Chemical Composition, Yield and Yield Components of Two Wheat
Cultivars in Response to Salt Stress
H Sadeghi1*and Y Emam2
Received: 15 July 2010 Accepted: 5 January 2011
1Assistant Professor of College of Agriculture, Shiraz University, Shiraz, Iran 2 Professor of College of Agriculture, Shiraz University, Shiraz, Iran
* Coresponding author : E-mail [email protected]
Komposisi Kimia, Hasil Panen dan Komponen Hasil Panen dari Dua
Gandum Kultivar dalam Responnya Terhadap Salt Stress
H Sadeghi1*and Y Emam2
ABSTRAK
Di kebanyakan provinsi di paling selatan Iran, salinitas tanah adalah masalah yang berkembang, khususnya di irigasi daerah pertanian, dan telah ditemukan bahwa hal ini mengurangi hasil panen gandum secara dramatis. Untuk meneliti efek dari Sodium Klorida pada dua gandum (Triticum aestivum L.) kultivar, empat tingkatan salinitas: 0, 4, 8, dan 12 dS / m, telah digunakan sebagai percobaan faktorial yang disusun dalam randomized complete block design dengan empat pengulangan dalam lingkungan yang terkendali di rumah kaca selama 2006-2007. Hasil penelitian menunjukkan bahwa peningkatan salinitas dari 0 sampai 12 dS / m, menurunkan persentase ketahanan hidup tanaman secara signifikan. Dua kultivar dari Kavir dan Shiraz memberi respon yang berbeda terhadap salinitas, secara signifikan Kavir menunjukkan tingkat hasil panennya lebih tinggi. Kultivar ini juga memiliki konten kalium lebih besar. Jumlah anakan dan daun pada tiap tanaman dan, juga, tinggi tanaman mengalami penurunan selama meningkatnya tingkat salinitas. Kandungan natrium itu, juga meningkat seiring dengan meningkatnya tingkat salinitas di kedua kultivar. Namun, natrium di dalam Kavir dibandingkan dengan Shiraz, lebih rendah, mungkin dikarenakan mekanisme pengecualian Na + dalam kultivar ini. Jumlah butir gandum tertinggi dan phytomass diperoleh dari Kavir di tingkat salinitas terendah. Phytomass dan hasil gabah, juga menurun secara signifikan sebagai akibat dari salinitas. Efek samping dari kurangnya salinitas di Kavir menunjukkan bahwa kultivar ini mungkin cocok untuk tanah yang mengandung kadar garam, sebuah objek yang perlu diteliti lebih lanjut.
Pengantar
Di kebanyakan provinsi di paling selatan Iran, salinitas adalah masalah yang berkembang terutama di irigasi daerah pertanian dengan meningkatnya permukaan air tanah, rendahnya kualitas air dan/ atau kurang baiknya kemampuan tanah untuk dilewati oleh air. Salinitas tanah telah mengurangi hasil panen gandum biasanya ketika nilai konduktivitas listrik di atas 6 dS / m diseluruh zona akar (Munns et al. 2006).
Salt stress adalah salah satu tekanan abiotik yang paling penting yang mempengaruhi produktivitas alami dan menyebabkan kerugian tanaman yang signifikan di seluruh dunia. Untuk tanaman, ion natrium (Na+) berbahaya, sedangkan ion kalium (K+) adalah ion penting. Sitosol pada sel tumbuhan normal mengandung 100-200 mM dari K+ dan 1-yang di dominasi oleh lingkungan garam di alam, akumulasi dari tingginya Na+ di sitosol, dan tingginya rasio Na+/ K+, mengganggu fungsi enzimatik yang biasanya diaktifkan oleh K+ di sel (Bhandal dan Malik 1988, Tester dan Davenport 2003, Munns et al. 2006). Oleh karena itu, sangat penting bagi sel untuk mempertahankan konsentrasi rendah sitosol Na+ atau untuk mempertahankan rasio rendah Na+/ K+ di dalam sitosol pada saat NaCl stress (Maathuis dan Amtmann, 1999).
Pada gandum, telah menunjukkan bahwa kedua tanggapan terjadi secara berurutan, sehingga menimbulkan respon pertumbuhan dua-tahap untuk salinitas
(Munns, 1993). Sebagai contoh, memiliki penurunan pertumbuhan yang sama untuk empat minggu pertama di 150 mM NaCl, dan itu tidak sampai perbedaan pertumbuhan antara genotipe jelas diamati (Munns et al. 1995). Namun, dalam dua minggu, daun-daun kering terlihat pada genotipe yang lebih sensitif dan tingkat kematian daun-daun tua jelas lebih besar pada genotipe yang lebih sensitif daripada genotipe toleran. Pernah terjadi, jumlah daun-daun kering meningkat sekitar di atas 20% dari jumlah total, pertumbuhan jauh lebih rendah daripada tanaman salt-sensitive. Di bidang pertanian lahan kering, meningkatkan toleransi garam dapat meningkatkan hasil panen gandum di tanah yang mengandung kadar garam.
penyakit dan kekurangan gizi); salinitas pada lapisan tanah tetap menjadi penghalang utama untuk pertanian di semua daerah semi-kering seperti kebanyakan di provinsi paling selatan Iran. Walaupun ketika pembukaan lahan di zona dengan curah hujan yang lebih tinggi telah menyebabkan bergeraknya permukaan air tanah dan garam, tetapi memungkinkan adanya peningkatan toleransi terhadap garam pada tanaman. Pengenalan pada 'deep-rooted perennial species' diperlukan untuk menurunkan permukaan air tanah, namun, toleransi garam akan diperlukan tidak hanya untuk 'de-watering' spesies, tetapi juga untuk tanaman tahunan, garam akan ditinggalkan dalam tanah ketika permukaan air tanah diturunkan (Francois
et al. 1994).
Gandum merupakan tanaman yang cukup toleran terhadap garam (Maas dan Hoffman 1977). Salah satu dari dua kultivar baru gandum, digunakan pada penelitian ini, Kavir, merupakan genotipe yang telah ditingkatkan yang direkomendasikan untuk daerah yang memiliki kadar garam seperti di provinsi paling selatan Iran, Namun, mekanisme NaCl pada komposisi kimia organ tanaman juga diteliti.
Bahan dan Metode
Lokasi, penerapan perlakuan dan pengumpulan data
Penelitian ini dilakukan untuk mengevaluasi efek dari empat tingkat salinitas (0,4, 8 dan 12 dS / m) pada dua kultivar gandum (Kavir, genotipe toleran garam dan Shiraz, kultivar sensitif garam). Salinitas yang ingin dikembangkan dengan mencampur jumlah NaCl dan CaCl2 (5: 1) yang diperlukan di tanah sebelum mengisi pot (0, 2.16, 4.32, 8.64 g/ kg tanah). Tanaman gandum ditaburkan pada 17 November 2006 dan dipanen pada 29 April 2007. Penelitian dilakukan dalam rumah kaca di Fakultas Pertanian, Shiraz University, Shiraz, Iran (52o 46'E, 29o
per tanaman dan penambahan panjang telah dicatat (Wilhelm et al. 1989).
Pengukuran Natrium dan Kalium
Sampel kering telah digiling menjadi bubuk dan sekitar 0.1 g dipindahkan ke tabung reaksi yang berisi 10 mL 0.1 N asam asetat, dan dipanaskan dalam water bath pada 80 ° C selama 2 ditentukan menggunakan serapan atom spektrometer (Munns dan James 2003).
Pengukuran Prolin
Lembaran jaringan daun segar (0.5 g) dihaluskan dalam nitrogen cair dan kemudian diekstraksi dalam 20 ml air panas selama 30 menit dengan moderat gemetar (moderate Shaking). Homogenat disentrifugasi pada 5000 g selama 10 menit. Konsentrasi prolin dihitung dengan aplikasi dari metode reagen asam ninhidrin (of the ninhydrin acid reagent method) Design Model menggunakan software
SAS (SAS Institute 1985). Berarti dibandingkan dengan uji jarak berganda Duncan (Duncan's multiple range test) pada p ≤ 0,05.
Hasil dan Pembahasan
Pengaruh natrium klorida terhadap pertumbuhan dan karakteristik morfologi
dan sub-seluler yang ada. Pemisah utama ion beracun oleh tanaman dengan toleransi salinitas, mungkin di vakuola, mungkin telah memungkinkan untuk mengatur metabolik sitoplasmiknya di kondisi stabil dan lebih mendekati normal daripada tanaman dengan sensitifitas salinitas tinggi. Oleh karena itu, ukuran yang benar dari sitoplasma tanaman dengan toleransi garam mungkin diperlukan. Fase pertama dari hasil respon pertumbuhan dari efek garam di luar tanaman contohnya garam
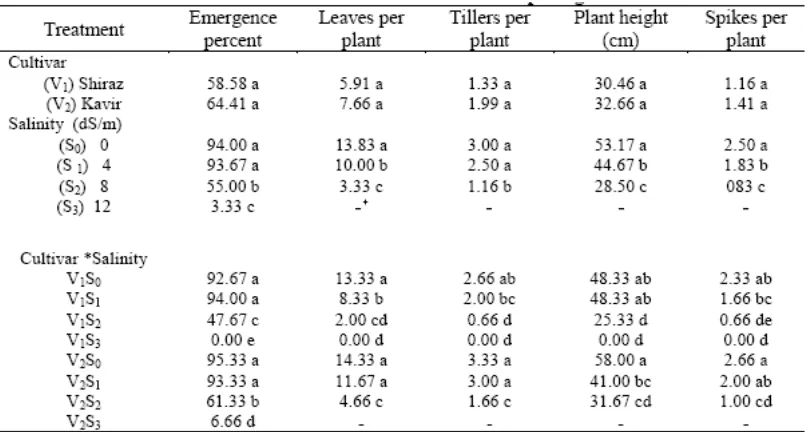
dalam larutan tanah (tekanan osmotik) mengurangi pertumbuhan daun seperti yang ditunjukkan pada Tabel 2. Memang, garam sendiri tidak banyak di jaringan yang sedang tumbuh pada konsentrasi dimana pertumbuhan terhambat, pemanjangan sel-sel dengan cepat dapat menampung garam yang tiba di xilem sampai mereka mencpai vakuola. Dengan demikian, garam diambil oleh tanaman tidak langsung menghambat pertumbuhan daun baru (Munns 1993).
Tahap kedua dari hasil respon pertumbuhan dari efek toksik garam di dalam tanaman. Garam diambil oleh konsentrat tanaman pada daun tua. Kelanjutan transportasi dari garam ke daun yang sedang melakukan transpirasi melalui jangka waktu yang panjang, akhirnya hasil di kondisi dengan konsentrasi Na+ dan Cl -yang sangat tinggi, dan daun mati seperti yang diamati pada percobaan kami (lihat Tabel 2 dan 4). Hal ini mungkin
bukti untuk ini dalam kultivar jagung yang berbeda dalam toleransi garamnya.
Hubungan Antara Salinitas dan Komponen Hasil Panen
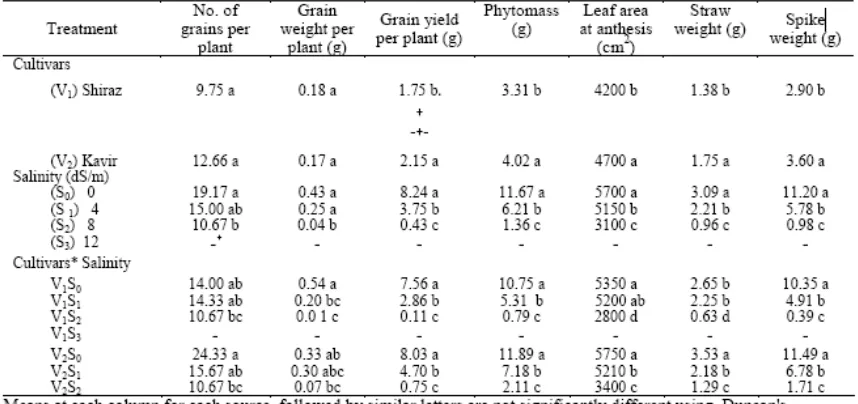
Hasil penelitian menunjukkan bahwa jumlah gandum tertinggi dan phytomass diperoleh dari Kavir pada tingkat salinitas terendah (Tabel 3). Phytomass dan hasil gabah juga menurun pada salinitas secara signifikan. Hasil Penurunan ini disebabkan, terutama untuk mengurangi berat seluruh bulir dan berat biji itu sendiri daripada jumlah seluruh bulir (Tabel 3). Temuan ini menegaskan hasil Francois et al. (1989). Hasil jerami
lebih sensitif terhadap salinitas dibandingkan hasil gabah (Tabel 3).
Hasil penelitian kami juga menunjukkan bahwa perkiraan hasil gabah mungkin menyebabkan kompleksitas lain terhadap respon salinitas, bukan hanya karena tanaman harus ditumbuhkan dalam lingkungan terkendali untuk jangka waktu yang lama, tetapi juga karena kompleksitas biomassa pucuk untuk mengkonversi menjadi gandum. Tingkat salinitas rendah tidak mungkin mengurangi berat gabah meskipun luas daun dan phytomass berkurang (Tabel 3), faktanya adalah hasil gabah tidak menurun sampai salinitas yang diberikan ('Threshold') tercapai (Maas dan Hoffman 1977).
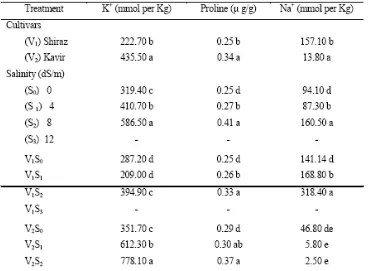
Pengaruh Natrium Klorida pada Komposisi Kimia
Hasil penelitian kami menunjukkan bahwa Kavir memiliki kandungan kalium lebih besar (Tabel 4). Konsentrasi natrium juga meningkat dengan jalan meningkatkan tingkat salinitas di kedua
pada fenomena fisiologis ini disebabkan perubahan kandungan ionik pada tanaman (Abdullah et al. 1978). Pendekatan lain untuk meningkatkan toleransi tanaman terhadap garam dalam gandum didasarkan pada mekanisme toleransi garam, menggunakan ciri-ciri fisiologis untuk memilih plasma nutfah. Dalam gandum, toleransi garam terkait dengan rendahnya tingkat transportasi Na+ ke tunas, dengan selektivitas tinggi untuk K+ dan Na+ (Chhipa dan Lal 1995, Ashraf dan O'Leary 1996, Ashraf dan Khanum 1997), meskipun hubungan ini tidak mencakup semua genotipe (Ashraf dan McNeilly 1988, El-Hendawy et al. 2005), menunjukkan bahwa mekanisme pengecualian Na+ bukan satu-satunya mekanisme toleransi garam (Colmer et al.,
2006). Secara umum, Kavir, yang ditandai dengan konsentrasi Na+ terendah, menghasilkan bahan kering yang lebih besar daripada kultivar Shiraz (Tabel 4). Genotipe Na+ rendah ini memiliki daun terluka yang
lebih sedikit, dan proporsi yang lebih besar dari daun hidup hingga daun-daun kering, seperti diamati selama percobaan. Efek pada pertumbuhan mungkin dikarenakan keseimbangan karbon yang lebih baik dalam genotipe Na+ rendah ini. Hubungan yang sama antara bahan kering pada tunas dan Na+ daun ditemukan pada populasi dari penyilangan antara genotipe Na+ tinggi dengan genotip rendah Na+ (Munns dan James 2003).
Gambar 1. Hubungan antara toleransi salinitas (% pertumbuhan dari kontrol) dan konsentrasi Na+ daun di kultivar Kavir. Konsentrasi Na+ diukur pada daun ketiga setelah 10 hari di 150
mM NaCl dan biomassa kecambah setelah 24 hari. Nilai dinyatakan sebagai persentase dari biomassa kecambah dalam kondisi control (R2 = 0,8252). Semua nilai-nilai didasarkan pada rata-rata (n = 5).
Kesimpulan
Hasil penelitian kami menunjukkan bahwa kedua kultivar, Kavir & Shiraz, memiliki respon yang berbeda terhadap salinitas, pada Kavir menunjukkan secara signifikan tingkat kemampuan hidup yang lebih tinggi. Kultivar ini (Kavir) juga memiliki kandungan kalium yang lebih besar. Jumlah anakan dan daun per tanaman dan juga tinggi tanaman menurun pada kedua kultivar setelah meningkatnya salinitas. Kandungan natrium dalam kedua kultivar juga meningkat denngan meningkatnya tingkat salinitas; Namun, kandungan natrium pada Kavir,
dibandingkan dengan Shiraz, mungkin lebih rendah dikarenakan mekanisme pengecualian Na+ pada kultivar ini. Hasil penelitian juga mengungkapkan bahwa jumlah gabah tertinggi dan phytomass diperoleh dari Kavir ditingkat salinitas terendah. Phytomass dan hasil gabah itu, juga menurun pada salinitas secara signifikan. Secara keseluruhan, tampak bahwa sedikitnya dampak buruk dari salinitas di kultivar Kavir ini membuatnya lebih cocok untuk dapat tumbuh di tanah yang mengandung kadar garam. Penelitian ini masih membutuhkan penyelidikan lebih lanjut.
Referensi
Abdullah Z, Ahmad R and Ahmad J, 1978. Salinity induced changes in the reproductive physiology of wheat plants. Plant and Cell Physiology 19: 99-106.
Ashraf M and Khanum A, 1997. Relationship between ion accumulation and growth in two spring wheat lines differing in salt tolerance at different growth stages. Journal of Agronomy and Crop Science 178: 39–51.
Ashraf M and McNeilly T, 1988. Variability in salt tolerance of nine spring wheat cultivars. Journal of Agronomy and Crop Science 160: 14–21.
Ashraf M and O'Leary JW, 1996. Responses of some newly developed salt-tolerant genotypes of spring wheat to salt stress. 1. Yield components and ion distribution. Journal of Agronomy and Crop Science176: 91–101.
Bates LS, Waldren RP and Teare ID, 1973. Rapid determination of free proline for water-stress studies. Plant Soil 39: 205-207.
Bhandal IS and Malik CP, 1988. Potassium estimation, uptake, and its role in the physiology and metabolism of flowering plants. International Review of Cytology 110: 205–254. Chhipa BR and Lal P, 1995. Na/K ratios as the basis of salt tolerance in wheat. Australian
Journal of Agricultural Research 46: 533–539.
Colmer TD, Epstein E and Dvorak J, 1995. Differential solute regulation in leaf blades of various ages in saltsensitive wheat and salt tolerant wheat x Lophopyrum elongatum
(Host) A. Löve amphiploid. Plant Physiology 108: 1715–1724.
Colmer TD, Flowers TJ and Munns R, 2006. Use of wild relatives to improve salt tolerance in wheat. Journal of Experimental Botany 57: 1059–1078.
El-Hendawy SE, Hu Y and Schmidhalter U, 2005. Growth, ion content, gas exchange, and water relations of wheat genotypes differing in salt tolerances. Australian Journal of Agricultural Research 56: 123–134.
Flowers TJ, Hajibagheri MA and Yeo AR, 1991. Ion accumulation in the cell walls of rice plants growing under saline conditions: evidence for the Oertli hypothesis. Plant, Cell and Environment 14: 319–325.
Francois LE, Donovan TJ, Lorenz K and Maas EV, 1989. Salinity effects on rye grain yield, quality, vegetative growth, and emergence. Agronomy Journal 81: 707-712.
Francois LE, Grieve CM, Maas EV, Donovan TJ and Lesch SM, 1994. Time of salt stress affects growth and yield components of irrigated wheat. Agronomy Journal 86: 100– 107.
Gorham J, Hardy C, Wyn Jones RG, Joppa LR and Law CN, 1987. Chromosomal location of a K/Na discrimination character in the D-genome of wheat. Theoretical and Applied Genetics 74: 584–588.
Gorham J, Wyn Jones RG and Bristol A, 1990. Partial characterization of the trait for enhanced K+-Na+ discrimination in the D-genome of wheat. Planta 180: 590–597. Kingsbury Ralph W, Epstein E and Pearcy W, 1984. Physiological responses to salinity in
selected lines of wheat. Plant Physiology 74: 417-423.
Läuchli A, 1984. Salt exclusion: an adaptation of legumes for crops and pastures under saline conditions. In: Staples RC (Ed). Salinity Tolerance in Plants: Strategies for Crop Improvement. Wiley, New York, pp. 171–187.
Maas EV and Hoffman GJ, 1977. Crop salt tolerance – current assessment. Journal of the Irrigation and Drainage Division of the American Society of Civil Engineering 103: 115–134.
Maathuis FJM and Amtmann A, 1999. K+ nutrition and Na+ toxicity the basis of cellular K+/ Na+ ratios. Annals of Botany 84: 123–133.
Moradi F and Ismail AM, 2007. Responses of photosynthesis, chlorophyll fluorescence and ROS-scavenging systems to salt stress during seedling and reproductive stages in rice. Annals of Botany 99: 1161-1173.
Mühling KH and Läuchli A, 2002. Effect of salt stress on growth and cation compartmentation in leaves of two plant species differing in salt tolerance. Journal of Plant Physiology 159: 137–146.
Munns R, 1993. Physiological processes limiting plant growth in saline soil: some dogmas and hypotheses. Plant, Cell and Environment 16: 15–24.
Munns R and James RA, 2003. Screening methods for salt tolerance: a case study with tetraploid wheat. Plant and Soil 253: 201–218.
Munns R, James AJ and Läuchli A, 2006. Approaches to increasing the salt tolerance of wheat and other cereals. Journal of Experimental Botany 57: 1025–1043.
Munns R, Schachtman DP and Condon AG, 1995. The significance of a two-phase growth response to salinity in wheat and barley. Australian Journal of Plant Physiology 22: 561–569.
SAS Institute, 1985. SAS user’s guide . Statistics. Version 5. SAS Inst., Cary, NC, USA. Schachtman DP, Munns R and Whitecross MI, 1991. Variation of sodium exclusion and salt
tolerance in Triticum tauschii. Crop Science 31: 992–997.
Taiz L. and Zeiger E, 2002. Plant Physiology. Sinauer Associates Inc., Sunderland, Massachusetts.
Tester M and Davenport R, 2003. Na+ tolerance and Na+ transport in higher plants. Annals of Botany 91: 503–527.
Wilhelm WW, Bouzerzour H and Power JF, 1989. Soil disturbance-residue management effect on winter wheat