Beradasarkan keparalelan Hukum Mendel dengan perilaku kromosom dalam proses
meiosis disimpulkan bahwa gen terletak pada kromosom (Bab 1 dan Bab 2). Pertanyaan
selanjutnya bagaimanakah struktur kimia dari bahan genetik yang ada pada kromosom
tersebut. Kromosom tersusun atas dua komponen yaitu DNA (asam deoksiribonukleat) dan
protein. Pada awalnya para ilmuwan menduga komponen proteinlah yang merupakan bahan
genetik; dengan alasan bahwa enzim (protein katalisastor) berperan dalam menjalankan
reaksi kimia metabolisme. Namun kemudian sejumlah penelitian membuktikan bahwa
komponen DNA pada kromosom yang merupakan bahan genetik. Pada saat sekarang
diketahui bahwa ada sebagian virus yang bahan genetiknya tersusun atas RNA (asam
ribonukleat). Percobaan-percobaan tersebut menunjukan bahwa (i) DNA merupakan senyawa
khas kromosom, (ii) DNA berperan dalam proses transformasi genetik pada bakteri; dan (iii)
Asam Nukleat yang berperanan dalam pewarisan sifat dari satu generasi ke generasi
berikutnya. dan penentuan sifat virus.
A. DNA Berperan Dalam Proses Transformasi Bakteri
Bukti DNA sebagai bahan dasar genetik pertama kali ditunjukkan pada tahun 1943 oleh
Oswald T. Avery, Colin McLeod, dan McLyn McCarthy dari Rockefeller Institute, yang
telah membuktikan bahwa DNA yang terlibat dalam proses transformasi bakteri.
Transformasi bakteri itu sendiri sebelumnya telah dikemukakan oleh F. Griffith (1928),
melalui percobaan pada Streptococcus pneumoniae, yaitu bakteri yang dapat menimbulkan
penyakit pneumonia pada manusia, dan menyebabkan kematian pada tikus.
Transformasi bakteri didefinisikan sebagai proses perubahan sifat bakteri akibat pengambilan DNA asing oleh sel bakteri tersebut secara langsung dari lingkungannya. DNA
asing tersebut dapat berintegrasi dengan DNA kromosom bakteri yang mengambilnya,
sehingga dapat diwariskan pada generasi berikutnya.
3
BAB
STRUKTUR MOLEKULAR
BAHAN GENETIK
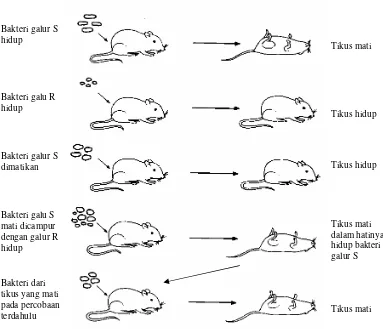
Griffith menunjukkan adanya proses transformasi ini melalui pencampuran satu galur
bakteri yang hidup dengan galur lainnya yang telah dimatikan. Diperoleh hasil bahwa galur
bakteri yang hidup itu dapat berubah sifat akibat masuknya bahan-bahan dari galur bakteri
yang telah dimatikan. Dalam percobaannya, Griffith telah menyuntikkan ke dalam tikus dua
galur S. pneumoniae yaitu yang berkulit halus (galur S) dan yang berkulit kasar (galur R).
Kulit halus pada galur S disebabkan oleh adanya mantel lendir polisakarida yang
menyelimuti bagian luar sel, sedangkan yang berkulit kasar tidak mempunyai lendir tersebut.
Adanya lendir itu menyebabkan galur S tahan terhadap antibodi yang dihasilkan oleh tikus,
sehingga bakteri tersebut tetap hidup dan dapat mematikan tikus. Galur R yang tidak
berlendir akan terbunuh oleh antibodi yang dihasilkan oleh tikus.
Skema percobaan Griffith ditunjukkan pada Gambar 3.1. Pada percobaan pertama tikus
disuntik dengan galur S atau galur R hidup, dan diperoleh hasil galur S mematikan, tetapi
galur R tidak mematikan. Dalam hati tikus yang mati terdapat sel bakteri galur R. Pada
Gambar 3.1. Rangkaian percobaan Griffith yang menunjukkan adanya transformasi bakteri, perubahan bakteri galur R menjadi seperti galur S akibat adanya bahan yang berasal dari galur S masuk ke sel galur R.
percobaan selanjutnya sebelum disuntikkan ke dalam tubuh tikus, galur R dan galur S bakteri
tersebut di atas dimatikan terlebih dahulu dengan cara pemanasan. Hasilnya seperti yang telah
diduga sebelumnya yaitu bahwa bakteri-bakteri tersebut tidak dapat mematikan tikus. Pada
percobaan ketiga dicampurkan galur R hidup dengan galur S yang telah dimatikan, dan
kemudian disuntikkan ke dalam tubuh tikus. Diperoleh bahwa tikus mati, dan ketika diperiksa
hatinya penuh dengan bakteri yang berlendir seperti galur S. Bakteri yang diambil dari tikus
yang mati itu ketika diinfeksikan kepada tikus lain ternyata juga mematikan.
Dari hasil rangkaian percobaan itu Griffith menarik kesimpulan bahwa telah terjadi
perubahan genetik dari sel galur R yang mulanya tidak mampu membentuk lendir menjadi
berkemampuan menghasilkan lendir seperti galur S. Kemampuan tersebut diperoleh karena
adanya kepingan bahan genetik dari galur S yang telah dimatikan yang masuk ke dalam sel
galur R. Proses masuknya bahan genetik dari pecahan galur S ke dalam sel galur R secara
langsung disebut transformasi genetik.
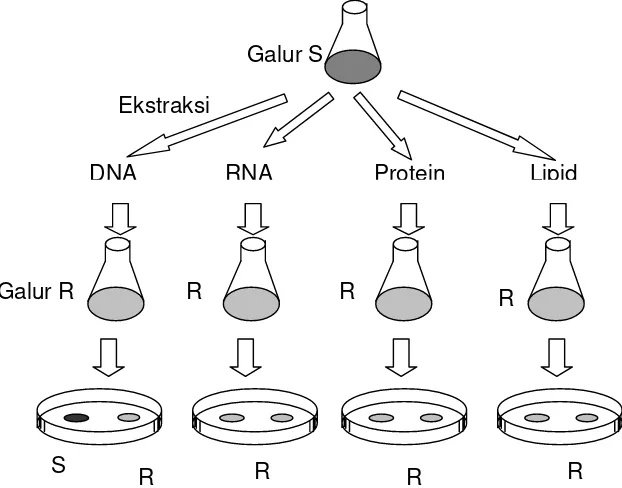
Pada tahun 1944, Avery, McLeod dan McCarthy berhasil membuktikan bahwa bahan
genetik yang terlibat dalam proses transformasi tersebut adalah DNA. Avery dkk melakukan
percobaan dengan cara mengisolasi berbagai molekul kimia dari pecahan sel galur S.
bahan yang berhasil diisolasi terdiri dari DNA, RNA, protein, polisakarida, dan lipid.
Bahan-Gambar 3.2. Hanya komponen DNA dari galur S yang dapat merubah galur R menjadi seperti galur S, berarti DNAlah yang merupakan bahan genetik yang terlibat dalam transformasi.
Galur S
Galur R
R R R R
S
R R R
DNA RNA Protein Lipid
bahan tersebut kemudian dicampurkan dengan sel galur R, ternyata hanya DNA yang dapat
merubah sel galur R menjadi sel yang bersifat seperti galur S (Gambar 3.2). Berarti DNAlah
yang bertanggung jawab dalam proses transformasi bakteri.
A. Asam Nukleat Merupakan Makromolekul
Asam nukleat merupakan salah satu makromolekul yang terdapat di dalam sel.
Makromolekul ialah molekul besar (berat molekul lebih dari 1000) yang disusun oleh
molekul-molekul dasar atau subunit yang lebih kecil. Dikenal empat jenis makromolekul
penting untuk kehidupan, yaitu:
1. Polinukleotida: Makromolekul dengan nukleotida sebagai molekul dasarnya. Asam
nukleat merupakan senyawa polinukleotida; dan terdapat dua jenis asam nukleat yaitu
Asam Deoksiribonukleat (DNA), dan Asam Ribonukleat (RNA)
2. Polipeptida: Makromolekul yang merupakan rangkaian asam amino. Polipeptida
merupakan subunit dasar penyusun molekul protein.
Struktur Dan Sifat Kimia DNA
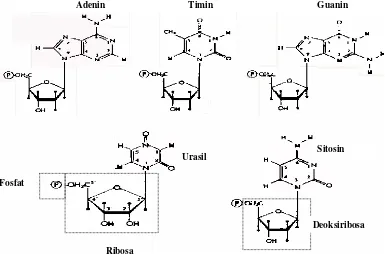
Gambar 3.3. Nukleotida adalah subunit penyusun rantai polinukleotida. Nukleotida disusun oleh Gula. Basa, dan fosfat. Terdapat lima jenis basa (Adenin, Guanin, Sitosin, Timin, Urasil) dan dua jenis gula (Ribosa dan Deoksiribosa)
Fosfat
Ribosa
Deoksiribosa
Urasil Sitosin
Guanin
3. Polisakarida: Makromolekul dengan sakarida sebagai bahan dasar penyuysun rantainya.
Polisakarida mencakup berbagai senyawa yang dilibatkan dalam reaksi metabolisme.
4. Lemak: merupakan rangkaian asam lemak.
Telah disebutkan bahwa nukleotida merupakan molekul dasar dari polinukleotida.
Nukleotida itu sendiri disusun oleh tiga komponen utama; gula, basa, dan fosfor (Gambar
3.3). Terdapat dua jenis gula, yaitu Ribosa dan Deoksiribosa; dan dua kelompok basa, yaitu
Purin dan Pirimidin. Kelompok purin terdiri dari Adenin (A) dan Guanin (G), sedangkan
pirimidin terdiri dari Sitosin (C), Timin (T), dan Urasil (U). DNA dan RNA dibedakan oleh
gula dan basa yang dikandungnya. Deoksiribosa dan Timin hanya dikandung oleh DNA;
sedangkan Ribosa dan Urasil dikandung oleh RNA.
Gula dengan basa membentuk ikatan antara Cl pada gula dengan N1 basa purin atau N6
basa pirimidin; dan senyawa yang dibentuknya disebut nukleosida. Selanjutnya fosfat
membentuk ester dengan nukleosida tersebut melalui pembentukan ikatan pada C5 gula.
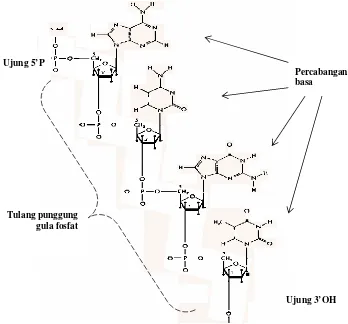
Gambar 3.4. Rantai polinukliotida, merupakan rangkaian nukleotida; fosfat pada C5 satu nukleotida dihubungkan dengan C3 dari nukleotida lain sehingga akan terbentuk tulang punggung gula fosfat, dan basa sebagai cabang. Rantai nukleotida mempunyai ujung C5 fosfat dan C3-OH
Tulang punggung gula fosfat Ujung 5’P
Esterfosfat-5-nukleosida ini disebut nukleotida. Berdasarkan kandungan gulanya terdapat dua
jenis nukleotida, yaitu ribonukleat (dikandung oleh RNA) dan deoksiribonukleotida (yang
dikandung DNA). Dalam keadaan bebas nukleotida-nukleotida berada dalam bentuk
nukleotida-trifosfat. Juga dapat ditemukan dalam bentuk nukleotida difosfat serta
monofosfat.
Dalam rantai polinukleotida, nukleotida-nukleotida digabungkan satu dengan yang
lainnya melalui ikatan fosfodiester yaitu antara fosfat pada C5 dari suatu nukleotida dengan
C3 dari nukleotida yang lain (Gambar 3.4). Dengan aturan seperti ini, maka pada ujung rantai
polinukleotida akan ditemukan fosfat pada ujung 5 dan radikal OH pada ujung 3.
B. Model Heliks Ganda DNA
Walaupun secara kimia, DNA telah diketahui sejak beberapa dekade yang lalu, namun
strukturnya baru dapat dijelaskan pada tahun 1953 oleh James D. Watson seorang pakar
genetika dan biokimia Amerika, dan Francis Crick seorang pakar kedokteran Inggris. Erwin
Chargaff dari Columbia University yang mengadakan penelitian terhadap kandungan DNA
melaporkan pada tahun 1949 bahwa kuantitas kandungan Adenin (A) dalam DNA berbagai
organisme adalah sama dengan Timin (T), sedangkan Guanin (G) sama dengan Sitosin (S).
Perbedaan antara DNA dari spesies yang berlainan terletak pada kandungan A+T/G+C.
Untuk lebih jelasnya pada Tabel 3.1 disajikan data hasil penelitian pada berbagai organisme.
Tabel 3.1 Komposisi Basa DNA Berbagai Organisme
Organisme Komposisi Basa (%) Nisbah Basa
A T G C A/T G/C A+T/G+C
Manusia 30.9 29.4 19.9 19.8 1.05 1.00 1.52
Kambing 29.3 28.3 21.4 21.0 1.03 1.02 1.30
Kecambah-gandum 27.3 27.1 22.7 22.8 1.04 1.00 1.19
Khamir 31.3 32.9 18.7 17.1 0.95 1.09 1.79
Escherichia coli 24.7 23.6 26.0 25.7 1.04 1.01 0.93
Brucella abortus 21.0 21.1 29.0 28.9 1.00 1.00 0.72
Diringkas dari Lehninger, A.L. 1982. Principle of Biochemistry, Worth Publisher, New York
Data Chargaff ini membawa Watson dan Crick kepada kesimpulan bahwa DNA tersusun
atas dua utas poliknukleotida, dan terjadi perpasangan basa antara kedua rantai tersebut
dengan mengikuti aturan Chargaff, yaitu A berpasangan dengan T, dan G berpasangan
dengan Timin terdapat dua ikatan hidrogen, dan Sitosin dengan Guanin terdapat tiga ikatan
hidrogen. Ikatan hidrogen ini selain mengikat juga memelihara jarak antara kedua basa.
Beberapa persyaratan kimia yang dituntut oleh perpasangan basa ini ialah arah runtunan gula
kedua polinukleotida harus berlawanan, sehingga ujung 5'P dari satu utas nukleotida akan
berpasangan dengan ujung 3'OH dari utas pasangannya (Gambar 3.5). Perpasangan seperti ini
disebut perpasangan anti paralel
Data lain yang digunakan Watson untuk menjelaskan struktur DNA ialah hasil
studi kristalografi DNA yang dilakukan oleh Rosalind Franklin dan Maurice Wilkins (1953).
Berdasarkan difraksi sinar-X ditemukan adanya kisi atau celah berukuran sekitar 3.4 A0, dan
selanjutnya terdapat struktur yang berulang setiap 34 A0 (Gambar 3.6). Watson dan Crick
(1953), menggabungkan data difraksi sinar-X ini dengan penemuan Chargaff, dan
menyimpulkan bahwa kedua pasangan rantai polinukleotida tersebut berputar membentuk
Gambar 3.5. Dalam satu molekul DNA terdapat dua utas polinukleotida dengan basanya
berpasangan sesuai aturan Chargaff yaitu A-T dan G-C. Pasangan A-T diikat oleh dua ikatan hidrogen, G-C oleh tiga ikatan hidrogen.
Timin Adenin
Sitosin Guanin
Sampel DNA
Film foto
Sinar terdifraksi. Lebar celah d=~1/θ
Sinar X
Gambar 3.6. Hasil difraksi sinar X oleh kristal DNA, dari Franklin dan Wilkins, yang membawa Watson dan Crick kepada Struktur heliks ganda DNA. Bagian luar merupakan hasil difraksi oleh kisi dengan celah 3.4 A0dan bagian dalam hasil difraksi oleh kisi berelah 34 A0.
Celah 3.4 A0
suatu pilinan mengelilingi satu sumbu dengan orientasi ke arah kanan. Ikatan hidrogen yang
terdapat pada pasangan basa akan membentuk garis tegak lurus terhadap sumbu tersebut di
atas (Gambar 3.7). Celah sebesar 3.4 A0 merupakan celah antara dua kisi yang terbentuk oleh
ikatan hidrogen dua pasang basa yang berdampingan. Pengulangan struktur yang sama setiap
34 A0 akan terjadi seandainya setiap runtunan sepuluh pasang basa terbentuk satu pilinan.
Jadi antara dua jari-jari pasangan basa yang berurutan terbentuk sudut sebesar 3600, dan
setelah sepuluh pasang basa akan terbentuk satu lingkaran (3600).
.Beberapa sifat kimia penting dari molekul DNA ialah (1) bahwa pasangan basa dari dua
utasan polinukleotida diikat oleh ikatan hidrogen. Ikatan ini memberikan kelenturan kepada
DNA, yaitu bahwa kedua rantai polenukleotida dapat berpisah dan bergabung kembali tanpa
mengalami kerusakan. Hal ini penting bagi DNA dalam proses replikasi (perbanyakan
molekul DNA) serta proses transkripsi (ekspresi gen). (2) Antara dua nukleotida dalam satu
rantai polinukleotida diikat dengan ikatan kovalen. Ikatan ini sangat kuat tetapi untuk
membentuknya diperlukan suatu reaksi kimia yang akan melibatkan energi. Sehingga
Gambar 3.7. Model heliks ganda DNA menurut Watson dan Crick. Pasangan dua utas polinukleotida akan berpilin mengitari sebuah sumbu dengan pilihan ke kanan. Dalam satu putaran penuh terdapat 10 pasang basa, antar pasangan basa terdapat jarak 3.4 A0 atau 34 A0 untuk sepuluh pasang basa.
A
Pasangan antiparalel Heliks ganda
3.4 A0
34 A0
20 A0
Relung mayor
kerusakan pada ikatan antara dua nukleotida dapat menyebabkan kerusakan fatal pada
molekul DNA. (3) Fosfat merupakan molekul yang hidrofil, sehingga akan berorientasi luar,
sedangkan basa-basa merupakan molekul hidrofob, sehingga akan berorientasi dalam.
Dengan posisi fosfat terletak di luar dan basa terletak di dalam heliks maka dapat dicegah
masuknya molekul air ke dalam DNA, sehingga dapat menghindarkan dari kerusakan akibat
reaksi kimia. Sifat ini cukup untuk memelihara kestabilan molekul DNA.
C. Implikasi Genetik dari Model Heliks Ganda
DNA sebagai bahan genetik, yang merupakan pusat pengendali sifat-sifat organisme,
harus mempunyai struktur yang stabil. Kestabilan ini meliputi pertama ketahanan terhadap
kerusakan akibat reaksi kimia, dan kedua kestabilan genetik yaitu tidak terjadinya perubahan
selama proses replikasi.
Secara kima di atas telah dijelaskan bahwa struktur heliks ganda dapat melindungi DNA
dari kerusakan akibat reaksi kimia yang terdapat dalam sel. Secara genetika menurut Watson
adanya utas ganda dalam suatu molekul DNA memberikan kemampuan kepada molekul
tersebut untuk membentuk dirinya sendiri. Hal ini penting dalam proses reproduksi atau
replikasi. Dalam kimia, sistem komplementasi berguna untuk mengontrol ketepatan
perpasangan dua molekul. Model antiparalel antara dua utasan polinukleotida merupakan
sistem komplementasi antara basa-basa dari dua rantai tersebut. Basa-basa inilah yang
menjadi kunci dalam proses perbanyakan diri DNA, yang memberikan kemampuan molekul
DNA untuk mencetak dirinya sendiri dan mengontrol ketepatan dari hasil cetakannya.
memung-Setiap utasan rantai polinukleotida akan dapat menjadi cetakan dalam pembentukan
molekul DNA baru, dan sistem komplementasi antar basa menjadi patokan dalam proses
pencetakan tersebut. Karena dalam setiap molekul ada dua rantai yang merupakan
komplemen satu terhadap yang lain, maka bila setiap utasan polinukleotida itu membentuk
komplemennya, akan diperoleh dua molekul yang strukturnya sama dengan molekul awal
(Gambar 3.8 Proses perbanyakan diri DNA seperti itu disebut proses replikasi
semikonservatif.
Genom ialah gugus atau himpunan gen yang lengkap dari suatu organisme yang dapat
mengendalikan jalannya keseluruhan metabolisme sehingga organisme tersebut dapat hidup
dengan sempurna. Banyaknya gen yang terdapat dalam suatu genom berbeda dari suatu
organisme ke organisme yang lain. Semakin rumit suatu organisme semakin banyak gen yang
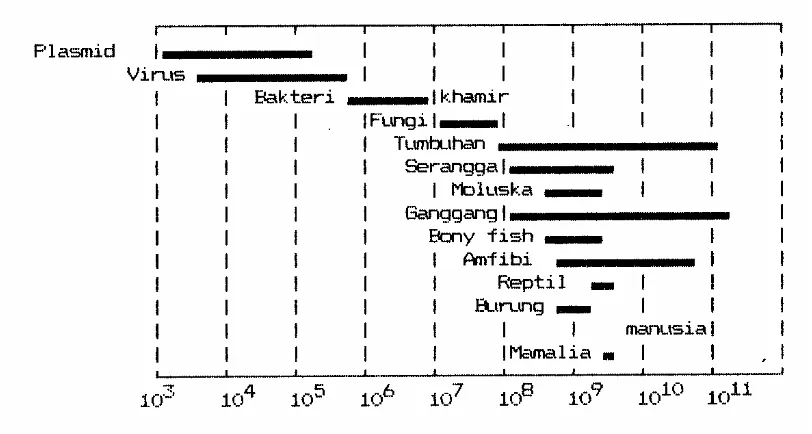
dikandung di dalam genomnya.Pada Gambar 3.9 diperlihatkan perbandingan ukuran genom
berbagai organisme berdasarkan kandungan DNAnya. Virus mempunyai genom kecil yang
terdiri dari sepotong asam nukleat yang mengandung beberapa gen. Bakteri mempunyai
ukuran genom puluhan ribu kali lebih besar ketimbang virus. Eukaroit mempunyai genom
paling besar; dalam inti eukariot diploid yang disebut genom setara dengan satu gugus
kromosom haploid
A. Genom Virus
Kebanyakan virus genomnya terdiri dari satu kromosom, terdapat sejumlah virus dengan
genom lebih dari satu kromosm. Berdasarkan bahan dasar penyusunnya genom virus dapat
dibagi menjadi dua jenis yaitu Virus DNA dan Virus RNA. Sebagian besar virus yang
menyerang tumbuhan adalah virus RNA. DNA atau RNA penyusun genom virus terdapat
dalam bentuk utas tunggal (ut) atau utas ganda (ug). Jadi terdapat virus DNAug, DNAut,
RNAug, dan RNAut. Khusus untuk molekul berutas tunggal (DNAut atau RNAut) terdapat
dalam dua jenis utasan yaitu utas positif (+) atau utas negatif (-). Yang dimaksud dengan utas
(+) ialah utasan yang basa-basanya homolog dengan basa-basa mRNA, sedang utas (-)
merupakan komplemen atau antiparalel terhadap mRNA.
Berdasarkan jenis genom serta cara reproduksinya, Baltimore (1971), mengajukan
klasifikasi virus, yang dikenal sebagai klasifikasi Baltimore (Tabel 3.2). Keterangan
mengenai jenis genom virus telah disampaikan pada paragraf terdahulu. Reproduksi atau
perbanyakan diri virus kelas I sampai kelas V dilakukan melalui proses replikasi, yaitu proses
pembentukan molekul baru, baik DNA maupun RNA, dengan menggunakan molekul lama
sebagai model cetakannya. Virus kelas VI memperbanyak diri melalui proses transkripsi
balik, yaitu pembentukan DNA dengan menggunakan RNA sebagai model cetakannya, dan
selanjutnya DNA yang terbentuk digunakan untuk membentuk RNA virus baru.
Tabel 3.2. Genom virus menurut klasifikasi Baltimore
Kelas Jenis Genom Contoh
I DNA ug fT4, fl, Adenovirus, virus herpes, SV40
II DNA ut(+) fX174, M13, MVM
III RNA ug Reovirus
IV RNA ut(+) MS2, virus polio, virus penyakit kuku dan mulut
ternak
V RNA ut(-) virus rabies
VI RNA ut(-) leukemia, virus AIDS
B. Genom Bakteri
Genom bakteri terdiri dari DNA kromosom dan DNA ekstrakromosom atau plasmid.
Karena bakteri tidak mempunyai pemisahan antara inti dan sitoplasma, maka kromosom dan
plasmid kedua-duanya terdapat dalam satu ruang plasma sel yang sama. Pembeda utama yang
paling mudah dilihat antara keduanya adalah dari segi ukurannya; plasmid mempunyai
ukuran yang relatif sangat kecil dibandingkan dengan kromosom. Ukuran kromosom bakteri
masih tergolong kecil bila dibandingkan dengan kromosom eukariot (lihat Gambar 3.10); dan
karena kecilnya kromosom ini tidak tampak di bawah mikroskop cahaya, tetapi sebaliknya
lebih mudah dipelajari secara molekular.
B1. Kromosom
Sebagian besar bakteri diketahui mempunyai satu kromosom yang disusun oleh satu
molekul DNA berbentuk sirkular. Pada akhir dekade delapanpuluhan abad duapuluh berbagai
hasil riset membuktikan adanya bakteri yang mempunyai kromosom linear, dan adanya
bakteri yang mempunyai dua kromosom.
Pembuktian pertama tentang bentuk kromosom bakteri dilakukan oleh Cairns (1960)
melalui percobaan pada Escherichia coli. Cairns berhasil mengembangkan teknik isolasi
DNA bakteri, dan memperoleh satu kromosom atau DNA yang utuh. Kemudian DNA
tersebut dipotret dengan teknik autoradiografi. Pada prinsipnya teknik autoradiografi terdiri
dari penandaan DNA dengan unsur radioisotop, dan kemudian DNA yang telah ditandai
tersebut diletakkan pada film negatif. Setelah dibiarkan dalam beberapa hari, radioisotop akan
menandai film tersebut. Dari jejak tanda yang terlihat dalam film dapat diketahui bahwa
kromosom bakteri E. coli mempunyai bentuk sirkular, dengan panjang sekitar 1360 mm.
E. coli mempunyai panjang sel sekitar 1 mm, sampai 2 mm sedangkan kromosomnya
mempunyai panjang sekitar 1360 mm, yang berarti panjang kromosomnya sekitar seribu kali
panjang selnya. Oleh karena itu untuk dapat berada di dalam sel molekul DNA tersebut harus
dibentuk dalam suatu kemasan yang lebih kompak. Dalam sel DNA, kromosom akan
berkondensasi membentuk struktur yang disebut nukleosid, yang menempati sekitar 10
persen dari volume sel bakteri. Proses kondensasi berlangsung dengan cara pelipatan dan
penggulungan.
B2. Plasmid
Plasmid merupakan DNA ekstrakromosom yang dikenal pada bakteri, merupakan DNA
berutas ganda dan berbentuk sirkular. Plasmid berukuran jauh lebih kecil dari kromosom
bakteri (BM 106 – 2 x 108), membawa sekitar 1-3 persen genom sel. Berbeda dari kromosom
keberadaan plasmid dalam sel tidak mutlak; dalam spesies yang sama suatu sel bakteri
mungkin mengandung satu jenis plasmid namun bakteri lain tidak memilikinya. dari segi
jumlahnya dalam suatu sel, plasmid dapat dibedakan menjadi plasmid dengan salinan yang
banyak (relaxed plasmid) atau dengan salinan terbatas (stringent plasmid)
Dari segi fungsinya plasmid dibagi menjadi plasmid konjugatif dan plasmid nirkonjugatif.
operon tra yang berperan dalam transfer plasmid ke sel bakteri lain. Sebagai contoh dari
plasmid konjugatif ialah plasmid F pada E. coli, yang berperan dalam pembentukan fili serta
replikasi tipe sigma. Dengan fili bakteri F+ akan dapat melakukan kontak dengan bakteri F-
dan menyalurkan DNA plasmid linear yang dihasilkan dari replikasi tipe sigma.
Plasmid menyandikan sejumlah sifat penting yang umumnya tidak disandikan oleh gen
pada kromosom bakteri. Sejumlah sifat yang diketahui disandikan oleh gen plasmid antara
lain resistensi terhadap antibiotik, produksi antibiotik, degradasi senyawa aromatik, produksi
haemolisin, fermentasi gula, produksi enterotoksin, resistensi terhadap logam berat, produksi
bakteriosin, merangsang tumor pada tumbuhan dan produksi hidrogen sulfida.
Agrobbacterium sp mempunyai kemampuan merangsang berkembangnya tumor pada
tumbuhan. Pada bakteri ini diketahui adanya satu plasmid, yaitu plasmid Ti yang berfungsi
dalam berhubungan dengan tumbuhan inang, yaitu dengan cara mentransfer satu fragmen
DNA (yaitu DNA-T) ke dalam sel inang, yang kemudian akan menyebabkan tumbuhnya
tumor pada inang. Kemampuan Agrobacterium mentransfer DNA T dan
mengintegrasikannya kedalam kromosom tanaman, sekarang telah dimanfaatkan dalam
rekayasa genetik. Juga plasmid secara umum banyak dimanfaatkan sebagai wahana atau
vektor dalam rekayasa genetik.
C. Genom Eukariot
Genom eukariot terbagi atas DNA-kromosom dan DNA-sitoplasma. Kromosom terdapat
di dalam inti sel dan DNA-sitoplasma terdapat di luar inti; oleh karena itu kedua jenis DNA
tersebut sering juga disebut sebagai DNA-inti dan DNA-sitoplasma. Pada sitoplasma, DNA
terdapat di dalam dua organel, yaitu mitokondria dan kloroplas.
C1. Kromosom Inti
Telah dijelaskan pada Bab 1 bahwa dalam inti eukariot terdapat sejumlah kromosom yang
terbagi ke dalam ploidi. Dalam satu ploidi terdapat satu set kromosom yang mencakum
seluruh jenis gen yang terdapat xdalam genom inti. Berdasrkan tingkat ploidinya satu sel
eukariot dapat merupak sel haploid, dipkoid dan poliploid. Pada sel diploid dan ploidi yang
lebih tinggi, setiap jenis kromosom terdapat dalam dua atau lebih salinan
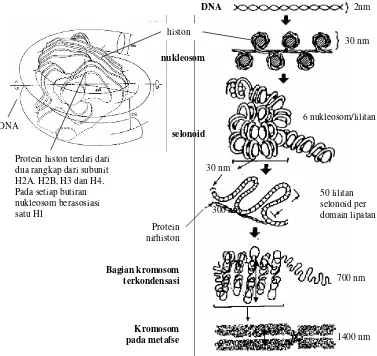
Dalam setiap kromosom eukariot terdapat satu molekul DNA yang berukuran sangat
panjang, yang berasosiasi dengan protein yang disebut histon. Protein histon disusun oleh
lima jenis subunit protein, yaitu H1, H2A, H2B, H3, dan H4. Asosiasi DNA dengan histon
bertujuan untuk melindungi molekul DNA dari kerusakan, misal pada saat migrasi kromosom
menggulung secara teratur sehingga ukurannya menjadi pendek dan tebal. seperti yang
tampak di bawah mikroskop cahaya
Di bawah mikroskop elektron, kromatin sering terlihat seperti serat yang mempunyai
rangkaian manik-manik yang diikat dengan serat halus, dengan butiran manik-maniknya
berukuran sekitar 100 A0. Serat halus tersebut adalah DNA dan manik-maniknya adalah
protein histon yang dililiti DNA. Asosiasi antara satu butiran histon dengan satu segmen
DNA itu disebut nukleosom (Gambar 3.10). Setiap butiran nukleosom disusun oleh unsur inti
dan subunit H1. Unsur inti disusun oleh 146 pasang basa DNA yang berasosiasi dengan
delapan subunit (tetramer) dari empat jenis subunit H2A, H2B, H3, dan H4 yang
masing-masing terdapat dalam dua rangkap (Gambar 3.10)
Asosiasi terjadi melalui interaksi antara protein yang bermuatan positif, berkat
Gambar 3.10 Asosiasi DNA dengan histon dalam kromosom eukariot. Menjelang pembelahan sel akan terjadi penggulungan DNA menjadi tebal sampai membentuk struktur kromosom yang tampak di bawah mikroskop.
DNA
kandungan Lisin dan Arginin, dengan fosfodiester DNA yang bermuatan negatif. Kemudian
kepada unsur inti nukleosom tersebut berasosiasi protein H1 serta 20 pasang basa DNA, yaitu
10pb masing-masing dihulu dan hilir DNA unsur inti nukleosom. Jadi pada satu nukleosom
keseluruhannya berasosiasi 166 pb DNA, dengan 5 jenis protein histon. Antara satu
nukleosom dengan yang lainnya dihubungkan oleh DNA penghubung yang mempunyai
panjang sekitar 34 pb. Sehingga untuk setiap nukleosom berasosiasi 200 pb. Satu nukleosom
mempunyai diameter sebesar 100 A0, dan dengan pembentukan nukleosom ini terjadi
pemendekan ukuran utasan DNA sekitar 7 kali dari panjang heliks bebas.
Pada awal pembelahan sel baik secara mitosis atau meiosis, akan terjadi kondensasi DNA
yang berlangsung melalui penggulungan bentuk rantai-rantai tersebut di atas. Penggulungan
ini berlangsung berkat adanya H1 yang mempunyai daya tarik ionik sehingga
nukleosom-nukleosom tersebut akan menempel membentuk suatu silinder dengan H1 di bagian tengah.
Silinder ini disebut solenoid dengan diameter sebesar 350 A0 (Gambar 3.9) Dengan
penggulungan solenoid ini terjadi pemendekan serat sebesar enam kali dibandingkan dengan
panjang serat nukleosom, atau sekitar 40 kali panjang DNA dalam bentuk heliks bebas.
Dari bentuk solenoid masih terjadi proses penggulungan lebih lanjut, tetapi
mekanismenya masih belum sejelas pada penggulungan solenoid. Salah satu model
penggulungan diajukan pada Gambar 3.9 dimana sebelum mencapai bentuk akhir kromosom
masih terdapat dua kali penggulungan setelah tingkat solenoid. Mula-mula solenoid digulung
dan menghasilkan tabung berlubang dengan garis tengah sekitar 300 A0. Selanjutnya pada
metafase saat peralihan dari bentuk kromosom ke kromatid, terjadi lagi proses penggulungan,
membentuk gulungan dengan garis tengah sekitar 14000 A0, yaitu bentuk akhir kromosom.
C2. DNA Sitoplasma
Dalam sitoplasma terdapat dua organel yang mengandung DNA, yaitu mitokondria dan
kloroplas. Mitokondria merupakan organel tempat metabolisme oksidatif dan fosforilasi;
dalam rangka respirasi. Sedangkan kloroplas merupakan organel tempat berlangsungnya
reaksi fotosisntesis. Dlam kedua organel tersebut terdapat DNA yang mengandung gen yang
masing-masing berperan untuk mendukung rsepirasi dan fotosintesis.
DNA mitokondria terdapat wilayah nukleoid mitokondria, yang terusun atas beberapa salinan molekul DNA sirkular bebas histon (kromosom). Molekul DNA mitokondria
mengandung 15 - 75 kilo pasangan basa, yang dapat menyandikan 16 - 80 macam protein,
yang diasumsikan masing-masing mempunyai panjang rata-rata 300 asam amino. DNA
mitokondria berperan menyandikan sistem enzim yang berhubungan dengan siklus Kreb.
besar dari protein mitokondria disandikan oleh DNA inti. Ekspresi gen mitokendria bebas
dari gen inti.
DNA kloroplas terdiri dari atas molekul DNA utas ganda sirkular. DNA ini menyandikan rRNA, tRNA dan sejumlah protein termasuk sejumlah besar subunit rubisco
dan sejumlah protein membran (protein klorofil fotosistem I). Replikasi DNA kloroplas
berjalan secara semikonservatif dan melibatkan model replikasi lingkaran berputar (rolling
circle)
Rangkuman
Melalui kejadian transformasi genetik pada bakteri telah dibuktikan bahwa DNA merupakan unsur genetik atau penentu sifat makhluk hidup. DNA merupakan suatu makromolekul yang tersusun oleh nukleotida sebagai molekul dasarnya; sedangkan nukleotida itu sendiri disusun oleh tiga komponen yaitu gula (deoksiribonukleotida), fosfat, dan basa. Ada empat jenis basa pada DNA yaitu Sitosin, Guanin, Adenin, dan Timin. Di samping DNA ada asam nukleat lain yaitu RNA, yang dibedakan dari DNA oleh gula dan basanya. Pada RNA terdapat Ribosa sebagai pengganti Deoksiribosa, dan Urasil sebagai pengganti Timin pada DNA.
Nukleotida yang satu dirangkaikan dengan nukleotida lain oleh ikatan 3' - 5' fosfodiester membentuk suatu rantai polinukleotida. Dalam satu molekul DNA terdapat dua utas polinukleotida yang diikat oleh ikatan hidrogen yang terbentuk antara basa-basanya. Perpasangan A-T diikat oleh dua ikatan hidrogen dan perpasangan G-C oleh tiga ikatan hidrogen. Pasangan kedua utasan tersebut berpilin membentuk heliks ganda; pada satu pilinan terdapat 10 pasang basa dengan jarak antar nukleotida sebesar 3.4 A0 atau satu pilinan mempunyai panjang sekitar 34 A0. Pilinan heliks ganda mempunyai garis tengah 20 A0.
Struktur heliks ganda dengan gula fosfat yang terletak di bagian luar heliks dan basa di bagian dalam heliks, menjamin kestabilan struktur DNA. Kestabilan ini meliputi pertama ketahanan DNA terhadap kerusakan akibat keadaan lingkungannya, dan yang kedua kestabilan genetik yang berhubungan dengan ketepatan dalam proses replikasi. Struktur heliks-ganda memungkinkan adanya replikasi semikonservatif yang menjamin ketepatan proses replikasi. Untuk lebih menjaga kestabilan DNA, di dalam sel DNA berasosiasi dengan protein, seperti histon pada eukariot.
Besarnya DNA yang menyusun genom beragam dari satu organisme ke organisme lain. Genom virus berkisar dari 3 x 103 sampai 105 pasang basa, bakteri dari 5 x 105 sampai 107 pasang basa, eukariot dari 107 sampai 1011 pasang basa. Genom bakteri terdiri dari kromosom dan plasmid; dan sebagian besar bakteri mempunyai satu kromosom. Genom eukariot terdiri dari gen inti yang disusun oleh sejumlah kromosom, dan gen sitoplasma yang terdapat dalam bentuk DNA mitokondria dan DNA kloroplas.
Pilih A bila 1&2 benar, B bila 1&3 benar, C bila 2&3, D bila 1&2&3
1) Makromolekul adalah suatu molekul besar yang tersusun oleh molekul kecil sebagai penyusunnya. Molekul di bawah ini adalah contoh dari makro molekul....
1) Protein tersusun oleh banyak asam amino
2) Polisakarida tersusun oleh banyak gula
3) Nukleotida tersusun oleh gula, basa, dan fosfat A, B, C, D
2) Yang bukan penyusun DNA adalah.... 1) Urasil
2) Histon 3) Sitosin
A, B, C, D
3) Dengan adanya ikatan hidrogen antara pasangan basa DNA, maka....
1) Pasangan heliks ganda dapat diudar dan dipilin tanpa risiko kerusakan tinggi 2) Heliks ganda dapat diudar dalam proses replikasi dan translasi
3) Heliks ganda mudah rusak karena kekuatan ikatan hidrogennya sangat lemah A, B, C, D
4) Pada DNA-B berukuran panjang 510 A0 akan terdapat.... 1) 150 pasang basa
2) 10 simpul pilinan 3) 15 simpul pilinan
A, B, C, D
5) Perbedaan plasmid dan kromosom antara lain....
1) Keberadaan kromosom dalam sel adalah mutlak sedangkan plasmid tidak mutlak 2) Plasmid berukuran lebih kecil dari kromosom
3) Kromosom tidak dapat melakukan konjugasi, sedangkan semua plasmid dapat ditransfer melalui konjugasi
A, B, C, D 6) Pada eukaroit,
1) Semua DNA sitoplasma terdapat dalam mitokondria
2) Dalam satu mitokondria terdapat satu kromosom berbentuk sirkular 3) DNA mitokondria mempunyai ukuran sekitar ukuran kromosom bakteri
A, B, C, D
Adams RLP, Knowler JT, and Leader DP. 1986. The Biochemistry of Nucleic Acids. Tenth Edition. Chapman and Hall. London, New York.
Albert B, Bray D, Lewis J, Raff M, Roberts K, and Watson JD. 1994. Molecular Biology of The Cell. Third Edition. Garland Publishing Inc. New York, London.
Chargaff E, Visher E, Doninger R, Green C, and Missani F. 1949. The Composition of the Deoxyribonucleic Acid of Thymus and Spleen. J. Biol Chem. 177: 405-416
Lehninger, A.L. 1982. Principle of Biochemistry, Worth Publisher, New York
Levine, L. (ed) 1971. Papers of Genetics. A book of readings. Mosby International Edition. The C.V. Mosby Cmpany Saint Lovis.
Singer M and Berg P. 1991. Genes and Genom. University Science Books, Mill Valley California. Blackwell Scientific Publications. Oxford, London, Edinburgh, Melbourne, London, Berlin, Vienna
Stryer L. 1995. Biochemistry. WH Freeman and Company. New York
Watson J.D. and Crick FHC. 1953. The Structure of DNA. Cold Spring Harbour Symposia Quant Biol XVIII :123-131
Watson JD and Crick FHC. 1953. Genetical Implication of The Structure of Deoxyribo-nucleic Acid. Nature 171:738-740
Zubay G and Marmur J. (Eds). 1973. Papers in Biochemical Genetics. Second Edition. Holt, Rinehart and Winston, Inc. New York, Chicago, San Francisco, Atlanta, Dallas, Montreal, Toronto, London.