MORFOTIPE EKTOMIKORIZA FAMILI DIPTEROCARPACEAE
DENGAN METODE LUBANG BOR DI HUTAN
PENELITIAN HAURBENTES, BOGOR
MUHAMMAD GINANJAR MULYANA
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
BOGOR 2014
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa skripsi berjudul Morfotipe Ektomikoriza Famili Dipterocarpaceae dengan Metode Lubang Bor di Hutan Penelitian Haurbentes, Bogor adalah benar karya saya bersama komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Mei 2014 Muhammad Ginanjar Mulyana NIM G34070075
* Pelimpahan hak cipta atas karya tulis dari penelitian kerja sama dengan pihak luar IPB harus didasarkan pada perjanjian kerja sama yang terkait.
ABSTRAK
MUHAMMAD GINANJAR MULYANA. Morfotipe Ektomikoriza Famili Dipterocarpaceae dengan Metode Lubang Bor di Hutan Penelitian Haurbentes, Bogor. Dibimbing oleh NAMPIAH SUKARNO dan UTUT WIDYASTUTI.
Di Indonesia, famili Dipterocarpaceae dapat hidup pada tanah masam yang miskin unsur hara dan aktivitas dekomposisi bahan organik yang rendah karena tumbuhan ini hidup bersimbiosis membentuk ektomikoriza. Penelitian ini bertujuan untuk mempelajari morfotipe ektomikoriza di Hutan Penelitian Haurbentes, Bogor. Sampel akar diperoleh dengan membuat lubang bor berdiameter 5 cm dengan kedalaman 15 cm. Analisis morfotipe dilakukan dengan mengamati bentuk percabangan, warna, mantel, dan rizomorf yang terbentuk pada akar yang terkolonisasi cendawan ektomikoriza. Berdasarkan hasil analisis dari 191 potong akar dari 48 sampel lubang bor menunjukkan bahwa delapan spesies tumbuhan dari famili Dipterocarpaceae dikolonisasi oleh cendawan ektomikoriza dengan rata-rata kolonisasi sebesar 34.71 % (2.15 x 102 cm) yang membentuk 12 morfotipe yang berbeda. Ke-12 morfotipe tersebut ialah tidak bercabang hitam lurus, tidak bercabang hitam bengkok, tidak bercabang krem bengkok, monopodial piramida cokelat lurus, monopodial menyirip hitam bergelombang, monopodial menyirip cokelat lurus, monopodial menyirip cokelat bengkok, monopodial menyirip cokelat bergelombang, menyirip tidak beraturan hitam bergelombang, menyirip tidak beraturan cokelat bergelombang, menyirip tidak beraturan cokelat bengkok, dan menyirip tidak beraturan krem bengkok. Kelimpahan kolonisasi akar tertinggi diperoleh dari morfotipe menyirip tidak beraturan cokelat bergelombang yaitu sebesar 32.54 %, sedangkan terendah dari morfotipe monopodial menyirip hitam bergelombang yaitu sebesar 0.37 %.
Kata kunci: Dipterocarpaceae, Ektomikoriza, Hutan Penelitian Haurbentes, Morfotipe
ABSTRACT
MUHAMMAD GINANJAR MULYANA. Root tip Morphotypes of Dipterocarpaceae Ectomycorrhizas Based on Coring Method in Haurbentes Conservation Forest, Bogor. Supervised by NAMPIAH SUKARNO and UTUT WIDYASTUTI.
In Indonesian, Dipterocarpaceae grows in nutrient-poor and acidic soils which usually characterized by low rates of organic matter decomposition, due to it’s selective advantage of ectomycorrizas association. This research aimed to study ectomycorrizal root tip morphotypes in Haurbentes conservation forest, Bogor. The root tip morphotype was determinited by observation of root branching pattern, colour, mantle, and rhizomorph. One hundred ninety one root tips from 48 core samples derived from 8 spesies of Dipterocarpaceae were observed for root colonization and morphotype. Data showed that roots of all plant species were colonized by ectomycorrizas with the average value 34.72% or 2.15 x 102 cm. They
produced 12 different morphotypes. They were unramified-simple black straight, unramified-simple black bent, unramified-simple cream colored bent, monopodial pyramide brown straight, monopodial pinnate black tortuous, monopodial pinnate brown straight, monopodial pinnate brown bent, monopodial pinnate brown tortuous, irregularly pinnate black tortuous, irregularly pinnate brown tortuous, irregularly pinnate brown bent, and irregularly pinnate cream colored bent. The highest value of root colonization abundance was observed from irregularly pinnate brown morphotype (32.54 %), while the lowest was found in monopodial pinnate black tortuous (0.37 %).
Keywords: Dipterocarpaceae, Ectomycorrhizas, Haurbentes conservation forest, Root tips morphotype
© Hak Cipta Milik IPB, Tahun 2014
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah, dan pengutipan tersebut tidak merugikan IPB
Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis ini dalam bentuk apa pun tanpa izin IPB
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains
pada
Departemen Biologi
MORFOTIPE EKTOMIKORIZA FAMILI DIPTEROCARPACEAE
DENGAN METODE LUBANG BOR DI HUTAN
PENELITIAN HAURBENTES, BOGOR
MUHAMMAD GINANJAR MULYANA
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
BOGOR 2014
Judul Skripsi : Morfotipe Ektomikoriza Famili Dipterocarpaceae dengan Metode Lubang Bor di Hutan Penelitian Haurbentes, Bogor
Nama : Muhammad Ginanjar Mulyana
NIM : G34070075
Disetujui oleh
Dr Ir Nampiah Sukarno Pembimbing I
Dr Ir Utut Widyastuti, MSi Pembimbing II
Diketahui oleh
Dr Ir Iman Rusmana, MSi Ketua Departemen
PRAKATA
Innalhamdalillah, puji syukur penulis panjatkan kehadirat Allah Jalla Jalaluhu atas limpahan nikmat dan karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian ini ialah ektomikoriza, dengan judul Morfotipe Ektomikoriza Famili Dipterocarpaceae dengan Metode Lubang Bor di Hutan Penelitian Haurbentes, Bogor. Kegiatan penelitian ini berlangsung dari bulan Juni-November 2013 di Laboratorium Mikologi Departemen Biologi IPB.
Terima kasih penulis ucapkan kepada Ibu Dr Ir Nampiah Sukarno selaku pembimbing I dan pemberi dana penelitian, Ibu Dr Ir Utut Widyastuti MSi selaku pembimbing II, Ibu Dr Ir Nunik Sri Ariyanti MSi dan Ibu Dr Ir Sri Listiyowati MSi yang telah banyak memberi motivasi dan saran, serta Prof Dr Alex Hartana selaku penguji atas saran dan masukkannya. Di samping itu, penghargaan penulis sampaikan kepada Staf Departemen Biologi, keluarga besar Laboratorium Mikologi IPB, keluarga besar Wisma Asri, keluarga besar Lembur Alit, keluarga besar Lab Biokontrol Balithi, keluarga besar Center of Tropical Animal Studies Lembaga Penelitian dan Pengabdian kepada Masyarakat (LPPM) IPB, keluarga besar Serang, keluarga besar Jampang, ayah, ibu, adik, neng Saprilian SH, serta keluarga atas doa dan kasih sayangnya.
Semoga karya ilmiah ini bermanfaat.
Bogor, Mei 2014
DAFTAR ISI
DAFTAR TABEL vi DAFTAR GAMBAR vi DAFTAR LAMPIRAN vi PENDAHULUAN 1 Latar Belakang 1 Tujuan Penelitian 2 METODE 2Bahan dan Alat 2
Prosedur 2
HASIL DAN PEMBAHASAN 3
Hasil 3 Pembahasan 7 SIMPULAN 9 DAFTAR PUSTAKA 10 LAMPIRAN 12 RIWAYAT HIDUP 15
DAFTAR TABEL
1 Keragaman morfotipe, persentase kolonisasi dan kelimpahan ektomikoriza pada akar tumbuhan, dan panjang akar yang terkolonisasi cendawan ektomikoriza famili Dipterocarpaceae dari 48 sampel lubang bor di Hutan Penelitian Haurbentes, Bogor dengan
menggunakan metode garis sentuh 6
DAFTAR GAMBAR
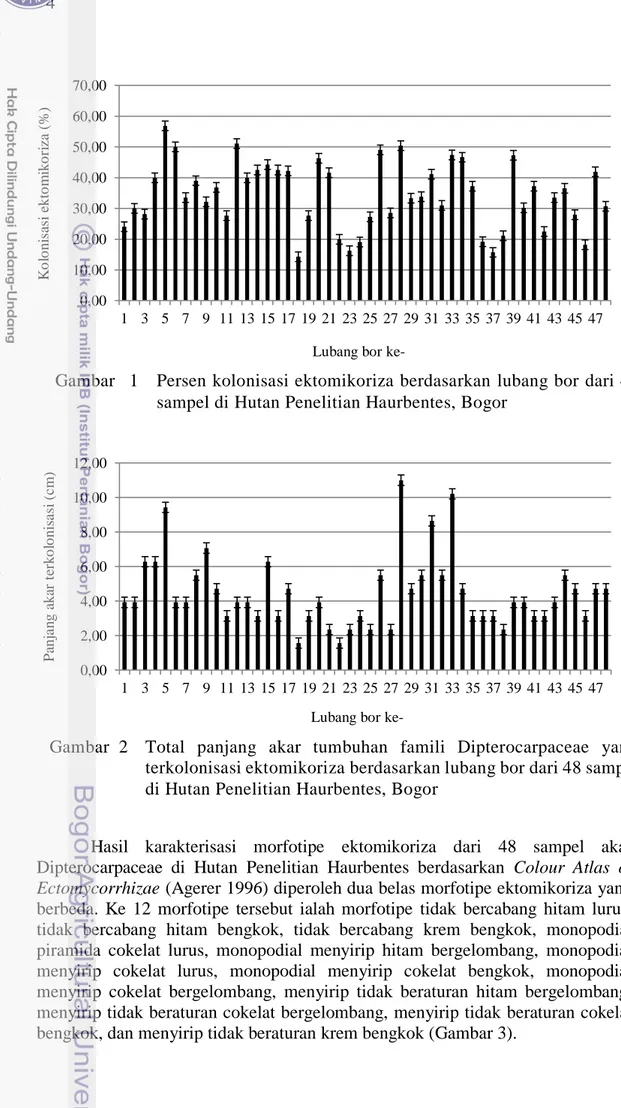
1 Persen kolonisasi ektomikoriza berdasarkan lubang bor dari 48 sampel
di Hutan Penelitian Haurbentes, Bogor 4
2 Total panjang akar tumbuhan famili Dipterocarpaceae yang terkolonisasi ektomikoriza berdasarkan lubang bor dari 48 sampel di
Hutan Penelitian Haurbentes, Bogor 4
3 Dua belas morfotipe ektomikoriza yang diperoleh dari 48 sampel lubang bor di Hutan Penelitian Haurbentes, Bogor 5
DAFTAR LAMPIRAN
1 Daftar lubang bor, nama tanaman inang, dan kondisi lingkungan
1 1
PENDAHULUAN
Latar Belakang
Famili Dipterocarpaceae secara alami tumbuh baik di hutan hujan tropis dan menjadi bagian yang sangat penting di dalam ekosistem tersebut. Dipterocarpaceae juga merupakan famili tumbuhan yang sering dijadikan pilihan utama dalam program reboisasi di Indonesia sebagai upaya menjaga kelestarian hutan yang terus terancam akibat eksploitasi hutan yang semakin tidak terkendali (Newman et al. 1999). Selain itu, spesies dari famili ini menghasilkan produk kayu dan non kayu yang bernilai ekonomi tinggi. Indonesia merupakan negara yang memiliki keragaman spesies tertinggi dari famili Dipterocarpaceae yang mencapai 13 genus dari total keseluruhan 16 genus di dunia (CIFOR 1998). Famili Dipterocarpaceae di Indonesia dapat dijumpai di Kalimantan, Sumatera, Jawa, Sulawesi, Maluku, dan Papua. Selain tumbuh di habitat aslinya, tumbuhan famili ini ditanam di Kawasan Hutan dengan Tujuan Khusus yaitu Hutan Penelitian Haurbentes, Bogor sebagai upaya konservasi plasma nutfah dan produksi benih dari berbagai spesies dari famili Dipterocarpaceae di Indonesia.
Indonesia merupakan salah satu negara yang memiliki keragaman mikroorganisme tertinggi di dunia, termasuk keragaman cendawan ektomikoriza. Keragaman cendawan ektomikoriza yang tinggi sangat menentukan keberhasilan dan efektifitas simbiosis pada kondisi tanah yang beragam. Kondisi lingkungan hutan yang memiliki tanah masam, miskin unsur hara, dan aktivitas dekomposisi bahan organik rendah bukan merupakan faktor pembatas untuk pertumbuhan dan dominasi famili tumbuhan ini di hutan hujan tropis karena tumbuhan tersebut dapat membentuk simbiosis ektomikoriza (Moyersoen et al. 2001). Ektomikoriza merupakan simbiosis mutualisme antara cendawan asal tanah dengan perakaran tumbuhan (Brundrett 2004). Simbiosis ini ditandai dengan terbentuknya struktur tubuh buah cendawan di luar akar, jalinan hifa yang menyelubungi akar rambut tumbuhan inang (mantel), hifa eksternal dan rizomorf yang mengeksplorasi tanah, dan jaringan hartig di daerah intraseluler epidermis serta korteks akar (Olsson et al. 2000). Interaksi antara cendawan dan inangnya merubah struktur fisik, anatomi, dan morfologi akar yaitu akar terlihat memendek, menebal, dan memiliki warna yang berbeda dari akar yang tidak terkolonisasi cendawan ektomikoriza (Brundrett et al. 1996).
Penelitian mengenai simbiosis ektomikoriza pada famili Dipterocarpaceae telah banyak dilakukan di Indonesia, namun hanya terbatas pada aspek penyerapan unsur hara oleh cendawan ektomikoriza dan pengaruhnya terhadap tumbuhan inang (Riniarti 2002; Prameswari 2004; Turjaman et al. 2005; Turjaman et al. 2006; Noor 2009; Turjaman et al. 2011). Aspek keragaman morfotipe ektomikoriza pada famili Dipterocarpaceae masih belum banyak dipublikasikan. Saat ini, ini tercatat hanya terdapat laporan morfologi ektomikoriza pada tanaman Dipterocarpaceae dari luar negeri yaitu keragaman ektomikoriza Dipterocarpaceae di Argentina (Becerra et al. 2004), Thailand (Amornpitak et al. 2006), Venezuela (Moyersoen 2006), dan Guyana (Brearley 2012; Smith et al. 2013). Oleh karena itu, penelitian untuk mempelajari keragaman morfotipe ektomikoriza famili Dipterocarpaceae di Indonesia khususnya Hutan Penelitian Haurbentes perlu dilakukan.
2
Tujuan Penelitian
Penelitian ini bertujuan untuk mempelajari morfotipe ektomikoriza famili Dipterocarpaceae di Hutan Penelitian Haurbentes, Bogor
METODE
Bahan dan Alat
Bahan yang digunakan ialah sampel akar dan tanah dari rizosfer tumbuhan famili Dipterocarpaceae di Hutan Penelitian Haurbentes, dan larutan FAA (Formaline -Acetic acid-Alcohol), sedangkan peralatan yang digunakan ialah mikroskop stereo, vorteks, tabung 1.5 mL, cawan petri, kamera mikroskop digital (Optilab Camera Microscope), dan plastik klip.
Lokasi Pengambilan Sampel Penelitian
Pengambilan sampel dilakukan di Hutan Penelitian Haurbentes, Bogor. Lokasi hutan terletak pada Kecamatan Curug-Jasinga, Kabupaten Bogor atau 60 km ke arah Rangkasbitung dengan letak astronomis 6°32’ LS - 6°33’ LS dan 106°26’ BT - 106°27’ BT.
Prosedur Koleksi Sampel Akar dan Tanah
Pengambilan sampel dilakukan dengan cara membuat petak percobaan seluas 100 m x 100 m, selanjutnya dilakukan pembuatan lubang bor berdiameter 5 cm dengan kedalaman 15 cm. Pembuatan lubang bor dilakukan secara acak sebanyak 48 lubang bor dengan radius sekitar 10 m. Sampel tanah yang berisi akar selanjutnya dimasukkan ke dalam plastik klip. Spesies tumbuhan dari famili Dipterocarpaceae yang tumbuh di sekitar pembuatan lubang bor dicatat sebagai data tanaman inang yang terkolonisasi ektomikoriza. Selanjutnya, sampel disimpan pada suhu 4-6 ºC hingga siap digunakan. Parameter yang diukur pada saat koleksi sampel ialah kondisi lingkungan berupa suhu, kelembapan, dan intensitas cahaya matahari.
Preparasi Sampel dan Analisis Morfotipe Ektomikoriza
Akar rambut yang diperoleh pada setiap lubang bor dipisahkan dari tanahnya. Sampel akar kemudian dicuci dengan air mengalir, dibersihkan dari tanah atau pasir yang tersisa, lalu dibilas dengan akuades. Akar yang telah bersih selanjutnya dianalisis morfotipe ektomikoriza yang terbentuk dengan menggunakan mikroskop stereo dan kunci identifikasi ektomikoriza Agerer (1996). Berbagai morfotipe yang diperoleh selanjutnya dikelompokkan berdasarkan bentuk percabangan, warna,
3
mantel, dan rizomorf yang terbentuk, kemudian dilakukan penghitungan terhadap persen kolonisasi dan dominasinya menggunakan metode garis sentuh (gridline intersect method) dengan petak berukuran 0.5 cm x 0.5 cm. Persen kolonisasi (C) dan kelimpahan (A) dihitung menggunakan metode Brundrett et al. (1996) dengan menggunakan rumus sebagai berikut :
C = Jumlah akar terkolonisasi
Total akar teramati x 100%
A = Jumlah morfotipe
Jumlah seluruh morfotipe x 100
%
HASIL DAN PEMBAHASAN
Hasil
Kondisi Lingkungan Hutan Penelitian Haurbentes, Bogor
Hasil pengukuran kondisi lingkungan di Hutan Penelitian Haurbentes menunjukkan bahwa kondisi tanah pada wilayah tersebut tergolong tanah masam dengan pH 3.8-5.8, rata-rata intensitas cahaya sebesar 638.65 lux, suhu 32.56 oC, dan kelembapan relatif sebesar 63.56 %. Jumlah spesies tumbuhan di dalam plot percobaan yang terdiri dari 48 lubang bor ialah Shorea selanica Blume, Shorea pinanga Scheff, Shorea stenoptera Burck, Shorea ovalis, Shorea leprosula Miq., Shorea seminis (de Vriese) Sloot., Hopea mengerawan Miq., dan Vatica odorata (Griff.) Sym. (Lampiran 1). Setiap lubang bor berada diantara satu sampai tiga spesies tumbuhan famili Dipterocarpaceae.
Analisis Morfotipe Ektomikoriza
Ke 48 lubang bor sampel memiliki akar yang bersimbiosis dengan cendawan ektomikoriza dengan nilai kolonisasi yang cukup bervariasi. Nilai terendah diperoleh pada lubang bor nomor 18 yaitu kolonisasi sebesar 14.29 % dengan spesies tumbuhan S. pinanga Scheff, sedangkan nilai tertinggi pada lubang bor nomor 5 yaitu sebesar 56.83% dengan tumbuhan inang S. stenoptera Burck dan S. selanica Blume (Gambar 1). Rata-rata persen kolonisasi ektomikoriza dari 48 lubang bor pada famili Dipterocarpaceae di Hutan Penelitian Haurbentes sebesar 34.71 % dengan total panjang akar yang terkolonisasi ektomikoriza sebesar 2.15 x 102 cm dari 6.18 x 102 cm sampel akar yang teramati. Total panjang akar tertinggi yang terkolonisasi ektomikoriza diperoleh dari lubang bor nomor 28 yaitu sebesar 11.00 cm dengan tanaman inang S. leprosula Miq., sedangkan total panjang akar terendah yaitu sebesar 1.57 cm berasal dari lubang bor nomor 18 dengan tanaman inang S. pinanga (Gambar 2).
4
Hasil karakterisasi morfotipe ektomikoriza dari 48 sampel akar Dipterocarpaceae di Hutan Penelitian Haurbentes berdasarkan Colour Atlas of Ectomycorrhizae (Agerer 1996) diperoleh dua belas morfotipe ektomikoriza yang berbeda. Ke 12 morfotipe tersebut ialah morfotipe tidak bercabang hitam lurus, tidak bercabang hitam bengkok, tidak bercabang krem bengkok, monopodial piramida cokelat lurus, monopodial menyirip hitam bergelombang, monopodial menyirip cokelat lurus, monopodial menyirip cokelat bengkok, monopodial menyirip cokelat bergelombang, menyirip tidak beraturan hitam bergelombang, menyirip tidak beraturan cokelat bergelombang, menyirip tidak beraturan cokelat bengkok, dan menyirip tidak beraturan krem bengkok (Gambar 3).
0,00 10,00 20,00 30,00 40,00 50,00 60,00 70,00 1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33 35 37 39 41 43 45 47 K o lo ni sa si e kto m iko ri za ( % )
Lubang bor ke-
P anj a n g aka r te rk o lo ni sa si ( c m )
Lubang bor ke-
Gambar 21 Persen kolonisasi ektomikoriza berdasarkan lubang bor dari 48 sampel di Hutan Penelitian Haurbentes, Bogor
0,00 2,00 4,00 6,00 8,00 10,00 12,00 1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33 35 37 39 41 43 45 47
Gambar32 Total panjang akar tumbuhan famili Dipterocarpaceae yang terkolonisasi ektomikoriza berdasarkan lubang bor dari 48 sampel di Hutan Penelitian Haurbentes, Bogor
5 a b c d e f g h i j k l 2.5 mm 5 mm 5 mm 5 mm 5 mm 5 mm 5 mm 5 mm 5 mm 5 mm 5 mm 5 mm
Gambar43 Dua belas morfotipe ektomikoriza yang diperoleh dari 48 sampel lubang bor di Hutan Penelitian Haurbentes, Bogor
Keterangan :
(a) morfotipe tidak bercabang hitam lurus, b) morfotipe tidak bercabang hitam bengkok, c) morfotipe tidak bercabang krem bengkok, d) morfotipe monopodial piramida cokelat lurus, e) morfotipe monopodial menyirip hitam bergelombang, f) morfotipe monopodial menyirip cokelat lurus, g) morfotipe monopodial menyirip cokelat bengkok, h) morfotipe monopodial menyirip cokelat bergelombang, i) morfotipe menyirip tidak beraturan hitam bergelombang, j) morfotipe menyirip tidak beraturan cokelat bergelombang, k) morfotipe menyirip tidak beraturan cokelat bengkok, dan l) morfotipe menyirip tidak beraturan krem bengkok
Data keragaman ektomikoriza dari famili Dipterocarpaceae di Hutan Penelitian Haurbentes berdasarkan jenis morfotipe yang diperoleh dari 48 sampel lubang bor tersaji pada Tabel 1. Berdasarkan jenis morfotipe yang terbentuk, morfotipe yang mempunyai nilai frekuensi persen kolonisasi tertinggi ialah morfotipe menyirip tidak beraturan cokelat bergelombang yaitu 30.21 %, sedangkan nilai terendah diperoleh dari morfotipe monopodial menyirip hitam bergelombang sebesar 0.60 %.
7 7 No Je ni s m or fo tipe Ju m la h po to nga n a ka r P anj an g aka r Fr eku en si T er ko lo ni sa si ut uh * Tot al T er ko lo ni sa si Tot al T er ko lo ni sa si R ata -ra ta sa tua n K olo ni sa si K eli m pa ha n (b ua h) -- -- ---- ( cm ) - (% ) - - --1 T ida k be rc ab an g hi ta m lur us 2 10 3 3. 93 1. 81 0. 55 0. 81 0. 55 2 T ida k be rc ab an g hi ta m b en gko k 4 25 4 9. 82 1. 57 0. 79 0. 90 0. 73 3 T ida k be rc ab an g kr em b en gko k 3 10 4 3. 93 1. 57 0. 52 1. 08 0. 73 4 M on op odi al pir am id a c oke la t lur us 2 10 4 3. 93 1. 57 0. 79 1. 08 0. 73 5 Mo no po di al m enyi rip hi ta m be rge lo m ba ng 2 18 4 7. 07 1. 57 0. 79 0. 60 0. 37 6 M on op odi al m enyi rip co ke la t lur us 82 490 122 192. 50 47. 93 0. 58 26. 64 22. 67 7 M on op odi al m enyi rip co ke la t be ngko k 6 28 8 11. 00 3. 14 0. 52 2. 29 1. 46 8 M on op odi al m enyi rip co ke la t be rge lo m ba ng 3 20 6 7. 86 2. 36 0. 79 0. 81 1. 10 9 M enyi rip tid ak be ra tur an hi ta m be rge lo m ba ng 27 367 140 144. 18 55. 00 2. 04 24. 66 25. 59 10 M enyi rip tid ak be ra tur an c oke la t be rge lo m ba ng 45 436 178 171. 29 69. 93 1. 55 30. 21 32. 54 11 M enyi rip tid ak be ra tur an c oke la t be ngko k 13 148 70 58. 14 27. 50 2. 12 9. 95 12. 80 12 M enyi rip tid ak be ra tur an kr em be ngko k 2 11 4 4. 32 1. 57 0. 79 0. 98 0. 73 Tot al 19 1 1573 54 7 617. 96 214. 89 11. 83 100. 00 100. 00 * ju ml ah tot al aka r t er ko lo ni sa si ekt om iko riz a y ang di pe ro le h da ri 48 lu ba ng bo r s eb elu m d ipo to ng -poto ng un tuk pe nghi tu nga n a ka r de nga n m eto de ga ris se ntuh 9 T a b e l 1 1 K e ra ga m a n m o rf o ti p e , p e rs e n ta se k o lo n is a si d a n k e lim p a h a n e k to m ik o ri za p a d a a k a r tu m b u h a n , d a n p a n ja n g a k a r y a n g te rk o lo n is a si c e n d a w a n e k o m ik o ri za f a m ili D ip te ro c a rp a c e a e d a ri 4 8 s a m p e l lu b a n g b o r d i H u ta n P e n e lit ia n H a u rb e n te s, B o go r d e n ga n m e n g gu n a k a n m e to d e g a ri s se n tu h
7 7 Frekuensi persentase kelimpahan morfotipe ektomikoriza yang diperoleh di Hutan Penelitian Haurbentes diperoleh nilai tertinggi dari jenis menyirip tidak beraturan cokelat bergelombang sebesar 32.54 % dan terendah yaitu sebesar 0.37 % dari jenis morfotipe monopodial menyirip hitam bergelombang. Panjang akar terkolonisasi berasal dari morfotipe menyirip tidak beraturan cokelat bergelombang sebesar 69.93 cm, sedangkan kolonisasi akar terendah sebesar 1.57 cm. Rata-rata panjang akar terkolonisasi ektomikoriza terbesar diperoleh dari morfotipe menyirip tidak beraturan cokelat bengkok yaitu sebesar 2.12 cm, sedangkan yang terpendek diperoleh ialah morfotipe monopodial menyirip hitam bergelombang sebesar 0.37 cm.
Pembahasan
Hutan daratan rendah atau hutan tropis basah di Indonesia dikenal dengan istilah hutan pamah atau kerangas. Karakteristik khusus pada kawasan ini ialah kondisi tanah yang miskin unsur hara, pH rendah, aktifitas dekomposisi bahan organik yang rendah, lapisan humus yang tipis, dan kandungan nitrogen, fosfor, serta air yang rendah (Moyersoen et al. 2001). Namun demikian, kondisi tersebut tidak mempengaruhi pembentukkan berbagai spesies tumbuhan dari famili Dipterocarpaceae untuk tumbuh di Indonesia, karena tumbuhan tersebut secara obligat membentuk simbiosis mutualisme ektomikoriza untuk mencukupi nutrisi yang dibutuhkan dalam pertumbuhannya (Smith dan Read 2008). Selain itu, famili Dipterocarpaceae mampu hidup pada jenis tanah podsolik merah kuning yang terdapat pada daerah dengan iklim basah dengan kelembaban tinggi dan ketinggian tempat 0 – 800 m dpl., serta curah hujan di atas 2000 mm/tahun dengan rentang musim kemarau yang pendek (CIFOR 1998). Hutan Penelitian Haurbentes merupakan tempat tumbuh yang memenuhi kriteria optimal tempat tumbuh berbagai spesies dari famili Dipterocarpaceae, terlihat dari aktivitas pembungaan dan produktivitas buah yang baik dari famili Dipterocarpaceae di Hutan Penelitian ini (Suzuki dan Gadrinab 1989). Fakta tersebut diperkuat dengan publikasi Balitbanghut (2007) yang menyatakan bahwa Hutan Penelitian Haurbentes merupakan hutan hujan tropis dataran rendah (250 m dpl) yang didominasi oleh jenis tanah podsolik merah kuning, regosol, dan acid brown forest soil (tanah dengan pH masam), dengan curah hujan rata-rata sebesar 4.276 mm/tahun. Tidak hanya itu, Mandang dan Kagemori (2003) mengungkapkan bahwa wilayah Jasinga-Rangkasbitung merupakan daerah hutan tua dan pusat perkembangan tanaman hutan famili Dipterocarpaceae, yang dibuktikan dengan ditemukannya batuan fosil tanaman hutan dari Dipterocarpoxylon sp. pada lapisan tanah Ploicene di wilayah Jasinga Bogor yang tidak jauh berbeda dengan beberapa fosil-fosil yang diperoleh di hutan Sumatra dan Kalimantan.
Famili Dipterocarpaceae yang terdapat di Hutan Penelitian Haurbentes meliputi H. mengerawan Miq., S. selanica Blume., S. stenoptera Burck., S. stenoptera Burck. forma, S. pinanga Scheff, S. seminis (de Vriese) Sloot., S. leprosula Miq., Shorea palembanica, Shorea mechisopteryx Ridley, dan Shorea platyclados V Sl (Istomo et al. 1999). Keberadaan beberapa spesies tersebut dapat dibuktikan dengan ditemukannya delapan spesies tumbuhan dari famili Dipterocarpaceae pada petak percobaan seluas 104 m2.
8
Hasil pengukuran kondisi lingkungan pada saat pengambilan sampel diperoleh kondisi yang sangat mendukung simbiosis ektomikoriza yakni pH tanah 3.8-5.8 dengan suhu lingkungan berkisar 32.56 oC. Kondisi ini sangat mendukung perkembangan cendawan ektomikoriza dan pertumbuhan tanaman famili Dipterocarpaceae di Hutan Penelitian Haurbentes. Umumnya ektomikoriza di wilayah hutan tropis berkembang dengan baik pada kondisi tanah masam dengan pertumbuhan optimal pada pH 4-6. Bahkan tidak sedikit cendawan yang mampu hidup pada pH sangat masam yaitu 2.7, namun dengan jumlah kolonisasi akar yang jauh menurun dibanding pH optimal (Dighton dan Skeffington 1987). Selain pH, faktor suhu lingkungan juga berpengaruh terhadap perkembangan ektomikoriza di alam. Suhu optimal untuk pertumbuhan cendawan ektomikoriza berkisar antara 18-27 oC, namun demikian terdapat ektomikoriza yang mampu hidup pada suhu ekstrim seperti suhu -2 oC atau bahkan 40 oC (Fakuara 1988; Smith dan Read 2008).
Marx et al. (1992) membagi persentase kolonisasi cendawan ektomikoriza menjadi empat kelompok, yakni sangat baik (75-100 %), baik (50-74 %), sedang (24-49 %), dan rendah (1-24%). Berdasarkan standarisasi tersebut, maka secara umum persen kolonisasi ektomikoriza dari 48 lubang bor famili Dipterocarpaceae di Hutan Penelitian Haurbentes, tergolong kelompok sedang (34.71%). Keragaman morfotipe ektomikoriza yang diperoleh pada Hutan Penelitian Haurbentes yaitu 12 morfotipe. Keragaman tersebut diperoleh berdasarkan tipe percabangan, bentuk ujung akar, warna akar, bentuk khas permukaan mantel, dan bentuk rizomorf atau hifa yang berada di sekitar akar yang terkolonisasi cendawan ektomikoriza.
Dalam penelitian ini tidak dilakukan isolasi cendawan yang bersimbiosis dengan akar tanaman Dipterocarpaceae yang diperoleh, namun karakterisasi morfotipe akar famili Dipterocarpaceae yang bersimbiosis dengan cendawan ektomikoriza dapat digunakan untuk menduga genus cendawan ektomikoriza yang bersimbiosis. Terdapat tiga macam keuntungan menggunakan metode karakterisasi morfotipe ektomikoriza dalam estimasi langsung kekayaan dan keragaman spesies ektomikoriza. Pertama, struktur yang berbeda pada akar berektomikoriza dapat saja menunjukkan bahwa cendawan yang bersimbiosis dengan akar tersebut berbeda spesies. Sehingga dapat dipastikan bahwa semakin beragam struktur morfotipe cendawan, maka semakin tinggi pula keragaman spesies cendawan yang diperoleh (Agerer 1996). Kedua, struktur reproduktif cendawan (tubuh buah) ektomikoriza biasanya ditemukan dalam jumlah yang sangat sedikit, jika pun ada hanya pada waktu-waktu tertentu, sehingga tidak efektif dalam penelitian mengenai kekayaan dan keragaman ektomikoriza, bahkan tidak sedikit ektomikoriza yang sama sekali tidak memproduksi tubuh buah. Ketiga, tanpa bergantung pada penemuan tubuh buah di alam, karena sangat sulit ditemukan tubuh buah di alam dalam kondisi yang segar (Horton dan Bruns 2001; Moser 2005; Nara 2006).
Selain nilai persentase kolonisasi dan kelimpahan ektomikoriza, parameter ektomikoriza yang penting lainnya ialah keragaman morfotipe. Hasil identifikasi keragaman ektomikoriza di Hutan Penelitian Haurbentes diperoleh 12 morfotipe berbeda dari 48 lubang bor. Apabila dibandingkan dengan hasil penelitian Sukarno et al. (2013) yang melakukan penelitian ektomikoriza di Hutan Penelitian Haurbentes meskipun dengan metode yang berbeda yakni metode runut akar, diperoleh beberapa morfotipe yang memiliki ciri-ciri serupa dengan hasil penelitian ini. Sukarno et al. (2013) mendapatkan 43 kelompok cendawan ektomikoriza dari 50 jenis morfotipe ektomikoriza di Hutan Penelitian Haurbentes. Beberapa
9
morfotipe tersebut diantaranya ialah morfotipe sederhana hitam yang dikolonisasi oleh cendawan Russula sp., morfotipe menyirip monopodial cokelat dikolonisasi Russula sp., dan morfotipe menyirip tidak beraturan dikolonisasi cendawan Sebacina sp. dan Tomentella sp. sebagai cendawan ektomikoriza yang mendominasi. Hal serupa juga dikemukakan Becerra et al. (2004) yang meneliti keragaman ektomikoriza berdasarkan morfotipenya pada tanah masam di Argentina, diperoleh beberapa morfotipe yang memiliki ciri-ciri serupa dengan hasil penelitian ini, beberapa diantaranya ialah morfotipe tidak bercabang umumnya dikolonisasi oleh Lactarius omphaliformis, Naucoria escharoides, atau Tomentella sp., morfotipe monopodial menyirip dikolonisasi cendawan Naucoria sp. Gyrodon sp., dan Lactarius sp., serta morfotipe menyirip tidak beraturan yang dikolonisasi cendawan Tomentella sp., Russula sp., dan Alnirhiza sp. Selanjutnya, Moser et al. (2009) melaporkan bahwa morfotipe monopodial piramida cokelat dikolonisasi oleh cendawan Boletus sp. atau Genea sp., morfotipe monopodial menyirip menuju menyirip tidak beraturan dikolonisasi oleh cendawan Inocybe sp., Sebacina sp., atau Tomentella sp.. Fakta lain menyebutkan bahwa genus ektomikoriza yang umumnya ditemukan pada hutan matang berupa genus Russula, Cortinarius, dan Amanita (Nara 2006). Hal ini memperkuat dugaan bahwa famili Dipterocarpaceae di Hutan Penelitian Haurbentes dikolonisasi ektomikoriza dengan morfotipe yang mendominasi berupa morfotipe monopodial menyirip menuju menyirip tidak beraturan.
SIMPULAN
Hasil penelitian menunjukkan bahwa delapan spesies tumbuhan dari famili Dipterocarpaceae di Hutan Penelitian Haurbentes dikolonisasi oleh cendawan ektomikoriza. Dua belas morfotipe ektomikoriza berhasil diperoleh dari 191 potong akar yang terkolonisasi ektomikoriza yang berasal dari 48 lubang bor. Ke-12 morfotipe tersebut ialah morfotipe tidak bercabang hitam lurus, tidak bercabang hitam bengkok, tidak bercabang krem bengkok, monopodial piramida cokelat lurus, monopodial menyirip hitam bergelombang, monopodial menyirip cokelat lurus, monopodial menyirip cokelat bengkok, monopodial menyirip cokelat bergelombang, menyirip tidak beraturan hitam bergelombang, menyirip tidak beraturan cokelat bergelombang, menyirip tidak beraturan cokelat bengkok, dan menyirip tidak beraturan krem bengkok. Persentase kolonisasi ektomikoriza termasuk ke dalam kelompok klasifikasi sedang (34.71 %) dengan kelimpahan kolonisasi akar tertinggi diperoleh dari morfotipe menyirip tidak beraturan cokelat bergelombang yaitu sebesar 32.54 %, sedangkan kelimpahan terendah dari morfotipe monopodial menyirip hitam bergelombang yaitu sebesar 0.37 %. Total panjang akar terkolonisasi ektomikoriza sebesar 2.15 x 102 cm dari 6.18 x 102 cm sampel akar famili Dipterocarpaceae yang teramati.
10
DAFTAR PUSTAKA
[Balitbanghut] Badan Penelitian dan Pengembangan Kehutanan. 2007. Kawasan Hutan dengan Tujuan Khusus Haurbentes. [Internet]. Bogor: [diunduh 2014 Mar 1]. Tersedia pada: http://www.dephut.go.id/index.php/news/details/951. Agerer R. 1996. Colour Atlas of Ectomycorrhizae. Munchen (DE): Einhorn,
Schwabish Gmund D-73525.
Amornpitak TW, Vichitsoonthonkul T, Tanticharoen M, Cheevadhanarak S, Ratchadawong S. 2006. Diversity of ectomycorrhizal fungi of Dipterocarpaceae in Thailand. J Biosci. 6: 1059-1064.
Becerra A, Pritsch K, Arrigo N, Palma M, Bartoloni N. 2004. Ectomycorrhizal colonization of Alnus acumiata Kunth. in Northwestern Argentina in relation to season and soil parameters. Ann For Sci. 65: 325-332. doi: 10.1051/forest:2005027.
Brearley FQ. 2012. Ectomycorrhizal association of the Dipterocarpaceae. Biotropica. 49: 637-648
Brundrett M. 2004. Diversity and classification of mycorrhizal associations. Biol Rev. 79:473–495.
Brundrett M, Bougher N, Dell B, Grove T, Malajczuk N. 1996. Working With Mycorrhizae in Forestry and Agriculture. Sydney (AU): Monograph ACIAR. [CIFOR] Center for International Forestry Research. 1998. A Review of
Dipterocarps. Appanah S, Turnbull JM, editor. Bogor (ID): CIFOR.
Dighton, Skeffington RA. 1987. Effects of artificial acid precipitation on the mycorrhizas of Scots pine seedlings. New Phytologist. 107: 191-202.
Fakuara MY. 1988. Mikoriza, Teori dan Kegunaan dalam Praktek. Bogor (ID): PAU IPB.
Horton TR, Bruns TD. 2001. The molecular revolution in ectomycorrhizal ecology: peeking into the black box. Mol Ecol. 10: 1855-1871.
Istomo, Cahyo W, Nurul H. 1999. Evaluasi pertumbuhan tanaman meranti (Shorea spp.) di Haurbentes BKPH Jasinga KPH Bogor Perum Perhutani Unit III Jawa Barat. J Man Hut Trop. 2: 13-32.
Mandang YI, Kagemori N. 2003. A fossil wood of Dipterocarpaceae from Pliocene deposit in the West Region of Java Island, Indonesia. Biodiversitas. 5 (1): 28-35.
Marx DH, Ruehle Jl, Cordel CE.1992. Methods for Studying Nursery and Field Response of Trees to Spesific Ectomycorrhizal. Di dalam: Norris JR, Read DJ, Varma AK. Methods in Microbiology: Techniques for Mycorrhizal Research. San Diego (US). Academic Pr. hlm 383-340.
Moser AM. 2005. Comparison of ectomycorrhizas of Quercus garryana (Fagaceae) on serpentine and non-serpentine soils in Southwestern Oregon. Am J Bot. 92 (2): 224-230.
Moser AM, Frank JL, D’Allura JA, Southworth D. 2009. Ectomycorrhizal communities of Quercus garryana are similar on serpentine and non-serpentine soils. Plant Soil. 305: 185-194.
Moyersoen B. 2006. Pakaraimaea dipterocarpacea is ectomycorrhizal, indicating an ancient Gondwana land origin for the ectomycorrhizal habit in Dipterocarpaceae. New Phytologist. 172: 759-762.
11
Moyersoen B, Becker P, Alexander IJ. 2001. Are ectomycorrhizas more abundant than arbuscular mycorrhizas in tropical health forest? New Phytologist. 150 :591-599.
Nara K. 2006. Ectomicorrhyzal networks and seedling establishment during early primary succession. New Phytologist. 169:169-178.
Newman MF, Burgess PF, Whitmore TC. 1999. Pedoman Identifikasi Pohon-pohon Dipterocarpaceae Jawa sampai Nugini. Bogor (ID): Prosea Indonesia. Noor M. 2009. Simbion jamur ektomikoriza pada anakan Shorea spp. di rumah kaca
pada umur 7 bulan. J P Diptero. 3 (1) 1-7.
Olsson PA, Munzenberger B, Mahmood S, Erland S. 2000. Molecular and anatomical evidence for three way association between Pinus sylvestris and the ectomycorrhizal fungi Suillus bovinus and Gomphidius roseus. J Mycol. 16 (1): 8-9.
Prameswari D. 2004. Pengaruh inokulasi cendawan ektomikoriza dan media tumbuh terhadap pertumbuhan Shorea javanica K & V [tesis]. Bogor (ID). Institut Pertanian Bogor.
Riniarti M. 2002. Perkembangan kolonisasi ektomikoriza dan pertumbuhan semai Dipterocarpaceae dengan pemberian asam oksalat dan asam humat serta inokulasi ektomikoriza [tesis]. Bogor (ID). Institut Pertanian Bogor.
Smith ME, Henkel TW, Wehling JK, Fremies AK, Clarke HD, Vilgalys R. 2013. The ectomycorrhizal fungal community in a Neotropical Forest dominated by the endemic Dipterocarpaceae. PLoS ONE 8(1): 1-13. doi: 10.1371/journal.pone.0055160.
Smith SE, Read DJ. 2008. Mycorrhizal symbiosis. 3th ed. London (GB): Academic Pr.
Sukarno N, Srilistiyowati, Nurcahyo OD, Retnowati A, and Nara K. 2013. Diversity of Shorea ectomycorrhiza. Japanese Mycorrhizal Symposium. 2013 Nov 16; Sendai (JP).
Suzuki E, Gadrinab LU. 1989. Fruit production of a six year old Shorea stenoptera plantation at Haurbentes, Bogor, Indonesia. Biotr. 2: 1-7.
Turjaman M, Tamai Y, Segah H, Limin SH, Cha JY, Osaki M, Tawaraya K. 2005. Inoculation with the ectomycorrhizal fungi Pisolithus arhizus and Scleroderma sp. improves early growth of Shorea pinanga nursery seedlings. J Int New Forest. 30:67-73.
Turjaman M, Tamai Y, Segah H, Limin SH, Cha JY, Osaki M, Tawaraya K. 2006. Increase in early growth and nutrient uptake of Shorea seminis seedlings inoculated with two ectomycorrhizal fungi. J Trop Forest Scie. 18(4):166-172.
Turjaman M, Tamai Y, Santoso E, Susanto A, Gaman S, Limin SH, Tamai Y, Osaki M, Tawaraya K. 2011. Ectomycorrhizal fungi promote Growth of Shorea balangeran in degraded peat swamp forest. Wetland Ecol Man. 19:331-339.
12
LAMPIRAN
Lampiran 1 Daftar lubang bor, nama tanaman inang, dan kondisi lingkungan di Hutan Penelitian Haurbentes, Bogor
No
Core Spesies tumbuhan
Intensitas (lux) Kelembapan (% RH) Suhu (oC) 1 S. selanica Blume 187 63.40 30.40 S. pinanga Scheff 2 S. selanica Blume 1350 65.30 30.40 S. pinanga Scheff 3 S. stenoptera Burck 1430 65.00 30.80 4 S. stenoptera Burck 1373 58.20 33.20 S. pinanga Scheff 5 S. stenoptera Burck 1429 58.20 32.70 S. selanica Blume
6 V. odorata (Griff.) Sym. 453 61.50 32.80 H. mengerawan Miq. 7 H. mengerawan Miq. 373 64.20 32.60 S. stenoptera Burck 8 S. ovalis 1770 67.60 32.90 S. pinanga Scheff 9 S. leprosula Miq. 1076 63.90 32.40 H. mengerawan Miq. 10 S. leprosula Miq. 575 62.70 32.20 H. mengerawan Miq. 11 S. pinanga Scheff 283 67.00 32.20 S. stenoptera Burck 12 S. pinanga Scheff 300 63.60 32.10 S. stenoptera Burck 13 S. pinanga Scheff 272 64.30 32.40 S. stenoptera Burck 14 S. stenoptera Burck 400 70.10 32.60 15 S. stenoptera Burck 499 66.90 33.00 16 S. stenoptera Burck 304 65.80 32.70 17 S. seminis (de Vriese) Sloot. 225 70.80 32.70 18 S. pinanga Scheff 248 62.10 33.00 19 S. seminis (de Vriese) Sloot. 545 64.80 32.70 20 S. stenoptera Burck 575 68.20 32.40 21 S. pinanga Scheff 1157 69.50 32.20 22 S. seminis (de Vriese) Sloot. 224 60.70 31.90 23 S. seminis (de Vriese) Sloot. 373 59.70 32.50 24 S. seminis (de Vriese) Sloot. 455 65.40 32.80 25 S. seminis (de Vriese) Sloot. 266 62.90 32.70
13
Lampiran 1 (Lanjutan) No
Core Spesies tumbuhan
Intensitas (lux) Kelembapan (% RH) Suhu (oC) 26 S. seminis (de Vriese) Sloot. 837 60.30 33.40 27 S. stenoptera Burck 299 65.10 34.00 28 S. leprosula Miq. 1958 60.00 33.50 29 S. leprosula Miq. 221 62.80 33.40 30 S. leprosula Miq. 215 59.40 33.80 31 S. leprosula Miq. 1249 62.00 33.20 32 S. leprosula Miq. 1201 65.30 32.60 33 S. leprosula Miq. 630 65.20 33.00 34 S. stenoptera Burck 493 60.30 33.40 35 S. stenoptera Burck 509 58.60 33.30 36 S. pinanga Scheff 298 60.70 33.20 37 S. pinanga Scheff 277 60.20 33.70 38 S. stenoptera Burck 991 59.80 33.10 39 S. stenoptera Burck 268 61.40 32.70 40 S. pinanga Scheff 872 61.40 31.60 41 S. pinanga Scheff 259 63.80 32.90 42 S. pinanga Scheff 492 62.90 32.70 43 S. pinanga Scheff 564 62.40 32.50 44 S. pinanga Scheff 709 63.70 31.40 45 S. pinanga Scheff 705 63.30 31.80 46 S. pinanga Scheff 466 66.00 31.80 47 S. pinanga Scheff 500 66.00 31.70 48 S. seminis (de Vriese) Sloot. 500 64.33 31.70
15
RIWAYAT HIDUP
Penulis dilahirkan di Cianjur pada tanggal 15 Juni 1990 dari ayah Tatang Mulyana dan ibu Ros Rosita. Tahun 2007 penulis lulus dari SMA Negeri 1 Sukaresmi Cianjur, dan pada tahun yang sama lulus seleksi masuk IPB melalui jalur Undangan Seleksi Masuk Institut Pertanian Bogor (USMI) dan diterima di Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam.
Penulis mempunyai pengalaman sebagai asisten peneliti di Laboratorium Biokontrol Balai Penelitian Tanaman Hias Departemen Pertanian RI, teknisi Laboratorium Mikrobiologi Center of Tropical Animal Studies Lembaga Penelitian dan Pengabdian kepada Masyarakat (LPPM) IPB, melaksanakan studi lapangan di Situ Gunung pada tahun 2009, melaksanakan praktik lapangan di Laboratorium Biokontrol Balithi pada tahun 2011. Penulis juga aktif menjadi staf Departemen Sosial dan Kesejahteraan Mahasiswa Badan Eksekutif Mahasiswa (BEM) TPB IPB Kabinet Oryza Sativa pada tahun 2007, staf Kementerian Kebijakan Nasional BEM KM IPB Kabinet IPB Bersatu pada tahun 2008, staf Kementerian Kebijakan Daerah BEM KM IPB Kabinet Generasi Inspirasi pada tahun 2009, Koordinator BEM se-Bogor pada tahun 2010, Koordinator Komunikasi dan Informasi Korps Sukarela (KSR) PMI Cianjur pada tahun 2008-2009, dan Ketua Karang Taruna Komplek Pertanian Landbauw 2011-sekarang.