3 MORFOGENETIKA PINUS MERKUSII
KANDIDAT BOCOR GETAH
3.1 Pendahuluan
Hasil analisis struktur produksi getah dan struktur pertumbuhan pinus kandidat bocor getah di KBS Perum Perhutani (Bab 2) menunjukkan adanya interaksi genotipe dengan lingkungan KBS tertentu (interaksi GxE). Analisis morfogenetika diperlukan untuk mengetahui besaran proporsi antara faktor genetika dan faktor lingkungan dalam mempengaruhi karakter produksi getah. Namun informasi karakter morfogenetika yang berhubungan dengan produksi getah di KBS Cijambu sampai saat ini belum diperoleh, padahal informasi karakter morfogenetik penting untuk kegiatan karakterisasi dan pemuliaan pinus kandidat bocor getah dimasa mendatang.
Informasi karakter morfogenetika kandidat bocor getah dapat diperoleh melalui serangkaian evaluasi fenotipik dan analisis genetika dengan bantuan penanda molekuler. Beberapa variabel genetika yang dapat digunakan sebagai acuan pertimbangan agar seleksi efektif dan efisien adalah keragaman genetika, heritabilitas, korelasi dan pengaruh dari karakter-karakter yang erat hubungannya dengan hasil pemuliaan yang diinginkan (Borojevic 1990; Falconer dan Mackay 1996). Adanya keragaman genetika mengindikasikan terdapatnya perbedaan nilai antar individu genotipe dalam populasi dan merupakan syarat keberhasilan seleksi terhadap karakter yang diinginkan. Nilai heritabilitas mampu menggambarkan efektivitas seleksi genotipe berdasarkan penampilan fenotipenya, sedangkan informasi korelasi antar karakter fenotipe diperlukan dalam seleksi tanaman, untuk mengetahui karakter yang dapat dijadikan petunjuk seleksi terhadap produktivitas getah yang tinggi (Tambarussi et al. 2010).
Sebagai langkah lanjutan dari evaluasi fenotipik, kegiatan verifikasi dengan bantuan penanda molekuler juga dilakukan pada penelitian ini. Hal tersebut sebagai langkah antisipasi untuk mengatasi kelemahan evaluasi fenotipik yang cenderung terpengaruh oleh tahap perkembangan tanaman dan lingkungan, sehingga sering tidak dapat membedakan antara genotipe yang diamati. Untuk mengatasi permasalahan tersebut penanda molekuler seperti RFLP, RAPD, AFLP dan mikrosatelit dapat digunakan. Penanda mikrosatelit dipilih pada penelitian ini karena merupakan kodominan marker, memiliki tingkat polimorfisme yang tinggi, dan terdistribusi secara acak pada genom (Tautz 1989; Powell et al.1996; Lehman 1998)
Penelitian mengenai evaluasi fenotipik P.merkusii pada awal pembangunan KBS yang dilakukan oleh Leksono (1996), pada P. pinaster (Tadesse et al. 2001)
dan P.eliotii (Zhang et al.2010) menunjukkan bahwa karakter produksi getah
memiliki heritabilitas tinggi, namun informasi di KBS Cijambu belum diperoleh, demikian juga dengan deteksi keragaman genetika dengan penanda mikrosatelit. Oleh karena itu, penelitian mengenai “Morfogenetika Pinus merkusii kandidat bocor getah” perlu dilakukan untuk memperoleh informasi mengenai heritabilitas, keterkaitan karakter fenotipe terhadap produksi getah serta variasi genetika
melalui penanda mikrosatelit dalam rangka pengembangan kegiatan pemuliaan tanaman dimasa mendatang.
3.2 Bahan dan Metode 3.2.1 Bahan
Bahan penelitian untuk evaluasi morfogenetik merupakan tanaman uji keturunan di KBS Cijambu tahun tanam 1978-1983 dengan tujuan pemuliaan awal kelurusan batang dan pertumbuhan yang cepat. Pada awal pembangunan KBS desain yang digunakan adalah rancangan acak lengkap berblok (RCBD) dengan 5 tree line plot dan 10 blok sebagai ulangan. Setiap tahun dilakukan penanaman 200 famili yang diperoleh dari eksplorasi hutan tanaman di Jawa, namun seiring dengan kegiatan penjarangan seleksi tahap 1 dilakukan dengan fokus produksi getah. Hasil seleksi bertahap yang telah dilakukan sampai tahun 2002, tersisa 2 pohon setiap blok atau 100 famili/blok. Selanjutnya hasil uji keturunan tersebut diseleksi kembali dengan tujuan untuk memperoleh produksi getah yang tinggi. Pada saat dilakukan penelitian ini (2011) tersisa 1-2 pohon/blok yang masuk kategori kandidat bocor getah.
Seleksi tahap 2 di lokasi yang sama dengan tujuan karakter produksi getah yang hanya menemukan 110 pohon plus yang memiliki produksi tinggi dari 6 tahapan penanaman uji keturunan (1978-1983). Pada penelitian ini bahan pengujian untuk perhitungan nilai heritabilitas berasal dari 2 set tanaman uji keturunan (tahun 1982 dan tahun 1983) dengan pertimbangan kecukupan data dan kurva sebaran yang lebih lebar dibandingkan set uji keturunan tahun lainnya. Tanaman uji keturunan tahun 1982 memiliki 20 famili dengan 8 famili yang memiliki ulangan di blok yang berbeda, sedangkan tahun tanam 1983 memiliki memiliki 25 famili dengan 7 famili yang memiliki ulangan pada blok berbeda. Adapun identitas famili tahun tanam 1982 dan 1983 yang digunakan untuk penelitian disajikan pada Tabel 3.1.
Tabel 3.1 Identitas famili untuk perhitungan nilai heritabilitas
Tahun tanam 1982 Tahun tanam 1983
Famili Blok Produksi getah (g/pohon/ 3hari) Tinggi (m) Diameter (m)
Famili Blok Produksi getah (g/pohon/ 3hari) Tinggi (m) Diameter (m) A 2 107.4 30 0.41 I 7 97.7 25 0.40 4 126.1 25 0.42 8 59.2 24.5 0.38 B 2 79.4 25 0.31 J 10 97.7 24.5 0.38 5 136.3 25 0.50 7 108.8 25 0.46 C 5 90 26 0.47 K 6 93.4 23 0.38 7 99.9 28 0.39 3 132.1 25 0.52 D 8 129.9 22.5 0.48 L 8 78.4 23 0.43 9 133.9 29 0.47 5 82.3 21 0.46 E 2 99.9 28 0.38 M 6 156.9 23 0.56 1 104.1 29 0.64 8 156.9 23.5 0.48 F 4 78.6 18.5 0.37 N 7 99.8 24.5 0.46 1 106.5 27 0.44 6 120.1 25 0.46 G 2 121.6 27 0.40 O 1 75.4 22 0.38 5 146.7 25 0.40 7 97.2 21 0.41 H 8 97.1 22 0.39 7 93.3 29 0.47
Untuk analisis DNA, sampel daun dan kayu dikumpulkan dari 70 pohon yang dibedakan menjadi pohon plus kelas produksi tinggi (≥ 150 g/pohon/3 hari) sebanyak 30 sampel, kelas produksi rendah (≤ 60 g/pohon/3 hari) sebanyak 30
sampel dan pohon normal (≤ 21 g/pohon/3 hari) sebanyak 10 sampel sebagai pembanding. Sampel yang mewakili kandidat bocor getah diambil dari KBS Cijambu Sumedang, sedangkan untuk produksi normal diambil dari Bogor. Sampel selanjutnya disimpan dalam kantong plastik tertutup yang telah diisi silica gel dan disimpan pada suhu ruangan hingga dilakukan kegiatan ekstraksi DNA. Kondisi lingkungan lokasi pengambilan sampel untuk analisis genetika disajikan pada Tabel 3.2.
Tabel 3.2 Kondisi lingkungan di lokasi pengambilan sampel di KBS Cijambu dan Bogor
Lokasi Sampel Posisi geografis Ketinggian (m dpl) Suhu (oC) KBS Cijambu 6052’"S dan107045’E 900-1300 17-22 Bogor 6°35'49"S
dan106°46'51"E
250 26-30
Pada penelitian ini digunakan 9 primer mikrosatelit yang terdiri dari 7 primer hasil pengembangan Nurtjahjaningsih et al. (2005) pada P.merkusii (pm01, pm04, pm05, pm07, pm08, pm09a , pm12) dan 2 primer dari P. densiflora
(pde5) dan P.sylvestris (SPAC 11.6) yang dikembangkan oleh Soronzo et al.
(1998). Primer SPAC 11.6 dan pde5 digunakan juga untuk mengetahui
transferability serta polimorfismenya pada P. merkusii (Tabel 3.3).
Tabel 3.3 Sembilan primer mikrosatelit untuk deteksi keragaman genetika
Primer Motif Pengulangan sekuens Asesi
GeneBank pm01 (TG)12 F: AGAGAAGGCACGATTTTGTC R: TCCCACTAATCACTTTGAAAG AB201535 pm04 (TG)10 F: CTCTAAGTAGGACAAGGCCT R: CATAATCCAAGGAGTCAAGG AB201536 pm05 (TG)9 F: GAGTCTAATTGCAAACCCCA R: TGGAGATCTACCACTTTTTC AB201537
pm07 (AC)8(AT)4 F: GAATCTAAGCATATGAAATGAG
R: CTTGTTAATGCTACTAGTTATG
AB201538
pm08 (AT)2(GT)1 F: GCTTCAATCTATTGACCCCAT
R: AAAGGGGCAGCTGCTACAACCAATGG
AB201539
pm09a (AT)5(GT)18(AT)2 F: CCTTCTCATTTCGATATGCAC
R: ATTAAAGGTTATATGGGGCT
AB201540
pm12 (GT)5CT(GT)5(AT)5 F: GAACAATCATTGCGGGTCCCG
R: ATGCTGCGTTTATATGTATAAGTGTC
AB201541
Pde5 (CA)5C(CA)51 F: AACGCACCTTTCTCAATGCAC
R: ATAAAGAGGCTACATGGTCCC
ABO38258
SPAC 11.6 (CA)29(TA)7 F: CTTCACAGGACTGATGTTCA
R: TTACAGCGGTTGGTAAATG
3.2.2 Metode
3.2.2.1 Evaluasi fenotipik
Sifat yang diamati pada evaluasi fenotik meliputi produksi getah dan 14 karakter kuantitatif yang diduga terkait dengan produksi getah meliputi karakter tinggi (tinggi total dan tinggi bebas cabang), karakter tajuk (rasio diameter cabang pembentuk tajuk, panjang tajuk, lebar tajuk, jumlah cabang pembentuk tajuk, sudut cabang pertama), karakter batang (diameter batang, rasio diameter cabang dan batang, volume total, volume bebas cabang, kelurusan batang) dan karakter kulit batang (tebal kulit dan kekasaran kulit). Teknik pengukuran untuk 14 karakter fenotipik tersebut mengikuti prosedur yang dilakukan oleh penelitian lain pada pohon-pohon kehutanan Bacilieri et al. (1995); Cantini et al. (1999); Kremer
et al. (2002); Ginwal et al. (2004); Weber dan Montes (2005); Baliuckas et al.
(2005); dan Devagiri et al. (2007). Data mengenai karakter fenotipik selanjutnya dianalisis untuk memperoleh informasi mengenai keragaman genetika, heritabilitas dan korelasi, serta komponen karakter yang memiliki keterkaitan dengan produksi getah dengan bantuan software statistik.
Analisis data
Perhitungan nilai heritabilitasdan variasi genetika
Nilai heritabilitas menggambarkan besaran proporsi antara varians genetika maupun varians lingkungan dalam menentukan suatu karakter. Pada penelitian di KBS Cijambu variabilitas diduga dengan menggunakan analisis komponen varians menurut Steel dan Torrie (1995) sedangkan untuk pendugaan heritabilitas dilakukan melalui pendekatan heritabilitas dalam arti sempit (narrow sense
heritability). Pemilihan pendekatan heritabilitas dalam arti sempit karena fokus
penelitian hanya pada satu lokasi (KBS Cijambu) dengan asumsi kondisi lingkungannya homogen. Pertimbangan lain, pendekatan ini digunakan oleh hampir semua pemulia pohon (Cornelius 1994) karena sesuai untuk kondisi kebun benih semai (White et al. 2007) serta sesuai untuk tujuan pemuliaan pohon untuk peningkatan daya gabung umum dengan pendekatan varians aditif (Zobel dan Talbert 1984).
Variabilitas diduga dengan menggunakan analisis komponen varians menurut Steel dan Torrie (1995). Varians fenotipe, varians genotipe dapat dihitung berdasarkan : δ2 f = (M2-M3)/r, δ2 e = M3, δ2b = M1-M3/r dimana: M1: kuadrat tengah blok; M2 = kuadrat tengah genotip; M3 = kuadrat tengah galat (Tabel 3.4).
Tabel 3.4 Analisis varians dan harapan kuadrat tengah dari single tree plot design untuk suatu karakter
Sumber keragaman
Derajat bebas
Kuadrat tengah F-Hitung Harapan Kuadrat
Tengah (EKT)
Blok (r ) r-1 M1 M1/M3 δ2 e + r (δ2 b)
Famili (g) g-1 M2 M2/M3 δ2 e + r (δ2 g)
Galat (r-1)(g-1) M3 - δ2 e
Analisis data untuk perhitungan parameter genetika dengan kondisi tegakan seperti yang telah dideskripsikan sebelumnya, dilakukan dengan pendekatan
single tree plot. Adapun model linear untuk pengamatan individu pohon dengan
pendekatan single tree plot (Isik 2008) pada setiap tanaman uji keturunan adalah sebagai berikut:
Yijk: µ+ Bi+ Fj+eijk
Keterangan: Yijk: pengamatan individu pohon ke-k pada blok ke-i dan famili
ke-j; µ:rerata umum; Bi: pengaruh blok ke-i; Fj: pengaruh
famili ke-j;galat acak pada Yijk. Hanya 2 ulangan yang
digunakan pada penelitian.
Pada penelitian ini, pendugaan nilai heritabilitas dilakukan dengan dua pendekatan. Pendekatan pertama dengan menggabungkan famili-famili tahun tanam 1982 dan 1983, dengan asumsi perubahan produksi getah dan karakter pertumbuhan pada kurun waktu 1 tahun tidak menunjukkan perbedaan yang signifikan. Pendekatan kedua dengan membuat set masing-masing tahun tanam dengan asumsi terjadi perubahan produksi getah dan karakter pertumbuhan dengan perbedaaan 1 tahun penanaman.
Heritabilitas dalam arti sempit untuk satu lokasi diduga menggunakan analisis komponen varians dan dihitung berdasarkan rumus Wright (1976); Zobel dan Talbert (1984); Falconer dan Mackay (1996); White et al. (2007); Leksono (1996) dan Hardiyanto (1996). Tanaman hasil uji keturunan dari masing-masing famili diasumsikan merupakan hasil half sib sehingga pendugaan nilai heritabilitas dan standar deviasinya mengikuti rumus untuk pendugaan heritabilitas satu lokasi:
Heritabilitas famili :
h2f : σ 2f/ (σ 2f +σ 2b/nb+σ 2e),
dengan S.E. h2f : (S.Eσ 2f)/ (σ 2f +σ 2b/nb+σ 2e),
Heritabilitas individu:
h2 : 4δ2 f/ (σ 2f+σ 2b+ σ 2e),
dengan S.E. h2 : (4S.Eσ 2f)/ ( δ2f+σ 2b+ σ 2e),
Keterangan: h2f:heritabilitas famili; h2:heritabilitas individu; σ 2f:komponen ragam famili; σ 2b: komponen ragam blok; σ 2e: komponen
ragam galat.
Kriteria nilai heritabilitas (h2) menurut Cotteril dan Dean (1990), yaitu: tinggi jika
h2> 0.50, sedang jika 0.20 ≤ h2 ≤ 0.50 dan rendah jika h2 < 0.20. Standar deviasi komponen ragam famili (S.Eδ2) dihitung menurut metode Anderson dan Bancroft (1952) dalam Hardiyanto (1996):
S.Eσ 2f: [2/k2 i(MSi)2/ (dfi +2)]0.5
Koefisien variasi genetika (KVG) dihitung menurut rumus Cornelius (1994) dengan kriteria nilai : tinggi (KVG ≥ 14.5%), sedang (5% ≤ KVG < 14.5%) dan
rendah (KVG < 5%). Koefisien korelasi sederhana digunakan untuk melihat keeratan antara karakter pertumbuhan dengan produksi getah
KVG = 100% 2 x x Gx
Korelasi fenotipik dan genetik antar karakter produksi getah
Analisis statistik dilakukan dengan menggunakan program MS Excel, SPSS versi 17 (SPSS inc 2007) untuk mendapatkan nilai rata-rata dan nilai tengah peubah pengamatan masing pohon. Selanjutnya dilakukan uji korelasi fenotipik dan genetik antar masing-masing karakter mengikuti rumus Singh dan Chaudary (1977): rP(xy) =
2 2 ) ( py px xy P Cov dan rG.(xy) =
2 2 ) ( Gy Gx xy G Cov Hasil pengujian nilai KVG, heritabilitas dan korelasi pada tahap evaluasi fenotipik, selanjutnya diverifikasi melalui analisis genetika dengan bantuan penanda molekuler mikrosatelit
3.2.2.1 Variasi genetika dengan penanda mikrosatelit
Kegiatan ekstraksi DNA dilakukan dengan menggunakan metode CTAB (Murray dan Thompson 1980) terhadap 70 sampel daun dan kayu yang telah dikumpulkan. Proses amplifikasi dilakukan dengan mesin PCR Thermal cycler
(PTC-100) terhadap 13.5 ml campuran (2 µl template DNA, 2 µl nuclease free
water, 7.5 green go taqdan 2 µl primer) dan dikondisikan pada suhu 95 0C selama
2 menit (predenaturasi), 95 0C selama 1 menit (denaturasi), 2 menit dengan suhu sesuai petunjuk primer (annealing), dan suhu 72 0C selama 5 menit (extension dan
final extension). Hasil PCR diamati dengan menggunakan gel poliakrilamid 1%
yang telah diwarnai dengan perak nitrat, selanjutnya didokumentasikan dan
discoring berdasarkan pola pita yang terbentuk.
Data yang diperoleh selanjutnya diolah dengan menggunakan software
POPGENE versi 1.31 (Yeh et al.1997), NTSYS 2.02 (Rohlf 1998) dan GenAlex
6.3 (Peakall dan Smouse 2006) untuk menduga beberapa variabel keragaman genetika meliputi persentase lokus polimorfik (PLP), jumlah alel yang teramati (na), jumlah alel efektif (ne), heterozigositas harapan (He). Analisis komponen utama (PCA) dan analysis of molecular variance (AMOVA) juga dilakukan untuk menduga pola penyebarannya. Selain itu digunakan juga software Treeview versi 32 dan Population versi 1.2.30 serta Structure versi 2.23 (Pritchard 2000) untuk melihat pola pengelompokan individu bocor getah.
3.3 Hasil dan Pembahasan
3.3.1 Sejarah pembangunan Kebun Benih Semai (KBS) P. merkusii di Cijambu, Sumedang
Kebun Benih Semai (KBS) Cijambu merupakan kebun benih kombinasi uji keturunan (half-sib) dengan kriteria pohon yang memiliki batang lurus dan pertumbuhannya cepat. Pembangunan KBS Cijambu dilakukan secara bertahap (1978-1983) bersama-sama dengan KBS Baturaden dan Sempolan. Pekerjaan ini diawali dengan pencarian pohon-pohon plus (batang lurus tinggi) dari hutan tanaman di Jawa, hutan alam di Aceh, Tapanuli dan Kerinci. Namun dalam pelaksanaannya benih-benih dari hutan alam tidak dapat diperoleh, sehingga pencarian pohon plus difokuskan pada hutan tanaman di Jawa.
Hasil eksplorasi pohon induk selama 5 tahun di hutan tanaman menemukan 1000 famili untuk selanjutnya digunakan sebagai materi penanaman. Desain yang dipilih untuk penanaman adalah rancangan acak lengkap berblok (RCBD) terdiri dari 5 tree plot, dalam 10 blok dan setiap tahunnnya ditanam 200 famili. Setelah sudah mulai berbuah 100 famili yang jelek dan satu pohon terjelek dari setiap 5
tree plot selanjutnya ditebang sehingga tertinggal 400 pohon/blok. Pada umur 10
tahun dikurangi 50 famili terjelek dan satu pohon terjelek dari setiap plot sehingga tertinggal 150 pohon/blok. Pada umur 15 tahun dilakukan penjarangan lanjutan dan setiap famili hanya diwakili oleh 2 pohon, sehingga dijumpai 100 pohon/blok dan masih dipelihara dengan baik sampai saat ini.
Seleksi dengan tujuan produksi getah dilakukan pada KBS Cijambu selama kurun waktu 2002 sampai 2009 dengan tujuan memperoleh pohon plus dengan karakter produksi getah yang tinggi (kandidat bocor getah). Hasil seleksi tersebut menemukan 110 pohon plus kandidat bocor getah yang tersebar dalam 10 blok dan 6 tahapan penanaman, namun informasi mengenai karakter morfogenetika berupa variasi genetika dan heritabilitas kandidat bocor yang dihasilkan dari seleksi tahap 2 di KBS Cijambu sampai saat ini belum diperoleh. Berdasarkan permasalahan tersebut dilakukan kajian informasi morfogenetika melalui pendekatan evaluasi fenotipik dan analisis genetika dengan penanda molekuler. 3.3.2 Varisi genetika dan heritabilitas pinus kandidat bocor getah
berdasarkan hasil evaluasi fenotipik
Penampilan fenotipik suatu karakter tanaman adalah merupakan hasil total dari faktor genetika, faktor lingkungan, dan interaksi antara faktor genetika dengan faktor lingkungannya (Falconer dan Mackay 1996). Pada penelitian ini besaran pengaruh faktor lingkungan dan genetika tersebut dikuantifikasi dengan menggunakan model perhitungan statistik ragam melalui pendugaan nilai koefisien variasi genetika (KVG) dan heritabilitas dalam arti sempit (narrow
sense heritability). Hasil pendugaan nilai-nilai tersebut disajikan pada sub bab
3.3.1.1 Variasi genetika
Nilai koefisien keragaman genetika mampu menggambarkan pengaruh faktor genetika maupun faktor lingkungan dalam menentukan suatu karakter. Pengetahuan mengenai variasi genetika karakter pertumbuhan diperlukan dalam kegiatan pemuliaan pohon karena menentukan strategi seleksi dan strategi perbanyakan yang akan dikembangkan. Berdasarkan klasifikasi Cornelius (1994) pada Tabel 3.5, variasi genetika di KBS Cijambu memperlihatkan nilai yang beragam dengan total kontribusi karakter antara 2.8% (karakter sudut cabang pertama tajuk) sampai kisaran 28.43% (karakter produksi getah). Hasil tersebut mengindikasikan bahwa tidak semua karakter pertumbuhan di KBS Cijambu dipengaruhi oleh faktor genetika.
Karakter tingkat keparahan serangan hama-penyakit dan produksi getah, memiliki nilai KVG tinggi berdasarkan klasifikasi Cornelius (1994) (KVG ≥
14.5%). Karakter diameter, tebal kulit, jumlah cabang, panjang tajuk, lebar tajuk, sudut percabangan set 2, volume bebas cabang dan volume total memiliki nilai KVG sedang (5% ≤ KVG < 14.5%), sedangkan karakter tinggi total, tinggi bebas cabang dan sudut cabang pada set 2 memiliki nilai KVG rendah (KVG < 5%).
Berdasarkan perhitungan nilai KVG menunjukkan variasi genetika tinggi diperoleh pada karakter tingkat keparahan serangan hama-penyakit (24.0- 28.70%), diikuti produksi getah (14.5-28.43%). Nilai KVG tinggi menandakan bahwa karakter-karakter tersebut sangat dipengaruhi oleh faktor genetika. Nilai KVG yang tinggi untuk karakter produksi getah juga telah diperoleh Roberds et al. (2003) di kebun benih P. taeda (KVG:13.7%). Nilai KVG yang tinggi untuk tingkat keparahan serangan hama penyakit juga sesuai dengan penelitian yang dilakukan oleh Bastein dan Alia (2000) pada P. sylvestris (KVG:26.8%) untuk serangan Hylobius abietis. Dengan demikian pada kegiatan pemuliaan pinus dengan tujuan produksi getah, kedua karakter tersebut (karakter produksi getah dan tingkat serangan hama penyakit) cukup penting untuk dipelajari karena sangat dipengaruhi oleh faktor genetika.
Keterkaitan antara produksi getah dengan tingkat keparahan serangan hama dan penyakit ditemukan pada sebagian besar konifer. Beberapa penelitian sebelumnya pada pinus di daerah temperate (Kleinhentz et al. 1998; Blada 2000;
Kim et al. 2003; Rafael et al. 2005) menyimpulkan serangan hama dan penyakit
menyebabkan terjadinya penurunan produksi getah yang cukup signifikan, oleh karena itu studi mendalam mengenai jenis serangan dan pengaruhnya terhadap produksi getah juga telah dilakukan. Hasil yang sama juga diperoleh Raffa dan Berryman (1982) yang menemukan tingkat serangan hama dan penyakit berhubungan erat dengan kuantitas produksi getah yang dihasilkan oleh pohon. Namun studi secara spesifik mengenai hubungan tingkat serangan hama dan penyakit dengan produksi getah P.merkusii sampai saat ini belum diperoleh. Hasil penelitian pada KBS Cijambu menunjukkan, karakter produksi getah dan tingkat serangan hama dan penyakit memiliki KVG yang tinggi, hal tersebut mengindikasikan kegiatan seleksi pohon dengan fokus produksi getah, dapat dilakukan secara beriringan dengan ketahanan terhadap serangan hama penyakit karena keduanya dipengaruhi oleh faktor genetik dan terkait satu dengan lainnya.
Tabel 3.5 Nilai ragam famili (σ2f), ragam blok (σ2b), ragam galat (σ2e) koefisien
variasi genetika (KVG), heritabilitas famili (h2f) dan individu (h2) beberapa sifat penting pada pinus kandidat bocor getah
Karakter σ2f σ2b σ2e KVG
(%)
h2f SE h2 SE
Umur 29 dan 30 tahun
Produksi getah (g/3 hari) 581.8 338.1 86.1 21.6 0.700.09 0.580.08
Tinggi total (m) 1.94 2.58 11.07 6.0 0.140.05 0.120.05
Tinggi bebas cabang 3.40 7.0 3.39 6.6 0.330.23 0.250.17
Diameter (m) 0.002 0.007 0.002 10.1 0.290.72 0.200.52
Tebal kulit (cm) 0.36 0.89 0.26 8.8 0.340..83 0.240.58
Jumlah cabang 55.92 69.15 33.17 12.8 0.450.10 0.350.08
Panjang tajuk (m) 1.507 3.122 1.74 10.6 0.310.30 0.240.23
Lebar tajuk (m) 1.272 2.96 1.55 12.5 0.300.30 0.220.21
Sudut cabang pertama tajuk 105.97 34.32 58.12 11.2 0.580.10 0.530.09 Volume bebas cabang (m3) 0.002 0.009 0.04 13.9 0.350.22 0.330.20
Volume total (m3) 0.007 0.34 0.14 9.4 0.180.60 0.120.42
Keparahan serangan hama dan penyakit
139.91 15.75 49.86 28.2 0.710.13 0.680.12
Set 1 (umur 29 tahun)
Produksi getah (g/3 hari) 200.70 53.94 46.82 14.5 0.72 0.13 0.670.19
Tinggi total (m) 0.67 3.5 21.83 3.3 0.040.01 0.030.03
Tinggi bebas cabang 1.69 3.5 6.08 4.89 0.210.11 0.150.13
Diameter (m) 0.001 0.0005 0.002 7.9 0.470.15 0.360.04
Tebal kulit (cm) 0.27 0.36 0.12 6.5 0.390.15 0.360.54
Jumlah cabang 15.73 77.16 40.13 13.2 0.140.03 0.120.09
Panjang tajuk (m) 2.42 3.02 1.13 13.0 0.400.48 0.370.18
Lebar tajuk (m) 1.11 1.56 3.26 10.3 0.260.18 0.190.05
Sudut cabang pertama tajuk 6.4 36.15 106.2 2.8 0.070.01 0.040.02 Volume bebas cabang (m3) 0.02 0.002 0.07 13.1 0.310.16 0.190.06
Volume total (m3) 0.06 0.07 0.23 13.8 0.250.15 0.170.37
Keparahan serangan hama dan penyakit
95.59 5.29 37.83 24.0 0.800.16 0.690.04
Set 2 (umur 30 tahun)
Produksi getah (g/3 hari) 1012.5 151.25 151.3 28.43 0.820.084 0.770.08
Tinggi total (m) 1.15 1.02 0.86 4.64 0.460.06 0.380.52
Tinggi bebas cabang 3.08 6.75 0.33 4.91 0.450.09 0.300.06
Diameter (m) 0.002 0.01 0.001 8.49 0.190.07 0.120.411
Tebal kulit (cm) 0.09 0.19 0.38 10.93 0.160.01 0.140.29
Jumlah cabang 66.50 14.09 5.61 8.12 0.840.05 0.700.41
Panjang tajuk (m) 0.82 0.05 1.007 7.81 0.440.06 0.440.55
Lebar tajuk (m) 0.50 0 0.48 7.84 0.510.09 0.510.01
Sudut cabang pertama tajuk 201.95 18.75 75 5.26 0.710.10 0.680.10 Volume bebas cabang (m3) 0.04 0.03 0.009 12.04 0.620.09 0.500.06
Volume total (m3) 0.14 0.33 0.009 13.06 0.380.08 0.260.07
Keparahan serangan hama dan penyakit
126.94 5
Nilai KVG untuk karakter tinggi total pohon, lebar tajuk dan panjang tajuk, jumlah cabang pembentuk tajuk masuk dalam kriteria rendah. Hal tersebut mengindikasikan bahwa faktor lingkungan lebih berpengaruh terhadap fenotipe karakter-karakter tersebut.
3.3.1.1 Heritabilitas
Nilai heritabilitas (h2 dan h2f) menggambarkan proporsi faktor genetika dan faktor lingkungan dalam menentukan suatu karakter. Nilai heritabilitas memiliki kisaran nilai 0-1. Nilai 0 mengindikasikan bahwa karakter tersebut lebih dipengaruhi oleh faktor lingkungan, sedang nilai 1 mengindikasikan bahwa karakter tersebut hanya dipengaruhi oleh faktor genetika. Nilai-nilai tersebut sangat penting bagi pemulia tanaman dalam menentukan strategi seleksi maupun strategi perbanyakan yang akan dilakukan agar perolehan genetikanya tetap stabil.
Hasil pendugaan nilai heritabilitas famili dan individu pada tanaman uji keturunan menunjukkan karakter produksi getah memiliki heritabilitas tinggi (h2f: 0.700.09-0.820.08) dan h2:(0.580.08-0.770.08), keparahan serangan hama penyakit juga menunjukkan nilai tinggi (h2f: 0.640.02-0.800.16 dan h2:0.640.13-0.690.04). Karakter diameter batang, tebal kulit, panjang tajuk, lebar tajuk, jumlah cabang pembentuk tajuk, sudut cabang pertama, volume pohon, dan volume batang) memiliki nilai heritabilitas individu pada kisaran rendah sampai sedang (Tabel 3.5).
Hasil perhitungan untuk nilai heritabilitas karakter produksi getah menunjukkan nilai yang cukup tinggi (0.580.08-0.770.08). Leksono (1996) melakukan penelitian serupa di KBS Cijambu dan Sempolan pada umur tanaman 12 tahun dan menemukan nilai heritabilitas untuk produksi getah sebesar 0.69. Hasil tersebut lebih tinggi dari penelitian Zhang et al. (2010) pada P.eliotii (0.37), Tadesse et al. (2001) pada P. pinaster (0.5) dan Roberds et al (2003) pada P.
taeda (0.44-0.59), demikian juga untuk karakter diameter batang, kekokohan
batang dan kualitas cabang. Nilai heritabilitas pada penelitian ini lebih tinggi dari penelitian awal yang dilakukan pada saat umur pohon 12 tahun, hal tersebut disebabkan karena sampel pohon yang digunakan pada penelitian ini merupakan hasil seleksi yang difokuskan untuk tujuan produksi getah. Untuk nilai heritabilitas yang tinggi pada karakter produksi getah, Wenger (1984); Birk dan Kanowski (1988); Burczyk et al. (1998); Kossuth (1984); Mergen et al. (1955); Gill (1998) menyatakan bahwa sifat tersebut dikendalikan oleh gen. Hal ini menandakan bahwa untuk pemuliaan sifat tersebut kegiatan seleksi akan memberikan perolehan genetika yang tinggi tanpa dipengaruhi oleh interaksi faktor lain.
Besaran nilai karakter tinggi pohon, diameter di KBS Cijambu sedikit berbeda dengan penelitian sebelumnya yang dilakukan oleh Hardiyanto (1996) dengan tujuan untuk memperoleh karakter batang yang bagus dan Leksono (1996) untuk tujuan produksi getah pada umur 12 tahun (h2f: 0.40 dan h2f: 0.43). Demikian juga untuk karakter percabangan (jumlah cabang dan sudut percabangan) di KBS Cijambu pada set 1 tergolong rendah (0.12 dan 0.07), namun pada set 2 masuk dalam kategori tinggi (0.44 dan 0.71). Hasil tersebut berbeda dengan penelitian sebelumnya pada P.sylvestris h2f: 0.24 (Kowalchzyk
2005); h2f: 0.23 dan pada P. taeda (Dean et al. 1986). Karakter percabangan menjadi salah satu variabel dalam evaluasi program pemuliaan pinus dengan fokus utama kayu karena terkait dengan kualitas kayu yang dihasilkan (Jhonson et al. 2009). Hasil pendugaan nilai KVG dan heritabilitas di KBS Cijambu menunjukkan karakter produksi getah memiliki nilai KVG dan heritabilitas yang tinggi, hal tersebut mengindikasikan bahwa faktor genetika lebih dominan mempengaruhi karakter tersebut. Berdasarkan perolehan nilai KVG dan heritabilitas, kegiatan pemuliaan untuk karakter produksi getah dapat diawali dengan melakukan kegiatan seleksi untuk memperoleh individu-individu dengan perolehan getah tinggi. Kegiatan seleksi yang direkomendasikan untuk karakter-karakter dengan nilai variasi genetika tinggi dan heritabilitas tinggi adalah melalui seleksi massa. Hal tersebut sesuai dengan Tadesse et al. (2001) yang mengemukakan jika suatu populasi memiliki nilai heritabilitas tinggi untuk suatu karakter maka seleksi massa/seleksi fenotipik akan lebih efisien dalam memperbaiki karakter tersebut. Seleksi massa merupakan kegiatan seleksi yang didasarkan keragaan fenotipik individu pohon tanpa memperhatikan informasi tentang kinerja induk, keturunan atau kerabatnya. Lebih lanjut White et al. (2007) mengatakan kegiatan seleksi massa merupakan satu-satunya teknik seleksi yang sesuai untuk diaplikasikan pada awal kegiatan pemuliaan pohon dan berguna untuk sifat-sifat dengan heritabilitas tinggi karena fenotipe yang ada merupakan cerminan yang baik dari genotipenya.
Berdasarkan struktur produksi getah dan pertumbuhan kandidat bocor getah (telah dibahas pada Bab 2), kondisi lingkungan di 3 KBS Perum Perhutani memiliki karakteristik kondisi lingkungan yang berbeda. Hasil studi pada Bab 2, menunjukkan adanya kesesuaian antara famili tertentu dengan lingkungan tertentu yang ditunjukkan dengan ketidakkonsistenan perolehan getah dari famili yang sama ketika ditanam di KBS yang berbeda. Ketidakkonsistenan perolehan getah dari famili yang sama di KBS berbeda dapat disebabkan karena sumber benih untuk materi penanaman berasal dari half sib, namun demikian kondisi lingkungan juga mempengaruhi cerminan genetik individu tersebut. Melalui pertimbangan tersebut seleksi secara famili akan sulit untuk dilakukan karena adanya kesesuaian famili tertentu dengan lokasi tertentu. Seleksi massa/seleksi fenotipe dapat mengatasi kelemahan interaksi GxE karena didasarkan pada keragaan fenotipe secara individual, sehingga individu kandidat bocor getah yang dihasilkan merupakan cerminan dari famili-famili yang adaptif di setiap KBS.
Selain informasi mengenai strategi seleksi, perolehan nilai KVG dan heritabilitas juga terkait dengan strategi perbanyakan yang akan dilakukan. Jika suatu karakter memiliki nilai heritabilitas tinggi, maka strategi perbanyakan melalui pembiakan generatif mampu memberikan kontribusi terhadap ketersediaan bibit dengan karakter target, sebaliknya jika karakter tersebut memiliki nilai yang rendah maka perbanyakan secara vegetatif sangat penting untuk menjaga stabilitas bibit yang akan diperoleh. Hasil penelitian ini menemukan nilai heritabilitas yang tinggi untuk karakter produksi getah, sehingga untuk pengembangannya dapat dilakukan melalui pembiakan generatif dari benih bocor getah famili yang adaptif di setiap KBS. Namun demikian cara tersebut terkendala karena penanaman uji keturunan untuk bocor getah baru dibangun pada tahun 2007 sehingga baru mampu menghasilkan materi generatif pada 2017.
Berdasarkan kecukupan pemenuhan kebutuhan bibit bocor getah dalam jangka pendek, maka perbanyakan secara vegetatif mampu mengatasi permasalahan tersebut. Bahasan mengenai strategi perbanyakan vegetatif kandidat bocor getah disajikan pada Bab 5.
Hasil penelitian ini juga memberikan informasi bahwa kegiatan seleksi yang dilakukan oleh Perum Perhutani dengan tujuan awal produksi kayu, juga mampu menghasilkan produksi getah tinggi. Hal tersebut ditunjukkan dengan kekonsistenan nilai heritabilitas hasil penelitian leksono (1996) sebesar 0.69 pada umur tanaman 12 tahun, dengan hasil penelitian yang dilakukan pada umur tanaman 30 tahun (h2: 0.580.08-0.770.08). Meskipun memiliki nilai yang relatif stabil, kegiatan pemeliharan (pemangkasan cabang, penjarangan, pembersihan tumbuhan bawah dan pemupukan ) masih tetap harus dilaksanakan untuk menjaga ekspresi genetika dari karakter target. Hal tersebut terkait dengan pernyataan Wright (1976) bahwa kondisi lingkungan yang kurang mendukung mampu merubah nilai heritabilitas suatu karakter ke arah yang kurang baik.
Perubahan nilai heritabilitas di KBS Cijambu dijumpai pada beberapa karakter yang diamati pada penelitian ini. Hal tersebut dapat disebabkan karena kondisi lingkungan yang kurang mendukung sehingga menyebabkan perubahan heritabilitas karakter suatu pohon sebanyak atau lebih besar daripada perubahan di susunan genetikanya. Lebih lanjut Namkoong et al. (1980) dan Franklin et al.(1979) menyebutkan perubahan nilai heritabilitas juga dapat disebabkan oleh perubahan umur dan perbedaaan fase pertumbuhan. Perubahan nilai heritabilitas karakter pertumbuhan seperti tinggi, diameter dan ketebalan kulit selain disebabkan adanya perubahan umur dan fase pertumbuhan kemungkinan juga dipengaruhi oleh adanya praktek silvikultur seperti penjarangan dan praktek pengelolaan lainnya. Keterkaitan antara praktek silvikultur dengan perubahan nilai heritabilitas karakter pertumbuhan juga ditemukan pada P. taeda (Gwaze et al.
1997; Lopez-Upton et al. 1999).
Pada penelitian ini juga dijumpai sedikit perbedaan nilai KVG dan heritabilitas beberapa karakter dengan 2 pendekatan yang berbeda. Pendekatan set tahun tanam pada penelitian ini lebih sesuai digunakan karena KBS Cijambu merupakan kebun benih yang ditanam secara bertahap dengan asal famili yang berbeda-beda dan kondisi tiap blok tahun tanaman yang relatif beragam. Meskipun memiliki sedikit perbedaan nilai KVG dan heritabilitas yang dihasilkan dari dua pendekatan tersebut, secara umum memiliki trend yang sama dan dapat disimpulkan bahwa nilai KVG dan heritabilitas untuk produksi getah di KBS Cijambu cukup tinggi, yang mengindikasikan faktor genetik lebih dominan menentukan karakter produksi getah. Selanjutnya, sebagai verifikasi hasil evaluasi fenotipik tersebut, dilakukan pengujian dengan menggunakan bantuan penanda molekuler mikrosatelit.
3.3.3 Variasi genetika kandidat bocor getah dengan penanda mikrosatelit Hasil evaluasi fenotipik terhadap kandidat bocor getah, memperlihatkan nilai KVG dan heritabilitas yang tinggi untuk karakter produksi getah yang mengindikasikan sifat produksi getah lebih dipengaruhi oleh faktor genetika. Sebagai langkah verifikasi terhadap hasil evaluasi fenotipik, selanjutnya dilakukan analisis secara genetika menggunakan penanda molekuler mikrosatelit.
3.3.2.1. Deskripsi DNA electropherogram
Penggunaan primer mikrosatelit pada konifer pertama kali dilakukan pada
P.radiata (Smith dan Devey 1994). Dibandingkan dengan gymnospermae,
transfer mikrosatelit pada jenis pinus relatif lebih sulit (Diapari 2006; Scott et al.
2001 dan Auckland 2002). Sebagai contoh pada P.taeda hanya sekitar 60% saja loci mikrosatelit yang teramplifikasi dengan pola polimorfik (Devey et al. 1999; Kutil dan William 2001).
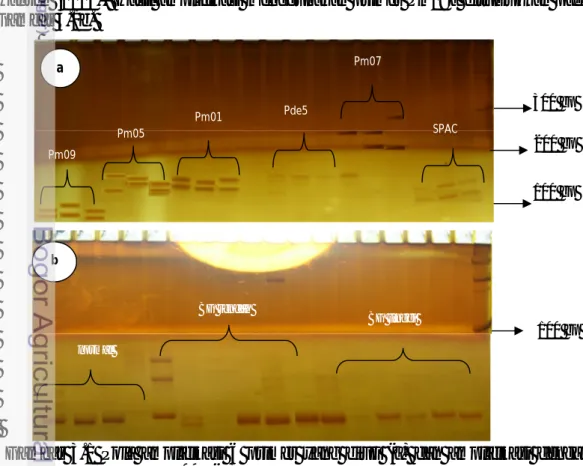
Hasil amplifikasi dengan menggunakan penanda mikrosatelit memperlihatkan semua primer yang digunakan memiliki kemampuan untuk mengamplifikasi pita DNA dengan baik. Terdapat 7 primer (pm01, pm05, pm07, pm09a, pm12, Pde5 dan SPAC 11.6) dengan kemampuan amplifikasi pita dengan pola polimorfik, dan 2 primer (pm04 dan pm08) dengan kemampuan amplifikasi namun dengan pola monomorfik (Gambar 3.1a). Kemampuan amplifikasi primer mikrosatelit untuk amplifikasi DNA telah teruji pada penelitian sebelumnya oleh
Li et al. (2002); Lagercrantz et al. (1993); Wang et al. (1994) dan Vendramin dan
Hansen (2005). Hasil amplifikasi menggunakan primer Pm09a ditunjukkan pada Gambar 3.1b.
Gambar 3.1 Pola amplifikasi 6 primer yang diuji (a) dan amplifikasi dengan primer pm09a (b) b normal BG rendah BG tinggi 100 bp Pm09 Pm05 Pm01 Pde5 Pm07 SPAC 100 bp 200 bp 300 bp a
Berdasarkan Gambar 3.1a terlihat bahwa primer mikrosatelit yang digunakan pada penelitian memiliki alel dengan perkiraan panjang fragmen antara 100-300 bp. Pm01 memiliki ukuran fragmen (111-117 bp), pm05 (112-118 bp), pm07 (284-309 bp), pm09 (81-99 bp, pde5 (116-235 bp) dan Spac 11.6 (165-200 bp). Hasil amplifikasi dengan menggunakan primer-primer mikrosatelit tersebut menunjukkan pola yang berbeda antara individu bocor getah dan normal. Amplifikasi dengan menggunakan primer pm09 memperlihatkan kandidat bocor getah tinggi dan rendah memiliki pola amplifikasi alel yang berbeda dengan pinus normal. Alel dengan perkiraan panjang alel 85 bp, 88 bp, 95 bp dan 99 bp dapat dijumpai pada kandidat bocor getah tinggi dan rendah, sedangkan pada pinus normal tidak dijumpai adanya alel 88 (Gambar 3.1b). Hasil amplifikasi dengan keenam primer lain (pm01, pm05, pm07, pm12, pde5 dan Spac 11.6) selanjutnya dianalisis untuk melihat struktur alelik dari masing-masing populasi produksi.
3.3.2.2 Struktur alelik kandidat bocor getah
Hasil DNA elektropherogram (Gambar 3.1) selanjutnya dianalisis untuk memastikan panjang fragmen (bp) pita hasil amplifikasi. Hasil amplifikasi tersebut selanjutnya discoring dan dianalisis dengan bantuan software Genalex
untuk mengetahui struktur alel pada ketiga populasi produksi. Hasil perhitungan struktur alel yang diperoleh menunjukkan jumlah alel keseluruhan populasi berkisar antara 3-5 alel (Tabel 3.6)
Tabel 3.6 Jumlah lokus dan perkiraan panjang fragment
No Lokus Alel Dugaan panjang fragment (bp)
1 Pm01 1 111 2 115 3 117 2 Pm05 1 108 2 116 3 118 3 Pm07 1 232 2 260 3 280 4 Pm09a 1 85 2 88 3 95 4 99 5 Pm12 1 128 2 132 3 135 6 Pde5 1 138 2 150 3 197 4 236 7 Spac 11.6 1 127 2 131 3 135 4 140
Jumlah alel sebanyak 4 buah dapat ditemukan pada lokus pm09, Pde5 dan SPAC 11.6, sedangkan pada lokus pm01, pm05, pm07 dan pm12 hanya dijumpai
3 alel. Jumlah alel tersebut sedikit berbeda dengan penelitian Nurtjahjaningsih et al. (2005) karena pada lokus pm07 hanya ditemukan 3 alel (referensi sebelumnya:5 alel) dan pada lokus pm09a hanya ditemukan 4 alel (referensi sebelumnya:6 alel).
Nilai frekuensi alel tertinggi dijumpai pada lokus pm05 alel 108 kandidat bocor getah tinggi dan lokus pm07 alel 232 populasi pinus normal (0.700). Frekuensi alel terendah dijumpai pada lokus pm01 alel 117 kandidat bocor getah tinggi dan bocor getah rendah (0.000) serta lokus pm05 alel 118dan lokus pm09 alel 88 pada pinus normal. Pada semua lokus mikrosatelit frekuensi alel dari masing-masing kelas produksi getah cukup beragam, sehingga memberikan peluang cukup besar untuk melakukan kegiatan seleksi untuk memperoleh karakter bocor getah di dalam populasi (Tabel 3.7).
Tabel 3.7 Frekuensi alel kandidat bocor getah dan normal berdasarkan 7 primer mikrosatelit dalam populasi bocor getah tinggi, bocor getah rendah dan normal Bocor getah tinggi Bocor getah rendah Normal Jumlah sampel (n) 30 (30) (10)
Lokus Panjang fragmen (bp) Jumlah Alel
Alel Frekuensi alel
pm01 111-117 3 1 0.567 0.500 0.200 2 0.433 0.500 0.400 3 0.000 0.000 0.400 pm05 112-118 3 1 0.700 0.433 0.400 2 0.233 0.567 0.600 3 0.067 0.000 0.000 pm07 284-309 3 1 0.567 0.600 0.700 2 0.233 0.067 0.100 3 0.200 0.333 0.200 pm09a 81-99 4 1 0.167 0.133 0.500 2 0.200 0.133 0.000 3 0.567 0.667 0.200 4 0.067 0.067 0.300 pm12 181-193 3 1 0.333 0.500 0.500 2 0.600 0.333 0.000 3 0.067 0.167 0.500 Pde5 116-235 4 1 0.067 0.167 0.600 2 0.667 0.467 0.100 3 0.033 0.300 0.200 4 0.233 0.067 0.100 Spac 11.6 165-200 4 1 0.067 0.067 0.400 2 0.300 0.133 0.300 3 0.300 0.133 0.100 4 0.333 0.667 0.200
Perbedaan jumlah lokus dan panjang fragmen dapat disebabkan karena terjadinya proses seleksi maupun mutasi serta perbedaan lokasi pengambilan sampel. Pada lokus pm01 dijumpai 1 alel (117) pada pinus normal, namun tidak dijumpai pada kandidat bocor getah. Pada lokus pm05, alel 118 hanya dijumpai
pada kandidat bocor getah tinggi, namun pada kandidat bocor getah rendah dan normal alel tersebut tidak dijumpai. Pada lokus pm09, alel 88dijumpai pada bocor getah namun tidak ada pada pinus normal demikian juga alel 132 pada pm12 (Tabel 3.8). Dijumpai juga adanya 1 private alel pada bocor getah tinggi dan1
private alel pada produksi normal, kemungkinan hal tersebut menjadi salah satu pembeda antara kandidat bocor getah dan normal.
Tabel 3.8 Hasil amplifikasi kandidat bocor getah dan normal berdasarkan 7 primer mikrosatelit
Alel
Dugaan panjang fragmen (bp)
Lokus Repeat 8 5 8 8 9 5 9 9 1 0 8 1 1 1 1 1 5 1 1 6 1 1 7 1 1 8 1 2 7 1 2 8 1 3 1 1 3 2 1 3 5 1 3 8 1 4 0 1 5 0 1 9 7 2 3 2 2 3 6 2 6 0 2 8 0 pm01 (TG)12 - - - + + - N - - - - pm05 (TG)9 - - - - + - - + - T - - - - pm07 (AC)8 (AT)4 - - - + - + + pm09a (AT)5 (GT)18 (AT)2 + T R + + _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ pm12 (GT)5CT (GT)5(AT)5 _ _ _ _ _ _ _ _ _ _ _ + + T R + _ _ _ _ _ _ _ _ Pde5 (CA)5C (CA)51 _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ + _ + + _ _ _ _ SPAC 11.6 (CA)29(TA) 7 _ _ _ _ _ _ _ _ _ _ + _ + _ + _ + _ _ _ _ _ _
Keterangan: +ditemukan pada semua populasi; -:tidak menunjukkan amplifikasi; N:hanya ditemukan pada populasi pinus normal; T:hanya ditemukan pada bocor getah tinggi; TR:ditemukan pada bocor getah tinggi dan rendah.
3.3.2.3 Variasi genetika dalam populasi
Keragaman genetika dalam populasi dihitung berdasarkan frekuensi alel dan dapat ditunjukkan oleh beberapa nilai variabel keragaman genetika (Tabel 3.9), jumlah alel yang diamati (na), jumlah alel yang efektif (ne), persen lokus polimorfik (PLP) dan heterozigositas harapan (He) dan heterozigositas aktual
(Ho). Berdasarkan perhitungan tersebut diperoleh nilai jumlah alel yang teramati pada setiap lokus antara 3.00-3.286 dengan rata-rata 3.143 dan jumlah alel efektif antara 2.244-2.403 dengan rata-rata 2.320. Nilai heterozigositas harapan berkisar antara 0.545-0.566 dengan rata-rata 0.554. Nilai rata-rata untuk heterozigositas aktual adalah 0.695 dengan interval nilai 0.648-0.771.
Berdasarkan nilai keragaman genetika (He), kandidat bocor getah tinggi memiliki nilai yang lebih tinggi (0.551) dibanding bocor getah rendah (0.545), namun lebih rendah dari pinus normal (0.566). Hasil tersebut sedikit berbeda dengan penelitian awal yang dilakukan Nurtjahjaningsih et al. (2007) mengenai keragaman populasi induk dan anakan di KBS Sumedang (0.501), KBS Baturaden (0.468) dan hampir sama dengan KBS Sempolan (0.545) dengan penanda mikrosatelit. Penelitian dengan menggunakan penanda molekuler lain telah dilakukan oleh (Kartikawati 1998; Na’iem 2000) dengan menggunakan penanda allozyme memperoleh nilai He:0.304, Siregar dan Hattemer (2005) pada hutan tanaman pinus di Jawa dengan penanda isozym memperoleh nilai He:0.389, Munawar ( 2002) dengan penanda allozyme pada strain Kerinci memperoleh nilai He: 0.042 dan pada strain Tapanuli (He: 0.219). Hasil penelitian dengan penanda mikrosatelit pada penelitian ini tidak jauh berbeda dengan jenis pinus lain di
daerah sub tropis menurut Echt et al. (1999) pada P. strobus (He: 0.515), Boys et al. (2005) pada P.resinosa (He:0.508) dan Bucci et al. (1998) pada P. halepensis
(He: 0.222), namun lebih rendah dari P. radiata (He: 0.625) menurut (Smith dan Devey 1994). Nilai tersebut masih dalam kisaran keragaman dengan penanda mikrosatelit umumnya pada pinus (He: 0.50-0.80) menurut Smith dan Devey 1994; Thomas et al. 1999; Keys et al. 2000). Di keseluruhan populasi produksi yang diamati (kandidat bocor getah dan pinus normal) nilai heterozigositas harapan (He) lebih rendah dibandingkan dengan heterozigositas aktual (Ho), hal tersebut mengindikasikan proporsi heterozigositas yang cukup tinggi di dalam populasi.
Tabel 3.9 Keragaman genetika dalam populasi Pinus merkusii kandidat bocor getah dan normal
Populasi Jumlah
sampel
PLP na ne Ho He
Bocor getah tinggi 30 100% 3.286 2.315 0.667 0.551 Bocor getah rendah 30 100% 3.143 2.244 0.648 0.545
Normal 10 100% 3.000 2.403 0.771 0.566
Rata-rata 100% 3.143 2.320 0.695 0.554
Keterangan:
PLP= persentase lokus polimorfik ; na = jumlah alel teramati ; ne = jumlah alel efektif;Ho: heterozigositas aktual He = heterozigositas harapan menurut Nei’s (1973) gene diversity
Proporsi heterozigositas dalam populasi kandidat bocor getah menunjukkan nilai tergolong sedang, hal tersebut terjadi karena individu-individu dalam KBS merupakan tanaman hasil uji keturunan yang berasal dari famili-famili pohon induk berbeda dan menyerbuk secara bebas di populasi alaminya (segregating
population). Menurut Novalina & Sagala (2011) pada tanaman tahunan yang
menyerbuk bebas, kemungkinan gen-gen yang terdapat pada suatu individu tanaman memiliki genotipe berbeda-beda sehingga menyebabkan keragaman genetika tanaman hasil uji keturunan tinggi. Hasil tersebut sesuai dengan hasil penelitian Feng et al. (2010) pada kebun benih P.koraiensis, dan El Kasaby (1991) pada kebun benih semai konifer umumnya. Kondisi tersebut juga dijumpai pada KBS P. merkusii di Jember hasil penelitian Nurtjahjaningsih et al. (2007).
Variasi genetika pada kandidat bocor getah di KBS Cijambu dan pinus normal tergolong sedang, mengindikasikan bahwa populasi tersebut memiliki kemampuan beradaptasi yang cukup tinggi terhadap perubahan lingkungan (White
et al. 2007). Hal yang perlu diperhatikan pada karakterisasi kandidat bocor getah
dengan bantuan penanda mikrosatelit ini adalah adanya kenyataan bahwa deteksi mikrosatelit pada karakter kuantitatif sangat ditentukan adanya proses seleksi dan mutasi yang terjadi maupun tekanan seleksi (Karhu 2001; Barton dan Turelli 1989). Adanya proses seleksi fenotipe yang dilakukan untuk pemilihan kandidat bocor getah, kemungkinan menjadi salah satu penyebab perbedaan variasi genetika kandidat bocor getah dibandingkan dengan pinus normal.
3.3.2.4 Variasi genetika antar populasi
Keragaman genetika antar populasi dapat diidentifikasi dengan menggunakan variabel jarak genetika. Jarak genetika mengukur perbedaan struktur genetika antar dua populasi pada suatu lokus gen tertentu. Pebedaan genetika dari dua atau lebih populasi pada umumnya dianalisis dengan sebuah matrik dengan elemen-elemennya berupa jarak genetika dan pasangan kombinasi dari masing-masing populasi (Finkeldey 2005). Data mengenai jarak genetika dalam penelitian ini disajikan pada Tabel 3.10
Tabel 3.10 Jarak genetika antar populasi produksi getah
Populasi Kode Jumlah
sampel Bocor getah tinggi Bocor getah rendah Normal
Bocor getah tinggi T 30 **** Bocor getah rendah R 30 0.114 ****
Normal N 10 0.483 0.322 ****
Analisis jarak genetika dan dendrogram menurut Nei (1972) menunjukkan pola pengelompokan yang berbeda antar kelas produksi getah. Kandidat bocor getah membentuk kelompok tersendiri yang terpisah dari pinus normal. Jarak genetika antara bocor getah tinggi dan rendah relatif kecil (0.114) sehingga dapat disimpulkan kedua populasi tersebut berkerabat dekat sedangkan jarak genetika antara kandidat bocor getah tinggi dan normal relatif besar (0.483). Nilai jarak genetika selanjutnya diolah berdasarkan metode pemasangan kelompok aritmatika tidak dibobot (Unweighted Pair-Grouping Method with Aritmatic Averaging, UPGMA) dengan menggunakan software NTSYS versi 2.02 menghasilkan pola pengelompokan seperti terlihat pada Gambar 3.2a. Hasil analisis jarak genetika, menunjukkan pengelompokan populasi yang sangat jelas menurut kelas produksinya. Hasil uji jarak UPGMA memperlihatkan populasi normal terpisah dengan populasi bocor getah.
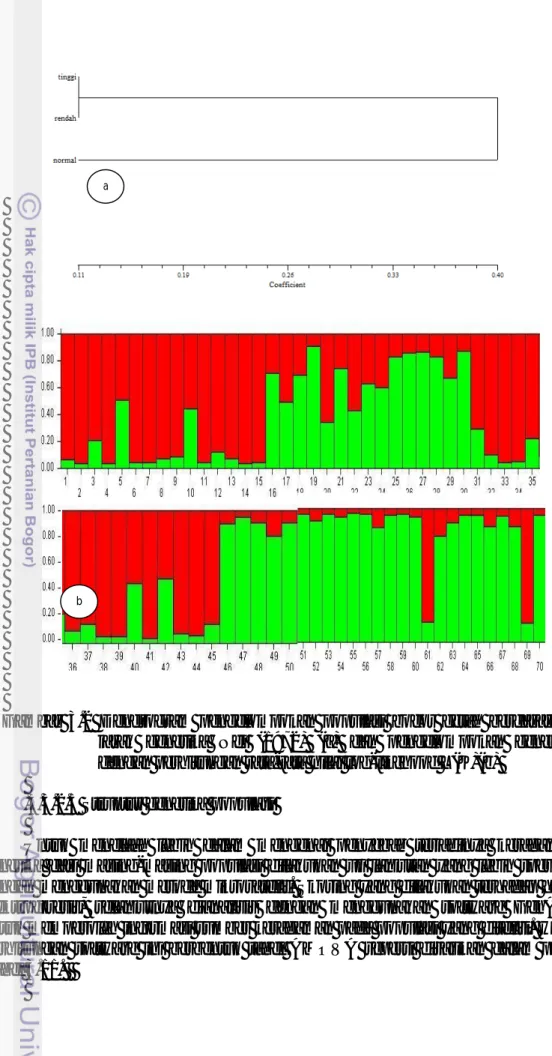
Hasil pengolahan data keragaman genetika pada populasi bocor getah dapat dilihat juga dengan melakukan pengujian model alogaritma melalui analisis struktur dengan menggunakan pendekat frekuensi alel (Pritchard 2000). Hasil pengujian dengan bantuan software structure menunjukkan pengelompokan populasi bocor getah paling optimal pada K:2. Hal tersebut terjadi karena hasil analisis logaritma memperlihatkan peluang L(K) meningkat secara liner pada K:2 Visualisasi dengan bantuan software R memperlihatkan pola warna yang berbeda, menandakan perbedaan populasi (Gambar 3.2b). Perhitungan struktur populasi menunjukkan populasi pinus hasil pengujian terpisah menjadi dua sub populasi. Hasil tersebut sesuai dengan pengelompokan berdasarkan jarak genetika yang telah dilakukan sebelumnya
Gambar 3.2 Dendrogram pengelompokan populasi bocor getah berdasarkan jarak genetika Nei (1972) (a) dan pengelompokan genetika dengan perhitungan rata-rata nilai log-likehood L(K)(b)
.3.3.2.5 Struktur genetika populasi
Untuk menelaah lebih dalam mengenai penyebab terjadinya keragaman genetika dari masing-masing populasi dilakukan uji lanjutan yang lebih spesifik dengan menggunakan metode mikrosatelit. Skoring yang dilakukan terhadap hasil elektroforesis, selanjutnya dianalisis dengan menggunakan software GenAlex
untuk memperoleh informasi sumber keragaman pada populasi yang diteliti. Hasil perhitungan software ini berbentuk tabel AMOVA seperti disajikan dalam pada Tabel 3.11.
a
Hasil perhitungan AMOVA (Tabel 3.11) memperlihatkan bahwa 79% keragaman terdapat di dalam populasi, sisanya (21%) disebabkan keragaman antar populasi. Hasil tersebut tidak jauh berbeda dibandingkan penelitian yang dilakukan oleh Moran et al. (1988) pada KBS Acacia auliculiformis; Millar et al.
(1991) pada KBS P.sylvestris dan Wu (1999) pada KBS P. tabulaeformis. Tabel 3.11 Hasil perhitungan analysis of molecular variance (AMOVA)
Sumber keragaman db Jumlah Kuadrat Kuadrat Tengah Persentase Ragam Indeks Fiksasi Antar populasi 2 25.590 12.795 21% Fst: 0.119
Dalam populasi 67 105.067 3.283 79% Fis:-0.242
Total 69 130.657 100%
Nilai diferensiasi genetika (Fst) memperlihatkan nilai yang lebih tinggi (0.1191) dibandingkan penelitian Nurtjahjanigsih (2007) sebesar 0.03 namun masih dalam kategori sedang, hal tersebut mengindikasikan bahwa adanya keseragaman genetika antar populasi, hal tersebut juga didukung oleh pernyataan Siregar dan Hattemer (2005); Hardiyanto dan Danarto (2000) yang menyatakan bahwa pohon-pohon di dalam KBS berasal dari material dengan keragaman genetika yang terbatas. Nilai indeks fiksasi (Fis) menggambarkan besarnya
inbreeding yang telah terjadi pada lokasi tersebut. Angka minus menunjukkan
bahwa persentase heterozigot lebih besar daripada homozigot, sedangkan nilai Fis (+) menunjukkan semakin besarnya homozigositasnya yang berarti kemungkinan terjadi inbreeding semakin besar (Munawar 2002).
Pada lokasi KBS Cijambu diperoleh angka Fis sebesar: -0.242 yang mengindikasikan bahwa peluang inbreeding cukup rendah sehingga sangat menjanjikan untuk kegiatan pemuliaan selanjutnya. Berdasarkan nilai nilai-tersebut strategi seleksi bocor getah selanjutnya sebaiknya lebih difokuskan dalam populasi.
3.3.2.6 Hubungan genetika antar individu populasi produksi
Pengelompokan dengan menggunakan matriks kovarian jarak genetika secara individual dengan menggunakan software Population versi 1.2.30 dan
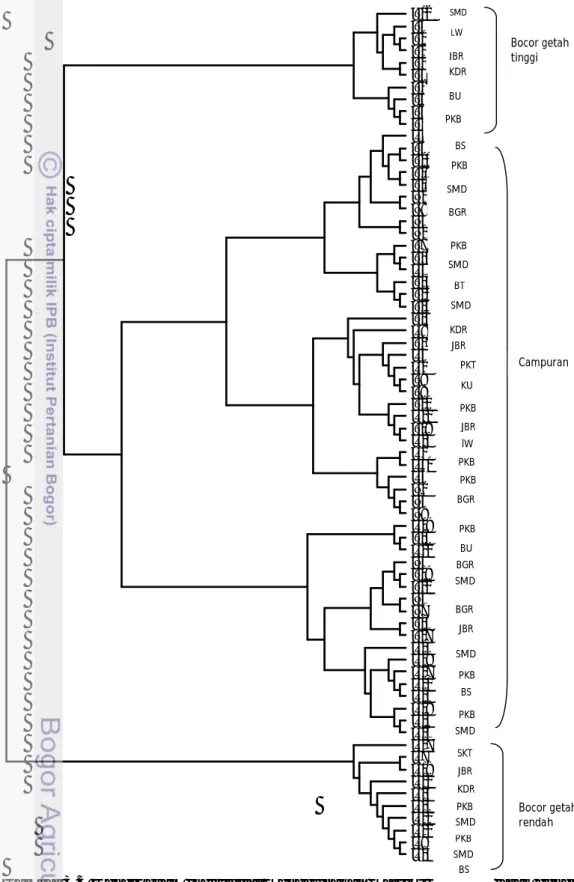
treeview 32 diperoleh dendrogram yang memperlihatkan adanya pengelompokan
individu-individu penyusun populasi bocor getah maupun normal menjadi 2 kelompok besar (Gambar 3.3). Kelompok pertama terdiri campuran individu bocor getah tinggi, rendah dan normal dengan interval produksi antara 87.2 - 271.2 g/pohon/3 hari. Kelompok kedua terdiri bocor getah rendah dengan interval produksi antara 60.7 – 134.6 g/pohon/3 hari.
Pengelompokan tersebut tidak mutlak karena perbedaan produksi getah namun cenderung dipengaruhi oleh asal sumber benih, sehingga individu-individu dari asal sumber benih yang sama akan mengelompok menjadi satu. Beragamnya pengelompokan individu populasi terjadi karena materi penanaman berasal dari famili berbeda yang ditanam secara bertahap selama kurun waktu 1978-1983. Hasil pengelompokan ini tidak berbeda dengan pengelompokan berdasarkan metode pemasangan kelompok aritmatika tidak dibobot sebelumnya.
"T12 "T4 "T1 "T2 "T10 "T30 "T24 "T3 "T6 "T15 "R5 "T5 "T21 "T28 "T29 "N1 "N9 "N4 "N10 "T7 "T13 "R4 "T14 "T11 "T23 "T26 "R8 "T17 "R3 "R2 "T8 "T9 "T20 "R22 "T18 "R16 "R1 "R30 "R6 "N2 "N6 "N8 "R19 "T16 "R20 "N3 "T19 "T22 "N5 "N7 "T25 "T27 "R23 "R29 "R17 "R21 "R24 "R28 "R25 "R26 "R27 "R7 "R18 "R12 "R15 "R13 "R11 "R10 "R9 "R14
Gambar 3.3 Dendrogram individual menggunakan matriks kovarian jarak genetika dengan 100 individual boothstap. BT:Banyumas Timur, Ku:Kuningan SKT:Surakarta,SMD:Sumedang,LW:Lawu,JBR:Jember,KDR:Kediri; BU:Bandung Utara; PKB:Pekalongan Barat;BS:Bandung Selatan.
BU PKB SMD LW JBR KDR Bocor getah tinggi BS PKB SMD BGR PKB BT SMD SMD KU PKB KDR PKT JBR BGR BGR JBR PKB SMD PKB JBR lW BU PKB PKB SMD BGR BS PKB SMD PKB SMD JBR SKT BS PKB KDR SMD Campuran Bocor getah rendah
3.3.4 Korelasi fenotipik dan genotipik antara karakter produksi getah dan komponen hasil lainnya
Seperti telah diketahui tingkat produksi resin pada pinus merupakan resultante dari peranan beberapa karakter pertumbuhan (Bab 2). Beberapa karakter fenotipik dilaporkan memiliki kaitan dengan produksi pada beberapa jenis pohon bergetah. Informasi mengenai keterkaitan karakter fenotipik dengan produksi getah di Indonesia banyak dilaporkan pada tanaman karet dan menunjukkan beberapa karakter seperti lilit batang, indeks penyumbatan, jumlah pembuluh lateks, diameter pembuluh lateks, kadar sukrosa, kadar fosfat, kadar thiol dan tebal kulit memiliki pengaruh langsung terhadap produksi lateks (Goncalves et al. 2005).
Koefisien korelasi genetipik dan fenotipik (Tabel 3.12) mempunyai nilai yang berbeda,mengindikasikan bahwa korelasi fenotipik yang ada tidak sepenuhnya merupakan ekspresi dari genetik. Adanya interaksi faktor lingkungan dan genetik berpengaruh terhadap penampilan fenotipik pohon dan tercermin dari nilai korelasi fenotipik yang dihasilkan. Hasil korelasi fenotipik menunjukkan karakter diameter batang, panjang tajuk dan tebal kulit berkorelasi positif nyata dengan produksi getah, sedang karakter tingkat keparahan serangan terhadap hama dan penyakit dan jumlah cabang pembentuk tajuk berkorelasi negatif nyata dengan produksi getah.
Korelasi yang positif antara produksi getah dengan beberapa karakter mengindikasikan bahwa produksi getah akan semakin tinggi dengan semakin tingginya nilai karakter tersebut. Sebaliknya, korelasi negatif antara produksi getah dengan beberapa karakter mengindikasikan bahwa produksi getah akan semakin menurun dengan tingginya nilai komponen tersebut. Korelasi positif produksi getah dengan karakter diameter dan tajuk sesuai dengan penelitian yang dilakukan oleh Westbork (2011); Tadesse et al. (2001); Zheng dan Xu (1992) dan Wang dan Zhu (1994) yang diduga terkait dengan jumlah saluran resin pada kayu dan luas bidang penyerapan fotosintesis. Korelasi negatif jumlah cabang dengan produksi getah sesuai Papajiannopoulos (2002) hal tersebut terkait dengan keterbukaan kanopi yang mendorong proses fotosintesis.
Korelasi genetik antara produksi getah dengan diameter, tebal kulit, dan panjang tajuk pada penelitian ini mengindikasikan bahwa produksi getah akan semakin meningkat dengan peningkatan diameter, tebal kulit dan panjang tajuk. Sebaliknya produksi getah akan menurun seiring peningkatan jumlah cabang dan serangan hama dan penyakit. Hasil berbeda ditemui pada famili set 2 (umur tanaman 30 tahun). Pada set uji keturunan ini karakter tinggi pohon juga memiliki korelasi genetik negatif dengan produksi getah, artinya produksi getah akan menurun dengan adanya peningkatan tinggi pohon.
Hasil verifikasi secara morfologi dan molekuler pada penelitian kandidat bocor getah memperlihatkan adanya hubungan antara karakter fenotipik dengan produksi getah, namun dengan keeratan yang sedang (r < 0.8) sesuai hasil pada Bab 2, hal tersebut sesuai dengan peneliti sebelumnya yang menyatakan intensitas produksi resin dipengaruhi oleh faktor genetika, dan menduga adanya gen yang berpengaruh terhadap karakter kuantitatif tersebut (Burczyk et al. 1998; Kossuth 1984 dan Mergen et al. 1955). Berdasarkan nilai korelasi genetika karakter
diameter, tebal kulit, panjang tajuk serta tingkat serangan memiliki korelasi yang nyata dengan produksi getah, sehingga untuk keperluan seleksi bocor getah selanjutnya karakter tersebut dapat dijadikan sebagai indikator seleksi.
3.3.5 Implikasi keragaman genetika terhadap kegiatan manajemen tegakan yang dilakukan untuk mendukung produksi getah.
Status keragaman genetika merupakan faktor penting dalam merumuskan strategi konservasi yang efektif dan manajemen plasma nutfah. Secara umum, pohon-pohon jenis konifer memiliki keragaman genetika yang tinggi dibandingkan dengan pohon jenis yang lain. Pengetahuan mengenai keragaman genetika dari suatu populasi sangat penting untuk kegiatan konservasi yang dilakukan karena keragaman genetika yang tinggi akan membantu populasi beradaptasi terhadap perubahan lingkungan di sekelilingnya dan terjaganya biodiversitas. Lebih lanjut Namkoong et al. (1996) dalam Finkeldey (2005) menyatakan derajat keragaman genetika merupakan salah satu indikator kelestarian pengelolaan hutan.
Untuk tujuan produksi getah, pengetahuan mengenai keragaman fenotipik dan genetika kandidat bocor getah sangat diperlukan dalam rangka identifikasi genotipe-genotipe superior dengan tujuan konservasi maupun pemuliaan di masa mendatang. Pada KBS Cijambu, individu-individu yang berada pada posisi menjulur ke kanan (produksi getah ≥ 150 g/pohon/3 hari) meliputi famili BG107, BG32, BG101, BG33, BG47 dan BG75 sangat perlu untuk dikonservasi dan dikembangkan karena memiliki keragaman yang sempit dan rawan terhadap kepunahan, sedangkan untuk perbanyakan massal individu-individu dengan frekuensi terbanyak (34%) dengan interval produksi 82-103 g/pohon/3 hari dapat digunakan untuk sumber materi perbanyakan (telah dibahas lebih mendalam pada Bab 2).
Hasil verifikasi genetika kandidat bocor getah menunjukkan bahwa produksi getah sangat dipengaruhi oleh faktor genetika, hal tersebut ditunjukkan oleh tingginya nilai heritabilitas, tingginya variasi genetika dan pengelompokan populasi produksi yang berbeda antara bocor getah tinggi, rendah dan normal. Namun demikian, studi morfogenetika yang dilakukan pada penelitian ini masih merupakan informasi awal mengenai karakter produksi getah pada P. merkusii
kandidat bocor getah, masih diperlukan adanya studi mendalam mengenai peranan gen dalam menentukan karakter bocor getah. Seperti halnya pada jenis pinus lain, karakter produksi getah memiliki heritabilitas tinggi, sehingga program pemuliaan sangat efektif diterapkan untuk peningkatan produktivitas getah. Beberapa pendekatan molekuler dapat dilakukan untuk memfasilitasi tujuan tersebut, salah satunya dengan marker assisted selection (MAS).
Marker assisted selection merupakan metode seleksi yang mengacu pada
pemanfaatan marka DNA yang berpautan dengan lokus target (genetic linkage
mapping) sebagai alat untuk menduga dan membantu seleksi fenotipe sifat yang
menjadi target pemuliaan. Peta tautan genetika (genetic linkage mapping) yang dihasilkan melalui pendekatan MAS akan memudahkan pemulia tanaman dalam mengidentifikasi lokus karakter kuantitatif tertentu (quantitative trait loci/ QTL). Menurut Krutovsky (2004), metode MAS cukup aplikatif untuk jenis-jenis konifer
yang memiliki ukuran genom cukup besar dan sangat bermanfaat untuk menemukan segmen kromosom homologus yang berperan mengontrol karakter tertentu. Teknik lain untuk deteksi gen pada jenis yang memiliki genom besar seperti pinus adalah dengan penanda SAMPL (Selective Amplification of
Microsatellite Polymorphic Loci). Penanda SAMPL merupakan kombinasi
kelebihan yang dimiliki oleh penanda AFLPs dan mikrosatelit, sehingga hasil amplifikasinya memiliki persentase polimorfik dan repeatibilty yang tinggi (Paglia & Morgenate 1998). Metode-metode tersebut telah digunakan pada P.
pinaster dan P.taeda dan memberikan hasil yang cukup menjanjikan. Pada P.
pinaster, De Miguel et al. (2012) telah menggunakan metode-metode tersebut dan
berhasil menemukan peta tautan gen pengendali produksi getah
Sampai saat ini kegiatan pemuliaan pada P.taeda dan P.pinaster lebih maju dibandingkan pada pinus lain. Kegiatan pemuliaan pada kedua pinus tersebut telah menggunakan pendekatan peta tautan genetika dengan informasi fungsional gen tertentu. Peta tautan genetika fungsional telah teruji memiliki kemampuan menyediakan penanda baru dari daerah coding tertentu (Komulainen et al. 2003, Tani et al. 2003) seperti EST-Ps (Expressed Sequence Tags Polymorphisms), EST-SSRs (EST derived microsatellites) dan SNPs (Single Nucleotide
Polymorphisms). Konstruksi-konstruksi marka tersebut dapat digunakan untuk
kegiatan pemuliaan konifer yang memiliki genom cukup luas. Berdasarkan informasi keberhasilan pada jenis P. taeda dan P. pinaster, marka-marka tersebut juga berpeluang untuk deteksi gen yang mengendalikan sifat produksi getah pada
P. merkusii.
3.4 Simpulan
Hasil perhitungan koefisien variasi genetika dan heritabilitas untuk karakter produksi getah menunjukkan nilai yang tinggi (KVG:14.5-28.43%; h2: 0.58 0.08-0.770.08), hal tersebut mengindikasikan karakter produksi getah dipengaruhi oleh faktor genetika. Berdasarkan nilai tersebut kegiatan seleksi massa untuk individu kandidat bocor sesuai lokasi KBS mampu memberikan harapan perolehan genetika yang baik, karena merupakan cerminan fenotipe pohon yang adaptif dengan KBS tertentu.
Evaluasi kandidat bocor getah dengan bantuan penanda molekuler mikrosatelit memberikan hasil sesuai evaluasi fenotipik yang telah dilakukan. Hal tersebut ditunjukkan dengan adanya perbedaan struktur alel, nilai heterozigositas dan pola pengelompokan terpisah . Hal tersebut mengindikasikan secara genetika kandidat bocor getah berbeda dengan pinus normal. Hasil perhitungan sumber keragaman menunjukkan sebanyak 79% terdapat keragaman berada di dalam populasi dan 21% berada antar populasi sehingga strategi seleksi bocor getah selanjutnya lebih difokuskan dalam populasi.
Hasil korelasi antara fenotipe dan genotipe menemukan karakter diameter batang, panjang tajuk, dan tebal kulit berhubungan positif nyata dengan produksi getah sedangkan karakter jumlah cabang dan tingkat serangan hama dan penyakit berhubungan negatif nyata dengan produksi getah. Sehingga selanjutnya karakter-karakter tersebut dapat digunakan sebagai indikator untuk seleksi awal kandidat bocor getah.
Tabel 3.12 Korelasi fenotipik (di atas diagonal) dan genetika (di bawah diagonal) antar karakter produksi getah dan komponen hasil lainnya
Karakter Prod TTB TBC D TKB PT LT CPT SCP HP VBC Vtot
Set 1 dan set 2
Prod 1 0.014 -0.112 0.584** 0.299 0.176 0.096 -0.496* -0.023 -0.517* -0.284 -0.254 TTB 0.121 1 0.204 -0.043 -0.283 0.316 0.716* 0.155 0.252 0.047 0.163 0.411 TBC 0.106 0.445 1 -0.344 -0.435 -0.187 0.033 0.240 -0.200 0.306 0.490* -0.0.81 D 0.650* 0.272 0.236 1 0.602** 0.128 -0.105 -0.096 -0.143 -0.241 0.183 0.387 TKB 0.186 0.774 0.672 0.415 1 -0.063 -0.144 -0.145 -0.011 -0.119 0.065 0.245 PT 0.129 0.540 0.649 0.290 0.248 1 0.323 -0.150 0.053 -0.204 -0.159 0.069 LT 0.135 0.174 0.490 0.302 0.254 0.500 1 0.141 0.281 0.198 0.030 0.286 CPT -0.052 0.219 0.190 0.117 0.334 0.233 0.243 1 0.047 0.357 0.321 0.256 SCP 0.044 0.186 0.162 0.100 0.284 0.198 0.207 0.08 1 0.114 0.038 0.233 HP 0.041 0.174 0.151 0.236 0.265 0.162 0.193 0.07 0.06 1 0.318 0.254 VBC 0.366 0.152 0.132 0.472 0.232 0.162 0.169 0.254 0.305 0.322 1 0.751** Vtot 0.280 0.116 0.101 0.623 0.178 0.124 0.124 0.204 0.409 0.400 0.356 1
Set 1 (umur 30 tahun)
Prod TTB TBC D TKB PT LT CPT SCP HP VBC Vtot Prod 1 0.329 0.297 0.429* 0.403* 0.426* 0.122 -0.355* 0.238 -0.321* 0.564 0.469 TTB 0.208 1 0.198 0.575 0.070 0.195 0.156 0.285 0.642 -0.179 0.498 0.838* TBC 0.164 0.681 1 0.059 -0.339 0.369 0.444 -0.188 -0.124 -0.086 0.720* 0.085 D 0.561* 0.424 0.336 1 0.469 0.082 0.160 -0.268 -0.428 -0.192 0.731* 0.933** TKB 0.345 0.109 0.664 0.533 1 -0.506 -0.097 0.190 0.560 0.349 0.097 0.338 PT 0.150 0.627 0.311 0.307 0.689 1 0.282 -0.290 -0.071 -0.451 0.058 0.281 LT 0.183 0.762 0.497 0.373 0.851 0.551 1 0.204 0.145 -0.047 0.157 0.549 CPT -0.094 0.311 0.311 0.192 0.619 0.284 0.345 1 0.174 -0.190 0.141 0.004 SCP 0.118 0.393 0.390 0.240 0.494 0.356 0.551 0.223 1 0.045 0.180 -0.059 HP -0.060 0.254 0.198 0.315 0.315 0.181 0.220 0.113 0.142 1 -0.179 0.174 VBC 0.252 0.219 0.174 0.505 0.276 0.159 0.193 0.667 0.124 0.63 1 0.683 Vtot 0.378 0.157 0.125 0.552 0.198 0.114 0.138 0.576 0.689 0.455 0.400 1
Karakter Prod TTB TBC D TKB PT LT CPT SCP HP VBC Vtot
Set 2 (umur 29 tahun)
Prod 1 0.045 -0.126 0.555* 0.094 0.397* 0.060 -0.714* -0.162 -0.666* 0.474 0.456 TTB -0.056 1 0.178 -0.491 0.356* 0.349 0.300* -0.029 0.550 -0.115 0.400 0.442* TBC -0.076 0.235 1 -0.423 -0.097 -0.516 -0.428 0.014 -0.641 -0.094 0.031 0.042 D 0.499* 0.107 0.296 1 0.653 -0.187 -0.218 -0.187 -0.259 -0.084 0.227 0.333 TKB 0.355 0.051 0.519 0.116 1 -0.283 -0.647 0.298 -0.181 0.428 0.664 0.580 PT 0.265 0.267 0.191 0.345 0.105 1 0.405 0.080 0.152 -0.125 -0.579 -0.254 LT 0.065 0.291 0.291 0.406 0.135 0.320 1 -0.205 0.630 0.115 -0.143 0.239 CPT -0.489 0.091 0.473 0.435 0.043 0.133 0.333 1 0.096 0.532* 0.301 0.326 SCP 0.075 0.087 0.087 0.335 0.132 0.094 0.201 0.0192 1 0.115 -0.143 0.239 HP -0.496 0.065 0.135 0.085 0.235 0.192 0.434 0.453 0.198 1 0.193 0.169 VBC 0.126 0.035 0.079 0.421 0.256 0.132 0.177 0.156 0.253 0.067 1 0.687 Vtot 0.278 0.230 0.217 0.435 0.299 0.343 0.192 0.257 0.198 0.099 0.188 1
Keteragan: **: sangat berbeda nyata pada selang kepercayaan99% * : berbeda nyata pada selang kepercayaan 95%