Keragaman Gen Major Histocompatibility Complex (MHC) DRB3 pada Sapi Perah Friesh Holland (FH) di Kabupaten Enrekang
Firman Zainal, Muhammad Ihsan Andi Dagong dan Lellah Rahim
Fakultas Peternakan Universitas Hasanuddin Jln. P. Kemerdekaan KM. 10 Makassar, 90245
Email : firmanzainalballu@gmail.com ABSTRAK
Major Histocompatibility Complex (MHC) merupakan kelompok lokus yang terdiri atas kumpulan gen penting (major) yang mengendalikan respon imun. Penelitian ini bertujuan untuk mengetahui keragamangen Gen Major Histocompatibility Complex (MHC) Sapi Perah Friesh Holland (FH) di Kabupaten Enrekang. Sebanyak 80 sampel darah dikoleksi dari ternak sapi FH dari kabupaten Enrekang yang berasal dari dua kecamatan (Kec. Cendana (n=30) dan Anggeraja (n=50)). Total DNA diekstraksi dari sampel darah dengan Kit GeneJet Genomic DNA extraction.Gen MHC kemudian diamplikasi dengan teknik PCR (Polymirase Chain Reaction). Untuk genotyping, produk PCR yang dihasilkan kemudian dipotong dengan menggunakan enzim retriksi HaeIII.Analisis polimorfisme meliputi frekuensi Alel dan Genotipe, heterozigositas pengamatan (Ho), heterozigositas harapan (He) dan keseimbangan Hardy Weinberg.Variasi genetic diantara populasi dihitung berdasarkan frekuensi alel dan genotipnya. Adapun frekuensi alel Gen MHC HaeIII yg ditemukan pada penelitian sapi perah FH yaitu alel B, C, D, E dan F. Alel C merupakan alel yang umum pada populasi ini sedangkan alel B dan D adalah alel yang langkah.Dari hasil penelitian, genotype yang ditemukan yaitu BD (0,02), BE (0,02), CC (0,35), CE (0,27), CF (0,13), EE (0,08), dan EF (0,12). Nilai heterozigositas pengamatan pada sapi Perah FH (0,56), Sedangkan nilai heterozigositas harapan yaitu (0,59).Nilai chi-square yaitu 163,9 (P<0,05) menunjukkan bahwa MHC HaeIII tidak berada dalam kesetimbangan Hardy-Weinberg.
Kata Kunci :SapiPerah FH, Gen MHC, Alel, PCR-RFLP
PENDAHULUAN
Sapi perah Friesian Holstein (FH) atau yang dikenal dengan namaFries Holland di Negara Belanda ini berasal dari nenek moyang sapi liar Bos (Taurus) Typicus Primigenius yang tidak berpunuk. Sapi FH ditemukan di Propinsi BelandaUtara dan Friesland Barat, Belanda (Schmidt & Vleck, 1974). Sapi ini di AmerikaSerikat disebut dengan Holstein Friesian atau disingkat Holstein dan di Eropadisebut Friesian, sedangkan di Indonesia terkenal dengan nama Friesian Holstein (Sudono, 2003).
Sapi perah merupakan ternak ruminansia yang dipelihara hanya pada daerah yang beriklim dingin demi tercapainya produksi susu yang maksimal, disebabkan sapi perah pada umumnya berasal dari daerah yang beriklim subtropis berbeda dengan iklim yang ada di indonesia yang beriklim tropis, oleh karena itu Kabupaten Enrekang merupakan salah satu daerah daerah yang ada di Sulawesi Selatan yang mengembangkan pemeliharaan sapi perah karena memiliki iklim yang sesuai dan memudahkan sapi perah dalam menyesuaikan diri, Salah satu jenis sapi perah yang
ada di Kabupaten Enrekang yaitu sapi FH yang berpotensi untuk dikembangkan karena adaptif terhadap lingkungan di Kabupaten Enrekang serta memiliki daya produksi susu yang tinggi. Akan tetapi hingga saat ini program pemuliaan dalam rangka meningkatkan kualitas genetik sapi perah FH belum menunjukkan hasil yang signifikan, informasi yang terkait tentang keragaman genetiknya sampai saat ini belum tersedia khususnya di Kabupaten Enrekang.
Sifat polimorphisme dari gen MHC dapat terjadi akibat dari kawin silang antara sapi perah FH dengan jenis sapi perah yang lainnya menimbulkan perubahan karakter-karakter genetik yang berbeda dengan induknya.Keragaman genetik dalam suatu populasi sapi perah akan mengakibatkan perbedaan reaktivitas imun pada setiap individu dalam suatu populasi sehingga berpengaruh terhadap ketahanan dan kerentanan individu terhadap penyakit. Dari permasalahan tersebut kemungkinan ada perbedaan imunogenetik.Oleh karena itu dilakukan penelitian untuk menguji seberapa besar terjadinya perubahan konstribusi genetik antara sapi perah FH yang ada di Kabupaten Enrekang, melalui analisis gen MHC.
MATERI DAN METODE Koleksi Sampel Darah
Sampel darah diperoleh dari Kabupaten Enrekang. Pengambilan sampel darah dilakukan dengan mengumpulkan sekitar 5 ml sampel darah dari sapi perah FH melalui vena jugularis dengan menggunakan venojet dan tabung vacuttainer dengan EDTA dan kemudian disimpan pada suhu 4oC.
Ekstraksi DNA
DNA diisolasi dan dimurnikan dengan menggunakan Kit DNA ekstraksi Genjet Genomic DNA Extraction (Thermo Scientific) dengan mengikuti protokol ekstraksi yang disediakan. Sebanyak 200 µl sampel darah dilisiskan dengan menambahkan 400 µl larutan lysis buffer dan 20 µl proteinase K (10 mg/ml), campuran kemudian diinkubasi pada suhu 56oC selama 60 menit didalam waterbath shaker. Setelah inkubasi larutan kemudian ditambahkan 200 µl ethanol absolute 96% dan disentrifugasi 6.000 x g selama 1 menit.
Pemurnian DNA dilakukan dengan metode spin column dengan penambahan 500 µl larutan pencuci wash buffer I yang kemudian dilanjutkan dengan sentrifugasi pada 8.000 x g selama 1 menit. Setelah supernatannya dibuang, DNA kemudian dicuci lagi dengan 500 µl wash buffer II dan disentrifugasi pada 12.000 x g selama 3 menit. Setelah supernatan dibuang, DNA kemudian dilarutkan dalam 200 µl elution buffer dan disetrifugasi pada 8.000 x g untuk selanjutnya DNA hasil ekstraksi ditampung dan disimpan pada suhu -20oC.
Tekhnik PCR-RFLP
Komposisi reaksi PCR dikondisikan pada volume reaksi 25 ul yang terdiri atas 100 ng DNA, 0.25 mM masing-masing primer, 150 uM dNTP, 2.5 mM Mg2+, 0.5 U Taq DNA polymerase dan 1x buffer. Kondisi mesin PCR dimulai dengan denaturasi awal pada suhu 94 oC x 2 menit, diikuti dengan 35 siklus berikutnya masing-masing denaturasi 94 oC x 45 detik, dengan suhu annealing yaitu : 64 oC x 60 detik, yang dilanjutkan dengan ekstensi : 72 oC x 60 detik, yang kemudian diakhiri dengan satu siklus ekstensi akhir pada suhu 72 oC selama 5 menit dengan menggunakan mesin
PCR (SensoQuest, Germany). Produk PCR kemudian dielektrofhoresis pada gel agarose 1.5 % dengan buffer 1x TBE (89 mM Tris, 89 mM asam borat, 2 mM Na2EDTA) yang mengandung 100 ng/ml ethidium bromide. Kemudian divisualisasi pada UV transiluminator (gel documentation system).
Produk PCR yang diperoleh dari masing-masing gen target kemudian dianalisis menggunakan RFLP melalui pemotongan menggunakan enzim restriksi yang memiliki situs pemotongan pada gen MHC HaeIII (gg cc). Sebanyak 5 l DNA produk PCR ditambahkan 0,3 l enzim restriksi (5U) ; 0,7 l buffer enzim dan 1 l milique water sampai volume 7 l, Selanjutnya dilakukan inkubasi selama 17 jam pada suhu 37oC. Lokus ditentukan dengan cara menginterpretasi pita (band) yang terbentuk paling jauh migrasinya ke kutub anoda sebagai lokus 1, lokus 2, dan seterusnya, cara yang sama dipakai juga untuk menentukan alel, yaitu pita yang bermigrasi paling jauh pada suatu lokus ditandai sebagai alel "a", berikutnya alel "b" dan seterusnya.
Analisa Data
Keragaman alel mikrosatelit ditentukan dari perbedaan migrasi alel pada gel dari masing-masing individu sampel. Karena alel mikrosatelit adalah kodominan maka genotip ditentukan berdasarkan variasi pita alel yang ada.Kemudian dihitung frekuensi masing-masing alel setiap lokus.Keseimbangan Hardy- Weinberg di uji dengan test chi-square (Nei dan Kumar, 2000).
HASIL DAN PEMBAHASAN Amplifikasi Gen MHC DRB3exon 2
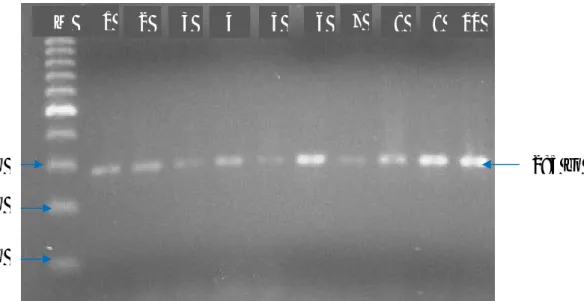
Berdasarkan penelitian yang telah dilakukan bahwa Gen MHC DRB3exon 2 berhasil diamplifikasi dengan menggunakan mesin thermocycler Senso Quest Germany, dengan suhu annealing 64oC. Hasil amplifikasi ruas gen dapat divisualisasikan pada gel agarose 1,5% yang dapat dilihat pada Gambar 1. Panjang produk hasil amplifikasi gen MHC DRB3exon 2 adalah 285 bp.66
6
Gambar 1. Hasil Amplifikasi Gen MHC DRB3exon 2 yang divisualisasi pada Gel Agarose 1,5%.M : Marker (100 bp) ; 1-10 : sampel Sapi Perah FH dari Kabupaten Enrekang ; bp : base pair
6
4
5
1
2
3
7
8
9
10
M
300 bp
200 bp
100 bp
285 bp
Panjang fragmen gen MHC DRB3exon 2 pada penelitian yang dihasilkan yaitu 285 bp. Hal ini sesuai dengan penelitian yang dihasilkan oleh Ahmed (2006), bahwa panjang produk PCR yang dihasilkan adalah 285 bp. Sedangkan Wu et al., (2010); Chu et al., (2012) menghasilkan panjang produk PCR untuk BoLA yaitu 284bp dan 280 bp pada penelitian yang dilakukan oleh Gilliespie et al., (1999).
Keragaman Gen MHC DRB3exon 2 dengan Metode PCR-RFLP
Hasil visualisasi menggunakan gel Polyacrylamide 8% dengan melihat panjang fragmen ruas gen MHC DRB3exon 2 dan enzim HaeIII sebagai enzim pemotong menunjukkan bahwa fragmen yang didapatkan adalah 7 macam genotipe, diantaranya, BD, BE, CC, CE CF, EE, EF. Terdapat dua macam genotipe yang homozigot dan lima genotipe yang heterozigot. Hasil tersebut menunjukkan bahwa gen MHC DRB3exon 2 pada sapi FH yang diamati sangat beragam. Hasil pendeteksian keragaman gen MHC HaeIII dapat dilihat pada Gambar 2.
Gambar 3. Hasil pendeteksian keragaman gen MHC DRB3 exon 2; 1-14: sampel sapi Perah Fries Holland dari Kabupaten Enrekang.
Hasil yang diperoleh pada penelitian ini yaitu masing-masing genotipe dibedakan berdasakan jumlah pita yang muncul dalam gel Polyacrylamide 8%.Jumlah alel dilihat berdasarkan jumlah potongan pita dengan panjang potongan yang berbeda seperti yang ditampilkan pada Tabel 2.
Tabel 2. Potongan Pita DNA Gen MHC HaeIII (Ahmed, 2006)
Alel Pita A / BoLA-DRB*0201 168/117 B / BoLA-DRB*0202 154/14/117 C / BoLA-DRB*0203 154/14/52/65 D / BoLA-DRB*0204 154/131 E / BoLA-DRB*0205 220/65 F / BoLA-DRB*0206 40/180/65
Pita-pita DNA yang terdapat dalam Tabel 1 memiliki jarak laju migrasi yang berbeda-beda sehingga dapat dibedakan menjadi 6 tipe pita (alel) dengan basis pasang
basa. Keenam tipe alel tersebut yaitu alel A (168/117), B (154/14/117), C (154/14/52/65), D (1544/131), E (220/65) dan F (40/180/65).Alel BoLA DRB3 exon 2 yang telah teridentifikasi dengan metode PCR-RFLP hingga saat ini ada 54 alel (Van Eijk et al., 1992; Gelhaus et al., 1995; Maillard et al., 1999).
Frekuensi Genotipe dan Alel
Hasil analisis frekuensi genotipe pada fragmen gen MHC sapi Perah FH dapat dilihat pada Tabel 3.
Tabel 3. Frekuensi Genotipe Gen MHC HaeIII
Populasi Jumlah (ekor)
Frekuensi Genotipe
CC BD CE EE CF BE EF FH 80 0,35 0,02 0,27 0,08 0,13 0,02 0,12
Frekuensi genotipe fragmen gen MHC pada sapi Perah FH memiliki 7 macam genotipe yaitu BD, BE, CC, CE CF, EE, dan EF. Sapi Perah FH memiliki genotipe CC yang lebih tinggi (0,35), sedangkan genotype yang paling rendah yaitu BD dan BE yaitu (0,02). Pada penelitian yang dilakukan oleh Chu et al., (2012) menemukan 7 macam genotipe dari sapi Beijing. Pendeteksian keragaman gen MHC DRB3 exon 2 dengan metode PCR-RFLP dengan HaeIII sebagai enzim pemotong (gg|cc).
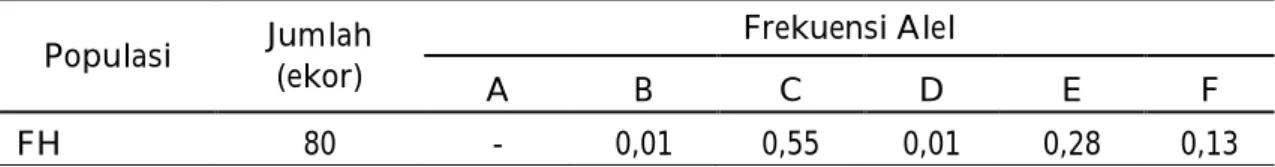
Tabel 4. Frekuensi Alel Gen MHC HaeIII Populasi Jumlah
(ekor)
Frekuensi Alel
A B C D E F
FH 80 - 0,01 0,55 0,01 0,28 0,13
Frekuensi alel yang tertinggi pada sapi Perah FH yang berasal dari Kabupaten Enrekang terdapat pada alel C (0,55) dan frekuensi alel terendah terdapat pada alel B dan alel D (0,01). Jumlah alel pada penelitian ini lebih sedikit dibandingkan alel yang ditemukan pada penelitian sapi bali sebelumnya oleh Puja et al.,(2011), yakni pada sapi Bali jantan asal Bali teridentifikasi sebanyak 7 alel dan pada sapi Bali jantan asal Nusa Penida sebanyak 9 alel. Sedangkan penelitian yang dilakukan oleh Trinayani et al., (2013) menemukan 10 alel.Jumlah alel yang ditemukan pada sapi Bali masih lebih sedikit dibandingkan pada bangsa sapi lainnya, seperti yang dilaporkan Acosta-Rodriguez et al.,(2005) yang mengidentifikasi 18 alel pada sapi persilangan antara bangsa sapi Eropa, Simmenthal, Holstein dan Zebu.
Nilai Heterozigositas
Keragaman genetik adalah penyimpangan sifat atau karakter dari individu yang terjadi karena perkawinan alami yang tidak terkontrol. Keragaman genetik dapat dilihat dari karakter alel dari lokus tertentu yang merupakan ekspresi dari gen tertentu (Johari, dkk., 2007). Hasil analisis nilai heterozigositas pada fragmen gen MHC sapi Perah FH dapat dilihat pada Tabel 5.
Tabel 5. Nilai Heterozigositas Gen MHC HaeIII
Populasi N Heterozigositas Nei X2
Ho He
FH 80 0,56 0,59 0,59 163,91
n= jumlah sampel (ekor) ; Ho= Heterozigositas pengamatan ; He= Heterozigositas harapan;X2= chi-square
Nilai heterozigositas pengamatan (HO) dan heterozigositas harapan (He) digunakan untuk menduga keragaman genetik. Heterozigositas harapan (He) merupakan penduga keragaman genetik yang tepat pada populasi hewan ternak karena perhitungannya langsung berdasarkan pada frekuensi alel (Moioli, et al.,2004) Menurut Nei dan Kumar (2000), keragaman genetik dapat diukur berdasarkan nilai heterozigositas.Nilai heterozigositas merupakan cara yang paling akurat untuk mengukur variasi genetik. Nilai heterozigositas pengamatan pada sapi Perah FH (0,56) .Sedangkan nilai heterozigositas harapan pada yaitu (0,59) . Menurut Tambasco, et al (2003), perbedaan antara nilai heterozigositas pengamatan (Ho) dan nilai heterozigositas harapan (He) dapat dijadikan indikator tidak adanya keseimbangan genotipe pada populasi yang diamati.
KESIMPULAN
Berdasarkan penelitian yang telah dilakukan maka dapat ditarik kesimpulan sebagai bahwa Gen MHC pada populasi sapi Perah Friesh Holland (FH)di Kabupaten Enrekang bersifat polimorfik, terdapat 7 macam genotipe Terdapat dua macam genotipe yang homozigot dan lima genotipe yang heterozigot, Frekuensi alel yang tertinggi pada sapi Perah FH yang berasal dari Kabupaten Enrekang terdapat pada alel C (0,55) dan frekuensi alel terendah terdapat pada alel B (0,01), Hasil analisis menunjukkan bahwa alel yang umum ditemukan pada sapi Perah FH yaitu alel C. Hasil uji chi square terhadap genotip MHC DRB3exon 2| HaeIII menunjukkan bahwa frekuensi genotipe tidak berada dalam keseimbangan Hardy-Weinberg.
DAFTAR PUSTAKA
Acosta-Rodriguez, R., R. Alonso-Morales, S. Balladares, H. Flores-Aguilar, Z. Garcia-Vazquez, and C. Gorodezky. 2005. Analysis of BoLA class II microsatellites in cattle infested with Boophilus microplus ticks: class II is probably associated with susceptibility. Vet. Parasitol., 127: 313–21.
Ahmed, S. and E. Othman. 2006. A PCR-RFLP method for the analysis of Egyptian goat MHC class ii DRB gene. Department of Cell Biology, National Research Center, Dokki, Egypt. Biotechnology 5 (1): 58-61
Chu, M.X., S. C. Ye, L. Qiao, J. X. Wang, T. Feng, D.W. Huang, G. L. Cao, R. Di, L. Fang, G. H. Chen. 2012. Polymorphism of exon 2 of BoLA-DRB3 gene and its relationshipwith somatic cell score in Beijing Holstein cows. Mol Biol Rep (2012) 39:2909–2914 DOI 10.1007/s11033-011-1052-3
Gilliespie, B. E. B. M. Jayarao, H. H. Dowlen and S. P. Oliver. 1999. Analysis and frequency of bovine lymphocyte antigen DRB3.2 Alleles in Jersey cows. Department of Animal Science, Institute of Agriculture, The University of Tennessee, Knoxville, 37996. Department of Veterinary Science. Pennsylvania State University, University Park, 16802-3500
Gelhaus, A., L. Schnittger, D. Mehlitz, R. D. Horstmann, and C. G. Meyer. 1995. Sequence and PCR-RFLP analysis of 14 novel BoLA-DRB3 alleles. Anim. Genet. 26:147–153.
Johari S., E. Kurnianto, Sutopo dan S. Aminah. 2007. Keragaman protein darah sebagai parameter biogenetik pada sapi Jawa. Journal Indonesian Tropical Agriculture, 32[2] Juni 2007. Universitas Diponegoro. Semarang. hal:112-118.
Maillard J. C., I. Chantal I, D. Berthier, I. Sidibe and H. Razandraibe. 2000. BoLA-DRB/DQB haplotypes as molecular markers of genetic susceptibility and resistance to bovine dermatophilosis. ISAG 2000 (Abstract). Proceedings ofthe 27th International Conference on Animal Genetics. July 22±26, 2000. Minneapolis, Minnesota, p. 62.
Moioli, B., F. Napolitano and G. Catillo. 2004. Genetic diversity between Peidmontese, Maremmana and Podolica cattle breeds. J. Hered. 95:250-256.
Nei M and Kumar S. 2000.Molecular Evolution and Phylogenetics. New York: Oxford University, Press.
Puja, I. K., I. N. Wandia, P. Suastika, I. N.Sulabd. 2011. Polimorfisme genetik DNA mikrosatellite gen BoLA lokus DRB3 pada sapi Bali (Bos indicus). Biota Vol. 16 (2): 336−341, Juni 2011 ISSN 0853-8670
Sudono, A. 2003. Ilmu Produksi Ternak Perah. Fakultas Peternakan Institut Pertanian Bogor, Bogor
Schmidt, G. H. dan L. D. Van Vleck. 1974. Principles of Dairy Science. Cornel Univ. San Fransisco.
Tambasco D. D., C. C. P. Paz, M. Tambasco-Studart, A. P. Pereira, M. M. Alencar, A. R. Freitas, L. L. Countinho, I.U. Packer and L. C. A. Regitano. 2003. Candidate genes for growth traits in beef cattle Bos Taurus x Bos Indicus. J. Anim. Bred. Genet. 120: 51-60
Trinayani, N. N., I. N. Wandia, I. K. Puja. 2013. Asosiasi keragaman lokus DNA mikrosatelit DRB3 gen BoLA dengan berat badan induk dan berat lahir pedet pada sapi bali. Vol . 1, No. 2: 58-63
Van Eijk M. J. T., Stewart-Haynes J. A. & Lewin H. A. (1992) Extensive polymorphism of the BoLA-DRB3 gene distinguished by PCRRFLP. Animal Genetics, 23, 483±96.
Wu, X. X., Z. P. Yang, X. L. Wang, Y. J. Mao, S. C. Li, X. K. Shi, Y. Chen. 2010. Restriction fragment length polymorphism in the exon 2 of the BoLA-DRB3 gene in Chinese Holstein of the south China. J. Biomedical Science and Engineering 3: 221-225.