L
Journal of Experimental Marine Biology and Ecology 246 (2000) 125–138
www.elsevier.nl / locate / jembe
Experimental evidence of predation by juvenile flounder,
Platichthys flesus, on a shallow water meiobenthic
community
* Katri Aarnio
˚ ˚
¨
Huso Biological Station, Abo Akademi University, FIN-22220 Emkarby, Aland Islands, Finland
Received 15 October 1999; received in revised form 22 October 1999; accepted 1 December 1999
Abstract
In the northern Baltic Sea juvenile flounder, Platichthys flesus, occur in high abundances on shallow sandy bottoms in late summer and autumn. They feed mainly on meiofauna and the ontogenetic switch to macrofaunal sized food occurs at a larger size here than in other areas, exemplifying the high relative importance of meiofauna. Consequently, juvenile P. flesus in the Baltic feed for a longer period on meiofauna, and could thus be expected to have a stronger predation effect on the meiofaunal assemblages. In this study the predation effects of juvenile P.
flesus on meiofaunal abundances and community structure were studied using microcosms that
were sampled repeatedly over a 3-week period. Significant differences between treatment and control were found for the total number of taxa, for abundances of harpacticoids, copepod nauplii and ostracods. The nematode community was not affected, but one genus, Axonolaimus, was negatively affected by predation. The predation affected meiofaunal community structure as the major taxon diversity was significantly reduced. The results suggest that the meiofauna on shallow sandy bottoms may be structured by juvenile P. flesus, and the combined predation pressure of juvenile flounder and other epibenthic predators in the area might be considerable. Microcosms are effective in testing natural predation, especially on meiofaunal communities, but field experiments should be conducted to account for the physical characteristics of the area studied. 2000 Elsevier Science B.V. All rights reserved.
Keywords: Experiment; Juvenile fish; Meiofauna; Platichthys flesus; Predation
1. Introduction
Shallow sandy bottoms are important feeding and nursery grounds for juvenile and
*Tel: 1358-18-37221; fax: 1358-18-37244. E-mail address: [email protected] (K. Aarnio)
small fish. In the northern Baltic Sea these areas are mainly utilized by gobies (Pomatoschistus sp) and juvenile flatfish (Platichthys flesus (L.), Scophthalmus maximus L.) (Nellbring, 1985; Jansson et al., 1985; Aarnio and Bonsdorff, 1993; Aarnio et al., 1996), where they may reach high abundances in summer and autumn when the recruitment of juveniles reaches a peak. Small and juvenile fish utilize primarily meiofauna as their food (Sogard, 1984; Fitzhugh and Fleeger, 1985; Tipton and Bell, 1988; Nelson and Coull, 1989; Coull et al., 1995). In the northern Baltic juvenile flounders have a variable diet, with food items from both meio- and macrofaunal groups. Meiofauna dominate the diet numerically by more than 90% for flounders less than 40 mm in size, and the most preferred food item is harpacticoid copepods, which constitute up to 85% of the diet (Aarnio et al., 1996). Effects of epibenthic predation on meiofaunal communities have been studied in both laboratory and field conditions, and usually no or only small effects of the predators on the meiofaunal abundance or community structure have been found (e.g. Reise, 1979; Alheit and Scheibel, 1982; Gee, 1987; St. John et al., 1989; Mattila et al., 1990; Webb and Parsons, 1991; Service et al., 1992). Other factors, such as predatory meiofauna, macroinfaunal activities or physical properties of the area studied, may be more important in structuring meiobenthic assemblages (Reise, 1979; Olafsson et al., 1990; Aarnio et al., 1998). It has also been suggested that rapid turnover rates and short generation times of the meiofauna may compensate for the predation effects (Gee et al., 1985; Coull, 1990).
In the northern Baltic Sea meiofauna is numerically more important than macrofauna as compared to fully marine areas (Elmgren et al., 1984). The ontogenetic shift of fish feeding from meio- to macrofauna sized organisms seems to occur at a larger size here than has been reported for other areas. In the Baltic the fish started to consume macrofauna at a size of 45 mm (Aarnio et al., 1996) compared to 30 mm in other areas (Hicks, 1984; Gee, 1987; Shaw and Jenkins, 1992). Consequently, juvenile P. flesus in the Baltic feed for a longer period on meiofauna, and could thus be expected to have a stronger predation effect on the meiofaunal assemblage. Further, the predation by P. flesus may be selective for harpacticoid copepods because these usually dominate the stomach contents, in contrast to the nematodes which dominate meiofaunal assemblages in the sediments but are rarely found in the guts (see Gee, 1989 for review). On the other hand, a fast digestion rate of the nematodes could be the reason why these are missing in the stomachs (Scholz et al., 1991).
The aim of this study was to investigate if predation by juvenile P. flesus may affect meiofaunal abundances and the structure of meiobenthic assemblages. This was studied by conducting an experiment using microcosms, which were sampled repeatedly over a 3-week period to determine the time scale over which possible effects became detectable. The following 0-hypotheses were tested:
1. predation by P. flesus does not affect meiofaunal abundances,
2. predation effects of P. flesus are not selective for any particular taxon, i.e. all prey are equally important for the flounders, and
2. Material and methods
The microcosm experiment was conducted in 1996 (September 22–October 14) by
2
using rectangular glass aquaria (11 l, surface area: 609 cm ) containing sediment taken
˚
from a shallow (1 m) sand bottom bay in the northwestern Aland archipelago, northern Baltic Sea (608179N, 198499E). The sediment had a medium grain size of 0.125–0.25
mm and an organic content of 0.5%. The area is exposed and frequently utilized by juvenile flounders especially in the autumn. The sampling was done with an
Ekman-2
Birge grab sampler (289 cm ) and the sediment was sieved through a 0.5 mm sieve in order to exclude the macrofauna. The sediment was stirred completely and allowed to settle after which a 3 cm thick layer was put in each aquarium. Filtered (0.063 mm) sea water was added to the aquaria (depth of overlying water: 15 cm), and they were equipped with an aerating system. The aquaria were left undisturbed for 3 days to allow the faunal assemblage in the sediment to stabilise before the experiment was started. To determine the initial faunal density in the microcosms five core samples ([1.0 cm) were
taken from each aquarium at the start of the experiment. The five samples from each microcosm were pooled and treated as one sample. As the core diameter was relatively small this procedure ensured that as many species / taxa as possible were included in the sample. Three juvenile flounders (length: 25–34 mm, weight: 115–330 mg) were added
2
to five randomly selected microcosms (corresp. to 49 ind / m ), while the other five served as controls and did not contain any fish. The natural density of flounders in this
2
area has been estimated to be about 1.5 ind / m (Aarnio et al., 1996), but much higher abundances have been detected in patches. The flounders were caught with a push net at 0.5–1 m depth 4 days prior to the experiment and kept in flow-through aquaria. They were starved for 24 h before they were put in the experimental aquaria. After 3, 7, 14 and 21 days of the start of the experiment five core samples were taken from each microcosm and treated in the same way as the initial samples. All samples were preserved in 4% formalin and dyed with Rose Bengal. Later they were sieved through 0.5, 0.25, 0.125 and 0.063 mm screens, and analysed under a stereomicroscope. The fauna were identified to species or taxa, and counted. A minimum of 100 nematodes were picked out from the samples taken from three randomly-chosen aquaria at the start and from three aquaria of both control and fish treatment after 3 weeks of experiment. They were transferred to glycerine, mounted on slides and identified to genus according to Platt and Warwick (1983). The hydrographical conditions (t8C, pH, O , salinity) were
2
monitored in every microcosm at the start of the experiment and every time core samples were taken, and daily in two randomly chosen aquaria of both control and fish treatment. Light conditions were standardized to 12 h dark:12 h light.
3. Numerical and statistical analyses
t-test (unpaired, 2 tail), after the data was checked for normality and homogeneity of variances. Log (x11) transformations were done where necessary to fulfill the
10
requirements for Student’s t-test. In the cases where transformation was unsuccessful the non-parametric Mann–Whitney U-test was used to compare the treatments (Sokal and Rohlf, 1995). To detect any faunal changes over time in the microcosms a comparison between start (day 0) and end (3 wk) of experiment was done for the control and fish treatment separately using paired Student’s t-test. Where a parametric test could not be used, the non-parametric Wilcoxon’s signed-ranks test was performed instead (Sokal and Rohlf, 1995). To detect any structuring effects of the predators on the meiofaunal assemblages, the Shannon-Wiener diversity index, H9, and its evenness component, J,
were calculated on taxon level for both the control and fish treatment at each date of sampling (Shannon and Waever, 1963). Comparisons between treatments at each date and comparisons for both treatments over time (start vs. end) were done using unpaired and paired Student’s t-test, respectively. For the nematode assemblage, the proportion of different nematode genera was compared between the control and fish treatment at the end of the experiment using unpaired Student’s t-test after an arcsin-transformation of the proportions were done (Sokal and Rohlf, 1995). Nematode diversity (H9) and
evenness (J ) in the end of the experiment was compared between the control and fish treatment using unpaired Student’s t-test.
4. Results
The hydrographical conditions in the aquaria were stable during the experimental time. The temperature was on average 128C, the salinity around 6, pH around 8 and
oxygen 95%. These figures are representative in autumn for the area from which the sediment and fish were sampled. At the end of the experiment the fish were killed and their stomach and gut contents analysed for prey items. The most frequently found prey were harpacticoids which constituted 78% of the stomach contents, while other prey items found were Nematoda (16%), Ostracoda (2%), Copepoda nauplii (2%) and Chironomidae (2%). One flounder had an empty stomach and gut when removed from the aquaria.
4.1. Initial fauna in the microcosms
When starting the experiment the total meiofaunal abundance (mean6S.E.) was 2
1143694 and 1132658 ind / 10 cm in the control and fish treatment, respectively. The
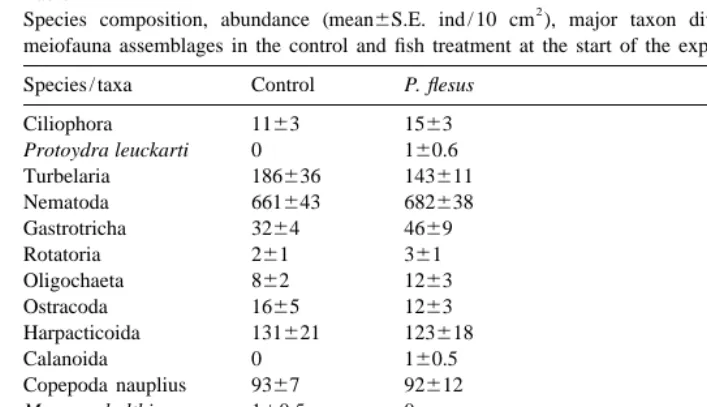
Table 1
2
Species composition, abundance (mean6S.E. ind / 10 cm ), major taxon diversity and evenness of the meiofauna assemblages in the control and fish treatment at the start of the experiment
Species / taxa Control P. flesus
Ciliophora 1163 1563
Protoydra leuckarti 0 160.6
Turbelaria 186636 143611
Nematoda 661643 682638
Gastrotricha 3264 4669
Rotatoria 261 361
Oligochaeta 862 1263
Ostracoda 1665 1263
Harpacticoida 131621 123618
Calanoida 0 160.5
Copepoda nauplius 9367 92612 Macoma balthica 160.5 0
Chironomidae 361 261
Total abundance 1143694 1132658 [species / taxa 9.460.2 1060.3 Diversity, H9 1.8960.04 1.8960.05 Evenness, J 0.5960.01 0.5760.01
4.2. Number of taxa and faunal abundances in the control and fish treatment after 3 days, 1 week, 2 weeks and 3 weeks of experiment
The number of taxa was 9.460.25 in the control and 1060.32 in the fish treatment
when starting the experiment. No difference was found between the control and fish treatment at the start, after 3 days or after 1 week of the experiment. A significant difference (P,0.05) was found between the control and treatment after 2 and 3 weeks
(Fig. 1a). After 3 weeks there were 960.2 and 760.4 taxa in the control and the fish
treatment, respectively. A significant difference (P,0.05) in the number of taxa was
found between the start and end of the experiment for the fish treatment, whereas no difference over time could be found for the control. The total meiofaunal abundance, however, did not differ between the control and fish treatment at any one date (Fig. 1b). The abundance decreased in both the control and treatment during the experimental time, but this reduction was significant only for the fish treatment (P,0.01). After the 3-week
2
experiment the meiofaunal abundance was 984635 ind / 10 cm in the control and 2
933647 ind / 10 cm in the fish treatment.
The abundance of nematodes did not differ significantly between the control and fish treatment at any one date (Fig. 1b). At the start their abundance was 661643 and
2
682638 ind / 10 cm in control and fish treatment, respectively. The nematodes
Fig. 1. Number of taxa (a), meiofaunal abundances (b–e) and major taxon diversity (f) in the control (–s–)
and in the treatment with P. flesus (–d–) over the 3-week experimental period. Error bars indicate S.E.
2
sampling occasion. After 3 weeks nematode abundance was 623630 ind / 10 cm in the 2
control and 659633 ind / 10 cm in the fish treatment.
2
The initial abundance of harpacticoids was 131621 and 123618 ind / 10 cm in the
control and treatment at the start or after 3 days of experiment. After 1 week the abundance of harpacticoids differed significantly (P,0.05) between the control and the
2
fish treatment and there were 12668 and 79615 ind / 10 cm in the control and the
treatment, respectively. A significantly lower number of harpacticoids (P,0.001) was
also found in the fish treatment after 2 and 3 weeks of the experiment (Fig. 1c). After 3
2 2
weeks there were 10465 ind / 10 cm in the control and 5967 ind / 10 cm in the fish
treatment. A significant difference could be found for the fish treatment when comparing the harpacticoids between the start and end of the experiment (P,0.05). In the control
no difference over time was found. The abundance of copepod nauplii differed significantly (P,0.005) between the control and fish treatment after 3 weeks of
2 2
experiment when there were 7565 ind / 10 cm in the control and 3367 ind / 10 cm in
the fish treatment (Fig. 1d). Their abundance decreased in the fish treatment over time and a significant difference (P,0.05) was found when comparing their abundance at
the start and end of the experiment. No difference over time was found for the control. The abundance of ostracods differed significantly between the control and treatment after 2 weeks (P,0.05, Mann–Whitney U-test) and 3 weeks (P,0.01, Mann–Whitney
2
U-test) of experiment (Fig. 1e). The initial ostracod abundance was 1665 ind / 10 cm in 2
the control and 1263 ind / 10 cm in the fish treatment. After 3 weeks there were 761 2
ind / 10 cm in the control, whereas no ostracods could be found in the fish treatment. The abundance of ostracods decreased in both the control and treatment during the experiment, but when comparing abundances at the start and the end of experiment a significant difference was found only for the fish treatment (P,0.05, Wilcoxon’s
signed-ranks test).
The abundances of turbellarians and gastrotrichs did not differ between the control and fish treatment at any dates. Turbellarian and gastrotrich abundances decreased somewhat during the experimental time, but no differences were found when the start and the end values were compared. Oligochaetes showed great fluctuations between samples, and no significant differences could be detected between the control and fish treatment or between start and end for either the control or treatment. Other taxa occurred in low abundances only and did not show any differences between the control and fish treatment, or over the experimental time in either control or treatment.
4.3. Structure of the benthic assemblages
The initial major taxon diversity (H9) in the meiofaunal assemblages was 1.960.04 in
the control and 1.960.05 in the fish treatment. The diversity did not differ between the
control and treatment at the start or after 3 days of the experiment. After one week a significantly lower diversity (P,0.05) was found in the fish treatment. The diversities
were also significantly different after 2 weeks (P,0.05) and 3 weeks (P,0.005) of
experimental time (Fig. 1f). After 3 weeks the diversity was 1.560.05 in the fish
treatment, and 1.860.03 in the control. When comparing the diversity values between
the start and end of the experiment a significantly lower value was found at the end of the experiment in both control (P,0.05) and the fish treatment (P,0.005). In the
evenness varied between 0.51 and 0.59 in both control and fish treatment, and no significant difference was found between the control and treatment at any one date or between the start and end of the experiment in either the control or treatment.
4.4. Nematode assemblage
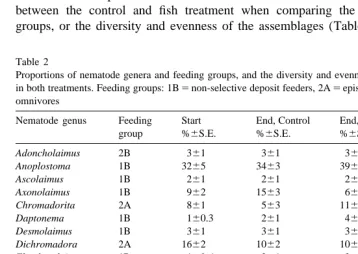
At the start of the experiment the nematodes consisted of 16 genera (Table 2). The most abundant was Anoplostoma comprising 32% of the nematode abundance, followed by Dichromadora (16%), Tripyloides (13%), Axonolaimus (9%), Chromadorita (8%) and Theristus (8%). 69% of the total nematode abundance belonged to non-selective deposit feeders, 27% to epistrate feeders and 4% were predators / omnivorous (Table 2). After 3 weeks the nematode assemblages consisted of 13 genera in both the control and fish treatment, and the dominating genus was Anoplostoma (34 vs. 39% in control vs. fish treatment). A significant difference (P,0.05) between the control and treatment
was found for Axonolaimus which constituted 6% of the nematode assemblage in fish treatment as compared to 15% in the control treatment. No differences could be found between the control and fish treatment when comparing the dominance of feeding groups, or the diversity and evenness of the assemblages (Table 2).
Table 2
Proportions of nematode genera and feeding groups, and the diversity and evenness at start and after 3 weeks in both treatments. Feeding groups: 1B5non-selective deposit feeders, 2A5epistrate feeders, 2B5predators / omnivores
Nematode genus Feeding Start End, Control End, P. flesus Sign. diff
group %6S.E. %6S.E. %6S.E. C vs. P. flesus
Adoncholaimus 2B 361 361 361 Anoplostoma 1B 3265 3463 3961 Ascolaimus 1B 261 261 261
Axonolaimus 1B 962 1563 661 P,0.5 Chromadorita 2A 861 563 1168
Daptonema 1B 160.3 261 462 Desmolaimus 1B 361 361 361 Dichromadora 2A 1662 1062 1062 Eleutherolaimus 1B 160.6 261 261 Enoplolaimus 2B 160 0 160.7 Microlaimus 2A 160.6 0 0 Paracanthonchus 2A 260 262 160.6 Sabatieria 1B 0 0.360.3 0 Sphaerolaimus 2B 0.360.3 0 0 Theristus 1B 862 761 661 Tripyloides 1B 1362 1363 1167
Feeding group 1B 6962 7963 7365
2A 2763 1862 2366
2B 461 361 562
Diversity (H9) 2.9960.12 2.8660.05 2.6860.15
5. Discussion
In this study the predation by juvenile flounder affected the meiofauna in the microcosms and all three 0-hypotheses could be rejected, i.e. (1) the predation affected meiofaunal abundances, (2) harpacticoids were selected, and (3) P. flesus did affect the meiofaunal community structure. The meiofaunal assemblages in the microcosms corresponded well with the natural assemblages (Aarnio et al., 1991; Aarnio and Bonsdorff, 1992), and the remarkable equal starting situation between the treatments (Table 1) further strengthen the results from this experiment and exemplifies the usefulness of microcosms in meiofaunal studies.
5.1. Predation on meiofauna is selective
The number of taxa was reduced in the presence of P. flesus after 2 weeks of predation, and the total meiofaunal abundance decreased compared to the controls. At taxon level the predation by P. flesus had negative effects on the abundances of crustaceans (i.e. harpacticoids, copepod nauplii and ostracods). Harpacticoids are the main food for many juvenile and small fish, including juvenile flounder (Gee, 1989; Coull, 1990; McCall, 1992; Aarnio et al., 1996). This indicates that if some effects of predation are to be found, they would most likely be found for harpacticoids. This was also the case in this study and harpacticoids were clearly selected by P. flesus. This selection for copepods may be an active process or it may only reflect the number of available prey in the sediments. Flatfish consume prey by moving over the bottom and biting prey when encountered (Ansell and Gibson, 1990) and mobile prey will stimulate the predators and increase their attack rate (Ansell, 1995). Harpacticoids are mobile and most species live epibenthically, and are thus susceptible to predation by epibenthic feeding fish. In sand habitats harpacticoids are thought to be more important as prey as there are only few alternative prey organisms of suitable size available (Gee, 1987). On the other hand, Smith and Coull (1987) suggested that predation by epibenthic predators is more effective in mud than in sand, because in sand the prey may have a deeper vertical distribution, whereas in mud all prey are concentrated in the uppermost sediment layer. Several studies conducted in microcosms have revealed negative effects on harpacticoids after a short experimental time, 10–24 h (Marinelli and Coull, 1987; Palmer, 1988; Ellis and Coull, 1989), while here no negative effects were detected until the experiment had run for one week. However, the predator density in my experiment
2 2
was much lower (49 ind / m ) than in the other experiments (180–625 ind / m ). The negative effects on Ostracoda were somewhat surprising, since it has been shown that a large proportion of them may survive the gut passage of fish, and consequently their abundance should not decline so much due to predation unless they are consumed in very large quantities. Vinyard (1979) showed that 26% of ostracods survived the gut passage of Lepomis machrochirus and even a higher percentage, 53–75%, survived the gut passage of juvenile P. flesus (Aarnio and Bonsdorff, 1997). However, the water temperature in my experiment was lower (128C) than in the above mentioned
experiments (15–208C). As water temperature affects the rate of food digestion (De
to a higher mortality of the ostracods. Ostracods are most active at the sediment surface and are thus available food for epibenthic feeding fish. They are also a preferred food item for juvenile flounder ,40 mm in size, and in the northern Baltic ostracods
constituted up to 18% of their diet (Aarnio et al., 1996).
5.2. The role of nematodes
In this study the nematode abundance was not reduced by predation, even though nematodes had been consumed by the fish; 16% of the flounders gut contents consisted of nematodes at the end of the experiment. It has been suggested that nematodes are more or less avoided by fish predators because they are very seldom found in the stomachs of the fish (Gee, 1989 and references therein). Since the abundance of nematodes did not differ between the control and fish treatment in this study, they were apparently not consumed by the flounders in any greater degree and only one nematode genus, Axonolaimus, was significantly reduced in the treatment with P. flesus. Ax-onolaimus is an euryhaline brackish-water genus, and shows a very wide range of occurrence, with regard to both depth and substrate type (Schiemer et al., 1983). It is also a surface dwelling species (Platt, 1977), which is probably the reason why it was affected by the predator. The most common nematode in this study, Anoplostoma, may occur in the deeper, anoxic sediment-layers (Surey-Gent, 1981), and would thus be unavailable food for juvenile P. flesus. The low consumption of nematodes by the fish was thus probably due to the deeper vertical distribution of the nematodes in the sediment (Ellis and Coull, 1989), and not due to an active avoidance of them. However, a few studies have been able to detect reductions in nematodes due to predation by fish (Fitzhugh and Fleeger, 1985; Smith and Coull, 1987; Coull et al., 1995). All these experiments were conducted in muddy sediments, where most prey organisms are situated in the surface layer, and are thus more vulnerable to predation (Smith and Coull, 1987). The flounders had no effects on nematode feeding groups and the assemblages were dominated by non-selective deposit feeders in both the control and fish treatment. The dominance of this trophic group is characteristic for a habitat with fine sand and mud (Giere, 1993).
5.3. Predation affects community structure
results. Epibenthic predation is generally thought to increase stability in benthic communities, leading to a higher diversity due to reduced competition (Peterson, 1979; Reise, 1985; Bonsdorff et al., 1986). Mattila (1992) and Mattila and Bonsdorff (1998) studying predation effects of benthic feeding fish, including juvenile flounder, suggested a stabilizing effect of epibenthic predation in the northern Baltic Sea. The size of their flounders were larger (mean length: 80 mm) than the ones I used in my experiment (mean length: 30 mm), and flounder larger than 50 mm have been shown to have a more variable diet consisting mainly of macrobenthic prey (Aarnio et al., 1996). The smallest flounders are probably more selective in their food choice because of a limited number of alternative prey organisms of suitable size. Thus, the dominance of one taxon, harpacticoids, in the diet of meiofauna-consuming predators may have a destabilizing effect on the meiofaunal assemblage, whereas predation by larger epibenthic predators may be stabilizing for the macrofauna.
5.4. Juvenile flounders are important
The results from this experiment suggest that the meiofaunal assemblages on shallow sandy bottoms in the northern Baltic Sea may be structured by juvenile P. flesus. The
2
density of flounders in this experiment (49 ind / m ) was higher than observed in nature, but as they seem to be irregularly distributed over the habitat the predator density was not unnatural for this region. The sand goby, Pomatoschistus minutus, is another major meiofaunal predator occurring in the same habitat as P. flesus. Juvenile, newly settled, P. flesus inhabits the area from late summer to late autumn, whereas P. minutus occurs in
2
the same habitats from spring to late autumn (12–14 ind / m ; Aarnio and Bonsdorff, 1993). The brown shrimp, Crangon crangon, occurs in the area (Sandberg et al., 1996) and the juveniles may also be considered as potential predators on meiofauna (Mattila et al., 1990). Thus, the combined predation on meiofauna by these epibenthic predators might be considerable during the ice-free period, and they may be able to structure the meiofaunal assemblages. This should however be further tested in field experiments, to account for the physical conditions in the area. Manipulative experiments conducted in laboratory microcosms are particularly appropriate for studies on meiobenthos, and they are an effective mechanism for testing natural predation / disturbance (Bell and Coull, 1978; Findlay and White, 1983; Warwick et al., 1990). As an entire benthic community is enclosed in a microcosm, predation effects on a complex system may be studied. Microcosm experiments differ from field experiments mainly in the lack of physical disturbance (caused by e.g. wave action, currents), and the inability of prey to emigrate from / immigrate to the studied area. As the hydrographical factors may be controlled by the experimenter, effects found on a community can be directly attributed to the predator. However, the restriction of the predator to move about freely in the microcosms may affect the feeding behaviour of the predator. Extrapolating the experimental results to natural conditions may be risky, depending on the characteristics of the area studied.
Acknowledgements
¨
I thank Huso Biological Station for providing excellent working conditions during this study. I am grateful to Prof. Erik Bonsdorff for advice during the work and for giving valuable comments on the manuscript. I am also indebted to Doc. Emil Olafsson, Stockholm University, for guiding me in nematode taxonomy. Financial support was
˚
¨ ¨
received from Stiftelsens for Abo Akademi forskningsinstitut and Kulturfonden for Sverige och Finland.
References
Aarnio, K., Bonsdorff, E., 1992. Colonization rates and community structure of benthic meiofauna in shallow Baltic archipelago waters. Aqua Fennica 22, 71–80.
Aarnio, K., Bonsdorff, E., 1993. Seasonal variation in abundance and diet of the sand goby, Pomatoschistus minutus (Pallas), in a northern Baltic archipelago. Ophelia 37, 19–30.
Aarnio, K., Bonsdorff, E., 1997. Passing the gut of juvenile flounder, Platichthys flesus: differential survival of zoobenthic prey species. Mar. Biol. 129, 11–14.
Aarnio, K., Sandberg, E., Bonsdorff, E., 1991. Benthic predation on shallow-water macro- and meiofauna in the Baltic Sea: an experimental comparison between Pomatoschistus minutus(Pisces) and Saduria entomon (Crustacea). Ann. Zool. Fennici 28, 41–48.
Aarnio, K., Bonsdorff, E., Rosenback, N., 1996. Food and feeding habits of juvenile flounder, Platichthys
˚
flesus (L.), and turbot, Scophthalmus maximus L., in the Aland archipelago, northern Baltic Sea. J. Sea Res. 36, 311–320.
Aarnio, K., Bonsdorff, E., Norkko, A., 1998. Role of Halicryptus spinulosus (Priapulida) in structuring meiofauna and settling macrofauna. Mar. Ecol. Prog. Ser. 163, 145–153.
Alheit, J., Scheibel, W., 1982. Benthic harpacticoids as a food source for fish. Mar. Biol. 70, 141–147. Ansell, A.D., 1995. Surface activity of some benthic invertebrate prey in relation to the foraging activity of
juvenile flatfish. In: Eleftheriou, A., Ansell, A.D., Smith, C.J. (Eds.), Biology and Ecology of Shallow Coastal Waters. Proc. 28th Europ. Mar. Biol. Symp, Olsen & Olsen, Fredensborg, pp. 245–252. Ansell, A., Gibson, R.N., 1990. Patterns of feeding and movement of juvenile flatfishes on an open sandy
beach. In: Barnes, M., Gibson, R.N. (Eds.), Trophic Relationships in the Marine Environment. Proc. 24th Europ. Mar. Biol. Symp, Aberdeen University Press, Aberdeen, pp. 191–207.
Bell, S.S., Coull, B.C., 1978. Field evidence that shrimp predation regulates meiofauna. Oecologia (Berl.) 35, 141–148.
¨ ¨
Bonsdorff, E., Mattila, J., Ronn, C., Osterman, C.-S., 1986. Multidimensional interactions in shallow soft-bottom ecosystems: testing the competitive exclusion principle. Ophelia, Suppl. 4, 37–44.
Coull, B.C., 1990. Are members of the meiofauna food for higher trophic levels? Trans. Am. Microsc. Soc. 109, 233–246.
Coull, B.C., Greenwood, J.G., Fielder, D.R., Coull, B.A., 1995. Subtropical Australian juvenile fish eat meiofauna: experiments with winter whiting Sillago maculata and observations on other species. Mar. Ecol. Prog. Ser. 125, 13–19.
De Groot, S.J., 1971. On the interrelationships between morphology of the alimentary tract, food and feeding behaviour in flatfishes (Pisces: Pleuronectiformes). Neth. J. Sea Res. 5, 121–196.
Ellis, M.J., Coull, B.C., 1989. Fish predation on meiobenthos: field experiments with juvenile spot Leiostomus ´ `
xanthurus Lacepede. J. Exp. Mar. Biol. Ecol. 130, 19–32.
¨
Elmgren, R., Rosenberg, R., Andersin, A.-B., Evans, S., Kangas, P., Lassig, J., Leppakoski, E., Varmo, R., 1984. Benthic macro- and meiofauna in the Gulf of Bothnia (northern Baltic). Finn. Mar. Res. 250, 3–18. Findlay, R.H., White, D.C., 1983. The effects of feeding by the sand dollar Mellita quinquiesperforata (Leske)
Fitzhugh, G.R., Fleeger, J.W., 1985. Goby (Pisces: Gobiidae) interactions with meiofauna and small macrofauna. Bull. Mar. Sci 36, 436–444.
Gee, J.M., 1987. Impact of epibenthic predation on estuarine intertidal harpacticoid copepod populations. Mar. Biol. 96, 497–510.
Gee, J.M., 1989. An ecological and economic review of meiofauna as food for fish. Zool. J. Linnean Soc. 96, 243–261.
Gee, J.M., Warwick, R.M., Davey, J.T., George, C.L., 1985. Field experiments on the role of epibenthic predators in determining prey densities in an estuarine mudflat. Est. Coast. Shelf Sci. 21, 429–448. Giere, O., 1993. Meiobenthology: the Microscopic Fauna in Aquatic Sediments, Springer-Verlag, Berlin. Hicks, G.R.F., 1984. Spatio-temporal dynamics of a meiobenthic copepod and the impact of
predation-disturbance. J. Exp. Mar. Biol. Ecol. 81, 47–72.
Jansson, B.-O., Aneer, G., Nellbring, S., 1985. Spatial and temporal distribution of the demersal fish fauna in a Baltic archipelago as estimated by SCUBA census. Mar. Ecol. Prog. Ser. 23, 31–43.
Kiørboe, T., 1978. Laboratory measurements of digestion rate in 0-group flounder (Platichthys flesus L.) at two temperatures. Ophelia 17, 155–161.
Marinelli, R.L., Coull, B.C., 1987. Structural complexity and juvenile fish predation on meiobenthos: an experimental approach. J. Exp. Mar. Biol. Ecol. 108, 67–81.
Mattila, J., 1992. Can fish predators regulate benthic communities on shallow soft bottoms in the Baltic Sea?
˚
The role of perch, ruffe and roach. PhD-thesis, Abo Akademi University.
Mattila, J., Bonsdorff, E., 1998. Predation by juvenile flounder (Platichthys flesus L.): a test of prey vulnerability, predator preference, switching behaviour and functional response. J. Exp. Mar. Biol. Ecol. 227, 221–236.
´
Mattila, J., Olafsson, E.B., Johansson, A., 1990. Predation effects of Crangon crangon on benthic infauna on shallow sandy bottoms – an experimental study from southern Sweden. In: Barnes, M., Gibson, R.N. (Eds.), Trophic Relationships in the Marine Environment. Proc. 24th Europ. Mar. Biol. Symp, Aberdeen University Press, Aberdeen, pp. 503–516.
McCall, J.N., 1992. Source of harpacticoid copepods in the diet of juvenile starry flounder. Mar. Ecol. Prog. Ser. 86, 41–50.
¨ Nellbring, S., 1985. Abundance, biomass, and seasonal variation of fish on shallow soft bottoms in the Asko
area, northern Baltic proper. Sarsia 70, 217–225.
Nelson, A.L., Coull, B.C., 1989. Selection of meiobenthic prey by juvenile spot (Pisces): an experimental study. Mar. Ecol. Prog. Ser. 53, 51–57.
Olafsson, E., Moore, C.G., Bett, B.Y., 1990. The impact of Melinna palmata Grube, a tube-building polychaete, on meiofaunal community structure in a soft-bottom subtidal habitat. Est. Coast. Shelf Sci. 31, 883–893.
Palmer, M.A., 1988. Epibenthic predators and marine meiofauna: separating predation, disturbance, and hydrodynamic effects. Ecology 69, 1251–1259.
Peterson, C.H., 1979. Predation, competitive exclusion and diversity in the soft-sediment benthic communities of estuaries and lagoons. In: Livingston, R.J. (Ed.), Ecological Processes in Coastal and Marine Systems, Plenum Publishing, London.
Pihl, L., 1982. Food intake of young cod and flounder in a shallow bay on the Swedish west coast. Neth. J. Sea Res. 15, 419–432.
Platt, H.M., 1977. Vertical and horizontal distribution of free-living marine nematodes from Strangford Lough, Northern Ireland. Cah. Biol. Mar. 18, 261–273.
Platt, H.M., Warwick, R.M., 1983. Free-living marine nematodes Part 1. British Enoplids. In: Kermack, D.M., Barnes, R.S.K. (Eds.), Synopses of the British Fauna (New Series), Vol. 38, Cambridge University Press, Cambridge.
¨ Reise, K., 1979. Moderate predation on meiofauna by the macrobenthos of the Wadden Sea. Helgolander wiss.
Meeresunters. 32, 453–465.
Reise, K., 1985. Tidal Flat Ecology, Springer, Berlin.
Sandberg, E., Tallqvist, M., Bonsdorff, E., 1996. The effects of reduced oxygen content on predation and siphon cropping by the brown shrimp, Crangon crangon. P.S.Z.N. I: Mar. Ecol. 17, 411–423.
Scholz, D.S., Matthews, L.L., Feller, R.J., 1991. Detecting selective digestion of meiobenthic prey by juvenile spot Leiostomus xanthurus (Pisces) using immunoassays. Mar. Ecol. Prog. Ser. 72, 59–67.
Service, S.S., Feller, R.J., Coull, B.C., Woods, R., 1992. Predation effect of three fish species and a shrimp on macrobenthos and meiobenthos in microcosms. Est. Coast. Shelf Sci. 34, 277–293.
Shannon, C.E, Weaver, H., 1963. The Mathematical Theory of Communication, University of Illinois Press, Urbana.
Shaw, M., Jenkins, G.P., 1992. Spatial variation in feeding, prey distribution and food limitation of juvenile ¨
flounder Rhombosolea tapirina Gunther. J. Exp. Mar. Biol. Ecol. 165, 1–21.
Smith, D.L., Coull, B.C., 1987. Juvenile spot (Pisces) and grass shrimp predation on meiobenthos in muddy and sandy substrata. J. Exp. Mar. Biol. Ecol. 105, 123–136.
Sogard, S.M., 1984. Utilization of meiofauna as a food source by a grassbed fish, the spotted dragonet, Callionymus pauciradiatus. Mar. Ecol. Prog. Ser. 17, 183–191.
Sokal, R.R., Rohlf, F.J., 1995. Biometry, WH Freeman, New York.
St. John, J., Jones, G.P., Sale, P.F., 1989. Distribution and abundance of soft-sediment meiofauna and a predatory goby in a coral reef lagoon. Coral Reefs 8, 51–57.
Surey-Gent, S.C., 1981. Distribution of Anoplostoma viviparum (Nematoda) in Southampton water sediments. Mar. Biol. 62, 157–160.
Tipton, K., Bell, S.S., 1988. Foraging patterns of two syngnathid fishes: importance of harpacticoid copepods. Mar. Ecol. Prog. Ser. 47, 31–43.
Vinyard, G., 1979. An ostracod (Cypriodopsis vidua) can reduce predation from fish by resisting digestion. Am. Midl. Nat. 102, 188–190.
Warwick, R.M., Clarke, K.R., Gee, J.M., 1990. The effect of disturbance by soldier crabs Mictyris platycheles. H. Milne Edwards on meiobenthic community structure. J. Exp. Mar. Biol. Ecol. 135, 19–33.