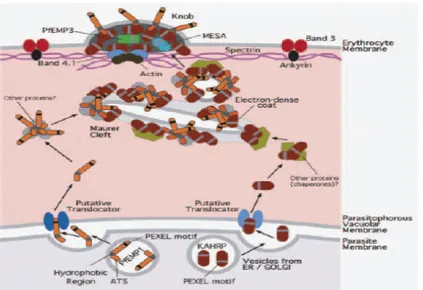
INTERVENSI CYTOADHERENCE SEBAGAI PELUANG UNTUK PENCEGAHAN DAN TERAPI MALARIA BERAT
Teks penuh
Gambar


Dokumen terkait
Panduan penggunamu.. Jika tampilan error selain yang dijelaskan di atas muncul, tekan tombol Reset*. Jika masalah tetap ada, matikan dan konsultasikan pada toko tempat Anda membeli.
Sebagai salah satu alternatif supaya proses pembelajaran dapat memberikan pengalaman langsung kepada siswa, melatih kemampuan berpikir tingkat tinggi dan dapat
Program Nasional Pemberdayaan Masyarakat Mandiri (PNPM Mandiri) dilakukan melalui berbagai kegiatan yang dilakukan oleh pengelola Program Nasional Pemberdayaan
Tingkat partisipasi angkatan kerja (usia 15-64 tahun) adalah proporsi penduduk usia kerja yang termasuk ke dalam angkatan.. kerja, yakni mereka yang bekerja dan mencari
Pada bagian ruang lingkup unit kerja, penulis akan menjelaskan tentang ruang lingkup pekerjaan yang menjadi tanggung jawab bagian processing di PT. Memproses dan
Dengan spesifikasi kepada sejarah lahirnya nasionalisme dari kalangan etnis Arab di Indonesia dan peran pan islamisme serta gerakan gerakan islam.. Selain itu, penulis juga
Sistem ini dibuat untuk memberikan informasi berupa SMS kepada BNPB (Badan Nasional Penanggulangan Bencana), Kepala Desa dan Tokoh Masyarakat mengenai pergerakan
Selain itu, penelitian Odewabi di Nigeria tahun 2014 menyatakan bahwa paparan radikal bebas pada pekerja SPBU meningkatkan kadar MDA dan menurunkan kadar antioksidan
