II TINJAUAN PUSTAKA
2.1 Ubi JalarUbi jalar atau ketela rambat atau “sweet potato” diduga berasal dari Benua Amerika. Para ahli botani dan pertanian memperkirakan daerah asal tanaman ubi jalar adalah Selandia Baru, Polinesia, dan Amerika bagian tengah. Nikolai Ivanovich Vavilov, seorang ahli botani Soviet, memastikan daerah sentrum primer asal tanaman ubi jalar adalah Amerika Tengah. Ubi jalar mulai menyebar ke seluruh dunia, terutama negara-negara beriklim tropika pada abad ke-16. Orang-orang Spanyol menyebarkan ubi jalar ke kawasan Asia, terutama Filipina, Jepang, dan Indonesia (Prihatman 2000).
Klasifikasi lengkap Ubi jalar (Ipomoea batatas L) adalah: kingdom : Plantae (tumbuh-tumbuhan) divisi : Spermatophyte (tumbuhan berbiji) sub divisi : Angiospermae (berbiji tertutup) kelas : Dycotyledoneae (biji berkeping dua)
ordo : Concolvulalesm
famili : Convolvuceae
genus : Ipomoea
spesies : Ipomoea batatas L
Menurut Prihatman (2000), plasma nutfah (sumber genetik) tanaman ubi jalar yang tumbuh di dunia diperkirakan berjumlah lebih dari 1000 jenis, namun baru 142 jenis yang diidentifikasi oleh para peneliti. Lembaga penelitian yang menangani ubi jalar, antara lain: International Potato centre (IPC) dan Centro Internationale de La Papa (CIP). Di Indonesia, penelitian dan pengembangan ubi jalar ditangani oleh Pusat Penelitian dan Pengembangan Tanaman Pangan atau Balai Penelitian Kacang-Kacangan dan Umbi-Umbian (Balitkabi), Departemen Pertanian. Varietas atau kultivar atau klon ubi jalar yang ditanam di berbagai daerah jumlahnya cukup banyak, antara lain: Lampenueng, Sawo, Cilembu, Rambo, SQ-27, Jahe, Kleneng, Gedang, Tumpuk, Georgia, Layang-layang, Karya, Daya, Borobudur, Prambanan, Sukuh, Mendut, dan Kalasan.
Menurut Lingga et al. (1986), umbi dari ubi jalar bermacam-macam tergantung dari varietas tanaman yang diusahakan. Tapi umumnya hasil umbi dibagi dua golongan yakni ubi yang berumbi keras (karena banyak mengandung tepung) dan ubi yang berumbi lunak (karena banyak mengandung air dan berdaging manis). Umbi putih mengandung kadar air yang lebih sedikit dibandingkan dengan ubi merah.
Ubi jalar memiliki waktu panen yang lebih singkat jika dibandingkan dengan beberapa tanaman umbi di indonesia. Tabel 1 menunjukkan perbandingan karakterisitik beberapa tanaman umbi di Indonesia. Umur panen ubi jalar lebih singkat (3–3.5 bulan) serta produktivitas yang cukup tinggi (10–30 ton/ha), selain itu ubi jalar merupakan tanaman yang cocok ditanam pada daerah yang marjinal yang tidak terlalu subur. Ubi jalar tidak membutuhkan pupuk yang banyak untuk tumbuhnya. Melihat beberapa keunggulan dari ubi jalar tersebut, maka diharapkan ubi jalar dapat digunakan sebagai salah satu alternatif bahan baku pada pembuatan bahan bakar yang bersumber dari alam (biofuel).
2.2 Sirup Glukosa
Sirup glukosa merupakan salah satu jenis dari gula cair, dimana komponen utamanya adalah glukosa sehingga disebut sirup glukosa. Namun bila komponen utamanya adalah fruktosa yang dipekatkan, maka dikenal sebagai HFS (High Fruktose Syrup) yang memiliki derajat kemanisan yang lebih tinggi.
Pembuatan sirup glukosa dilakukan dengan menghidrolisa pati yang merupakan bahan baku pembuatnya. Hidrolisa pati dapat dilakukan dengan cara hidrolisa dengan katalis asam, kombinasi asam dan enzim, serta kombinasi enzim dan enzim (Judoamidjojo, 1989). Pada hidrolisa pati dengan asam, diperlukan suhu tinggi yaitu 140–160 0C. Asam akan memecah molekul pati secara acak dan gula yang dihasilkan sebagian besar merupakan gula pereduksi.
Hidrolisa pati dengan asam mempunyai kelemahan antara lain yaitu diperlukan peralatan yang tahan korosi, menghasilkan sakarida dengan spektra-spektra tertentu saja karena katalis asam menghidrolisa secara acak. Kelemahan lain, jika nilai ekuivalen dekstrosa ditingkatkan, disamping terjadi degradasi
7
Tabel 1 Perbandingan karakteristik tanaman umbi
Karakteristik Ubi Jalar Singkong Tebu Kentang Talas
Jenis Tanah Cocok untuk semua jenis tanah
Cocok untuk semua jenis tanah
Tanah lembab Tanah lembab Tanah lembab
pH 5.5 – 7.5 4.5 – 8 5 - 6 5.5-6.0 5 - 6
Kebutuhan pupuk rendah Rendah tinggi Tinggi tinggi
Masa panen (bulan) 3 – 3.5 6 – 12 8 -14 3-7 6 - 10
Kandungan karbohidrat (%) bk 98.13 51.36 Kadar sukrosa = 10% 18 21-27 Produktivitas (ton/ha) 11 - 30 10 -13 90 13,7 30 Sumber: Deptan 2005
karbohidrat, juga terjadi rekombinasi produk degradasi yang dapat mempengaruhi warna, rasa, bahkan menimbulkan masalah teknis.
Hidrolisa pati dengan enzim dapat menghasilkan sirup dengan DE lebih dari 95 persen. Selain itu, penggunaan enzim dapat mencegah terjadinya reaksi sampingan karena sifat enzim sangat spesifik, sehingga dapat mempertahankan flavour dan aroma bahan dasar. Pembuatan sirup glukosa dengan menggunakan enzim terdiri atas dua tahap penting yaitu likuifikasi dan sakarifikasi.
Pada proses likuifikasi, pati diubah menjadi glukosa, maltosa, maltotriosa dan oligosakarida. Proses ini berlangsung pada suhu yang tinggi dengan menggunakan enzim yang tahan serta lebih efektif bekerja pada suhu tinggi yaitu α-amilase. Tahap selanjutnya adalah proses sakarifikasi. Pada tahap ini, larutan hasil likuifikasi diturunkan suhunya hingga mencapai 55–58 0C, setelah itu ditambahkan enzim AMG (amiloglukosidase). Pada tahap ini, enzim akan merubah semua karbohidrat menjadi glukosa. Proses pada tahap sakarifikasi ini berlangsung pada suhu 55–58 0C selama 48–96 jam.
Setelah proses sakarifikasi selesai, selanjutnya hidrolisat yang diperoleh disaring dan dihilangkan warnanya dengan menggunakan karbon aktif. Untuk mendapatkan konsentrasi sirup glukosa yang tinggi maka dilakukan proses evaporasi.
Enzim yang digunakan pada pembuatan sirup glukosa dimulai dari tahap likuifikasi hingga sakarifikasi adalah sebagai berikut:
α- Amilase
Menurut Judoamidjojo et al. (1989), α-amilase termasuk enzim pemecah dari dalam molekul, bekerja menghidrolisa dengan cepat ikatan α-1,4 glukosida pati yang telah mengalami gelatinisasi. Pada proses likuifikasi, α-amilase akan memecah ikatan 1,4 pada amilosa dan menghasilkan dekstrin. Aktivitas α-amilase pada amilopektin menghasilkan oligosakarida dengan jumlah monomer dua sampai enam.
Mekanisme kerja enzim ini pada amilosa dibagi dalam dua tahap, yaitu: (1) Degradasi secara cepat molekul amilosa menjadi maltosa dan maltotriosa yang
(2) Pembentukan glukosa dan maltosa dengan laju lebih lambat tidak secara acak. Degradasi α-amilase pada amilopektin akan menghasilkan glukosa, maltosa dan berbagai jenis α-limit dekstrin.
Sumber enzim α-amilase sangat beragam, mulai dari tanaman, jaringan mamalia sampai mikroorganisme. Saat ini sumber enzim α-amilase yang paling potensial dan banyak digunakan di industri berasal mikroorganisme. Salah satu bakteri penghasil enzim α-amilase adalah yang berasal dari spesies Bacillus. Bakteri ini menghasilkan enzim yang tahan terhadap panas (termostabil) (Judoamidjojo, 1989).
Amiloglukosidase
Enzim amiloglukosidase atau disebut juga glukoamilase (1,4 glucan glucohydrolase, EC. 3.2.1.3) mengkatalis pemotongan gugusan glukosa dari ujung non reduksi dari polimer pati, hasil akhir dari hidrolisa adalah senyawa glukosa (Schwimmer, 1981). Enzim amiloglukosidase dapat menghidrolisis ikatan α-1,6 glukosida namun reaksinya berlangsung lambat. Aktivitas optimal dari enzim amiloglukosidase sangat dipengaruhi oleh pH dan suhu.
Secara komersial, enzim amiloglukosidase diproduksi dari Aspergillus niger dan Rhizopus sp. Amiloglukosidase dapat menghidrolisa pati sampai mencapai DE 95 – 98 persen. Produk hidrolisa yang dihasilkan bersifat lebih manis, namun cenderung membentuk kristal disebabkan kandungan dekstrosanya yang tinggi (Judoamidjojo, 1989).
2.3 Bioetanol
Bioetanol merupakan produk yang dihasilkan dari fermentasi gula, walaupun sebenarnya, etanol juga dapat dibuat dengan menggunakan reaksi kimia dengan menggunakan etilene. Etanol atau etil alkohol (C2H5OH) merupakan
cairan jernih tidak berwarna, biodegradable, rendah kadar racunnya, dan sangat sedikit menimbulkan polusi bagi lingkungan apabila tertumpah
(www.indobiofuel.com).
Jika dibakar, etanol menghasilkan karbondioksida dan air. Dengan mencampurkan etanol dan bensin, maka dapat dihasilkan bahan bakar campuran yang dapat terbakar dengan sempurna dan dapat mengurangi emisi pencemaran
udara. Bahan bakar campuran ini telah banyak digunakan secara luas di Amerika Serikat. Kendaraan yang tanpa modifikasi dapat menggunakan bahan bakar campuran 10% etanol dan 90% bensin atau yang lebih dikenal dengan gasohol (Gasoline - alcohol fuels). Sekarang telah ada kendaraan yang dapat menggunakan bahan bakar campuran dengan kandungan 85% etanol dan 15% bensin (www.ristek.go.id)
Gambar 1 Rumus bangun etanol.
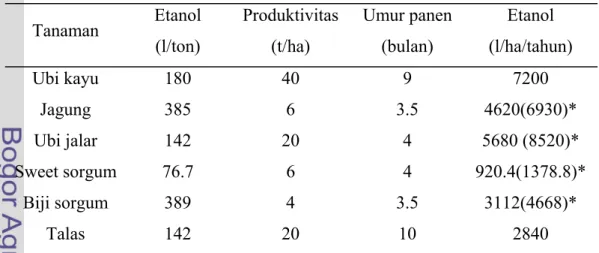
Bioetanol memiliki beberapa keuntungan diantaranya berasal dari bahan yang dapat diperbaharui, serta tidak menghasilkan emisi gas yang dapat menimbulkan polusi udara. Keuntungan lainnya yaitu biodegradabel, serta cara pembuatannya yang sederhana yaitu melalui fermentasi gula–gula sederhana yang berasal dari tumbuhan (www.ristek.go.id). Tabel 2 berikut memperlihatkan beberapa tanaman penghasil bioetanol.
Tabel 2 Tanaman Penghasil Bioetanol
Tanaman Etanol (l/ton) Produktivitas (t/ha) Umur panen (bulan) Etanol (l/ha/tahun) Ubi kayu 180 40 9 7200 Jagung 385 6 3.5 4620(6930)* Ubi jalar 142 20 4 5680 (8520)* Sweet sorgum 76.7 6 4 920.4(1378.8)* Biji sorgum 389 4 3.5 3112(4668)* Talas 142 20 10 2840
* diganti oleh penulis dengan 3 kali panen ubi jalar dalam setahun Sumber: Shintawaty 2006
Menurut Judoamidjojo et al. (1989), proses fermentasi pembentukan etanol membutuhan bantuan yeast atau khamir. Untuk bahan yang mengandung gula dalam bentuk polisakarida atau oligosakarida, terlebih dahulu harus dirubah dulu dalam bentuk yang lebih sederhana yaitu monosakarida (fruktosa atau glukosa). Yeast tersebut akan merubah gula–gula sederhana yaitu fruktosa atau glukosa (C6H12O6) menjadi etanol (C2H5OH) dan karbondioksida (CO2). Reaksi yang
terjadi adalah sebagai berikut (Judoamidjojo, 1990).
C6H12O6 2C2H5OH + 2CO2 Monosakarida Etanol Karbondioksida
Etanol memiliki berat jenis pada 15 ºC adalah sebesar 0.7937 g/ml dan titik didihnya 78.32 ºC pada tekanan 766 mmHg. Sifatnya yang lain adalah larut dalam air dan eter dan mempunyai panas pembakaran 328 Kkal. Penggunaan etanol yang terbanyak adalah sebagai pelarut sebesar 40 persen, untuk membuat asetildehid 36 persen, untuk penggunaan secara kimiawi yang lain 15 persen, serta eter, glikol eter, etil asetat dan khoral 9 persen (Paturau, 1981).
Menurut Paturau (1981), fermentasi etanol membutuhkan waktu 30–72 jam. Frazier dan Westhoff (1978) menambahkan suhu optimum fermentasi 25–30 oC dan kadar gula 10–18 persen.
Beberapa jenis substrat telah digunakan untuk memproduksi bioetanol diantaranya adalah dapat dilihat pada Tabel 3 berikut:
Tabel 3 Jenis Substrat serta Mikroorganisme yang digunakan untuk memproduksi Etanol
Substrat Kadar etanol tertinggi Mikroorganisme Ubi Jalar(a) 9.8 g/l (1.23% v/v) S. cerevisiae
Gandum (b) 6.9 g/l (0.869% v/v) S. cerevisiae rekombinan kode CR2
Molases (c) 1.8 – 2.3 g/l (0.29% v/v)
S. cerevisiae yang diimobilisasi Typha latifolia(d) 28.5 g/l (3.59% v/v) Candida shehatae Daun tebu (e) 1.6 g/l (0.20% v/v) S. cerevisiae
Mahula(f) 33.99 g/l (4.27% v/v) S. cerevisiae strain CTCRI Onggok tapioka(g) 1.635% v/v Schizosaccharomyces sp.
(a) Yu et al. (1996). (b) Zaldivar et al. (2005).
(c) Gaptista et al. (2006). (d) Chandel et al. (2006). (e) Krishna et al. (1998). (f) Swain et al. (2007).
(g) Judoamidjojo et al. (1987).
2.4 Khamir
Khamir adalah mikroorganisme bersel tunggal, berbentuk oval tidak beraturan dan berukuran antara 5-20 μ (Paturau, 1981). Fungsi utama khamir dalam pembuatan etanol adalah mengubah gula dalam substrat menjadi etil alkohol dan CO2. Menurut Frazier (1977), enzim yang dihasilkan oleh khamir
adalah enzim invertase yang berfungsi sebagai pemecah sukrosa menjadi monosakarida (glukosa dan fruktosa) serta enzim zimase yang mengubah monosakarida tersebut menjadi etanol pada proses fermentasi.
Khamir termasuk kelompok kapang dan umumnya membelah diri dengan pertunasan. Tunas ini tumbuh sampai ukurannya mendekati ukuran induk, dan kemudian sel anak memisahkan diri. Pada kondisi tumbuh tertentu, tunas tidak memisahkan diri dari sel induk, sehingga suatu sel yang panjang dan bercabang terbentuk. Pada kondisi optimal khamir dapat membagi diri setelah 45 menit, tetapi kadang – kadang juga setelah 90 – 120 menit.
S. s cerevisiae merupakan “Top Yeast” di dunia. Hal ini dikarenakan, S. cerevisiae banyak digunakan dalam membantu kehidupan manusia baik untuk industri maupun rumah tangga. Penggunaan S. cerevisiae diantaranya untuk membuat beer atau minuman beralkohol maupun roti.
Produktivitas sel khamir merupakan fungsi dari konsentrasi glukosa, konsentrasi oksigen dan konsentrasi etanol (beberapa variabel kontrol juga berpengaruh seperti suhu dan pH). Glukosa merupakan reaktan dasar untuk metabolisme khamir. Menurut Judoamidjojo et al. (1989), pemecahan zat gula seperti glukosa oleh khamir dapat terjadi secara anaerobik (fermentasi) atau aerobik (respirasi). Tetapi proses yang lebih tipikal adalah pemecahan anaerobik yang dikenal juga sebagai fermentasi alkohol.
Pada kondisi aerobik, pemecahan gula mengikutsertakan oksigen atmosfir melalui beberapa lintasan proses. Pada respirasi oksidasi sempurna dari glukosa
menghasilkan CO2 dan air, sedang oksidasi tidak sempurna diikuti oleh akumulasi
asam dan lain-lain produk intermediet.
Khamir dapat tumbuh pada suhu yang cukup bervariasi dari 0 0C sampai 47
0C. Secara umum dianggap bahwa khamir dapat tumbuh dengan baik pada
suasana asam, yaitu pH 3.5 sampai 3.8 yang dapat menghambat sebagian bakteri. Toleransi asamnya adalah selang pH 2.2 sampai 8.0.Mekanisme pembentukan etanol oleh khamir melalui sebuah jalur Embden-Meyerhof-Parnas Pathway atau lebih dikenal dengan jalur glikolisis. Alur dari tahap glikolisis tersebut dapat dilihat pada Gambar 2.
Hasil dari glikolisis atau EMP pathway adalah memecah glukosa menjadi dua molekul piruvat. Proses yang terjadi dalam jalur glikolisis adalah sebagai berikut:
1. Langkah pertama pada tahap glikolisis adalah reaksi phosphorilasi dari glukosa menjadi glukosa-6-phosphat (G-6P) dengan bantuan enzim hexokinase; pada tahap ini, satu molekul ATP digunakan dan satu molekul ADP dihasilkan;
2. Selanjutnya glukosa-6-phosphat (G-6P) dikonversi menjadi fruktosa-6-phosphat (F-6P) dengan menggunakan enzim phosphoglukose isomerase; 3. Fruktosa-6-phosphat (F-6P) selanjutnya dikonversi menjadi fruktosa-1,6-bisphosphat oleh enzim phosphofructokinase; pada tahap ini pun satu molekul ATP digunakan dan satu molekul ADP dihasilkan;
4. Tahap selanjutnya adalah pemecahan fruktosa-1,6-bisphosphat oleh enzim aldolase menjadi dihidroxyacetone phosphat (DHAP) dan glyceraldehydes-3-phosphat (GA-3P); tahap ini merupakan salah satu tahap penting dalam proses glikolisis (dimana C6 dirubah menjadi 2C3);
DHAP dan GA-3P merupakan senyawa yang memiliki susunan molekul yang sama; GA-3P langsung digunakan dalam tahap selanjutnya glikolisis dan DHAP dikonversi menjadi GA-3P oleh enzim triose phosphat isomerase;
5. Glyceraldehydes-3-phosphat (GA-3P) dioksidasi dengan penambahan phosphat inorganic (Pi) menjadi 1,3-diphospho-glycerat (1,3-dP-GA) oleh enzim glyceraldehyde-3-phosphat dehydrogenase;
Glucose Hexokinase Glucose 6-phosphate ATP ADP Fructose 6-phosphate Phosphofructokinase aldolase ATP ADP Phosphoglucose isomerase Fructose 1,6-bisphosphate Dihydroxyacetone phosphate Glyceraldehhyde 3-phosphate (2 molecules) Triose phosphate isomerase 2 NAD++ 2P1 2 NADH + 2H+ O OH H H H O H OH H OH H CH2OH O OH H H H O H OH H OH H CH2-OPO3 2-O CH2OH OH OH H H OH CH2-OPO3 2-H O CH2-OPO3 2-OH OH H H OH CH2-OPO3 2-H C H C OH HC O H H OPO3 2-1,3-Bisphosphoglycerate (2 molecules) 2 ATP 2 ADP Glyceraldehide 3-phosphate dehydrogenase Phosphoglycerate kinase 3-Phosphoglycerate (2 molecules) 2-Phosphoglycerate (2 molecules) 2 H2O Phosphoenolpyruvate (2 molecules) 2 ATP 2 ADP Pyruvate (2 molecules) Phosphoglycero-mutase Enolase Pyruvate kinase C H C OH C O H H OPO3 2-2- O3PO C H C OH C O H H OPO3 2--O C H C 2- O3PO C O H H OPO3 2--O C H C 2- O3PO C O H H -O C O C C O H H H -O C O C C H OH H H OPO3 H-H
Gambar 2 Tahap glikolisis (Embden-Meyerhof-Parnas Pathway) (Crueger & Anneliese 1984).
CH3 C COOH O CH3 HC O acetaldehyde CH3 CH2OH CO2 ethanol pyruvate
+
Pyruvate decarboxylase NADH NAD+ Alcohol dehydrogenase6. 1,3-diphospho-glycerat melepaskan satu grup phosphat untuk membentuk ATP dari ADP dan kemudian dikonversi menjadi 3-phosphoglycerate (3P-GA) oleh enzim 3-phosphoglycerate kinase;
7. 3-phosphoglycerate (3P-GA) selanjutnya dikonversi menjadi 2- phosphoglycerate (2G-PA) oleh enzim phosphoglyceromutase;
8. Langkah selanjutnya adalah proses dehidrasi dari 2- phosphoglycerate (2G-PA) menjadi phosphemol pyruvate (PEP) oleh enzim enolase.
9. Langkah terakhir dari glikolisis adalah dephosphorelasi phosphemol pyruvate (PEP) menjadi pyruvat oleh enzim pyruvat kinase; pada tahap ini dibentuk sebuah molekul ATP.
Reaksi setelah pembentukan DHAP dan GA-3P selama proses glikolisis berlangsung sebanyak dua kali. Pyruvat yang merupakan produk akhir dari tahap glikolisis ini merupakan metabolit kunci pada proses metabolisme. Secara keseluruhan, reaksi dari proses glikolisis adalah sebagai berikut:
Setelah melalui tahap glikolisis, pyruvat yang terbentuk kemudian dirubah menjadi asetaldehide dan CO2 oleh enzim pyruvate decarboxylase, setelah itu oleh
enzim alcohol dehydrogenase, asetaldehide tersebut dirubah menjadi alkohol.
Gambar 3 Proses pembentukan etanol dari pyruvat.
Ln konsentrasi bi om assa Waktu Fase adaptasi Fase log/exponensial F as e per tum buhan l am
bat Fase stasioner
2.5 Kinetika Fermentasi
Pertumbuhan mikroorganisme secara batch yang ditumbuhkan pada medium tertentu memiliki kurva pertumbuhan seperti yang ditunjukkan oleh Gambar 4 berikut.
Gambar 4 Kurva pertumbuhan mikroorganisme pada kultur batch (Stanbury & Whitaker 1993).
Menurut Fardiaz (1988), beberapa fase pertumbuhan yang umumnya terjadi pada mikroorganisme secara umum adalah sebagai berikut:
a. Fase Adaptasi
Jika mikrrorganisme dipindahkan ke dalam suatu medium, mula-mula akan mengalami fase adaptasi untuk menyesuaikan dengan kondisi lingkungan di sekitarnya. Lama fase adaptasi ini dipengaruhi oleh beberapa faktor diantaranya medium dan lingkungan sekitar serta jumlah inokulum..
b. Fase Logaritmik/eksponensial
Pada fase ini mikroorganisme membelah dengan cepat dan konstan mengikuti kurva logaritmik. Pada fase ini kecepatan pertumbuhan sangat dipengaruhi oleh medium tempat umbuhnya seperti pH dan kandungan nutrient, juga kondisi lingkungan termasuk suhu dan kelembaban udara. Pada
x
t
d
x
d
μ
=
N
t
d
N
d
nμ
=
fase ini mikroorganisme membutuhkan energi lebih banyak daripada fase lainnya. Pada fase ini kultur paling sensitif terhadap keadaan lingkungan. c. Fase Pertumbuhan Lambat
Pada fase ini pertumbuhan populasi mikroorganisme diperlambat karena beberapa sebab: (1). zat-zat nutrisi dalam medium sudah sangat berkurang; (2). adanya hasil-hasil metabolisme yang mungkin beracun atau dapat menghambat pertumbuhan mikroorganisme. Pada fase ini jumlah populasi masih meningkat karena jumlah sel yang tumbuh masih lebih banyak daripada jumlah sel yang mati.
d. Fase Stasioner
Pada fase ini jumlah populasi sel tetap karena jumlah sel yang tumbuh sama dengan jumlah sel yang mati. Ukuran sel pada fase ini menjadi lebih kecil karena sel tetap membelah meskipun zat-zat nutrisi sudah habis. Karena kekurangan zat nutrisi, sel kemungkinan mempunyai komposisi berbeda dengan sel yang tumbuh pada fase logaritmik. Pada fase ini, sel lebih tahan terhadap keadaan ekstrim seperti panas, dingin, radiasi, dan bahan-bahan kimia.
Pertumbuhan mikroorganisme biasanya dicirikan dengan waktu yang dibutuhkan untuk menggandakan massa sel atau jumlah sel. Waktu ganda massa dapat berbeda dengan waktu ganda sel, karena massa sel dapat meningkat tanpa peningkatan dalam jumlah sel. Namun demikian bila dalam suatu lingkungan tertentu interval antara massa sel atau penggandaan jumlah sel adalah konstan dengan waktu, maka organisme itu tumbuh pada kecepatan eksponensial. Pada keadaan ini maka pertumbuhan dinyatakan sebagai:
…… (1) atau ……. (2)
dimana: x = konsentrasi sel (g/l)
N = konsentrasi sel dalam keseluruhan (sel/l)
t = waktu
μ = laju pertumbuhan spesifik untuk massa (jam-1) μn = laju pertumbuhan spesifik untuk jumlah (jam-1)
∫
∫
=
2 1 2 1 t t x xdt
x
x
d
μ
t
x
x
Ln
=
μ
Δ
1 2μ
μ
693
.
0
2
ln =
=
td
S
K
S
s maks+
=
μ
μ
Persamaan (1) menggambarkan peningkatan massa sel dan kesetimbangan, sedang persamaan (2) menggambarkan peningkatan jumlah sel dengan fungsi waktu. Pada umumnya pertumbuhan diukur dengan peningkatan massa sehingga μ dapat digunakan. Jika persamaan (1) diintegralkan maka akan menjadi:
Bila laju pertumbuhan spesifik tetap, maka menghasilkan
Untuk kasus dimana Δ t = td, maka
Dimana td adalah waktu ganda sel.
Menurut Stanbury & Whitaker (1993), selama fermentasi batch, laju pertumbuhan spesifik adalah konstan dan tidak tergantung pada perubahan konsentrasi nutriennya. Bentuk hubungan antara laju pertumbuhan dan konsentrasi substrat telah diteliti oleh Monod (1942), dengan persamaan sebagai berikut:
Dimana:
μ = laju pertumbuhan spesifik
μmaks = laju pertumbuhan spesifik yang maksimal
S = konsentrasi substrat
Ks = suatu konstanta yang sebanding pada konsentrasi substrat bila μ = ½
μmaks.
Pertumbuhan dan pembentukan produk oleh mikroorganisme merupakan proses – proses biokonversi. Dimana nutrien kimiawi yang diumpankan pada fermentasi dikonversi menjadi massa sel dan metabolit–metabolit. Setiap konversi dapat dikuantifikasikan oleh suatu koefisien hasil yang dinyatakan sebagai massa
S
x
Y
s xΔ
Δ
=
S
p
Y
s pΔ
Δ
=
waktu x atau p waktu x atau p waktu x atau psel atau produk yang terbentuk per unit massa nutrien yang dikonsumsi, yaitu Yx/s
untuk sel dan Yp/s untuk produk yang dihitung sebagai:
Dimana:
Yx/s = rendemen pembentukan sel oleh substrat Yp/s = rendemen pembentukan produk oleh substrat
Menurut Wang et al.(1978), pertumbuhan juga dapat dihubungkan dengan konsumsi nutrien selain dihubungkan dengan pembentukan produk. Pembentukan produk tidak mungkin terjadi tanpa adanya sel. Oleh karena itu diharapkan pertumbuhan dan pembetukan produk terpadankan dengan erat dengan penggunaan nutrien, dan ini tergantung pada kontrol pengaturan metaboliknya.
Hubungan kinetika di antara pertumbuhan dan pembentukan produk tergantung pada peranan produk dalam metabolisme sel. Dua buah kinetika yang umum digunakan adalah kinetika yang menggambarkan sintesis produk selama pertumbuhan dan kinetika yang menggambarkan sintesis produk setelah pertumbuhan terhenti. Suatu pola lain yang jarang diterapkan adalah bila pertumbuhan terjadi tanpa adanya pembentukan produk, tetapi setelah melalui selang waktu tertentu produk mulai muncul dan pertumbuhan terus berlangsung. Kinetika pola ini diperlihatkan pada Gambar 5 berikut.
(a) (b) (c)
Gambar 5 Pola kinetika pertumbuhan dan pembetukan produk pada fermentasi batch: (a) pertumbuhan – berasosiasi dengan pembentukan produk; (b) campuran pertumbuhan – berasosiasi dengan pembentukan produk; (c) bukan (pertumbuhan terhenti) pertumbuhan – berasosiasi dengan pembentukan produk (Wang et al. 1978).