ENTOMOPATOGEN DARI BERBAGAI INANG DI ALAM
DAN PATOGENISITASNYA TERHADAP RAYAP TANAH
COPTOTERMES GESTROI WASMANN
Abstrak
Cendawan entomopatogen dari berbagai inang atau inokulum di alam yaitu dari ulat krop kubis (Crocidolomia pavonana F.), ulat grayak (Spodoptera litura F.), walang sangit (Leptocorisa oratorius F.), penghisap polong (Riptortus
linearis L.), rayap tanah (Coptotermes curvignathus Holmgren.), tanah dan pasir
telah diisolasi dan identifikasi untuk menentukan patogenisitasnya terhadap rayap tanah Coptotermes gestroi Wasmann, kemudian masing-masing isolat ditempatkan pada suhu 4 0C. Sebelum digunakan, semua isolat dikulturkan
kembali pada media agar Sabouraud Dextrose Agar with Yeast Extract (SDAY) dan data yang diperoleh dianalisis berdasarkan Rancangan Acak Lengkap satu faktor dengan uji sidik ragam. Perbedaan antara isolat diuji lanjut menggunakan Duncan's Multiple Range Test. Hasil isolasi mengindikasikan keragaman spesies cendawan lebih tinggi pada inang yang berasal dari hama tanaman terinfeksi dibandingkan dari sumber inokulum yang berasal dari tanah dan pasir, setelah diidentifikasi beberapa spesies cendawan yang ditemukan adalah: Beauveria
bassiana (Bals.) Vuill., Metarhizium anisopliae (Metsch.) Sorok, Metarhizium brunneum Petch, Paecilomyces fumosoroseus (Wize) Brown dan Smith, Penicillium citrinum Thom. Verticilium lecanii (Zimmermann), Aspergillus flavus Link., Myrothecium roridum Tode ExFR, Fusarium oxysporum Link., dan Fusarium solani Link., spesies B. bassiana paling dominan. Uji patogenisitas
mengindikasikan bahwa umumnya isolat bersifat patogen terhadap rayap dan dapat menyebabkan mortalitas lebih dari 60% setelah 6 hari inokulasi. Mortalitas rayap tertinggi disebabkan oleh M. anisopliae dari inang penghisap polong, M.
brunneum dari pasir, M. roridum dari tanah, B. bassiana dari inang walang
sangit, F. oxysporum dari inang ulat grayak, dan A. flavus dari inang rayap tanah yaitu dapat menyebabkan mortalitas rayap 100%, dan diikuti oleh kemampuan bersporulasi secara in vivo yang cukup tinggi (76,25% - 96,25%) kecuali spesies
M. anisopliae dan M. roridum. Kemudian isolat-isolat terseleksi disiapkan untuk
dipelajari karakterisasi fisiologisnya, hasilnya mengindikasikan: kemampuan berkecambah tertinggi oleh M. brunneum dari pasir, diameter koloni tertinggi oleh F. oxysporum dari inang ulat grayak dan kemampuan bersporulasi secara in
vitro tertinggi oleh B. bassiana dari inang walang sangit.
Kata kunci: Bio-kontrol, uji tapis, cendawan entomopatogen, patogenisitas,
Pendahuluan
Cendawan entomopatogen termasuk genera cendawan yang berasosiasi dengan serangga dan beberapa spesies arthropoda lainnya (seperti laba-laba dan kutu) dengan berbagai cara, yaitu sebagai saprofit, kommensalistik, parasit atau patogen (Boucias & Pendland 1998). Di pertanian dan perkebunan di Indonesia juga banyak ditemukan serangga terinfeksi oleh cendawan entomopatogen. Diharapkan cendawan tersebut dapat diisolasi dan dibiakkan pada media sintetik untuk dimanfaatkan sebagai agens pengendalian rayap secara hayati. Untuk tujuan ini diperlukan koleksi dan uji tapis cendawan entomopatogen dari berbagai inang atau sumber inokulum di alam untuk mengetahui tingkat keragaman spesies dan keefektifannya terhadap rayap. Menurut Keller dan Zimmermann (1989), cendawan entomopatogen hanya dapat menginfeksi satu atau beberapa jenis serangga saja, karena jenis serangga secara umum jarang ditemukan mempunyai tingkat kerentanan yang serupa.
Di samping hal tersebut juga perlu dipelajari kemampuan cendawan bersporulasi secara in vivo dan in vitro serta informasi tentang karakter fisiologis lainnya. Hal ini penting dilakukan karena cendawan entomopatogen bersifat spesifik dalam hubungannya dengan inang dan potensinya untuk dapat diperbanyak secara massal, serta dapat tersebar luas setelah bersporulasi pada inang sasaran di dalam koloni. Menurut Behle et al. (1999), cendawan entomopatogen mempunyai keuntungan-keuntungan dibanding migro organisme patogen lainnya diantaranya: cendawan cenderung mempunyai sebaran inang yang lebih luas, beberapa cendawan entomopatogen menghasilkan spora yang toleran terhadap proses pengawetan lewat pengeringan dan menghasilkan stabilitas yang bagus dalam masa penyimpanan.
Pada penelitian pendahuluan, penggunaan cendawan entomopatogen yang diperoleh dari hama tanaman dengan kerapatan konidia 108 konidia/ml dapat menyebabkan mortalitas rayap C. gestroi 6 hari setelah inokulasi. Pada penelitian berikut, dicoba penggunaan cendawan entomopatogen dari berbagai inang atau inokulum di alam yang meliputi isolasi, identifikasi, dan uji tapis cendawan
dengan tujuan untuk mendapatkan isolat cendawan entomopatogen yang efektif dimanfaatkan sebagai pengendali rayap C. gestroi.
Bahan dan Metode
Penelitian ini telah dilakukan di Laboratorium Pengawetan Kayu UPT Balai Litbang Biomaterial Lembaga Ilmu Pengetahuan Indonesia (LIPI) Cibinong dan Laboratorium Patologi Serangga Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor (IPB).
Jenis Rayap yang Digunakan
Rayap yang digunakan terdiri dari kasta pekerja dan kasta prajurit spesies rayap tanah C. gestroi (Benson 2005) yang dipelihara di UPT Balai Litbang Biomaterial LIPI Cibinong selama 2 tahun.
Persiapan Isolat Cendawan
Sumber isolat cendawan entomopatogen dikoleksi dan diisolasi dari hama tanaman terinfeksi, tanah dan pasir mengacu pada Bab III.
Prosedur Isolasi
Prosedur isolasi telah dijelaskan pada Bab III.
Prosedur Identifikasi
Prosedur identifikasi telah dijelaskan pada Bab III.
Prosedur Perbanyakan
Perbanyakan dilakukan pada media SDAY dengan cara menginokulasikan konidia kultur murni cendawan di dalam cawan petri. Biakan diinkubasikan selama 3 minggu dalam inkubator dengan suhu 24 0C dan kelembaban relatif 95%. Untuk menjaga tingkat virulensinya sebelum perlakuan cendawan diremajakan dengan cara menginfeksikan ke serangga sasaran (rayap). Rayap
yang terinfeksi cendawan dibiarkan selama 2 minggu sampai bersporulasi (Ansari
et al. 2004), kemudian cendawan dari tubuh rayap diisolasi lagi sampai
didapatkan lagi kultur murni. Kultur murni tersebut dibiarkan bersporulasi sempurna kurang lebih 3 minggu sehingga siap digunakan sebagai bahan uji. Pada percobaan ini cendawan yang digunakan untuk pengujian adalah pada perbanyakan kedua (F2). Keberhasilan peremajaan ditentukan oleh berhasilnya cendawan menginfeksi dan bersporulasi pada tubuh rayap.
Penyediaan Suspensi Konidia
Suspensi konidia disiapkan dengan menambahkan 2 ml akuades steril yang telah mengandung 0,05% Triton X-100 ke dalam cawan petri berisi biakan cendawan yang telah berumur 3 minggu. Cawan digoyang-goyang, kemudian dengan bantuan kuas kecil steril konidia dapat terlepas. Konidia tersebut disaring dengan kain kasa dan dilakukan 4 kali pengenceran, selanjutnya dilakukan pengenceran dengan air sampai diperoleh konsentrasi yang diinginkan dengan kerapatan sebagai berikut: a). 107 konidia/ml untuk uji patogenisitas dan uji sporulasi in vivo, b). 106 konidia/ml untuk uji daya kecambah dan diameter koloni, c). 105 konidia/ml untuk uji sporulasi in vitro. Konidia dihitung dengan menggunakan haemocytometer.
Uji Patogenisitas
Suspensi konidia masing-masing isolat disiapkan seperti diterangkan di atas. Sebanyak 80 ekor kasta pekerja dan delapan ekor kasta prajurit rayap C.
gestroi dicelupkan kedalam 0,50 ml suspensi konidia yang telah diaduk. Kontrol
hanya dicelupkan dalam larutan 0,05% Triton X-100. Kertas saring Whatman no. 40 ditempatkan kedalam cawan petri sebagai sumber makanan kemudian rayap (20 ekor pekerja dan dua ekor prajurit) yang telah diinokulasi cendawan ditempatkan ke dalamnya. Semua perlakuan dipelihara pada ruangan dengan kondisi gelap dan mortalitas dihitung setiap hari selama satu minggu. Rayap yang mati diinkubasi pada suhu 24 0C dan kelembaban relatif 95% selama 5-7 hari untuk melihat sporulasi cendawan pada tubuh rayap yang telah mati. Percobaan diulang 4 kali.
Sporulasi in Vivo
Cendawan yang menyerang serangga akan bersporulasi yaitu miselianya tumbuh di permukaan tubuh rayap yang mati. Data untuk pengujian sporulasi in
vivo diambil dari data rayap yang mati pada uji patogenisitas yang diinkubasi
pada suhu 24 0C dan RH 95% selama 5-7 hari. Setiap perlakuan diulang 4 kali.
Persentasi sporulasi dihitung dengan rumus:
Sporulasi = Rayap terkolonisasi X 100% Jumlah rayap perlakuan
Karakterisasi Fisiologi Cendawan Entomopatogen in Vitro
Evaluasi daya kecambah konidia
Pengamatan daya kecambah konidia menggunakan metode yang dilakukan oleh Junianto dan Sukamto 1995 dalam Trizelia (2005). Media SDAY (Ø 0,5 cm tebal 1-2 mm) yang telah ditetesi suspensi konidia berkerapatan 106 konidia/ml
diletakkan di atas objek gelas steril, kemudian dimasukkan kedalam cawan petri dan diinkubasi pada suhu 24 0C selama 12 - 24 jam. Masing-masing perlakuan diulang 4 kali. Pengamatan dilakukan dibawah mikroskop dengan perbesaran 400 kali. Persentase konidia yang berkecambah dihitung dari 100 konidia. Konidia dinyatakan telah berkecambah apabila tabung kecambah (germ tubes) telah muncul lebih panjang dari diameter konidia.
Diameter koloni
Media SDAY yang telah ditumbuhi miselia masing-masing isolat berumur 5 hari dengan dimeter 0,8 cm ditumbuhkan pada media SDAY baru di dalam cawan petri dan diinkubasi dalam inkubator dengan suhu 24 0C. Diameter koloni dari masing-masing isolat diukur setelah hari ke 15.
Sporulasi in vitro
Untuk menghitung sporulasi masing-masing isolat pada media SDAY, disiapkan kerapatan suspensi konidia cendawan 105 konidia/ml. Suspensi konidia masing-masing isolat sebanyak 0,1 ml dikulturkan pada media SDAY dalam
cawan petri (Ø 9 cm), dan diinkubasi selama 15 hari pada suhu 24 0C. Kemudian biakan cendawan dimasukkan ke dalam wadah erlenmeyer dan ditambahkan 40 ml aquades steril, dikocok dengan vortex selama 5 menit, disaring dan dilakukan pengenceran sampai 4 kali. Kerapatan konidia dihitung menggunakan
haemocytometer dan rata-rata konidia untuk setiap cawan petri dibandingkan
antar isolat. Analisis Data
Data mortalitas, sporulasi in vivo dan karakterisasi fisiologis in vitro di analisis berdasarkan Rancangan Acak Lengkap (Completely Randomize Design) satu faktor (keragaman isolat atau spesies cendawan entomopatogen yang terdiri dari 16 isolat untuk uji mortalitas dan sporulasi in vivo, 6 spesies untuk uji karakterisasi fisiologis in vitro) dengan 4 x ulangan menggunakan analisis ragam (ANOVA), kemudian dilanjutkan dengan uji Duncan's Multiple Range Test. Bentuk umum dari persamaannya adalah sebagai berikut:
Dimana: i = 1,2,....17, t dan j= 1, 2, ....4, r
Yij = pengamatan pada perlakuan ke-i dan ulangan ke-j µ = rataan umum
δi = pengaruh perlakuan ke-i
εij = pengaruh acak pada perlakuan ke-i ulangan ke-j
Hasil dan Pembahasan
Isolasi dan Identifikasi
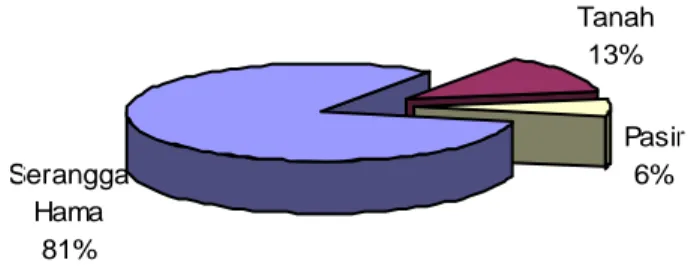
Hasil isolasi cendawan entomopatogen dari berbagai inang atau sumber inokulum di alam (Gambar 4.1) mengindikasikan keragaman spesies cendawan entomopatogen lebih tinggi pada inang yang berasal dari hama tanaman (ulat krop kubis, ulat grayak, penghisap polong, walang sangit dan rayap tanah sebanyak 81%) dibandingkan dengan yang berasal dari tanah (13%) dan pasir (6%). Berdasarkan hasil ini, untuk mendapatkan keragaman spesies cendawan
entomopatogen lebih mudah dilakukan dengan cara mengisolasi dari serangga inang yang terinfeksi di alam dibandingkan dengan mengisolasi dari sumber lainnya (tanah dan pasir). Diperkirakan spesies cendawan yang ada pada inang yang berasal dari hama tanaman umumnya bersifat sebagai cendawan entomopatogen. Menurut MacLeod dan Muller-Kogler 1973 dalam Butt et al. (2001), serangga inang dari cendawan entomopatogen dijumpai pada lebih dari 32 famili yang tersebar pada ordo Hemiptera, Homoptera, Diptera, Lepidoptera, Coleoptera, Orthoptera, dan Hymenoptera. Beberapa spesies mempunyai rentangan inang yang luas, dan yang lainnya terbatas pada satu spesies atau terbatas pada kelompok spesies tertentu saja.
Gambar 4.1. Persentase isolat cendawan entomopatogen yang berhasil diisolasi dari berbagai sumber inokulum di alam.
Cendawan entomopatogen dari sumber inokulum tanah dan pasir mempunyai keragaman spesies yang lebih rendah. Hal ini dimungkinkan karena cendawan entomopatogen selain hidup sebagai patogen obligat juga banyak yang hidup sebagai patogen fakultatif yaitu disamping dapat meneruskan siklus hidupnya sebagai patogen pada serangga inang juga mampu bertahan hidup sebagai sapropit pada berbagai media di alam. Tanada dan Kaya (1993) menyatakan bahwa beberapa cendawan entomopatogen hidup sebagai patogen obligat yang siklus hidupnya selalu pada serangga inang namun kebanyakan cendawan entomopatogen juga sebagai patogen fakultatif dan mampu hidup tanpa inang dan melanjutkan siklus hidupnya sebagai sapropit
Menurut Keller dan Zimmermann (1989) cendawan entomopatogen juga dapat dikoleksi dari tanah dengan menggunakan serangga umpan. Metode ini terbukti sangat bermanfaat untuk mendeteksi cendawan entomopatogen
Serangga Hama 81% Tanah 13% Pasir 6%
khususnya untuk mempelajari sejarah penyebarannya. Walaupun tanah merupakan habitat yang sangat menguntungkan untuk interaksi serangga dan cendawan entomopatogen, hanya sedikit spesies cendawan entomopatogen yang ditemukan pada tanah. Beberapa spesies cendawan penting yang ditemukan pada tanah tercatat sebagai genera: Conidiobolus, Beauveria, Metarhizium dan
Paecilomyces.
Lebih lanjut dinyatakan keberhasilan cendawan entomopatogen di dalam tanah telah berkembang dengan melakukan adaptasi khusus, dimana siklus kehidupan cendawan entomopatogen terdiri dari fase parasit dan sapropit. Bila serangga terinfeksi cendawan mati dan secara umum siklus hidup cendawan akan berahir bila fase vegetatif atau organ reproduksi seksual terbentuk di luar inang. Namun fase kehidupan selanjutnya sebagai sapropit akan mengkolonisasi inang mati dan untuk sementara waktu berhenti pada pembentukan pseudosclerotium; tingkatan dormansi yang memungkinkan cendawan bertahan pada kondisi yang tidak menguntungkan. Jika faktor eksternal telah menguntungkan, hifa akan muncul lewat kutikula serangga.
Beberapa spesies cendawan seperti Paecilomyces spp., B. bassiana, B.
brongniartii atau Cordyceps spp. memproduksi untaian hifa, synnemata atau stromata di luar serangga inang. Elemen-elemen hifa ini sebagai adaptasi
morfologi terhadap kondisi tanah yang memungkinkannya menginfeksi inang baru.
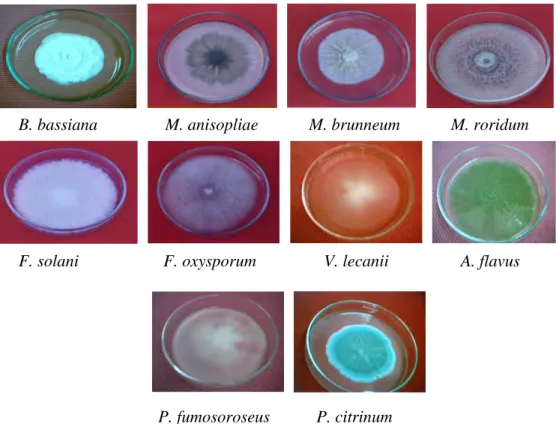
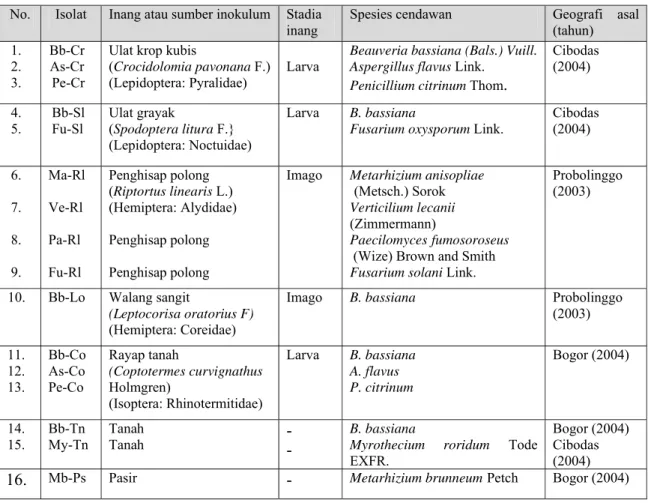
Hasil isolasi cendawan entomopatogen dari masing-masing inang atau sumber inokulum di alam dikoleksi untuk selanjutnya diidentifikasi berdasarkan warna, pertumbuhan secara in vivo dan in vitro serta bentuk organ seksual masing-masing spesies. Berdasarkan hasil identifikasi pada penelitian ini ditemukan 16 isolat (10 spesies) (Gambar 4.2 dan Table 4.1).
Keragaman antar spesies cendawan entomopatogen sangat luas sekali. Cendawan terdiri dari dua Divisi yaitu Myxomycota dan Eumycota. Cendawan entomopatogen ditemukan pada Divisi Eumycota yang terdiri dari sub-divisi: Mastigomycotina, Zygomycotina, Ascomycotina, Basidiomycotina dan Deuteromycotina. Umumnya mikroorganisme ini ada pada semua habitat tingkat perkembangan serangga (Ainsworth 1973 dalam Butt et al. 2001). Boucias dan
Pendland (1998) menyatakan beberapa genus cendawan entomopatogen yang tergolong ke dalam divisi Eumycota, sub-divisi Deuteromycota, klas Hyphomycetes dan ordo Moniliales adalah: Aspergilus, Beauveria, Fusarium,
Metarhizium, Paecilomyces dan Verticilium.
B. bassiana M. anisopliae M. brunneum M. roridum
F. solani F. oxysporum V. lecanii A. flavus
P. fumosoroseus P. citrinum
Gambar 4.2. Koloni cendawan entomopatogen dari konidia tunggal ber-umur 3 minggu pada media SDAY kecuali F. oxysporum (13 hari)
Spesies B. bassiana adalah spesies cendawan entomopatogen yang paling dominan ditemukan. Menurut Scholte et al. (2004), cendawan entomopatogen
B. bassiana mempunyai sebaran inang yang luas dan mudah diperbanyak secara in vitro pada media sintetik, penyebarannya hampir di semua lokasi di dunia dan
mempunyai spesies serangga inang lebih banyak dibandingkan spesies cendawan entomopatogen lainnya; kebanyakan inangnya berasal dari ordo Lepidoptera, Coleoptera, Hemiptera, Diptera dan Hymenoptera.
Spesies B. bassiana menunjukkan pertumbuhan yang tidak cepat tetapi dengan mudah tumbuh pada media SDAY, permukaan biakan rata seperti tepung
dan bewarna putih, cendawan ini menginfeksi rayap dengan penyebaran yang tidak teratur, kemudian mengalami penyempurnaan yang tidak menyebar rata. Setelah kematian inang konidiofora tumbuh keluar dari tubuh rayap dan diiringi dengan munculnya konidia.
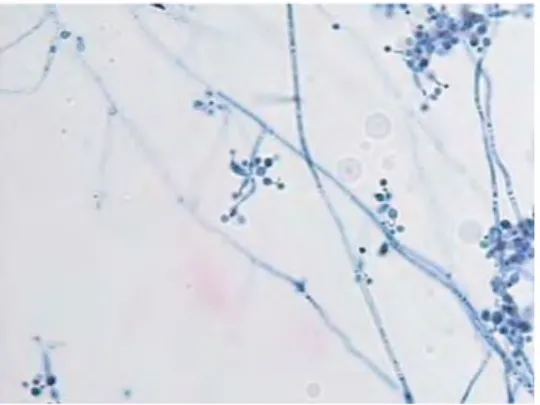
Barron (1968) menjelaskan tentang deskripsi dari B. bassiana sebagai berikut: konidiofora tidak begitu jelas; sel sporogenous sympodulae, muncul langsung dari hipa vegetatif, sederhana, hyaline - sub hyaline, secara berulang menghasilkan kuntum konidia berkelompok atau zigzag, terdiri dari satu sel basal yang sering mengembung, dan menghasilkan sympodulospores pada suksessi
acropetal; muncul spora pada ujung, seperti rachis, kadang-kadang berliku; spora
sangat kecil, bundar atau oval, hyaline - sub hyaline (Gambar 4.3)
Tabel 4.1 Cendawan entomopatogen yang ditemukan pada berbagai inang atau sumber inokulum di alam
No. Isolat Inang atau sumber inokulum Stadia
inang Spesies cendawan Geografi asal (tahun) 1. 2. 3. Bb-Cr As-Cr Pe-Cr
Ulat krop kubis
(Crocidolomia pavonana F.) (Lepidoptera: Pyralidae) Larva
Beauveria bassiana (Bals.) Vuill. Aspergillus flavus Link.
Penicillium citrinum Thom.
Cibodas (2004) 4. 5. Bb-Sl Fu-Sl Ulat grayak (Spodoptera litura F.} (Lepidoptera: Noctuidae) Larva B. bassiana
Fusarium oxysporum Link.
Cibodas (2004) 6. 7. 8. 9. Ma-Rl Ve-Rl Pa-Rl Fu-Rl Penghisap polong (Riptortus linearis L.) (Hemiptera: Alydidae) Penghisap polong Penghisap polong
Imago Metarhizium anisopliae
(Metsch.) Sorok
Verticilium lecanii
(Zimmermann)
Paecilomycesfumosoroseus (Wize) Brown and Smith Fusarium solani Link.
Probolinggo (2003)
10. Bb-Lo Walang sangit
(Leptocorisa oratorius F)
(Hemiptera: Coreidae)
Imago B. bassiana Probolinggo
(2003) 11. 12. 13. Bb-Co As-Co Pe-Co Rayap tanah (Coptotermes curvignathus Holmgren) (Isoptera: Rhinotermitidae) Larva B. bassiana A. flavus P. citrinum Bogor (2004) 14. 15. Bb-Tn My-Tn Tanah
Tanah - - B. bassiana Myrothecium roridum Tode
EXFR.
Bogor (2004) Cibodas (2004)
Isolat lainnya yang diisolasi dari inang atau sumber inokulum di alam seperti terlihat pada Tabel 4.1. juga diidentifikasi berdasarkan bentuk organ seksual, warna massa konidia secara in vitro dan in vivo, dan tipe pertumbuhannya (Deskripsi selengkapnya pada Lampiran 1).
Gambar 4.3. Cendawan B. bassiana pada media SDAY dengan perbesaran 1000 x
Patogenisitas dan Virulensi
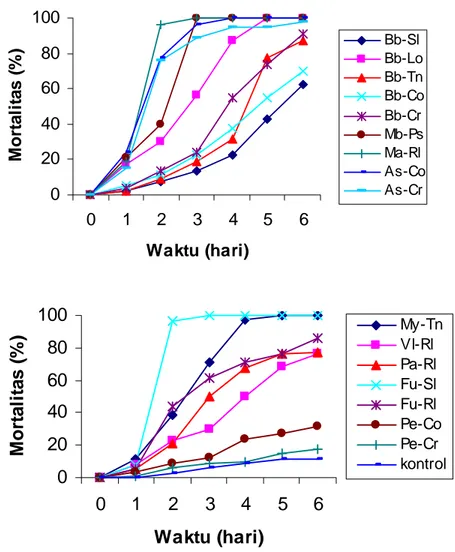
Tingkat patogenisitas antar spesies cendawan entomopatogen dan tingkat virulensi antar isolat di dalam satu spesies cendawan yang ditemukan pada penelitian ini umumnya berbeda nyata pada uji DUNCAN pada tingkat keragaman 5%. Spesies cendawan M. anisopliae dari inang penghisap polong,
M. brunneum dari pasir, M. roridum dari tanah, B. bassiana dari inang walang
sangit, F. oxysporum dari ulat grayak, dan A. flavus dari rayap tanah dapat menyebabkan mortalitas 100% setelah 6 hari inokulasi (Gambar 4.4). Hal ini berarti beberapa isolat cendawan entomopatogen bersifat sangat patogen dan berpotensi dijadikan sebagai agens hayati untuk pengendalian rayap tanah
C. gestroi.
Perbedaan tingkat patogenisitas antar spesies dan perbedaan virulensi antar isolat cendawan entomopatogen diperkirakan disebabkan oleh perbedaan sifat dasar internal (genetik) antar spesies dan perbedaan sumber inang asal isolat. Selain hal ini juga disebabkan oleh pengaruh lingkungan sebagai faktor eksternal yang dapat berpengaruh terhadap kemampuan cendawan tumbuh dan berkembang serta melumpuhkan mekanisme pertahanan serangga inang. Menurut
0 20 40 60 80 100 0 1 2 3 4 5 6 Waktu (hari) M o rt a lit a s ( % ) Bb-Sl Bb-Lo Bb-Tn Bb-Co Bb-Cr Mb-Ps Ma-Rl As-Co As-Cr
0
20
40
60
80
100
0
1
2
3
4
5
6
Waktu (hari)
M
o
rt
a
lit
a
s
(
%
)
My-TnVl-Rl Pa-Rl Fu-Sl Fu-Rl Pe-Co Pe-Cr kontrolTanada dan Kaya (1993) biasanya cendawan menyebabkan mortalitas dengan satu atau lebih cara seperti: defisiensi nutrisi, menyerang dan merusak jaringan, dan melepaskan toksin. Beberapa di antaranya bersifat virulen dan membunuh serangga dalam waktu yang singkat dan yang lainnya menghasilkan infeksi kronik yang lama.
Gambar 4.4. Laju mortalitas rayap C. gestroi oleh cendawan entomopatogen (107 konidia/ml) dari berbagai inang 6 hari setelah inokulasi (kontrol 11,25%).
Keterangan:
Bb-Sl = B. bassiana dari ulat grayak, Bb-Lo = B. bassiana dari walang sangit, Bb-Tn = B. .bassiana dari tanah, Bb-Co =
B. bassiana dari rayap tanah, Bb-Cr = B. bassiana dari ulat krop kubis, Mb-Ps = M. brunneum dari pasir, Ma-Rl = M. anisopliae
dari penghisap polong, As-Co = A. flavus dari rayap tanah, As-Cr = A..flavus dari ulat krop kubis, My-Tn = M. roridum dari tanah, Vl-Rl = V. lecanii dari penghisap polong, Pa-Rl = P. fumosoroseus dari penghisap polong, Fu-Sl = F. oxysporum dari ulat grayak, Fu-Rl = Fusarium solan dari penghisap polong, Pe-Co = P. citrinum dari rayap tanah, Pe-Cr = P. citrinum dari ulat krop kubis,
Isolat cendawan entomopatogen yang ditemukan dari berbagai inang umumnya bersifat patogen terhadap rayap, dan dapat menyebabkan mortalitas lebih dari 60%. Yoshimura dan Takahashi (1998), menjelaskan bahwa penggunaan cendawan entomopatogen sebagai pengendalian rayap telah 25 tahun lebih menjadi target penelitian dan banyak spesies cendawan telah diuji tingkat patogenisitasnya terhadap rayap. Setelah tahun 1960-an banyak peneliti memulai investigasi patogenisitas cendawan terhadap rayap menggunakan A. flavus,
Absidia coerulea Bainier, B. bassiana, Entomophthora sp. M. anisopliae, Conidiobolus coronatus dan Penicillium sp.
Setiap spesies cendawan entomopatogen mempunyai sifat dan kemampuan spesifik untuk tumbuh dan berkembang pada rayap. Perbedaan ini juga terlihat pada karakterisasi fisiologisnya secara in vivo dan in vitro terutama pada kemampuan berkecambah, laju pertumbuhan koloni dan konidiogenesis (Tabel 4.2). Diduga setiap spesies juga menghasilkan jenis metabolit sekunder (toxin) yang bervariasi sehingga mempunyai daya toksisitas yang berbeda satu sama lainnya.
Boucias dan Pendland (1998) mengemukakan, cendawan entomopatogen dicirikan oleh kemampuannya untuk menempel dan menembus kutikula inang dan dapat tumbuh ke bahagian internal inang (hemocoel) dan mengkonsumsinya sehingga nutrient di dalam hemolymph habis oleh pertumbuhan cendawan yang begitu cepat. Di samping itu cendawan dapat menghancurkan jaringan lainnya atau dengan melepaskan zat beracun yang mengganggu perkembangan inang secara normal. Beberapa di antara zat beracun (toxin) yang dihasilkan cendawan yang dapat membunuh serangga adalah: 1) Beauvericin oleh B. bassiana,
Paecilomyces dan Fusarium, 2) Bassianolide oleh B. bassiana, 3) Cyclosporin A
oleh B. bassiana, Verticillium, Fusarium dan Tolypocladium, 4) Oosporein oleh
B. bassiana, 5) asam Oxalic oleh B. brongniartii, 6) Destruxins oleh M. anisopliae dan Aspergillus ochraceus, 7) Cytochalasins oleh M. anisopliae, 8)
Swainsonine oleh M. anisopliae, 9) Aflatoxins oleh Aspergillus, 10) Asam kojic oleh A. flavus dan 11) Restrictocin oleh Aspergillus fumigatus.
Selanjutnya dinyatakan bahwa penempelan mungkin melibatkan kekuatan
N-acetylglucosamine merupakan substansi yang ditemukan pada permukaan spora, spora berkecambah dengan cepat tergantung pada kelembaban lingkungan, temperatur, kondisi cahaya yang kurang serta nutrisi lingkungan. Penetrasi terutama tergantung pada sifat kutikula serangga (ketebalan, sclerotization, dan kehadiran zat anti cendawan) dan substansi nutrisi. Setelah berkecambah hifa masuk ke dalam integumen serangga dan terus ke hemocoel, dan menghasilkan tubuh hifa (hyphal bodies), yang pada hakekatnya berupa blastospores yang berkembang dengan tunas (budding).
Menurut Scholte et al. (2004), siklus cendawan entomopatogen hingga menyebabkan inangnya mati adalah sebagai berikut: konidia menempel pada kutikula kemudian berkecambah dan menembus kutikula. Kemudian di dalam hemocoel miselia tumbuh terus menerus pada inang membentuk tubuh hifa (blastospores). Kematian serangga sering disebabkan oleh kombinasi dari aksi toksin, terhalangnya sirkulasi darah, komsumsi nutrisi atau penyerangan organ dari serangga.
Virulensi antar isolat juga menunjukkan perbedaan, seperti pada 5 isolat cendawan B. bassiana yang berasal dari sumber inang yang berbeda, isolat walang sangit (Bb-Lo) paling virulen dibanding isolat B. bassiana yang lainnya. Dua isolat cendawan A. flavus yang juga berasal dari inang berbeda, isolat rayap tanah (As-Co) lebih virulen dibanding isolat ulat krob kubis (As-Cr). Diperkirakan sifat virulensi yang berbeda ini dipengaruhi oleh kondisi morfologi dan fisiologi serangga inang asal isolat dan lingkungan asal isolat yang berbeda.
Menurut Tanada dan Kaya (1993), virulensi adalah kemampuan penyakit yang dihasilkan oleh suatu mikroorganisme, dalam hal ini adalah kemampuan suatu organisme untuk menyerang dan menyebabkan luka pada inang, yang berhubungan dengan kesanggupan suatu mikroorganisme untuk mengatasi mekanisme pertahanan inang. Suatu patogen mungkin bersifat sangat virulen sebab rendahnya ketahanan atau tingginya kerentanan dari inang, dan sebaliknya patogen dapat mempunyai virulensi yang rendah sebab tingginya ketahanan atau rendahnya kerentanan dari inang.
Patogenisitas secara dekat merupakan sinonim terhadap virulensi yaitu berkenaan dengan kemampuan menghasilkan penyakit oleh mikroorganisme.
Perbedaannya adalah bahwa patogenisitas diaplikasikan terhadap kelompok atau spesies dari organisme; sedangkan, virulensi digunakan pengertian tingkat dari patogenisitas di dalam kelompok atau spesies. Patogenisitas kadang-kadang dipandang sebagai kemampuan penetapan secara genetika untuk menghasilkan penyakit, dan virulensi tidak sebagai hasil secara genetika. Dengan demikian, kita dapat mengatakan bahwa patogenisitas B. bassiana adalah tinggi untuk isolat
L. oratorius (Bb-Lo), tetapi virulensinya berbeda tergantung pada kondisi, seperti
metode pembiakan, penyimpanan, formulasi, dan faktor lingkungan. Sebagai contoh di bawah kondisi nutrisi tertentu, virulensi dari patogen lebih tinggi dibanding pada nutrisi lainnya. B. bassiana mempunyai banyak strains, yang pada suatu waktu dipertimbangkan untuk berbeda spesies sebab berbeda karakteristik morfologinya. Strains akan bervariasi virulensinya dan tergantung pada kerentanan spesies serangga sasaran.
Perbedaan mortalitas rayap sebagai akibat perbedaan tingkat virulensi disebabkan oleh asal dan kondisi sumber inang yang berbeda pula, hal ini menyebabkan perbedaannya dalam karakter menyerang rayap. Disamping juga tergantung pada lingkungan pada daerah asal, karakter dan struktur dari konidia juga berakibat terhadap perkecambahan. Perkecambahan yang sukses dan berpenetrasi pada inang tergantung pada total persentase perkecambahan, lamanya waktu berkecambah, cara dari perkecambahan, agresivitas cendawan, dan kerentanan inang (Samson et al. 1988 dalam Tanada dan Kaya 1993).
Spesies cendawan mempunyai banyak strains yang berbeda virulensinya. Pada umumnya, strains dari spesies yang diisolasi dari inang yang spesifik lebih virulen untuk inang yang sama dibandingkan isolat dari inang yang lainnya, dan suksesi penularan di dalam suatu inang dapat juga menghasilkan peningkatan virulensi atau menghasilkan strain yang lebih virulent. Pernyataan ini dipertegas oleh hasil penelitian ini yaitu pada kasus perbedaan virulensi antar isolat
A. flavus, yang berasal dari serangga inang rayap tanah (As-Co) lebih virulen
dibandingkan yang berasal dari serangga inang ulat krop kubis (As-Cr) terhadap mortalitas rayap C. gestroi. Namun tidak demikian pada kasus perbedaan virulensi antar isolat B. bassiana, isolat walang sangit (Bb-Lo) menunjukkan
tingkat virulensi yang lebih tinggi dibanding isolat dari rayap tanah (Bb-Co), ulat krop kubis (Bb-Cr), ulat grayak (Bb-Sl) dan tanah (Bb-Tn).
Pada kasus seperti diterangkan di atas, tingkat virulensi ternyata tidak hanya dipengaruhi oleh asal isolat yang secara umum ditentukan oleh faktor eksternal (lingkungan), akan tetapi juga oleh spesies cendawan yang secara umum lebih dipengaruhi oleh sifat internal, dengan pengertian bahwa setiap spesies cendawan entomopatogen masing-masingnya mempunyai spesies inang yang spesifik. Pernyataan ini sejalan dengan penelitian Trizelia (2005), bahwa pada spesies cendawan B. bassiana, isolat yang virulen terhadap serangga hama tidak selalu berasal dari hama yang sama. Hasil uji virulensi dari 13 isolat B. bassiana menunjukkan bahwa isolat B. bassiana yang sangat virulen terhadap ulat krop kubis C. pavonana dari Cibodas berasal dari serangga dan daerah lain yang bukan serangga inang uji yaitu berasal dari inang L. oratorius dari Cianjur.
Mortalitas rayap C. gestroi yang disebabkan cendawan entomopatogen
M. anisopliae dan F. oxysporum lebih cepat dibandingkan yang disebabkan oleh
spesies lainnya yaitu dapat menyebabkan mortalitas rayap 100% dalam periode waktu yang lebih singkat. Hal ini diperkirakan spesies cendawan ini disamping kemampuannya mendegradasi inang, juga dengan sangat cepat menyebarkan metabolit sekundernya yang bersifat racun bagi rayap sehingga terjadi ketidak seimbangan fungsi organ tubuh. Agens hayati penyebab penyakit ”green
muscardine” M. anisopliae merupakan spesies patogen yang secara alami
menginfeksi lebih dari 200 jenis serangga, termasuk rayap (Tanada dan Kaya 1993 dalam Strack 2003).
Ferron 1981 di dalam Scholte et al. (2004), menyatakan bahwa di dalam studi histopatologi pada jaringan elaterid yang diinfeksi M. anisopliae memperlihatkan bahwa toksin (destruxin) membunuh serangga inang dengan merangsang atau memacu terjadinya kemerosotan jaringan serangga inang sehingga kehilangan keutuhan struktural membran dan kemudian terjadi dehidrasi sel. Dalam hal ini penyumbatan spirakel dimungkinkan terjadi sehingga dapat menyebabkan kematian sebelum serangan pada hemocoel.
Menurut MacLeod 1963 dalam Tanada dan Kaya (1993), periode dari infeksi sampai pada kematian serangga dapat dalam waktu yang singkat 3 hari
dan selama-lamanya 12 hari, dan kebanyakan terjadi dalam rentangan 5 - 8 hari. Periode dapat bervariasi dan juga tergantung pada ukuran dari inang. Kebanyakan serangga mati setelah sore hari antara 1500 dan 1900 jam. Virulensi dan patogenisitas dari cendawan entomopatogen dapat berasosiasi dengan produksi enzim collagenolytic dan mycotoxins.
Spesies cendawan M. anisopliae telah sangat populer keefektifannya di dalam pengendalian rayap, bahkan sampai saat sekarang telah banyak biopestisida yang berbasiskan konidia dari cendawan M. anisopliae diedarkan secara komersil. Namun spesies cendawan Fusarium spp. masih diragukan dalam penggunaan sebagai biopestisida walaupun mempunyai daya tinggi dalam membunuh serangga hama, hal ini sehubungan dengan sifatnya yang juga sebagai patogen pada tanaman.
Teetor-Barsch dan Roberts (1993) menyatakan cendawan Fusarium diketahui kelimpahannya di alam dan juga keragaman tempat ia berasosiasi di antaranya pada tanaman hidup dan yang telah mati serta pada banyak hewan. Cendawan ini terutama ditemukan berasosiasi dengan serangga. Perhatian khusus diberikan terhadap rentangan inang, teristimewa antara inang tanaman dan serangga, dan memungkinkan potensi cendawan ini untuk mengendalikan hama. Beberapa jenis Fusarium spp. yang bersifat entomopatogen bersifat lemah dan sebagai patogen fakultatif khususnya pada ordo Lepidoptera dan Coleoptera. Cendawan akan mengkolonisasi inangnya yang mati sebagai sapropit. Pada segolongan kecil kasus tingkat patogenisitas terhadap tanaman dan serangga oleh satu isolat juga ditemukan. Tingkat potensi isolat Fusarium yang menyebabkan mortalitas tinggi terhadap serangga juga memperlihatkan spesifikasi inang yang tinggi dan tidak berbahaya terhadap jenis tanaman.
Periode waktu kematian rayap oleh cendawan entomopatogen secara umum tidak menunjukkan tanda dan gejala yang nyata pada awal tingkatan infeksi. Hanya setelah infeksi menyebar ke dalam tubuh, rayap menjadi kurang aktif atau menunjukkan kegelisahan. Pada tahapan akhir infeksi, rayap kehilangan tenaga gerak dan diam ditempat lalu kemudian mati, pada tahapan paling ahir dari proses infeksi, kadang-kadang rayap dapat berganti warna menjadi gelap dan tidak dikolonisasi oleh miselia seperti pada kasus rayap yang diinfeksi oleh cendawan
M. anisopliae. Kadang-kadang rayap yang telah mati menjadi keras dan 3 - 4 hari
setelahnya, rayap berubah warna sesuai warna konidia yang dihasilkan, rayap mati diselimuti warna konidia putih mengindikasikan terserang oleh B. bassiana. Sporulasi in Vivo
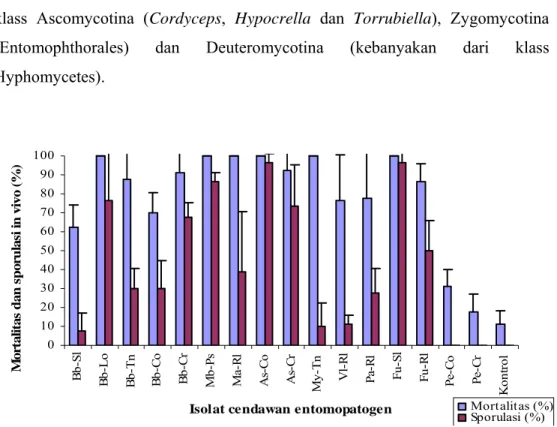
Pada penelitian ini, sporulasi cendawan pada permukaan tubuh rayap (in
vivo) tidak semua berkorelasi dengan mortalitas namun spesies cendawan yang
menunjukkan tingkat patogenisitas yang tinggi seperti A. flavus dari inang rayap tanah, F. oxysporum dari ulat grayak, M. brunneum dari pasir dan B. bassiana dari walang sangit juga mampu bersporulasi yang tinggi secara in vivo ( 78,25% – 97,25%) kecuali M. anisopliae dan M. roridum dari tanah. Dalam hal ini
M. roridum menunjukan kemampuan bersporulasi in vivo paling rendah
(Gambar 4.5). Diperkirakan hal ini juga dipengaruhi oleh spesies cendawan, sumber isolat dan faktor lingkungan; cendawan dapat tumbuh pada kondisi kelembaban yang tinggi namun tidak semua dapat bersporulasi dengan baik dan terlihat dengan jelas.
Menurut Prior dan Perry 1980 di dalam Butt et al. (2001) dan Tanada dan Kaya (1993), pertama kali mikroorganisme ditemukan sebagai penyebab penyakit pada serangga adalah cendawan sebab pertumbuhannya secara makroskopis nampak dengan nyata pada permukaan serangga inang. Namun beberapa cendawan entomopatogen bentuk pertumbuhannya tipis atau jarang dan tidak nyata karena struktur eksternalnya yang sangat kecil dan sulit dideteksi oleh peneliti. Pertumbuhan dan perkembangan cendawan terutama dibatasi oleh kondisi lingkungan eksternal, kususnya kelembaban yang tinggi atau kelembaban dan temperatur yang memadai untuk bersporulasi dan perkecambahan spora.
Rombach (1988) menyatakan bahwa banyak spesies cendawan dapat ditemukan tumbuh pada serangga mati, namun kebanyakan merupakan cendawan saprofit yang menyerang setelah serangga mati. Akan tetapi hanya spesies entomopatogen yang dapat secara aktif menyerang serangga hidup, membunuh inang dan bersporulasi pada inang yang telah mati. Secara keseluruhan lebih dari 700 spesies cendawan dari perkiraan 90 genera adalah merupakan patogen terhadap serangga. Spesies cendawan entomopatogen tersebut ditemukan pada
0 10 20 30 40 50 60 70 80 90 100 Bb -S l Bb -L o Bb -T n Bb -C o Bb -C r Mb -P s Ma -R l As -C o As-C r My -T n Vl -R l Pa -R l Fu -S l Fu -R l Pe -C o Pe -C r Ko nt ro l
Isolat cendawan entomopatogen
M o rt alit as d a n s p or u las i in viv o ( % ) Mortalitas (%) Sporulasi (%)
klass Ascomycotina (Cordyceps, Hypocrella dan Torrubiella), Zygomycotina (Entomophthorales) dan Deuteromycotina (kebanyakan dari klass Hyphomycetes).
Gambar 4.5. Sporulasi cendawan entomopatogen pada tubuh rayap (in vivo) dan mortalitas rayap C. gestroi 6 hari setelah diinokulasi dengan suspensi cendawan entomopatogen (107 konidia/ml).
Di samping virulensi, kemampuan bersporulasi pada inang (in vivo) menjadi sangat penting bila untuk tujuan penularan di dalam koloni karena cendawan yang mampu bersporulasi pada inang dengan baik akan dapat membentuk infective propagul baru dan dapat tersebar luas ke seluruh individu di dalam koloni. Goettel dan Inglis 1997 dalam Scholte et al. (2004) menyatakan bahwa setelah inang mati, hifa biasanya muncul dari bangkai serangga dan di bawah kondisi abiotik yang menguntungkan, konidia dapat dihasilkan pada tubuh bahagian luar inang, ini kemudian akan tersebar oleh angin atau air.
Menurut Yoshimura et al. (1992), pertumbuhan hifa dari tubuh rayap setelah diamati sampai mati tidak nyata, namun dua sampai 3 hari setelah kematian banyak hifa yang tumbuh ke luar, kira-kira 2 - 10 hari setelah kematian
Keterangan:
Bb-Sl = B. bassiana dari ulat grayak, Bb-Lo = B. bassiana dari walang sangit, Bb-Tn = B. .bassiana dari tanah, Bb-Co =
B. bassiana dari rayap tanah, Bb-Cr = B. bassiana dari ulat krop kubis, Mb-Ps = Metarhizium brunneum dari pasir, Ma-Rl = M. anisopliae dari penghisap polong, As-Co = A. flavus dari rayap tanah, As-Cr = A..flavus dari ulat krop kubis, My-Tn = Myrothecium roridum dari tanah, Vl-Rl = V. lecanii dari penghisap polong, Pa-Rl = P. fumosoroseus dari penghisap polong,
Fu-Sl = F. oxysporum dari ulat grayak, Fu-Rl = Fusarium solan dari penghisap polong, Pe-Co = P. citrinum dari rayap tanah, Pe-Cr = P. citrinum dari ulat krop kubis,
dibutuhkan untuk pembentukan konidia baru. Pada kasus cendawan C. coronatus, banyak konidia terlihat setelah 2 hari pada rayap yang telah di ekspose selama 6 jam.
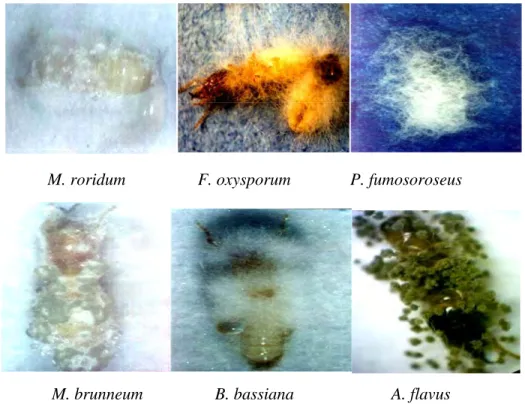
M. roridum F. oxysporum P. fumosoroseus
M. brunneum B. bassiana A. flavus
Gambar 4.6. Sporulasi in vivo beberapa spesies cendawan entomopatogen pada tubuh rayap 9 hari setelah diinokulasi
Karakterisasi Fisiologi Cendawan Entomopatogen Terseleksi pada Media SDAY (in vitro): Daya Kecambah, Diameter Koloni dan Sporulasi
Karakter fisiologi adalah kemampuan cendawan untuk tumbuh dan berkembang pada media sintetik atau inang, dalam hal ini yang diamati antara lain adalah: daya kecambah, diameter koloni dan kemampuan berseporulasi. Di samping patogenisitas, karakterisasi fisiologi cendawan secara in vitro akan menjadi penting bila cendawan tersebut diperbanyak secara massal untuk tujuan bio-termitisida komersil.
Umumnya cendawan entomopatogen yang mempunyai tingkat patogenisitas tinggi dapat direkomendasikan pada penelitian selanjutnya, walaupun dari hasil
analisis statistik ada perbedaan diameter koloni, daya berkecambah dan jumlah konidia dari masing-masing spesies atau isolat cendawan entomopatogen.
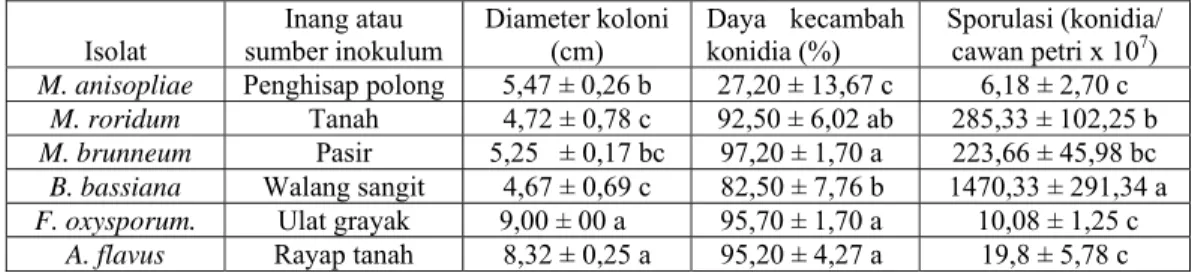
Tabel 4.2. Karakterisasi fisiologi beberapa spesies cendawan entomopatogen terseleksi: diameter koloni, daya kecambah konidia dan sporulasi.
Isolat sumber inokulum Inang atau Diameter koloni (cm) Daya kecambah konidia (%) Sporulasi (konidia/ cawan petri x 107)
M. anisopliae Penghisap polong 5,47 ± 0,26 b 27,20 ± 13,67 c 6,18 ± 2,70 c M. roridum Tanah 4,72 ± 0,78 c 92,50 ± 6,02 ab 285,33 ± 102,25 b M. brunneum Pasir 5,25 ± 0,17 bc 97,20 ± 1,70 a 223,66 ± 45,98 bc B. bassiana Walang sangit 4,67 ± 0,69 c 82,50 ± 7,76 b 1470,33 ± 291,34 a F. oxysporum. Ulat grayak 9,00 ± 00 a 95,70 ± 1,70 a 10,08 ± 1,25 c
A. flavus Rayap tanah 8,32 ± 0,25 a 95,20 ± 4,27 a 19,8 ± 5,78 c
Angka-angka yang diikuti oleh huruf yang sama pada baris dan kolom yang sama tidak berbeda nyata menurut uji DNMRT pada taraf nyata 5%.
Diameter koloni (cm)
Diameter koloni dari masing-masing spesies setelah dua minggu berkisar antara 4,4 cm – 9,0 cm, tertinggi diperoleh pada F. oxysporum (9 cm) dan kemudian diikuti oleh A. flavus (8,3 cm). Analisis statistik memperlihatkan kedua spesies cendawan ini tidak berbeda nyata, cendawan yang menunjukkan pertumbuhan koloni yang lebih cepat ini menunjukkan pertumbuhan yang lebih tipis dipermukaan media kultur SDAY. Hal ini ternyata berkaitan dengan jumlah konidia yang dihasilkan juga tidak berbeda nyata dan menghasilkan jumlah konidia lebih sedikit.
Olsson (1997) mendefinisikan koloni cendawan adalah suatu kelompok hifa yang berasal dari satu spora atau satu individu atau yang berasal dari satu sumber miselia. Berdasarkan pernyataan ini, pertumbuhan koloni diawali oleh perkecambahan dan pertumbuhan miselia, cepatnya pertumbuhan miselia akan menyebabkan nutrisi yang diperoleh dari media tumbuh akan dimanfaatkan terlebih dahulu untuk pertumbuhan miselia sehingga diperkirakan pembentukan konidia akan dapat terhambat.
Daya kecambah konidia (%)
Daya kecambah cendawan entomopatogen dari masing-masing spesies cendawan lebih dari 80% kecuali M. anisopliae (27,2%) setelah 12 - 24 jam
inkubasi. Hasil penelitian menunjukan secara umum semua isolat terseleksi tergolong mempunyai daya kecambah tinggi, hal ini akan sangat menguntungkan didalam keberhasilan cendawan menginfeksi inangnya, karena keberhasilan cendawan untuk dapat menyerang inang terutama sangat ditentukan oleh kemampuan cendawan menempel dan berkecambah pada kutikula serangga. Boucias dan Pendland (1998), mengemukakan cendawan entomopatogen dicirikan oleh kemampuannya untuk menempel dan menembus kutikula inang serta dapat tumbuh ke bahagian internal inang (hemocoel).
Menurut Leland 2001; Jenkins et al. 1998; Kassa 2003 dalam Trizelia (2005) evaluasi daya kecambah konidia cendawan entomopatogen perlu dilakukan terutama apabila cendawan tersebut akan dikembangkan sebagai bioinsektisida. Daya kecambah konidia merupakan salah satu kriteria dalam pemilihan isolat; cendawan yang memiliki daya kecambah konidia di atas 80% telah memenuhi syarat untuk dikembangkan sebagai bioinsektisida. Selanjutnya dinyatakan bahwa dalam pemilihan isolat yang akan digunakan, kecepatan perkecambahan juga harus diperhitungkan. Isolat yang berkecambah lebih cepat lebih berpotensi untuk menimbulkan infeksi, karena isolat ini dapat terhindar dari pengaruh kekeringan, pengaruh dari mikroorganisme lain dan terlepas dari kutikula serangga pada waktu ekdisis.
Kemampuan konidia untuk berkecambah merupakan suatu faktor penting untuk berhasilnya melakukan penetrasi pada inang, namun hal ini akan sangat tergantung pada faktor lingkungan asal spesies atau isolat dan sifat genetik masing-masing spesies cendawan. Hal ini biasanya ditunjukkan oleh perbedaan viabilitas antar isolat di dalam spesies yang sama, pada penelitian ini terlihat bahwa daya kecambah M. anisopliae berbeda sangat nyata dengan M. brunneum (Tabel 4.2). Menurut Ekesi et al. (2003) viabilitas 4 isolat M. anisopliae yang berasal dari Kenya dan Congo berkisar antara 85 – 90%.
Viabilitas antar spesies cendawan M. brunneum, F. oxysporum dan A. flavus tidak berbeda nyata. Hal ini ditunjukkan dengan kemampuan tingkat patogenisitas yang tidak berbeda (Gambar 4.4). Adanya keterkaitan antara tingkat patogenisitas dengan kemampuan berkecambah merupakan salah satu hal yang sangat menunjang di dalam adanya kemungkinan spesies cendawan ini untuk
dikembangkan ke penelitian yang lebih mendalam sehingga berpotensi sebagai biopestisida.
Sporulasi (konidia / cawan petri).
Sporulasi adalah kemampuan cendawan entomopatogen untuk menghasilkan konidia pada media sintetik atau inang. Untuk dapat melakukan proses infeksi dan menurunkan generasi berikutnya dengan cepat akan sangat tergantung pada kemampuan cendawan menghasilkan konidia. Pada penelitian ini setelah 2 minggu, B. bassiana menghasilkan jumlah konidia tertinggi (1,47 x 1010 konidia / cawan petri). Hasil ini berbeda nyata dengan jumlah konidia yang dihasilkan oleh spesies cendawan lainnya. Kemudian diikuti oleh M. brunneum,
A. flavus, F. oxysporum dan M. anisopliae (Tabel 4.2).
Spesies cendawan B. bassiana, menunjukkan laju pertumbuhan koloni yang lambat namun memperlihatkan pertumbuhan koloni yang lebih tebal, diperkirakan hal ini salah satu faktor penentu spesies cendawan ini dapat menghasilkan jumlah konidia lebih banyak dibandingkan spesies cendawan yang diujikan, yang bearti ada korelasi dengan karakter fenotip cendawan disamping juga ditentukan oleh faktor genetik. Hasil penelitian ini menunjang penelitian yang dilakukan oleh Trizelia (2005), 13 isolat B. bassiana dapat menghasilkan jumlah konidia antara 1,27 X 1010 – 1,79 X 1010 konidia / cawan petri. Hasil ini
juga memperlihatkan perbedaan nyata antar isolat dan tertinggi ditemukan pada isolat yang berasal dari inang C. pavonana (1,79 X 1010 konidia/cawan petri)
Kemampuan cendawan menghasilkan jumlah konidia yang banyak, merupakan salah satu faktor yang menguntungkan di dalam pemanfaatannya sebagai agens pengendalian hayati, karena konidia sangat penting untuk infeksi dan penyebaran cendawan. Sun et al. 2003 dalam Trizelia (2005) menyatakan bahwa Isolat B. bassiana yang lebih cepat bersporulasi dengan total sporulasi yang tinggi menghasilkan epizootik yang lebih baik dalam koloni rayap C.
formosanus.
Karakter fisiologis (diameter koloni, viabilitas dan kemampuan bersporulasi) cendawan entomopatogen menunjukan keragaman antar spesies yang diuji. Selain faktor-faktor penentu yang telah diuraikan di atas, faktor media
tumbuh diperkirakan juga menjadi salah satu faktor penentu pertumbuhan secara fisiologis. Pada penelitian ini media yang digunakan adalah SDAY dengan komposisi dekstrosa 10 g, pepton 2,5 g, ekstrak khamir 2,5 g, agar 20 g dalam akuades 1 liter yang mengandung 250 ppm chloromphenicol (Samuels et al. 2002). Dalam hal ini masing-masing spesies cendawan entomopatogen memungkinkan dapat tumbuh lebih baik pada media yang lebih spesifik.
Kesimpulan
Hasil isolasi menunjukkan keragaman spesies yang didapatkan dari sumber inokulum di alam. Tertinggi ditemukan pada inang yang berasal dari serangga terinfeksi dibanding dari sumber tanah dan pasir. Spesies cendawan entomopatogen yang ditemukan pada berbagai inang atau sumber inokulum di alam adalah B. bassiana, M. anisopliae, M. brunneum, P. fumosoroseus,
P. citrinum, M. roridum, V. lecanii, F. oxysporum, F. solanii dan A. flavus. Hasil
uji patogenisitas menunjukkan pada umumnya cendawan bersifat patogen terhadap rayap C. gestroi.
Cendawan entomopatogen yang ditemukan umumnya dapat dimanfaatkan sebagai agens pengendalian hayati rayap C. gestroi, khususnya: M. anisopliae dari inang penghisap polong, M. brunneum dari pasir, M. roridum dari tanah,
B. bassiana dari walang sangit, F. oxysporum dari ulat grayak dan A. flavus dari
rayap tanah; pada penggunaan kerapatan konidia cendawan 107 konidia/ml dapat membunuh rayap C. gestroi 100% setelah 6 hari inokulasi.
Kemampuan bersporulasi in vivo antar spesies umumnya tidak berkorelasi dengan mortalitas, namun spesies cendawan yang menunjukkan tingkat patogenisitas yang tinggi seperti A. flavus dari inang rayap tanah, F. oxysporum dari ulat grayak, M. brunneum dari pasir dan B. bassiana dari walang sangit juga menghasilkan kemampuan bersporulasi secara in vivo tinggi (78,25% – 97,25%).
M. roridum mempunyai kemampuan bersporulasi secara in vivo paling rendah
(10%).
Cendawan entomopatogen terseleksi jika ditumbuhkan pada media SDAY memperlihatkan daya kecambah konidia lebih dari 80% kecuali M. anisopliae.
Diameter koloni tertinggi setelah 2 minggu dicapai oleh F. oxysporum. (9 cm) dan sporulasi tertinggi dicapai oleh B. bassiana (1,47 X 1010 konidia/ml).
Berdasarkan hasil uji tapis tingkat patogenisitas (laju mortalitas), sporulasi
in vivo dan karakterisasi fisiologis, cendawan entomopatogen A. flavus dari
inang rayap tanah, F. oxysporum dari ulat grayak, M. brunneum dari pasir,
B. bassiana dari walang sangit dan M. anisopliae dari penghisap polong kedele