J. Biol. Indon. Vol 6, No.3 (2010)
Vol. 6, No. 3, Desember 2010 Akreditasi: No 816/D/08/2009 BOGOR, INDONESIA
JURNAL
BIOLOGI
INDONESIA
ISSN 0854-4425
JURNAL
BIOLOGI
INDONESIA
ISSN 0854-4425
Zingiberaceae of the Ternate Island: Almost A Hundread Years After Beguin’s Collection Marlina Ardiyani
293
Production of Acid Phosphatase in Bacillus sp. Isolated from Forest Soil of Gunung Salak National Park
Maman Rahmansyah & I Made Sudiana
313
Eksplorasi Keanekaragaman Aktinomisetes Tanah Ternate Sebagai Sumber Antibiotik Arif Nurkanto, Febrianti Listyaningsih, Heddy Julistiono & Andria Agusta
325
Komposisi Flora dan Struktur Hutan Alami Di Pulau Ternate, Maluku Utara Edi Mirmanto
341
Penapisan Mikroba Laut Perombak Senyawa Nitril dan Protein yang Diisolasi Dari Sponge di Perairan Ternate
Rini Riffiani & Nunik Sulistinah
353
Perbandingan Tiga Metode Transformasi Agrobacterium Untuk Pencarian Gen-gen Terkait Toleransi Kekeringan Menggunakan Transposon Ac/Ds pada padi cv. Batutegi E.S.Mulyaningsih, H.Aswidinnoor, D.Sopandie, P.B.F.Ouwerkerk, S. Nugroho, &I.H. Slamet Loedin
367
Kajian Pakan Bersumber Energi Tinggi pada Pembentukkan Monyet Obes
Ria Oktarina, Sri Supraptini Mansjoer, Dewi Apri Astuti, Irma Herawati Suparto & Dondin Sajuthi
J. Biol. Indon. Vol 6, No. 3 (2010)
Jurnal Biologi Indonesia diterbitkan oleh Perhimpunan Biologi Indonesia. Jurnal ini memuat hasil penelitian ataupun kajian yang berkaitan dengan masalah biologi yang diterbitkan secara berkala dua kali setahun (Juni dan Desember).
Editor Pengelola Dr. Ibnu Maryanto Dr. I Made Sudiana Deby Arifiani, S.P., M.Sc
Dr. Izu Andry Fijridiyanto
Dewan Editor Ilmiah Dr. Abinawanto, F MIPA UI Dr. Achmad Farajalah, FMIPA IPB
Dr. Ambariyanto, F. Perikanan dan Kelautan UNDIP Dr. Aswin Usup F. Pertanian Universitas Palangkaraya Dr. Didik Widiyatmoko, PK Tumbuhan, Kebun Raya Cibodas-LIPI
Dr. Dwi Nugroho Wibowo, F. Biologi UNSOED Dr. Parikesit, F. MIPA UNPAD
Prof. Dr. Mohd.Tajuddin Abdullah, Universiti Malaysia Sarawak Malaysia Assoc. Prof. Monica Suleiman, Universiti Malaysia Sabah, Malaysia
Dr. Srihadi Agung priyono, F. Kedokteran Hewan IPB Y. Surjadi MSc, Pusat Penelitian ICABIOGRAD
Drs. Suharjono, Pusat Penelitian Biologi-LIPI Dr. Tri Widianto, Pusat Penelitian Limnologi-LIPI
Dr. Witjaksono Pusat Penelitian Biologi-LIPI Alamat Redaksi
Sekretariat
Oscar efendi SSi MSi
d/a Pusat Penelitian Biologi - LIPIJl. Ir. H. Juanda No. 18, Bogor 16002 , Telp. (021) 8765056 Fax. (021) 8765068
Email : [email protected]; [email protected] Website : http://biologi.or.id
Jurnal ini telah diakreditasi ulang dengan nilai A berdasarkan SK Kepala LIPI 816/ D/2009 tanggal 28 Agustus 2009.
J. Biol. Indon. Vol 6, No.3 (2010)
KATA PENGANTAR
Jurnal Biologi Indonesia yang diterbitkan oleh PERHIMPUNAN BIOLOGI INDONESIA edisi volume 6 nomer 3 tahun 2010 memuat 13 artikel lengkap. Penulis pada edisi ini sangat beragam yaitu dari Departemen Kementerian Pertanian, IPB, Puslit Biologi LIPI, Bioteknologi-LIPI dan Institute of Biology IBL Leiden Univer-sity Netherlands. Topik yang dibahas pada edisi ini meliputi 5 topik dalam bidang Botani, tiga topik tentang mikrobiologI, empat topik tentang zoologi dan satu topik campuran yang mebahas bidang botani dan zoologi. Pada edisi ini yang menarik 6 makalh merupakan hasil kajian kawaasan pulau-pulau Kecil di Ternate Maluku Utara. Selanjutnya artikel yang memuat serangga pengunjung bunga raflesia dapat dipastikan merupakan artikel sangat jarang dijumpai sehubungan dengan populasi bunganya yang sangat sulit diperoleh.
J. Biol. Indon. Vol 6, No. 3 (2010)
UCAPAN TERIMA KASIH
Jurnal Biologi Indonesia mengucapkan terima kasih dan penghargaan kepada para pakar yang telah turut sebagai penelaah dalam Volume 6, No 3, Juni 2010: Prof.Dr. Woro.A.Noerdjito Puslit Biologi-LIPI
Drs. M. Noerdjito, Puslit Biologi-LIPI Dr Yulin Lestari F MIPA-IPB Awal Riyanto, Puslit Biologi-LIPI Drs. Roemantyo, Puslit Biologi-LIPI Ir. Titi Juhaeti MSi, Puslit Biologi-LIPI Dr. Nuril Hidayati, Puslit Biologi-LIPI
J. Biol. Indon. Vol 6, No.3 (2010)
DAFTAR ISI
Zingiberaceae of the Ternate Island: Almost A Hundread Years After Beguin’s Collection Marlina Ardiyani
293
Production of Acid Phosphatase in Bacillus sp. Isolated from Forest Soil of Gunung Salak National Park
Maman Rahmansyah & I Made Sudiana
313
Eksplorasi Keanekaragaman Aktinomisetes Tanah Ternate Sebagai Sumber Antibiotik Arif Nurkanto, Febrianti Listyaningsih, Heddy Julistiono & Andria Agusta
325
Komposisi Flora dan Struktur Hutan Alami Di Pulau Ternate, Maluku Utara Edi Mirmanto
341
Penapisan Mikroba Laut Perombak Senyawa Nitril dan Protein yang Diisolasi Dari Sponge di Perairan Ternate
Rini Riffiani & Nunik Sulistinah
353
Perbandingan Tiga Metode Transformasi Agrobacterium Untuk Pencarian Gen-gen Terkait Toleransi Kekeringan Menggunakan Transposon Ac/Ds pada padi cv. Batutegi E.S.Mulyaningsih, H.Aswidinnoor, D.Sopandie, P.B.F.Ouwerkerk, S. Nugroho, &I.H. Slamet Loedin
367
Kajian Pakan Bersumber Energi Tinggi pada Pembentukkan Monyet Obes
Ria Oktarina, Sri Supraptini Mansjoer, Dewi Apri Astuti, Irma Herawati Suparto & Dondin Sajuthi
383
Pengaruh Laju Eksploitasi Terhadap Keragaan Reproduktif Ikan Tembang (Sardinella gibbosa) di Perairan Pesisir Jawa Barat
Yunizar Ernawati & Mohammad Mukhlis Kamal
393
Keragaman Genetik Amfibia Kodok (Rana nicobariensis) di Ecology Park, Cibinong Berdasarkan Sekuen DNA dari Mitokondria d-loop Dwi Astuti & Hellen Kurniati
405
Model Pemanfaatan Lahan Pulau Moti, Kota Ternate, Maluku: Suatu Analisis Tata Ruang Berbasis Vegetasi
Roemantyo
415
Komunitas Serangga pada Bunga Rafflesia patma Blume (Rafflesiaceae) di Luar Habitat Aslinya Kebun Raya Bogor Kota Bogor Provinsi Jawa Barat Indonesia
Sih Kahono, Sofi Mursidawati & Erniwati
J. Biol. Indon. Vol 6, No. 3 (2010)
Kajian Hubungan Tutupan Vegetasi dan Sebaran Burung di Pulau Moti, Ternate, Maluku Utara Hetty I.P. Utaminingrum & Eko Sulistyadi
443
Pengujian 15 Genotipe Kedelai pada Kondisi Intensitas Cahaya 50% dan Penilaian Karakter Tanaman Berdasarkan Fenotipnya
Gatut Wahyu Anggoro Susanto & Titik Sundari
353 Jurnal Biologi Indonesia 6 (3): 353-365 (2010)
Penapisan Mikroba Laut Perombak Senyawa Nitril dan Protein yang
Diisolasi Dari Spons di Perairan Ternate
Rini Riffiani & Nunik Sulistinah
Bidang Mikrobiologi, Pusat Penelitian Biologi-LIPI, Cibinong Science Center
Jl. Raya Jakarta-Bogor Km 46, Cibinong. E-mail : [email protected]
ABSTRACT
Screening of Nitrile and Protein-Degrading Marine Bacteria Isolated from Sponge in Ternate Sea Water. Thirty three marine bacteria have been isolated from marine sponge in Ternate by enrichment culture. Screening bacteria-degrading nitrile was done by microtitter plate method based on growth ability tested by Iodonitrotetazolium chloride. Product of nitrile degradation was determined by Gas Chromatography (GC) and the potential bacteria-degrading protein was also screened by using selected media which contained casein. The results showed that twenty one isolates were able to show the clearing zone in selected media. Five isolates capable of utilizing acetamide as the sole source of carbon and nitrogen. Acetate and ammonia produced for hydrolysis acetonitrile by using resting cell of Lysobacter sp.
Key words: Nitrile, bacterium, sponge, Ternate
PENDAHULUAN
Lautan Indonesia merupakan bagian wilayah Indopasifik yang mempunyai keanekaragaman biota laut terbesar di dunia. Dilaporkan bahwa jenis biota laut di daerah tropis Indonesia 2-3 kali lebih besar dibandingkan biota laut di daerah sub tropis dan daerah beriklim dingin (Sujatmiko 2000). Terumbu karang merupakan salah satu sumber daya alam tersebut dan ekosistem terumbu karang merupakan bagian dari ekosistem laut yang menjadi sumber kehidupan bagi beraneka ragam biota laut. Di dalam ekosistem terumbu karang hidup lebih dari 300 jenis karang, 200 jenis ikan dan beberapa jenis moluska, krustasea, spounge, algae dan biota lainnya (Var Soest 1994). Spons merupakan salah satu organisme laut yang mempunyai potensi
cukup besar karena kandungan senyawa bioaktifnya. Binatang laut ini mampu hidup sampai kedalaman 50 meter di bawah permukaan laut. Hasil penelitian juga menunjukkan bahwa pada porus sponge (Microcionia prolifera) ditemukan beberapa jenis bakteri, seperti genus Pseudomonas, Aerosomonas, Vibrio, Achromobacter, Flavobac-terium, CorynebacFlavobac-terium, dan Micro-coccus (Madri et al. in Reinheimer 1991) Saat ini eksplorasi mikroba banyak dilakukan karena mikroba mempunyai kemampuan mensintesis enzim-enzim tertentu yang dapat digunakan untuk kepentingan industri seperti industri pangan, obat, kimia maupun sebagai agen bioremediasi lingkungan tercemar. Beberapa mikroba perombak senyawa nitril baik alifatik maupun aromatik telah banyak ditemukan dan dimanfaatkan
354
Riffiani & Sulistinah
untuk mensintesis senyawa-senyawa penting secara komersial, seperti misalnya untuk memproduksi akrilamida dan S-(+)-Ibuprofen (Yamamoto et al.
1990, Nawas et al. 1992). Dilaporkan juga oleh Sunarko et al. 2007 mikroba potensial pendegradasi nitril dapat dikembangkan dengan meningkatkan kemampuan region, stereo, dan kemose-lektifitasnya untuk menghasilkan produk yang lebih spesifik.
Disamping mikroba pendegradasi nitril, beberapa mikroba yang mampu mensintesis enzim protease juga banyak dimanfaatkan untuk kepentingan industri, seperti industri deterjen, pengolahan susu, farmasi, dan makanan (Moon & Parulekar 1993). Enzim protease dilaporkan memiliki nilai ekonomi cukup tinggi karena aplikasinya sangat luas.
Penelitian ini merupakan studi awal untuk mengisolasi dan menapis mikroba laut potensial yang mampu mensintesis enzim pendegradasi nitril dan protease. Dari penelitian ini diharapkan diperoleh mikroba dari ekosistem laut yang nantinya dapat dimanfaatkan dan dikembangkan sebagai agen untuk mensintesis senyawa-senyawa organik maupun dimanfaatkan dibidang pangan,
serta dapat memberikan informasi mengenai data mikroba laut potensial perairan laut, khususnya di Ternate. BAHAN DAN CARA KERJA
Isolasi mikroba laut dari spons
dilakukan dengan 3 metode. Metode pertama dilakukan dengan mengisolasi langsung spons(tanpa sterilisasi) melalui pengenceran bertingkat hingga 10-3 yaitu
± 1,0 gram sampel diimasukkan ke dalam 9 ml larutan NaCl 0,85%, dihomogenkan dengan menggunakan vortex selama 10 menit, kemudian 50 μl sampel yang telah dihomogenkan diambil dan diinokulasikan ke dalam media Marine Agar. Koloni yang tumbuh diisolasi dan dimurnikan untuk pengujian lebih lanjut. Metode kedua dilakukan dengan mensterilisasi permukaan spons
dengan cara merendam potongan spons
ke dalam larutan alkohol 70% selama 10 menit, kemudian dibilas dengan akuades steril sebanyak 2 kali. Selanjutnya sampel diperlakukan sama dengan metode I. Metode ketiga dilakukan dengan memotong spons yang telah disterilkan menjadi bagian-bagian yang lebih kecil kemudian langsung ditanam pada media Marine Agar.
Media yang digunakan untuk isolasi mikroba laut adalah Marine Agar dengan komposisi sebagai berikut : 5,0 g pepton, 1,0 g yeast extract, 0,1 g ferric citrate, 19,45 g sodium chloride, 8,8 g MgCl, 3,24 g sodium sulfate, 1,88 g calcium chloride, 0,55 pottasium chloride, 0,16 g sodium bicarbonate, 15,0 g agar, 34,0 mg stronsium chloride, 22,0 mg boric acid, 4,0 mg sodium sillicate, 2,4 mg sodium flouride, 1,6 mg ammoniumnitrat, dan 8 mg disodium phosphate, aquadest 1000 ml. pH media diatur 7,4±0,2
Media penapisan yang digunakan adalah media mineral dengan komposisi sebagai berikut : Na2HPO4 0,357g, KH2PO4 0,1g, MgSO4.7H2O 0,1g, CaCl2.2H2O 0,01g, FeSO4.7H2O 0,001g, Yeast Extract 0,01g, Mikroelemen 1,0 ml, Aquadest ditambahkan sampai volume 1000 ml (Meyer & Schlegel 1983;
355
Penapisan Mikroba Laut Perombak Senyawa
Pfennig 1974). Adapun komposisi mikro-elemen : ZnSO4.7H2O, MnCl2.4H2O, H3BO3, CoCl2.6H2O, CuCl2.2H2O, NiCl2.6H2O, Na2MO4.2H2O, Na2SeO3, Aquadest 1000 ml.
Penapisan mikroba pendegradasi nitril dilakukan dengan menumbuhkan isolat bakteri hasil isolasi ke dalam microtitter plate steril yang berisi 1 ml media mineral yang mengandung senya-wa nitril dengan konsentrasi yang berbeda (dalam hal ini asetonitril/ asetamida 100 mM dan benzonitril/benza-mida 25 mM). Microtitter plate yang telah diinokulasi tersebut diinkubasi di atas mesin pengocok pada kecepatan 110 rpm pada suhu 28 oC selama 72 jam.
Pada 72 jam inkubasi, pertumbuhan mikroba diuji dengan larutan INT (Iodoni-trotetrazolium) dan aktivitasnya diuji de-ngan reagen Nessler (Oliver et al. 1989). Sebanyak 100 μl kultur ditambah dengan 14 μl larutan INT 0,5 mg/ml. Perubahan warna yang terbentuk mengindikasikan pertumbuhan mikroba. Semakin pekat warna yang terbentuk mengindikasikan pertumbuhan semakin tinggi.
Sebanyak 198 μl NaOH 0,1 N ditambahkan ke dalam 2 μl kultur, dihomogenkan kemudian ditambahkan 4 μl reagen Nessler. Selanjutnya diinkubasi pada suhu ruang ± 28oC selama 20 menit.
Aktivitas yang tinggi ditandai dengan terbentuknya warna kuning tua hingga kecoklatan.
Biomassa dari isolat bakteri terpilih diproduksi dalam erlenmeyer (1000 ml) yang berisi 500 ml media tumbuh yang mengandung 100 mM asetamida. Selanjutnya kultur diinkubasi di atas
shaker pada suhu ruang (± 28 oC)
selama 72 jam. Sel dipanen dengan mensentrifuse kultur pada kecepatan 9000 rpm selama 30 menit pada suhu 4oC.
Sel yang diperoleh dicuci dengan 50 mM buffer fosfat (KH2PO4)pH 7,2 sebanyak 2 kali. Sel tersebut selanjutnya digunakan untuk penentuan aktivitas enzim dan penentuan produk degradasi nitril.
Konsentrasi substrat dan produk degradasi ditentukan dengan menggu-nakan GC-Shimadzu 14 B, detektor FID, kolom Porapax Q, suhu kolom 225oC,
suhu injektor 240 oC, suhu detektor 240 oC (Sunarko et al. 2000). Konsentrasi
sampel ditentukan dengan rumus sebagai berikut: (sampel) = Luas area sampel/ Luas area standar X (standar)
Pengujian aktivitas enzimatik secara kualitatif dilakukan dengan menumbuh-kan isolat bakteri terpilih pada media protease agar dan diinkubasi pada suhu ruang (280C) selama 48 jam.
Terbentuk-nya atau zona bening pada media mengindikasikan bahwa mikroba yang diuji mampu mensintesis protease. Pengujian aktivitas enzim secara kuanti-tatif dilakukan dengan menambahkan substrat casein 2% (480 ì l) ke dalam 20 mM bufer fosfat pH 7 dan crude enzim sebanyak 120 ì l, kemudian campuran reaksi tersebut dihomogenkan dan diinkubasi di atas shaker pada suhu 300C
selama 30 menit. Selanjutnya ditambah-kan 600 ì l TCA 10% dan diinkubasi dalam es selama 30 menit, disentrifugasi dengan kecepatan 15.000 rpm, selama 5 menit pada suhu 40C dan supernatan
diukur pada panjang gelombang 420 nm
(Secades & Guijarro 1999).
Identifikasi dilakukan secara molekular dengan 16S rDNA. Analisis
356
Riffiani & Sulistinah
molekuler yang dilakukan berupa ekstraksi DNA dan PCR amplifikasi, purifikasi PCR produk dan sekunsing. Ekstraksi DNA menggunakan intragene matrix kit (Biorad) dilanjutkan dengan amplifikasi. Hasil optimasi PCR diperoleh komposisi per reaksi sebesar 25 μL dengan menggunakan Primer 9F (5‘GAGTTTGATCCTGGCTCG) dan 1510R (5‘GGCTACCTTGTTACGA CTT).
Analisis DNA menggunakan program BioEdit dan dilakukan blast pada Bank Gen NCBI dataLibrary. Analisis Filogenetik menggunakan program multiple aligment Clustal X versi 1.83. Konstruksi pohon filogenetik berdasarkan jarak kekerabatan genetik dengan metode Neighbor joining dan jarak evolusi dalam derajat kepercayaan menggunakan bootstrap value pada program NJ plot.
HASIL
Mikroba Pendegradasi Nitril
a.Isolasi dan Penapisan Bakteri Laut Pendegradasi Nitril
Diperoleh 33 isolat bakteri yang di isolasi dari spons yang diperoleh dari perairan laut Ternate. Penapisan mikroba pendegradasi nitril dengan teknik microtitter plate dan uji aktivitas bakteri secara kualitatif ditampilkan pada Tabel
1 dan Gambar 1 dan menunjukkan bahwa dari keseluruhan isolat yang diuji kemampuan tumbuhnya pada asetamida hanya 15 isolat bakteri yang mampu tumbuh. Sedangkan isolat bakteri yang tumbuh pada benzamida relatif lebih sedikit dan aktivitas enzimnya juga relatif lebih rendah dibandingkan pada asetamida (Tabel 1). Warna pink yang terbentuk pada saat penambahan indikator Iodonitrotetrazolium chloride (INT) pada kultur cair mengindikasikan pertumbuhan mikroba, semakin pekat warna yang terbentuk mengindikasikan pertumbuhan semakin tinggi (Gambar 1). Secara umum dari hasil pengujian menunjukkan adanya korelasi positif antara pertumbuhan dan aktivitas enzim. Isolat yang tumbuh dengan baik memiliki aktivitas enzim yang relatif cukup baik pula. Hal ini mengindikasikan, bahwa isolat bakteri yang mampu tumbuh pada amida kemungkinan besar mampu mensintesis enzim pendegradasi senyawa nitril.
b. Pola Pertumbuhan Isolat bakteri ID 04TR dan ID 13TR pada Aseta-mida 100 mM
Pola pertumbuhan dua isolat bakteri terpilih ID.04.TR dan ID.13.TR pada asetamida 100 mM ditampilkan dalam Gambar 2.
Gambar 1. Pengujian pertumbuhan bakteri pada asetamida dan benzamida sebagai substrat
357
Penapisan Mikroba Laut Perombak Senyawa
Setelah masa pertumbuhan ± 72 jam pertumbuhan nampaknya mencapai maksimal. Sedangkan bakteri ID.13.TR memerlukan fase lag yang cukup panjang yaitu ± 20 jam kemudian masuk ke fase eksponesial. Pertumbuhan isolat ID.04.TR lebih baik dibanding isolat ID.13.TR. Dengan demikian untuk pengujian selanjutnya dipilih satu isolat (ID.04.TR) yang mempunyai kemampuan tumbuh dan aktivitas yang lebih baik.
No Isolat Asetamida 100 mM Benzamida 25 mM
Pertumbuhan Aktivitas Pertumbuhan Aktivitas
1 ID.02.TR ++ ++ ++ _ 2 ID.03.TR + + _ _ 3 ID.04.TR ++++ ++++ _ _ 4 ID.06.TR _ ++ + - 5 ID.07.TR + + _ _ 6 ID.11.TR + _ _ _ 7 ID.13.TR ++ + _ _ 8 ID.14.TR _ _ _ _ 9 ID.15.TR + + _ _ 10 ID.16.TR + _ ++ + 11 ID.21.TR + _ _ _ 12 ID.22.TR ++ + ++ + 13 ID.23.TR + ++ - - 14 ID.24.TR ++ + + _ 15 ID.29.TR _ ++ _ _ 16 ID.30.TR - ++ _ _ 17 ID.31.TR + + + + 18 ID.32.TR _ ++ _ - 19 ID.33.TR _ ++ _ -
Tabel 1. Pertumbuhan dan aktivitas isolat bakteri pada asetamida dan benzamida dengan
teknik penapisan menggunakan microtitter plate
c. Pola Degradasi Isolat bakteri ID04TR dan penentuan produk degradasi.
Pola degradasi dan produk degradasi asetonitril oleh isolat bakteri ID04TR ditampilkan pada Gambar 3. Degradasi asetonitril dengan menggunakan sel utuh (whole cell) tampaknya melalui 2 alur reaksi yang melibatkan enzim nitril hidratase dan amidase. Penurunan konsentrasi asetonitril selama proses degradasi dan terbentuknya asam asetat dan ammonia memperkuat dugaan terjadinya proses degradasi asetonitril oleh aktivitas enzim pendegradasi nitril.
358
Riffiani & Sulistinah
Profil degradasi asetonitril menggu-nakan gas chromatografi (GC) memperlihatkan adanya penurunan konsentrasi asetonitril selama 180 menit inkubasi (Gambar 4, Tabel 2).
Mikroba Perombak Protein a. Penapisan Mikroba Laut Peng-hasil Enzim Protease
Hasil penapisan mikroba laut penghasil enzim protease terhadap 36 isolat bakteri menunjukkan bahwa dijumpai ada 21 isolat diantaranya
memberikan hasil positif yaitu terbentuknya clearing zone atau zona bening disekeliling koloni. Zona bening yang terbentuk merupakan indikasi awal bahwa isolat bakteri tersebut mampu mensintesis protease. Dari 21 isolat hanya 4 isolat yaitu ID.06.TR, ID.09.TR, ID.24.TR dan ID.31.TR mempunyai aktivitas tinggi dipilih untuk pengujian lebih lanjut secara kuantitatif. Selanjutnya, keempat isolat yang diduga mempunyai aktivitas protease tinggi ditumbuhkan dalam media cair yang
Waktu (menit) 0 50 100 150 200 Aset on it ri l (mM) 20 30 40 50 60 70 80 Asetonitril (CH3CN) A sam A set at ( m M) 0 1 2 3 Asam asetat (mM) A m m oni um ( m M ) 0 2 4 6 8 10 Ammonium
Gambar 3. Degradasi asetonitril menggunakan resting cell Lysobacter sp. dan pembentukan
produk metabolitnya. Waktu (Jam) 0 20 40 60 80 Per tum buhan ( O D 43 6 nm ) 0,01 0,1 1 Pertumbuhan Lysobacter sp. Kontrol (media + Lysobacter sp.) Pertumbuhan Enterobacteriaceae bacterium Kontrol (media + Enterobacteriaceae bacterium)
Gambar 2. Pertumbuhan Lysobacter sp. dan Enterobacteriaceae bacterium pada Asetamida
359
Penapisan Mikroba Laut Perombak Senyawa
Standar asetonitril Inkubasi 15 menit 30 menit 150 menit 180 menit
peak asetonitril (standar)
peak asetonitril sampel
Gambar 4. Penurunan konsentrasi asetonitril selama proses degradasi menggunakan Gas Chromatografi (GC)
Tabel 2. Konsentrasi substrat/produk biotransformasi kelima isolat Waktu
inkubasi
Konsentrasi substrat/produk biotransformasi (mM)
asetonitril asam asetat amonium
15 53,614 1,3754 7,2
60 53,167 0,2960 1,55
90 47,522 0,4232 2,22
120 37,692 0,323 1,90
180 26,144 0,1860 9,70
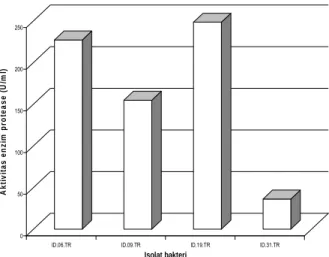
mengandung casein dan diinkubasi selama 48 jam dengan pengocokan. Hasil pengujian aktivitas memperlihatkan bahwa isolat bakteri ID.19.TR memiliki aktivitas enzim protease yang paling tinggi yaitu sebesar 226,85 (U/ml) dan aktivitas terendah ditunjukkan oleh isolat bakteri ID.31.TR sebesar 36,100 U/ml (Gambar 5)
b. Identifikasi dan Analisis Filogene-tik Isolat Terpilih
Berdasarkan analisis penjajaran urutan nukleotida parsial gen pengkode
16S rDNA menggunakan program
BLAST ditampilkan pada Tabel 3 . PEMBAHASAN
Penapisan bakteri laut perombak senyawa nitril dan amida
Hasil penapisan 33 isolat bakteri spons menggunakan asetamida dan benzamida menunjukkan bahwa isolat isolat bakteri ID.02.TR (Micrococcus luteus strain G3-6-08)ID.04.TR (Lyso-bacter sp.), ID.13.TR ( Enterobacte-riaceae bacterium B12), ID.22.TR
360
Riffiani & Sulistinah
(Arthrobacter sp. WPCB190) ID.24.
TR (Sphingomonas sp.) dan ID.31.TR
(Spons bacterium IS8) mampu tumbuh dalam 100 mM asetamida dan mempunyai aktivitas enzim yang relatif lebih baik dibandingkan isolat-isolat yang lainnya. Dari Tabel 1 tampak bahwa isolat yang mempunyai kemampuan tumbuh yang baik memiliki aktivitas enzim yang baik mampu tumbuh pada asetamida kemungkinan juga mampu mensintesis enzim pendegradasi senyawa nitril. Tabel 1 tersebut juga menunjukkan, bahwa Lysobacter sp. menunjukkan kemampu-an tumbuh dkemampu-an aktivitas ykemampu-ang paling baik dibandingkan tiga isolat lain. Dengan demikian bakteri Lysobacter sp. merupa-kan isolat potensial untuk pengujian lebih lanjut.
Penapisan mikroba dengan menggunakan benzamida menunjukkan bahwa hanya 2 isolat bakteri yang mampu
0 50 100 150 200 250 A k ti v it a s en zi m pr o tea se ( U /m l)
ID.06.TR ID.09.TR ID.19.TR ID.31.TR Isolat bakteri
Gambar 5. Aktivitas enzim protease isolat ID.06.TR, ID.09.TR, ID.19.TR, ID.31.TR tumbuh dengan baik pada benzamida yaitu ID.02.TR (Micrococcus luteus strain
G3-6-08), dan ID.22.TR (Arthrobacter
sp. WPCB190) dengan aktivitas enzim yang relatif rendah. Ketidakmampuan tumbuh isolat pada benzamida disebabkan karena benzamida merupakan senyawa turunan benzonitril dan tergolong dalam amida aromatik yang sangat toksik. Disamping itu benzamida dan benzonitril mempunyai kelarutan yang sangat rendah dibandingkan asetamida atau asetonitril, sehingga sulit didegradasi dan dimetabo-lisme sebagai sumber energi, karbon dan nitrogen untuk pertumbuhan mikroba. Dilaporkan sedikit mikroba yang mampu tumbuh pada benzonitril dan benzamida (Sunarko et al. 2000). Dengan demikian ditunjukkan bahwa kemampuan tumbuh isolat bakteri pada asetamida lebih tinggi dibandingkan pada benzamida karena asetamida merupakan senyawa alifatik
361 Penapisan Mikroba Laut Perombak Senyawa
No ID Isolat Hasil identifikasi (16S rDNA) Homologi (100 %)
Kemampuan 1 ID.02.TR Micrococcus luteus strain G3-6-08 98 amidase +
protease +
2 ID.04TR Lysobacter sp. 99 amidase +
protease +
3 ID.12TR Vibrio harveyi 96 protease +
4 ID.13.TR Enterobacteriaceae bacterium B12 89 amidase +
5 ID.19.TR Sphingomonas phyllosphaerae strain FA2
90 protease +
6 ID.22TR Arthrobacter sp. WPCB190 98 amidase +
7 ID.23.TR Bacillus pumilus strain ST277 96 protease +
8 ID.24. TR Sphingomonas sp. 98 amidase +
9 ID.31.TR Sponge bacterium IS8 99 amidase + protease +
Tabel 3.
Identifikasi 9 isolat bakteri laut potensial hasil penapisan
amida yang cenderung lebih mudah dihidrolisis oleh mikroba.
Biodegradasi asetonitril
Degradasi asetonitril dengan menggunakan resting cell Lysobacter sp. yang ditumbuhkan dalam 100 mM tampaknya melalui 2 jalur reaksi yang melibatkan enzim nitril hidratase dan amidase. Hal ini terbukti dengan menurunnya konsentrasi asetonitril selama proses degradasi tersebut dengan terbentuknya asam asetat dan amonia memperkuat terjadinya proses degradasi asetonitril oleh aktivitas enzim pendegradasi nitril. Secara umum, biodegradasi senyawa nitril dilaporkan melalui dua alur reaksi yang menghasilkan asam karboksilat dan ammonia (NH4+) sebagai produk akhirnya (Nagasawa et al. 1987; Kobayashi1991). Pada alur reaksi pertama melibatkan enzim nitril hidratase
(E.C. 4.3.2.84) dan amidase (E.C. 3.5.1.4), sedangkan reaksi kedua melibatkan enzim nitrilase (E.C.3.5.5.1). Diketahui bahwa degradasi senyawa nitril alifatik melibatkan nitril-hidratase dan amidase, sedangkan degradasi nitril aromatik hanya melibatkan enzim nitrilase saja (Nagasawa et al. 1987; Banerjee et al. 2002).
Dari hasil persamaan regresi
menunjukkan bahwa Lysobacter sp
mampu menurunkan konsentrasi
asetonitril (190 mM) sampai konsentrasi
0 mM dalam waktu 5 jam 35 menit
(Gambar 6).
Berdasarkan Uji Korelasi menggu-nakan program SPSS versi 12 (P=0,05), menunjukkan bahwa penurunan konsen-trasi asetonitril ternyata tidak berkorelasi dengan terbentuknya asetat dan amonia. Selanjutnya selama penelitian ini ditunjukkan bahwa penurunan asetonitril
362
Riffiani & Sulistinah
tidak seiring dengan peningkatan asam asetat dan amonia. Hal ini kemungkinan karena asetat dan amonia yang terbentuk digunakan mikroba sebagai sumber karbon dan nitrogen untuk pertumbuhan. Dilaporkan oleh Langdahl et.al. (1996), asetat yang dihasilkan dari degradasi asetonitril oleh Rhodococcus erythro-polis BL 1 digunakan sebagai sumber karbon untuk pertumbuhan.
Kemampuan tumbuh Lysobacter sp. pada 100 mM asetamida relatif lebih rendah bila dibandingkan Rhodococcus erythropolis BL 1 yang diisolasi dari sedimen laut. R erythropolis mampu tumbuh pada asetronil hingga konsentrasi 1 M, fase lag dicapai ± 50 amonia pada fase eksponensial (Langdahl et al. 1996). Dilaporkan juga oleh Sunarko et al. 2000 bahwa Corynebacterium sp. D5 yang diisolasi dari limbah kimia ASTRA juga mampu tumbuh pada asetonitril hingga 5% (± 950 mM). Nampak mikroba yang diisolasi dari lingkungan yang ekstrim mempunyai toleransi yang tinggi terhadap senyawa pencemar. y = -0,18x + 60,37 R2 = 0,9175 0 10 20 30 40 50 60 70 0 50 100 150 200
Bakteri Laut Perombak Protein Beberapa bakteri yang memiliki enzim amidase juga memiliki aktivitas protease yaitu Micrococcus luteus, Lysobacter sp. dan Spons bacterium. Isolat bakteri dengan ID.19.TR teridentifikasi sebagai Sphingomonas phyllosphaerae strain FA2 memiliki aktivitas enzim protease tertinggi yaitu 226,85 (U/ml) . Akan tetapi kemampuan S. phyllosphaerae menghasilkan enzim protease jauh lebih rendah bila dibandingkan dengan Bacillus subtilis. Safey & Raouf (2004) melaporkan bahwa B. subtilis memiliki aktivitas spesifik ekstraselular enzim protease sebesar 6381,71 (unit/mg prot/ml-1).
Protease adalah enzim yang menghi-drolisis ikatan peptida pada protein. Enzim ini seringkali dibedakan menjadi proteinase dan peptidase. Protease mengkatalisis hidrolisis molekul protein menjadi fragmen-fragmen besar, sedangkan peptidase mengkatalisis hidrolisis fragmen polipeptida menjadi asam amino. Protease memegang peranan utama dalam banyak fungsi hayati, mulai dari tingkat sel, organ,
363 Penapisan Mikroba Laut Perombak Senyawa
sampai organisme, yaitu dalam melangsungkan reaksi metabolisme dan fungsi regulasi (Mell et al. 2000).
Berdasarkan hasil yang diperoleh menunjukkan beberapa bakteri laut yang telah diisolasi dari sampel spons dari perairan Ternate nampaknya selaras dengan penelitian yang dilakukan oleh Suryati et al (2000) bahwa kelimpahan jenis bakteri yang diisolasi dari spons di perairan Spermonde, Sulawesi Selatan didominasi oleh bakteri Vibrio sp. Pseudomonas sp., Spingomonas sp, Flavobacterium sp, Acinetobacter sp, dan Bacillus sp. Meyers et al. (2001) melaporkan bahwa terjadi hubungan simbiosis antara spons dan sejumlah
bakteri, dimana spons menyediakan dukungan dan perlindungan bagi bakteri sebagai simbion dan simbion menyedia-kan mamenyedia-kanan bagi spons.
Hasil analisis filogenik bakteri laut potensial yang diisolasi dari spons asal Perairan Ternate dapat dilihat pada Gambar 7. Pohon filogeni menunjukkan bahwa bakteri potensial yang diisolasi dari spons terbagi menjadi enam kelompok bakteri yaitu kelompok bakteri
Arthrobacter, Bacillus, Lysobacter,
Micrococcus , Spingomonas, dan
Spons bacterium.
364
Riffiani & Sulistinah
KESIMPULAN
Isolat bakteri ID04TR teridentifikasi sebagai Lysobacter sp mampu tumbuh dan memanfaatkan (100mM) asetonitril sebagai satu-satunya sumber karbon, energi, dan nitrogen. Proses degradasi asetonitril dengan menggunakan resting sel Lysobacter sp melibatkan enzim nitril hidratase dan amidase. Asam asetat dan amonium merupakan produk degradasi. Isolat bakteri dengan ID 19TR teridentifikasi sebagai Sphingomonas phyllosphaerae strain FA2 mampu merombak protein dan memiliki aktivitas enzim protease tertinggi.
UCAPAN TERIMA KASIH
Penulis mengucapakan terima kasih kepada Prof. Dr. Ibnu Maryanto sebagai koordinator Projek IPTEKDA KHUSUS yang telah mendanai kegiatan penelitian ini. Ucapan terima kasih juga disampaikan Arief Nurkanto, S.Si dan Munir yang telah banyak membantu dalam penelitian ini.
DAFTAR PUSTAKA
Banerjee, A., R. Sharma, UC. Banerjee. 2002. The nitrile degrading enzymes: current status and future prospect. Appl. Microbiol. Biotechnol., 60 : 33-34
Kobayashi, M. 1991. Studies on Enzymes Involved in Nitrile Metabolism in
Rhodococcus rhodochrous. PhD
[Thesis].
Langdahl, BR., P. Bisp & K. Ingvorsen. 1996. Nitrile hydrolysis by
Rhodococcus erythropolis BL1,
an acetonitrile-tolerant strain isolated from a marine sediment.
Microbiol. Letter. 142 : 145-154 Mel, SF, K. Fuller, S. Wimer-Mackin, W.
Lencer., & J. Mekalanos. 2000. Association of protease activity in Vibrio cholerae Vaccine Strains With Decreases in Transcellular Epithelial Resistance of Polarized T84 Intestinal Epithelial Cells. Infect Immune 68(11):6487-92.
Moon, SH & SJ Parulekar. 1993. Some observation on protease producing in continuous suspension cultures of Bacillus firmus. Biotech. Bioengin. 41, 43-54
Meyer, O. & HG. Schlegel.1983. Biology of aerobic carbon monoxide oxidizing bacteria. Ann. Rev. Microbiol. 37 :277-310.
Nagasawa, T., H. Nanba, K. Ryuno, K. Takeuchi & H. Yamada. 1987. Nitrile Hydratase of Pseudomonas chlororaphis B23. Eur. J. Biochem. 162 : 1305-1312.
Nawaz, MS., TM. Heinze & C.A Cerniglia. 1992. Metabolism of Benzonitrile and Butyronitrile by
Klebsiella pneumoniae. Appl. Environ. Microbiol. 38: 27-31
Oliver, MH., NK. Harison, JE. Bishop, PJ. Cole & GJ. Lauren. 1989. A rapid and convenient assay for counting cells cultured in microwell plates : Application for assessment
365
Penapisan Mikroba Laut Perombak Senyawa
of growth factors. J. Cell. Scie.
92 : 513-518.
Pfennig, N. 1974. Rhodopseudomonas globiformis sp.A new species of Rhodospirillaceae. Arch. Micro-biol. 100 : 197-206.
Reinheimer, G. 1991. Aquatic Microbiology. 4 th Ed. John Wiley and Sons, Chichester and New York Secades P., & J.A Guijarro.1999. Purification and Characterization of an Extrascellular Protease from the Fish Pathogen Yersinia ruckeri and Effect of Culture Condition on Production. App.Envir. Microbiol. 65 (9): 3969-3975
Sujatmiko, W. 2000. Inventarisasi Jenis Spons Disekitar Perairan Pulau Lombok dan Garam di Pulau Sumbawa Nusa Tenggara Barat, Kerjasama: Yayasan Rinjani Bahari, Badan Perencanaan dan Penerapan Teknologi, Badan Perencanaan dan Pembangunan Propinsi Nusa Tenggara Barat. Sunarko, B., Adityarini, USF. Tambunan
& N. Sulistinah. 2000. Isolasi, seleksi dan karakterisasi mikroba pendegradasi asetonitril. Berita Biologi 5(2) : 177-185.
Sunarko, B., TU. Harwati, AT. Utari, D. Setianingrum & L. Nurhayani. 2007. Penapisan mikroba potensial untuk biokatalis produksi senyawa obat antiinflamasi nonsteroid (AINS). Laporan Teknik Kegia-tan Pusat Penelitian Biotekno-logi DIPA tahun 2006.
Suryati, E., A Parenrengi, & Rosmiati. 2000. Penapisan Serta Analisis Kandungan Bioaktif Spons Clathria sp. yang efektif sebagai Antibiofouling pada teritif (Balanus amphitrit). J.Penel.Perik. Indo. 5 (3)
Van Soest, RMW. 1994. Desmospons Distribution Pattern, In: Eds. Van Soest, R.W.M Van Kompen . T.M.G . Braekman, J.C. Spons in Time and Space. A.A Balkema, Rotterdam
Yamamoto, K., Y. Ueno, K. Otsubo, K. Kawakami & K. Komatsu. 1990. Production of S- (+)-Ibuprofen from a nitrile compound by Acinetobacter sp. Strain AK226. J. App.Envir. Miro. 56 : 3125-3129.
Memasukkan: Desember 2009 Diterima: Mei 2010
J. Biol. Indon. Vol 6, No.3 (2010)
PANDUAN PENULIS
Naskah dapat ditulis dalam bahasa Indonesia atau bahasa Inggris. Naskah disusun dengan urutan: JUDUL (bahasa Indonesia dan Inggris), NAMA PENULIS (yang disertai dengan alamat Lembaga/ Instansi), ABSTRAK (bahasa Inggris, maksimal 250 kata), KATA KUNCI (maksimal 6 kata), PENDAHULUAN, BAHAN DAN CARA KERJA, HASIL, PEMBAHASAN, UCAPAN TERIMA KASIH (jika diperlukan) dan DAFTAR PUSTAKA.
Naskah diketik dengan spasi ganda pada kertas HVS A4 maksimum 15 halaman termasuk gambar, foto, dan tabel disertai CD. Batas dari tepi kiri 3 cm, kanan, atas, dan bawah
masing-masing 2,5 cm dengan program pengolah kata Microsoft Word dan tipe huruf Times New Roman
berukuran 12 point. Setiap halaman diberi nomor halaman secara berurutan. Gambar dalam bentuk grafik/diagram harus asli (bukan fotokopi) dan foto (dicetak di kertas licin atau di scan).
Gambar dan Tabel di tulis dan ditempatkan di halam terpisah di akhir naskah. Penulisan simbol α,
β, χ, dan lain-lain dimasukkan melalui fasilitas insert, tanpa mengubah jenis huruf. Kata dalam
bahasa asing dicetak miring. Naskah dikirimkan ke alamat Redaksi sebanyak 3 eksemplar (2 eksemplar tanpa nama dan lembaga penulis).
Penggunaan nama suatu tumbuhan atau hewan dalam bahasa Indonesia/Daerah harus diikuti nama ilmiahnya (cetak miring) beserta Authornya pada pengungkapan pertama kali.
Daftar pustaka ditulis secara abjad menggunakan sistem nama-tahun. Contoh penulisan pustaka acuan sebagai berikut :
Jurnal :
Hara, T., JR. Zhang, & S. Ueda. 1983. Identification of plasmids linked with polyglutamate
production in B. subtilis. J. Gen. Apll. Microbiol. 29: 345-354.
Buku :
Chaplin, MF. & C. Bucke. 1990. Enzyme Technology. Cambridge University Press. Cambridge.
Bab dalam Buku :
Gerhart, P. & SW. Drew. 1994. Liquid culture. Dalam : Gerhart, P., R.G.E. Murray, W.A. Wood,
& N.R. Krieg (eds.). Methods for General and Molecular Bacteriology. ASM., Washington.
248-277.
Abstrak :
Suryajaya, D. 1982. Perkembangan tanaman polong-polongan utama di Indonesia. Abstrak Pertemuan Ilmiah Mikrobiologi. Jakarta . 15 –18 Oktober 1982. 42.
Prosiding :
Mubarik, NR., A. Suwanto, & MT. Suhartono. 2000. Isolasi dan karakterisasi protease ekstrasellular dari bakteri isolat termofilik ekstrim. Prosiding Seminar nasional Industri Enzim dan Bioteknologi II. Jakarta, 15-16 Februari 2000. 151-158.
Skripsi, Tesis, Disertasi :
Kemala, S. 1987. Pola Pertanian, Industri Perdagangan Kelapa dan Kelapa Sawit di Indonesia.[Disertasi]. Bogor : Institut Pertanian Bogor.
Informasi dari Internet :
Schulze, H. 1999. Detection and Identification of Lories and Pottos in The Wild; Information for surveys/Estimated of population density. http//www.species.net/primates/loris/ lorCp.1.html.
J. Biol. Indon. Vol 6, No. 3 (2010)
Pengaruh Laju Eksploitasi Terhadap Keragaan Reproduktif Ikan Tembang (Sardinella gibbosa) di Perairan Pesisir Jawa Barat
Yunizar Ernawati & Mohammad Mukhlis Kamal
393
Keragaman Genetik Amfibia Kodok (Rana nicobariensis) di Ecology Park, Cibinong Berdasarkan Sekuen DNA dari Mitokondria d-loop Dwi Astuti & Hellen Kurniati
405
Model Pemanfaatan Lahan Pulau Moti, Kota Ternate, Maluku: Suatu Analisis Tata Ruang Berbasis Vegetasi
Roemantyo
415
Komunitas Serangga pada Bunga Rafflesia patma Blume (Rafflesiaceae) di Luar Habitat Aslinya Kebun Raya Bogor Kota Bogor Provinsi Jawa Barat Indonesia
Sih Kahono, Sofi Mursidawati & Erniwati
429
Kajian Hubungan Tutupan Vegetasi dan Sebaran Burung di Pulau Moti, Ternate, Maluku Utara Hetty I.P. Utaminingrum & Eko Sulistyadi
443
Pengujian 15 Genotipe Kedelai pada Kondisi Intensitas Cahaya 50% dan Penilaian Karakter Tanaman Berdasarkan Fenotipnya
Gatut Wahyu Anggoro Susanto & Titik Sundari