kentang 59 77 55 42 69 - 98 74 Tomat 57 73 54 37 70 91 - 74 Bayam 62 66 56 53 69 63 66 - Keterangan: Persentase di bagian bawah diagonal adalah tingkat kesamaan basa nulkeotida sekuen
lengkap JJ3, dan bagian atas diagonal adalah tingkat kesamaan berdasarkan asam amino.
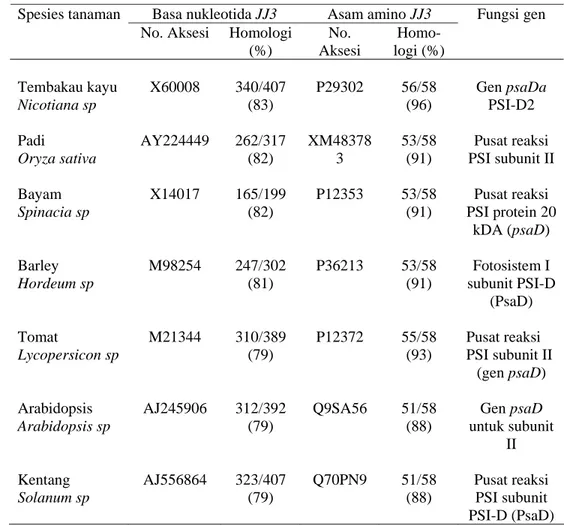
Hasil pensejajaran menggunakan BLAST terhadap bagian terkonservasi (conserved domain), tingkat homologi cDNA JJ3 dengan tanaman lain baik berdasarkan sekuen DNA atau asam amino menunjukkan tingkat homologi yang lebih tinggi dibanding hasil pensejajaran berdasarkan sekuen lengkap (Tabel 17). Tingkat homologi tertinggi cDNA JJ3 adalah dengan gen psaD pada tanaman tembakau kayu (Nicotiana sylvestris) (X60008) yaitu 83% berdasarkan sekuen nukleotida dan dengan tanaman yang sama tembakau kayu (Nicotiana sylvestris) (P29302) yaitu 96% berdasarkan sekuen asam amino.
Tabel 17 Tingkat homologi cDNA JJ3 dengan gen psaD beberapa spesies tanaman lain menggunakan domain terkonservasi sekuen basa nukleotida dan asam amino
Basa nukleotida JJ3 Asam amino JJ3
Spesies tanaman
No. Aksesi Homologi (%) No. Aksesi Homo-logi (%) Fungsi gen Tembakau kayu Nicotiana sp X60008 340/407 (83) P29302 56/58 (96) Gen psaDa PSI-D2 Padi Oryza sativa AY224449 262/317 (82) XM48378 3 53/58 (91) Pusat reaksi PSI subunit II Bayam Spinacia sp X14017 165/199 (82) P12353 53/58 (91) Pusat reaksi PSI protein 20 kDA (psaD) Barley Hordeum sp M98254 247/302 (81) P36213 53/58 (91) Fotosistem I subunit PSI-D (PsaD) Tomat Lycopersicon sp M21344 310/389 (79) P12372 55/58 (93) Pusat reaksi PSI subunit II (gen psaD) Arabidopsis Arabidopsis sp AJ245906 312/392 (79) Q9SA56 51/58 (88) Gen psaD untuk subunit II Kentang Solanum sp AJ556864 323/407 (79) Q70PN9 51/58 (88) Pusat reaksi PSI subunit PSI-D (PsaD)
Hasil pensejajaran ganda atau multiple aligment menggunakan program clustalW (http://www.ebi.ac.uk/clustalW/index.html) menunjukkan bahwa cDNA JJ3 memiliki elemen konsensus yang cukup tinggi baik berdasarkan basa nukleotida maupun asam amino seperti disajikan pada Gambar 27 dan 28. Elemen konsensus atau daerah terkonservasi (consensus element atau conserved region) merupakan bagian, elemen atau sekuen basa nukleotida atau asam amino yang sama atau paling sering muncul pada bagian fungsional suatu gen tertentu (Khal 2001). Adanya elemen yang tidak terkonservasi menunjukkan bahwa pada organisme tersebut telah terjadi mutasi dalam bentuk penambahan, pengurangan, atau pertukaran elemen.
Gambar 27 Sebagian hasil pensejajaran (multi-alignment) sekuen basa nukleotida cDNA JJ3 dengan spesies tanaman lain menggunakan progran clustalW. Tanda aktris (*) menunjukkan nukleotida terkonservasi.
Hasil pensejajaran ganda (multiple aligment) sekuen asam amino cDNA JJ3 dengan tanaman lain, menunjukkan bagian konsensus atau conserved region yang lebih tinggi dari pada hasil pensejajaran sekuen basa nukleotida (Gambar 27). Hal ini karena beberapa kodon atau triplet yang memiliki basa nukleotida berbeda dapat membentuk asam amino yang sama, misalnya valin (V) oleh triplet GTT, GTC, GTG atau GTA (Dale dan von Schantz 2002).
Hasil ini juga membuktikan bahwa JJ3 yang diisolasi dari kedelai toleran merupakan cDNA yang homolog dengan gen psaD yang merupakan conserved domain mengkode protein PsaD PSI subunit. Meskipun demikian, ada beberapa asam amino cDNA JJ3 yang berbeda, misalnya dengan tembakau kayu seperti arginin (R) yang menggantikan posisi glutamin (Q) atau alanin (A) yang menggantikan prolin (P) pada padi dan barley. Adanya perbedaan ini diduga karena adanya mutasi.
JJ3 AACACCCCTTCCCCGATCTTCGGGGGCAGCACCGGCGGGCTCCTGCGCAAGGCACAGGTG 355 Tembakau AACACACCTTCCCCAATCTTCGGTGGCAGCACCGGTGGGCTTCTCCGCAAGGCCCAAGTT 811 Padi AACACGCCGTCCCCGATCTTCGGCGGGAGCACGGGGGGACTCCTCCGGAAGGCGCAGGTG 267 Bayam ACACTCCCTCCCCCATCTTTGCTGGAAGCACAGGTGGGCTATTGAGGAAGGCACAAGTAA 354 Barley TCCACGCCGTCCCCGATCTTCGGCGGCAGCACCGGCGGGCTGCTCCGCAAGGCCCAGGTC 312 Tomat AACACACCCTCCCCAATCTTCGGTGGCAGCACCGGTGGGCTTCTTCGCAAAGCCCAAGTG 323 Arabidopsis AACACACCGTCTCCGATCTTCGCTGGAAGCACCGGTGGTCTTCTACGTAAAGCGCAAGTG 282 kentang AACACACCTTCCCCAATCTTCGGTGGCAGCACTGGTGGGCTTCTTCGCAAAGCCCAAGTA 316 *** ** ** ** ***** * ** ***** ** ** ** * * ** ** ** ** JJ3 GAGGAGTTTTATGTCATTACGTGGGACTCACCCAAAGAACAGATCTTTGAAATGCCCACT 415 Tembakau GAGGAGTTTTACGTAATTACTTGGGAATCACCTAAAGAACAGATCTTTGAGATGCCAACT 871 Padi GAGGAGTTCTACGTCATCACATGGACGTCGCCCAAGGAGCAGGTGTTCGAGATGCCCACG 327 Bayam GAGGAGTTTTACGTGATAACATGGGAATCACCAAAAGAGCAAATATTCGAGATGCCAACA 414 Barley GAGGAGTTTTACGTCATCACCTGGACCTCCCCCAAGGAGCAGGTCTTCGAGATGCCCACC 372 Tomat GAAGAATTCTACGTCATCACATGGGAATCACCAAAGGAACAGATCTTTGAGATGCCAACA 383 Arabidopsis GAAGAGTTCTACGTTATCACGTGGAACTCACCGAAAGAACAGATCTTTGAGATGCCGACA 342 kentang GAAGAATTCTACGTCATCACATGGGAATCACCAAAGGAACAGATCTTCGAGATGCCAACA 376 ** ** ** ** ** ** ** *** ** ** ** ** ** * ** ** ***** ** JJ3 GGCGGCGCCGCTATCATGAGGGAGGGTCCTAACCTTCTCAAGTTGGCCAGGAAGGAGCAG 475 Tembakau GGTGGTGCAGCTATTATGAGGGAAGGTGCTAATTTGCTGAAATTGGCGAGGAAAGAGCAG 931 Padi GGCGGCGCCGCCATCATGCGCGAGGGCCCCAACCTGCTGAAGCTGGCCAGGAAGGAGCAG 387 Bayam GGAGGAGCAGCAATAATGAGGGAAGGACCAAACTTGCTAAAATTAGCACGTAAAGAGCAA 474 Barley GGCGGCGCCGCCATCATGCGCGAGGGCCCCAACCTCCTCAAGCTCGCCCGCAAGGAGCAG 432 Tomat GGTGGTGCTGCTATAATGAGACAAGGACCAAATTTGTTGAAATTGGCAAGGAAAGAACAG 443 Arabidopsis GGAGGAGCAGCGATCATGAGAGAAGGTCCGAATCTTCTGAAGCTAGCGAGGAAAGAGCAG 402 kentang GGTGGTGCTGCTATAATGAGAGAAGGACCCAATTTGTTGAAATTGGCCAGGAAAGAACAG 436 ** ** ** ** ** *** * * ** * ** * * ** * ** * ** ** **
Gambar 28 Hasil pensejajaran sekuen lengkap asam amino cDNA JJ3 dengan tanaman tingkat tinggi lain. Tanda aktris (*) menunjukkan daerah konsensus, (:) menunjukkan bagian yang di dalamnya mempunyai ukuran dan hidrofobisitas yang sama, dan (.) menunjukkan bagian atau kolom dimana ukuran atau hidrofobisitasnya tidak pernah berubah (preserved) selama proses evolusi.
Pada Gambar 28 selain dapat diketahui daerah terkonservasi dan daerah beragam, juga dapat diketahui posisi yang memiliki profil hidrofobisitas yang tidak pernah berubah selama proses evolusi atau memiliki profil yang sama.
Berdasarkan hasil pensejajaran di atas dapat dinyatakan bahwa JJ3 yang diisolasi dari kedelai toleran naungan memiliki tingkat kesamaan nukleotida yang cukup tinggi (sekitar 80%) begitu juga dengan sekuen asam amino (sekitar 95%) dengan gen psaD pada tanaman tembakau kayu (Nicotiana sylvestris), kentang (Solanum tuberosum), tomat (Lycopersicon esculentum), bayam (Spinacia
JJ3 MAMATQAS-LLTP-PLSGLKASDR-ASVPWKQNSSLSFSSPKPLKFSRTIRAAAADETTE 57 Nicotiana MAMATQAS-LFTP-ALSAPKS---SAPWKQ-SLASFS-PKQLKSTVSAPRPIRAMAEE 51 Oryza MAMATQAS-AAKCHLL-AAWAP---AKPRSSTLSMPTS-RAPTSLRAAAED-QPA 48 Spinacia MAMATQAT-LFSPSSLSSAKPIDTRLTTSFKQPSAVTFA-SKPASRHHSIRAAAAAEGKA 58 Hordeum MAMATQAS-AATRHLITAAWSPS---AKPRPATLAMPSSARGPAPLFAAAPD-TPA 51 Lycopersicon MAMATQAS-LFTP-PLSVPKST----TAPWKQ-SLVSFSTPKQLKSTVSVTRPIRAMAEE 53 Arabidopsis --MATQAAGIFSP-AITTTTS---AVKKLHLFSSSHRPKSLSFTKTAIRAEKTESSS 51 Solanum MAMATQAS-LFTP-ALSVPKS---TAPWKQ-SLLSFSTPKQLKSTVSVTRPIRAMAEE 52 *****: . : . : .. . . JJ3 AP--AKVEAAPVGFTPPELDPNTPSPIFGGSTGGLLRKAQVEEFYVITWDSPKEQIFEMP 115 Nicotiana A--ATKEAEAPVGFTPPQLDPNTPSPIFGGSTGGLLRKAQVEEFYVITWESPKEQIFEMP 109 Oryza AAATEEKKPAPAGFVPPQLDPNTPSPIFGGSTGGLLRKAQVEEFYVITWTSPKEQVFEMP 108 Spinacia AA-ATETKEAPKGFTPPELDPNTPSPIFAGSTGGLLRKAQVEEFYVITWESPKEQIFEMP 117 Hordeum PAAPP-AEPAPAGFVPPQLDPSTPSPIFGGSTGGLLRKAQVEEFYVITWTSPKEQVFEMP 110 Lycopersicon APAATEEKPAPAGFTPPQLDPNTPSPIFGGSTGGLLRKAQVEEFYVITWESPKEQIFEMP 113 Arabidopsis AA--PAVKEAPVGFTPPQLDPNTPSPIFAGSTGGLLRKAQVEEFYVITWNSPKEQIFEMP 109 Solanum APAATEEKPAPAGFTPPQLDPNTPSPIFGGSTGGLLRKAQVEEFYVITWESPKEQIFEMP 112 ** ** **:***.******.******************** *****:**** JJ3 TGGAAIMREGPNLLKLARKEQCLALGTRLRSKYKIKYQFYRVFPNGEVQYLHPKDGVYPE 175 Nicotiana TGGAAIMREGANLLKLARKEQCLALGTRLRSKYKINYRFYRVFPNGEVQYLHPKDGVYPE 169 Oryza TGGAAIMREGPNLLKLARKEQCLALGTRLRSKYKINYQFYRVFPNGEVQYLHPKDGVYPE 168 Spinacia TGGAAIMREGPNLLKLARKEQCLALGTRLRSKYKIKYQFYRVFPSGEVQYLHPKDGVYPE 177 Hordeum TGGAAIMREGPNLLKLARKEQCLALGNRLRSKYKIAYQFYRVFPNGEVQYLHPKDGVYPE 170 Lycopersicon TGGAAIMRQGPNLLKLARKEQCLALGTRLRSKYKINYQFYRVFPNGEVQYLHPKDGVYPE 173 Arabidopsis TGGAAIMREGPNLLKLARKEQCLALGTRLRSKYKITYQFYRVFPNGEVQYLHPKDGVYPE 169 Solanum TGGAAIMREGPNLLKLARKEQCLALGTRLRSKYKINYQFYRVFPNGEVQYLHPKDGVYPE 172 ********:*.***************.******** *:******.*************** JJ3 KVNAGRQGVGQNFRSIGKNVSPIEVKFTGKQPYDL 210 Nicotiana KVNAGRQGVGQNFRSIGKNKSPIEVKFTGKQVYDL 204 Oryza KVNAGRQGVGQNFRSIGKNVSPIEVKFTGKNVFDI 203 Spinacia KVNPGRQGVGLNMRSIGKNVSPIEVKFTGKQPYDL 212 Hordeum KVNAGRQGVGQNFRSIGKNVSPIEVKFTGKNSFDI 205 Lycopersicon KVNPGREGVGQNFRSIGKNKSAIEVKFTGKQVYDI 208 Arabidopsis KANPGREGVGLNMRSIGKNVSPIEVKFTGKQSYDL 204 Solanum KVNPGREGVGQNFRSIGKNKSAIEVKFTGKQVYDI 207 *.*.**:*** *:****** *.********: :*:
oleracea), Arabidopsis (Arabidopsis thaliana), barley (Hordeum vulgare), dan padi (Oryza sativa). Tingkat homologi ini lebih besar dari yang dilaporkan sebelumnya (Hiyama 1997). Tanaman-tanaman tersebut berpotensi untuk dikembangkan di daerah dengan kondisi lingkungan intensitas cahaya rendah karena memiliki gen psaD.
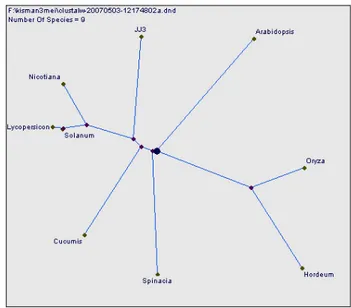
Hasil analisis pensejajaran di atas juga didukung oleh hasil analisis kekerabatan menggunakan urutan asam amino seperti pada Gambar 29. terlihat bahwa, cDNA JJ3 lebih dekat dengan gen psaD pada tanaman tembakau kayu, tanaman tomat, kentang, mentimun, bayam, Arabidopsis, padi dan barley.
Gambar 29 Filogenetik cDNA JJ3 dengan gen psaD pada beberapa tanaman berdasarkan urutan asam amino
Tingkat homologi dan kekerabatan yang tinggi mengindikasikan bahwa mekanisme adaptasi tanaman kedelai terhadap intensitas cahaya rendah berhubungan erat dengan efisiensi proses elektron transport pada reaksi cahaya yang terjadi di dalam membran tilakoid. Dengan kata lain, keberadaan cDNA JJ3 akan meningkatkan laju transpor elektron fotosintetik sehingga dalam kondisi kurang cahaya pun fiksasi energi dari fotosintesis berjalan dengan normal. Hal ini sejalan dengan yang dilaporkan oleh Muhuria (2007) bahwa laju transport elektron pada genotipe toleran naungan (Ceneng) lebih tinggi dibanding genotipe
positif antara struktur membran tilakoid dengan ekspresi gen lhcp pada kedelai yang tahan naungan. Beberapa genotipe tanaman yang toleran naungan telah dilaporkan antara lain pada tanaman padi gogo (Sopandie et al. 2001, 2003b, 2003c, 2006; Khumaida 2002) dan Arabidopsis (Carabelli et al. 1996; Franklin et al. 2003; Vandenbussche et al. 2005).
Dengan demikian dapat dinyatakan bahwa JJ3 merupakan kandidat gen fotosintetik inti, homolog dengan gen psaD yang bekerja di dalam pusat reaksi PSI subunit untuk mengkode protein PsaD fotosistem I subunit II. Menurut Hiyama (1997), gen psaD merupakan gen fotosintetik inti yang mengkode protein PsaD yang ditransfer ke kloroplas.
JJ3 telah terdaftar di database di GenBank dengan nomor aksesi EF628505 sebagai kandidat gen yang menyandi PsaD pada tanaman kedelai (Lampiran 5).
KESIMPULAN
1. Sekuen lengkap cDNA JJ3, kandidat gen yang terkait dengan mekanisme adaptasi kedelai terhadap intensitas cahaya rendah, memiliki coding sequence (CDS) 633 bp dengan 210 asam amino deduksi tersusun atas 136 asam amino terkonservasi dan 77 asam amino beragam.
2. Sekuen CDS JJ3 homolog dengan PsaD PSI subunit, fungsinya terkait dengan transport elektron fotosintesis pada pusat reaksi PSI.
3. cDNA JJ3 homolog dengan gen psaD pada tanaman tembakau liar, padi, barley, Arabidopsis, kentang, tomat, dan bayam.