koloni secara mikroskopis serta pigmentasinya.
Penapisan aktivitas lipase dari bakteri endofit. Bakteri non aktinomiset endofit murni hasil isolasi dari daun, batang dan akar jati belanda, jambu biji serta rimpang temulawak diambil dengan sedotan steril berdiameter 0,8 cm sebanyak 3 bulatan dan diinokulasikan pada media TGY yang mengandung (per liter) 5 g tryptone, 5 g ekstrak khamir, 1 g dekstrosa dan 1 g K2HPO4 dengan pH 7.2, sedangkan aktinomiset endofit murni diinokulasikan pada media ISP2. Masing-masing media tersebut mengandung 3% minyak zaitun dan 2 mL 0,1% pewarna Rhodamine B yang telah disterilisasi meng-gunakan milipore (Swinnex) 0,2 µm, kemudian isolat bakteri diinkubasi pada suhu 30 0C selama 48 jam sedangkan isolat aktinomiset diinkubasi pada suhu 25 0C selama 120 jam. Aktivitas lipase pada media penapisan dapat dideteksi di bawah sinar UV sebagai zona berpendar berwarna oranye.
Penentuan kurva tumbuh dan produksi enzim. Isolat terpilih yang memiliki aktivitas lipase terbesar diinokulasikan pada 50 mL media tryptone soy broth (TSB) lalu diinkubasi pada suhu 50 0C selama 24 jam. Biakan kemudian diinokulasikan sebanyak 50 mL pada media produksi yang mengandung (per 100 mL) 1% minyak zaitun, 0,02% CaCl2. 2H2O, 0,01% MgSO4. 7H2O, dan 0,04% FeCl3. 6H2O lalu diinkubasi menggunakan inkubator bergoyang pada suhu 50 0C dengan kecepatan 150 rpm. Setiap 12 jam dilakukan pengambilan kultur sel bakteri non aktinomiset untuk diukur densitas selnya pada panjang gelombang 620 nm (Lampiran 3) yang berlangsung selama 96 jam, sedangkan kultur sel aktinomiset diambil setiap 24 jam sekali selama 240 jam. Kultur sel tersebut kemudian disentrifugasi selama 10 menit pada suhu 4 0C dengan kecepatan 10000 rpm. Supernatan yang diperoleh merupakan enzim ekstrak kasar yang selanjutnya diukur aktivitas lipasenya
Pengukuran aktivitas lipase dan kadar protein. Enzim ekstrak kasar yang diperoleh diukur aktivitas lipasenya dengan menggunakan metode menurut Kwon dan Rhee (1986). Satu mL enzim ekstrak kasar dicampurkan ke dalam larutan yang mengandung 2,5 mL minyak zaitun + 0,005 M buffer fosfat (1:1) dan 20 µL CaCl2, kemudian diinkubasi menggunakan inkubator bergoyang
dengan kecepatan 200 rpm pada suhu 60 0C selama 30 menit. Campuran ditambah 1 mL 6 N HCl dan 5 mL larutan isooktan lalu dihomogenasi menggunakan vortex selama 1 menit. Empat mL lapisan atas isooktan kemudian dipindahkan ke dalam tabung yang baru, ditambah dengan 1 mL reagen cupric
acetate-pyridine (Lampiran 4) dan
dihomogenasi menggunakan vortex selama 1 menit. Selanjutnya campuran diukur dengan spektrofotometer pada absorbansi 715 nm. Satu unit aktivitas lipase didefinisikan sebagai jumlah enzim yang membebaskan 1 µmol asam lemak per mL/menit pada suhu 60 0C. Pengukuran aktivitas lipase dilakukan dengan meng-gunakan kurva standar asam margarat (asam heptadekanoat) yang diukur absorbansinya pada 715 nm (Lampiran 5).
Asam margarat adalah salah satu jenis asam lemak dan merupakan monomer penyusun trigliserida yang dihasilkan dari hidrolisis trigliserida oleh lipase. Asam margarat ini kemudian juga direaksikan dengan reagen cupric-acetate pyridine. Besarnya aktivitas lipase ditandai dengan banyaknya asam lemak yang dibebaskan per menit, sehingga penggunaan asam margarat pada penelitian ini bertujuan untuk mendekati jumlah asam lemak yang dibebaskan dari hidrolisis trigliserida (minyak zaitun) oleh enzim ekstrak kasar (sampel). Kadar protein diukur berdasarkan metode Bradford (1976) dengan menggunakan standar bovine serum albumin (BSA) (Lampiran 6).
Pengukuran aktivitas spesifik lipase. Aktivitas spesifik lipase diukur dengan menggunakan rumus sebagai berikut; Aktifitas spesifik lipase (unit/mg) = aktivitas lipase (unit/mL) / kadar protein (mL/mg). Aktivitas spesifik lipase yang diperoleh kemudian disetarakan dengan konsentrasi (ppm) lipase tipe VII sebagai kontrol positifnya (Lampiran 7).
HASIL
Isolasi bakteri endofit. Isolasi bakteri endofit dari daun, batang dan akar jati belanda, jambu biji, serta rimpang temulawak pada media HV agar berhasil memperoleh 69 isolat bakteri endofit, 55 diantaranya dimasukkan ke dalam golongan non aktinomiset sedangkan 14 diantaranya dimasukkan ke dalam golongan aktinomiset. Isolasi dari tanaman jati belanda diperoleh 22 isolat, dan dari tanaman jambu biji diperoleh
AJB 4(4)
C
27 isolat, sedangkan dari rimpang temulawak diperoleh 20 isolat. Keenam puluh sembilan isolat tersebut memiliki karakteristik morfologi koloni dan pigmentasi yang sangat beragam (Lampiran 8). Isolasi aktinomiset pada media HV agar memperlihatkan adanya per-tumbuhan aktinomiset endofit pada media tersebut (Gambar 1). Selain itu, isolat aktinomiset yang berhasil dipurifikasi pada media ISP2 juga menunjukkan keragaman yang tinggi dalam morfologi koloni dan pigmentasinya (Gambar 2). Pengamatan mikroskopis juga dilakukan terhadap morfologi rantai spora koloni aktinomiset endofit baik yang diduga termasuk ke dalam genus Streptomyces maupun non Streptomyces.Isolat AJB 4(3), AJB 4(5), AJB 4(4), AJB 4(1), dan APG 5(4) merupakan isolat aktinomiset endofit yang termasuk ke dalam genus Streptomyces karena mampu membentuk rantai spora dengan morfologi yang beragam (seperti kait hingga spiral), sedangkan CX 12(1) merupakan isolat aktinomiset endofit yang termasuk ke dalam genus non Streptomyces karena berdasarkan pengamatan mikroskopis diketahui tidak membentuk rantai spora (Gambar 3). Dua puluh tujuh dari 55 isolat bakteri non-aktinomiset endofit terbukti mampu mem-bentuk endospora di dalam sel vegetatifnya seperti isolat AJB 3 dan DPG 3(2) (Gambar 4).
Gambar 1 Koloni aktinomiset yang tumbuh pada media
HV agar.
Gambar 2 Keragaman morfologi koloni dan pigmentasi aktinomiset endofit pada media ISP2.
Gambar 3 Pengamatan mikroskopis (400 x) morfologi
rantai spora Streptomyces; AJB 4(3) (a), AJB
4(5) (b), AJB 4(4) (c), AJB 4(1) (d), APG 5(4)
(e), dan non Streptomyces CX 12(1) (f).
Gambar 4 Endospora yang dibentuk di dalam sel vegetatif isolat AJB 3 (a) dan DPG 3(2) (b).
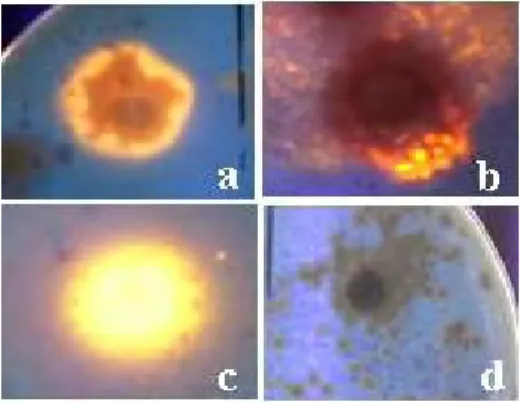
Penapisan aktivitas lipase dari bakteri endofit. Hasil penapisan aktivitas lipase terhadap 69 isolat bakteri endofit pada media agar Rhodamine B menunjukkan delapan isolat bakteri endofit memiliki aktivitas lipase. Hal ini terlihat dari kedelapan isolat endofit mampu membentuk pendaran berwarna oranye dengan kemampuan yang berbeda-beda. Dua isolat diantaranya yaitu DPG 3(2) dan AJB 4(4) memiliki kemampuan tertinggi diantara keenam isolat lainnya (Gambar 5). Isolat DPG 3(2) merupakan bakteri dari golongan non-aktinomiset yang diisolasi dari daun tanaman jambu biji, sedangkan isolat AJB 4(4) merupakan bakteri dari golongan aktinomiset yang diisolasi dari akar tanaman jati belanda. Kedua isolat ini kemudian dipilih untuk diuji aktivitas lipasenya.
Gambar 5 Isolat DPG 3(2) (a), AJB 4(4) (b) dan lipase tipe VII (1000 ppm) (c) memperlihatkan aktivitas lipase, sedangkan AJB 4(3) (d) tidak memperlihatkan aktivitas lipase pada media
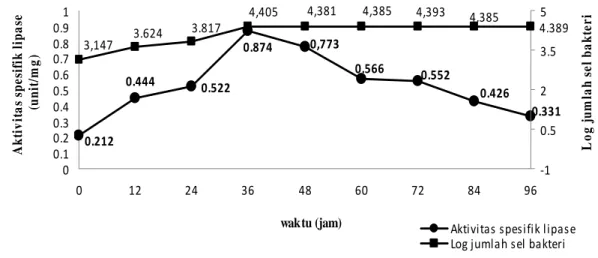
Kurva tumbuh dan uji aktivitas enzim. Pengukuran pertumbuhan dan uji aktivitas lipase isolat DPG 3(2) dilakukan setiap 12 jam sekali selama 96 jam. Untuk isolat AJB 4(4) uji aktivitas lipase dilakukan setiap 24 jam sekali selama 240 jam. Jumlah sel tertinggi dan aktivitas spesifik lipase tertinggi isolat DPG 3(2) terlihat pada jam ke-36 yaitu sebesar 4,405 log jumlah sel dan 0,874 unit per mg (Gambar 6). Jumlah sel terlihat mulai menurun pada jam ke-48 yaitu sebesar 4,381 log jumlah sel, begitu juga dengan aktivitas spesifik yang terlihat menurun pada jam ke-48 sebesar 0,773 unit per mg. Aktivitas spesifik lipase tertinggi isolat AJB 4(4)
terlihat pada jam ke-72 yaitu sebesar 1,139 unit per mg, pada jam ke-96 aktivitas spesifik lipase terlihat mulai menurun sebesar 0,734 unit per mg (Gambar 7). Aktivitas spesifik lipase isolat DPG 3(2) dan AJB 4(4) ini kemudian disetarakan dengan konsentrasi kontrol positifnya yaitu lipase tipe VII (Sigma chemical co). Berdasarkan data pada Tabel 1 dan 2, diketahui bahwa aktivitas spesifik lipase tertinggi isolat DPG 3(2) setara dengan konsentrasi lipase tipe VII 10, 172 ppm, sedangkan aktivitas spesifik lipase tertinggi isolat AJB 4(4) setara dengan konsentrasi lipase tipe VII 13, 260 ppm.
0.522 0.331 0.426 0.552 0.566 0,773 0.874 0.444 0.212 4.389 4.385 4,393 4,385 4,381 4,405 3.817 3.624 3,147 0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1 0 12 24 36 48 60 72 84 96 waktu (jam) A k ti v it a s spe si fi k l ipa se (u n it /m g ) ‐1 0.5 2 3.5 5 L og j u m lah s e l b ak te r i
Aktivitas spesifik lipase Log jumlah sel bakteri
Gambar 6 Pertumbuhan dan aktivitas spesifik isolat DPG 3(2) pada media yang mengandung minyak zaitun 1%.
0 0 0.935 1.139 0.734 0.632 0.651 0.637 0.568 0.431 0.414 0 0.2 0.4 0.6 0.8 1 1.2 0 24 48 72 96 120 144 168 192 216 240 Waktu (jam) A k ti v it a s s p es if ik li p a se (uni t/ m g )
Tabel 1 Kesetaraan aktivitas spesifik lipase isolat DPG 3(2) dengan konsentrasi lipase tipe VII Waktu (jam) Aktivitas spesifik lipase (unit/mg) Konsentrasi lipase tipe VII (ppm) 0 0,212 2,479 12 0,444 5,177 24 0,522 6,080 36 0,874 10,172 48 0,773 8,995 60 0,566 6,592 72 0,552 6,434 84 0,426 4,970 96 0,331 3,826
Tabel 2 Kesetaraan aktivitas spesifik isolat AJB 4(4) dengan konsentrasi lipase tipe VII
Waktu (jam) Aktivitas spesifik lipase (unit/mg) Konsentrasi lipase tipe VII (ppm) 0 0 0 24 0 0 48 0,935 10,888 72 1,139 13,260 96 0,734 8,552 120 0,632 7,363 144 0,651 7,577 168 0,637 7,415 192 0,568 6,616 216 0,431 5,022 240 0,414 4,823
PEMBAHASAN
Bakteri endofit yang berhasil diisolasi dari daun, batang dan akar jati belanda, jambu biji, serta rimpang temulawak dengan mengguna-kan media HV agar berjumlah 69 isolat. Keenam puluh sembilan isolat bakteri endofit tersebut menunjukkan keragaman yang tinggi baik dalam hal morfologi koloni maupun pigmentasinya. Berdasarkan morfologi koloninya, 55 isolat diantaranya dimasukkan ke dalam golongan non aktinomiset, sedangkan 14 isolat lainnya termasuk ke dalam golongan aktinomiset. Bakteri endofit paling banyak ditemukan di akar tanaman jati belanda dan jambu biji termasuk di bagian rimpang temulawak yaitu sebanyak 45 isolat.
Bakteri endofit masuk ke dalam jaringan tumbuhan melalui bagian akar lateral kemudian menyebar ke dalam ruang interseluler dan berkas pembuluh, sehingga bagian ini merupakan bagian yang paling banyak dihuni oleh bakteri dibandingkan dengan bagian lainnya pada tumbuhan (Sharma et al. 2005). Pernyataan tersebut didukung oleh Lamb et al. (1996) yang menyatakan bahwa secara umum, populasi bakteri endofit sangat melimpah di bagian akar dan kelimpahannya menurun di bagian
batang dan daun. Variasi jumlah ini sangat dipengaruhi oleh jenis tanaman inangnya, umur tanaman, jenis jaringan tanaman, waktu pengambilan sampel, dan kondisi lingkungannya. Selain itu, Leben et al. (1968) juga menyatakan bahwa bakteri endofit dapat masuk ke dalam jaringan tanaman melalui berbagai bagian tanaman yang terbuka seperti bunga, batang, daun, kotiledon, stomata dan bagian tanaman yang terluka. Bakteri ini kemudian tinggal di dalam jaringan tanaman dan menyebar ke dalam jaringan ruang interseluler atau di dalam berkas pembuluh.
Media HV agar merupakan media yang digunakan untuk mengisolasi aktinomiset. Media ini biasanya disuplementasi dengan berbagai macam antibiotik seperti asam nalidiksat untuk menekan pertumbuhan bakteri Gram negatif dan sikloheksamida untuk menekan pertumbuhan cendawan. Hasil isolasi aktinomiset pada media HV agar menunjukkan bahwa koloni aktinomiset endofit mampu tumbuh pada media tersebut. Koloni aktinomiset yang muncul kemudian dipurifikasi pada media ISP2 untuk dilakukan observasi terhadap morfologi koloni dan pigmentasinya. Koloni aktinomiset endofit yang berhasil dipurifikasi pada media ISP2 memiliki keragaman morfologi dan pigmentasi yang sangat tinggi. Aktinomiset dapat dibedakan dari bakteri lain dengan mudah dengan melihat bentuk koloninya di medium padat. Koloni Aktinomiset nampak keras seperti tumbuh akar di dalam agar-agar, berbeda dengan bakteri lain yang koloninya lunak di atas agar-agar. Selain itu, identifikasi koloni aktinomiset pada genus Streptomyces dapat dilakukan dengan mengamati morfologi rantai sporanya secara mikroskopis (Kudo 1997). Karakteristik mikroskopis ini secara jelas tampak pada isolat-isolat aktinomiset endofit yaitu isolat AJB 4(3), AJB 4(5), dan AJB 4(1) memiliki morfologi rantai spora berbentuk spiral yang berbeda-beda. Hal ini mengindikasikan bahwa isolat-isolat tersebut merupakan isolat dari genus Streptomyces dengan spesies yang berbeda. Sebaliknya, isolat AJB 4(1) dan APG 5(4) memiliki morfologi rantai spora yang mirip. Hal ini mengindikasikan bahwa kedua isolat tersebut memiliki kekerabatan yang dekat. Kudo (1997) menjelaskan bahwa bentuk permukaan spora dan rantai spora, warna miselium, warna pigmen serta penggunaan berbagai karbohidrat merupakan standar taksonomi yang dapat digunakan untuk identifikasi sampai dengan tingkat spesies.
Isolat AJB 4(3), AJB 4(5), AJB 4(1), AJB 4(4) dan APG 5(4) merupakan isolat aktinomiset yang termasuk genus Streptomyces. Hal ini berdasarkan ciri-ciri kelima isolat tersebut yang mampu membentuk miselium vegetatif dan mempunyai banyak sekali hifa serta membentuk rantai spora yang tersusun seperti rantai keriting atau heliks dari pucuk belahan hifanya. Isolat CX 12(1) merupakan isolat aktinomiset yang termasuk kedalam genus non Streptomyces karena diketahui hanya membentuk miselium vegetatif tanpa membentuk hifa sehingga koloninya terlihat lembab di atas media agar.
Berdasarkan pewarnaan Gram, diketahui bahwa ke-69 isolat bakteri endofit dapat dikelompokkan ke dalam bakteri Gram negatif (17 isolat) dan Gram positif (52 isolat). Berdasarkan teknik ini juga diketahui bahwa tiap koloni bakteri endofit memiliki morfologi sel yang berbeda-beda. Koloni bakteri endofit tersebut diketahui memiliki bentuk sel batang hingga kokus. Hal serupa juga dilaporkan oleh Zinniel et al. (2002) bahwa bakteri Gram positif maupun Gram negatif dapat diisolasi dari berbagai jenis jaringan tanaman pada sejumlah spesies tanaman yang berbeda. Keragaman ini juga dilihat dari kemampuan ke-55 isolat bakteri non-aktinomiset dalam menghasilkan endospora. Hal tersebut dibuktikan melalui kemampuan 27 dari 55 isolat bakteri endofit dalam membentuk endospora di dalam sel vegetatifnya. Posisi endospora yang dibentuk di dalam sel vegetatif juga beragam, seperti isolat AJB 3 yang mampu membentuk endospora pada posisi sub-terminal di dalam sel vegetatifnya, sedangkan isolat DPG 3(2) membentuk endospora pada posisi terminal di dalam sel vegetatifnya. Delapan dari 69 isolat hasil isolasi tersebut telah diketahui memiliki aktivitas lipase pada media penapisan agar Rhodamine B.
Enzim lipase dihasilkan oleh berbagai mikroorganisme baik secara tunggal maupun dengan senyawa-senyawa lain seperti esterase (Carboxylic-ester hydrolase EC 3.1.1.1). Penapisan bakteri penghasil lipase pada media agar biasanya dilakukan dengan menggunakan tributyrin atau Tween 80 sebagai substrat enzim. Namun, substrat tersebut tidak cocok digunakan untuk mendeteksi lipase yang sebenarnya karena senyawa tersebut juga dapat dihidrolisis oleh enzim-enzim esterase (Peled & Krenz 1981). Hofelmann et al. (1983) menyatakan bahwa lipase dapat divisualisasi pada agar yang mengandung
trioleoylglyserol dan pewarna berpendar Rhodamine B. Metode penapisan pada penelitian ini diadaptasi dari metode tersebut dengan minyak zaitun 3% digunakan sebagai substitusi trioleoylgliserol. Hasil penapisan aktivitas lipase pada media agar Rhodamine B menunjukkan sebanyak 8 isolat memiliki aktivitas lipase. Kouker dan Jaeger (1987) berpendapat bahwa proses pengikatan Rhodamine B dengan asam lemak dan mono atau digliserida sangat spesifik dan sensitif. Selain itu, pembentukan dimer kompleks antara Rhodamine B dengan asam lemak dan mono atau digliserida menghasilkan pendaran yang dapat dideteksi di bawah sinar UV. Spesifitas dari metode ini ditunjukkan ketika enzim esterase diuji pada media penapisan yang mengandung minyak zaitun 3% dan Rhodamine B, tidak ada zona berpendar yang dihasilkan. Sensitifitas metode ini telah dibuktikan melalui fakta bahwa aktivitas lipase dapat dideteksi hingga 60 nmol asam lemak yang dilepaskan per menit, sementara penapisan aktivitas lipase menggunakan metode titrimetric dapat dideteksi apabila aktivitas lipase mencapai 1200 nmol asam lemak yang dilepaskan per menit. Berdasarkan pernyataan tersebut, ke-8 koloni bakteri endofit (dua diantaranya dari golongan aktinomiset dan 6 lainnya dari golongan non aktinomiset) yang telah ditumbuhkan pada media yang mengandung minyak zaitun 3% dan pewarna Rhodamine B dapat dideteksi kemampuannya dalam menghasilkan lipase.
Isolat DPG 3(2) dan AJB 4(4) dipilih sebagai isolat uji karena mampu menghasilkan pendaran yang kuat diantara keenam isolat penghasil lipase lainnya. Hal ini serupa dengan yang dinyatakan oleh Hou dan Johnston (1992) bahwa bakteri penghasil lipase yang baik dapat dilihat dari kekuatan pendaran dan besarnya diameter zona fluorescent halo yang dihasilkan.
Isolat DPG 3(2) merupakan bakteri dari golongan non aktinomiset yang diketahui mampu mensekresikan protein yang memiliki aktivitas lipase pada media produksi yang ditambah dengan minyak zaitun 1%. Hal serupa terjadi dengan isolat Bacillus sp. strain 42 yang juga memiliki aktivitas lipase pada media yang juga ditambah dengan 1% minyak zaitun (Eltaweel et al. 2005). Aktivitas spesifik lipase tertinggi isolat DPG 3(2) didapatkan pada saat akhir fase log ketika densitas sel mencapai jumlah yang paling tinggi (4,405 log jumlah sel) pada jam ke-36 yaitu sebesar 0,874 unit per mg. Setelah jam ke-36, aktivitas lipase terlihat mulai menurun
hingga akhir pengujian yaitu saat jam ke-96 sebesar 0,331 unit per mg.
Aktinomiset merupakan mikroorganisme penghasil senyawa bioaktif baru yang paling banyak ditemukan. Lestari (2006) melaporkan bahwa isolat Streptomyces spp. yang diisolasi dari tanah di daerah Sukabumi, Kepulauan Seribu, Cipanas, dan Kalimantan Timur mampu menghasilkan senyawa antibakteri yang mampu menghambat pertumbuhan bakteri patogen seperti Bacillus subtilis, dan Xanthomonas axonopodis. Selain itu, isolat lokal aktinomiset seperti Streptomyces sp. SKK1-8 yang diisolasi dari daerah Sukabumi juga dilaporkan mampu menghasilkan enzim xylanase (Meryandini et al. 2006). Hasegawa et al. (2006) menyatakan bahwa beberapa isolat Streptomyces juga diketahui mampu memproduksi enzim-enzim hidrolitik seperti selulase, hemiselulase, kitinase, amilase, dan glukanase. Hasil yang diperoleh dari penelitian ini, diketahui bahwa isolat aktinomiset AJB 4(4) memiliki aktivitas spesifik lipase tertinggi pada jam ke-72 yaitu sebesar 1,139 unit per mg. Aktivitas lipase terlihat mulai menurun setelah jam ke-72 hingga akhir pengujian yaitu saat jam ke-240 yaitu sebesar 0,414 unit per mg. Nilai aktivitas spesifik lipase isolat DPG 3(2) dan AJB 4(4) berkorelasi positif dengan berbagai konsentrasi lipase tipe VII sebagai kontrol positifnya. Semakin tinggi nilai aktivitas spesifik lipasenya, maka semakin besar pula konsentrasi lipasenya. Hal tersebut terlihat dari nilai aktivitas spesifik lipase tertinggi isolat DPG 3(2) pada jam ke-36 setara dengan konsentrasi lipase tipe VII 10,172 ppm sedangkan nilai aktivitas spesifik lipase yang menurun pada jam ke-48 diketahui setara dengan konsentrasi lipase tipe VII 8,995 ppm. Hal yang sama juga terjadi pada isolat AJB 4(4) bahwa aktivitas spesifik lipase tertinggi pada jam ke-72 setara dengan konsentrasi lipase tipe VII 13,260 ppm dan menurunnya aktivitas spesifik lipase pada jam ke-96 diketahui setara dengan konsentrasi lipase tipe VII 8,552 ppm. Menurunnya aktivitas spesifik lipase isolat DPG 3(2) pada jam ke-48 hingga jam 96 serta isolat AJB 4(4) pada jam ke-96 hingga jam ke-240 diduga disebabkan oleh adanya metabolit yang dapat menghambat aktivitas lipase yang dikeluarkan bersama-sama dengan lipase kedua isolat tersebut pada media produksinya.
Lipase yang dihasilkan dari penelitian ini diharapkan mampu berperan dalam lipolisis cadangan lemak yang menumpuk pada jaringan adiposa. Akibat berkurangnya
cadangan lemak pada jaringan adiposa ini, secara efektif menyebabkan terjadinya penurunan berat badan.
SIMPULAN
Sebanyak 69 isolat bakteri endofit berhasil diisolasi dari tanaman jati belanda (22 isolat), jambu biji (27 isolat), dan rimpang temulawak (20 isolat). Delapan isolat diantaranya diketahui memiliki aktivitas lipase dan dua isolat diantaranya merupakan bakteri penghasil lipase terbaik yaitu isolat DPG 3(2) dan AJB 4(4). Aktifitas spesifik lipase tertinggi isolat DPG 3(2) terlihat pada jam ke-36 yaitu sebesar 0,874 unit per mg. Sedangkan aktivitas spesifik lipase tertinggi isolat isolat AJB 4(4) terlihat pada jam ke-72 yaitu sebesar 1,139 unit per mg. Kedelapan isolat berpotensi menghasilkan senyawa antihiper-lipidemia melalui aktivitas lipasenya. Isolat AJB 4(4) memiliki potensi yang lebih besar dibandingkan dengan isolat DPG 3(2).
SARAN
Perlu dilakukan optimasi kondisi pertumbuhan terkait dengan produksi senyawa antihiperlipidemia. Selain itu, perlu juga dilakukan uji in-vivo untuk melihat pengaruhnya terhadap enzim gastrointestinal lipase.
DAFTAR PUSTAKA
Bradford MM. 1976. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein dye binding. Anal Biochem. 72: 248-254
Castillo et al. 2002. Munumbicins, wide-spectrum antibiotics produce by Streptomyces NRRL 30562, endophytic on Kennedia nigrisca. Microbiology 148: 2675-2685
Dalimartha S. 2001. 36 Resep Tumbuhan Obat untuk Menurunkan Kolesterol. Edisi ke-3. Jakarta : Penebar Swadaya
Dugi et al. 2000. In vivo evidence for both lipolytic and nonlipolytic function of hepatic lipase in the metablism of HDL. Arterioscler Thromb Vasc Biol. 20: 793-800
Eltaweel MA, Rahman RNZ, Salleh AB, Basri M. 2005. An organic solvent-stable lipase from Bacillus sp. strain 42. Annals of Microbiol. 55(3): 187-192