*)korespondensi : yati@unidar.ac.id
ANATOMICAL RESPONSES HOTONG (Setaria italiaca L. Beauv)
TO DROUGHT STRESS
Yati Tuasamu
*)Staf Pengajar pada Fakultas Keguruan dan Ilmu Pendidikan Universitas Darussalam Ambon
Diterima 09-03-2013, diterbitkan 01-05-2013
ABSTRACT
Hotong (Setaria italica L. Beauv) is one of local excellent crop in Buru Island which becomes an important food alternative. However, because this plants are normally cultivated during dry season, droughtstressis one of the major factor limiting plant growth and production. The objective of the study was to examine the adverse effects of drought stress on 3 accessions of hotong (Buru 1 accession (A1), Buru 2 accession (A2), and AGH accession (A3) based on anatomical responses. Plants were grown in 6 kg pot containing soil and sand (1:1, v/v) in the greenhouse. Drought was imposed at 14 days after the plants were 6 weeks old. The anatomy parameters were anatomy characteristics of leaf, stem and root were observed using paradermal and transversal section. The results showed that leaf anatomical characteristics of the plants subjected to drought stress decreased significantly in primary vena, vein rib, leaf, and mesophyll thickness, but the lower reduction was showed by A3 compared to other accessions especially in leaf and mesophyll thickness. The A1 showed the higher decrease in xilem vessels diameter of leaf, as compared to A2 and A3. The xilem vessels diameter of stems and root of all plants were relatively stable and equal to the plant control. Stomata density decreased significant in A2, but A1 and A3 it was stable. The results showed the A3 more tolerant than A1 and A2, indicated by stable of anatomical characteristics especially in in the leaf and mesophyll thickness that suported of photosynthetic rate during the drought stress.
Keyword: drought stress, anatomical responses, Setaria italica L. Beauv Pendahuluan
Pulau Buru memiliki beberapa komoditi tanaman pangan unggul lokal, salah satunya adalah Hotong (Setaria italica L. Beauv). Biji tanaman ini dikonsumsi oleh masyarakat Pulau Buru sebagai pangan alternatif penghasil karbohidrat, karena selain mengadung karbohidrat tinggi,bijinya juga memiliki kandungan nutrisi yang cukup tinggi yaitu mengandung protein sebesar 11,18%, lemak 2,36%, karbohidrat 73,36%, air sebesar 11,78%, dan abu sebesar 1,32%. Energi yang dihasilkan dalam 100 g biji adalah 359 kalori. Dilihat dari kandungan gizinya yang cukup tinggi, maka tanaman hotong dapat dijadikan komoditi alternatif dalam program diversifikasi pangan penghasil karbohidrat. Tanaman ini juga mempunyainilai ekonomis penting antara lain dapat dijadikan sebagai bahan baku pembuatan mi instan, minuman berenergi, dan makanan balita. (Andrawulan, 2003). Untuk itu perlu dibudidayakan secara luas serta di jaga kelestariannya sebagai komoditi unggul lokal.
Pemerintah Kabupaten Buru telah melakukan upaya budidaya tanaman hotong yaitu pada lahan-lahan kering dengan pola penanaman tadah hujan terutama di bagian barat dan utara Pulau Buru. Upaya pengembangan budidaya tanaman hotong di Pulau Buru hingga saat ini masih menemukan banyak kendala. Kondisi kekeringan dan ketersediaan benih unggul yang toleran kekeringan sejauh ini merupakan kendala utama. Data dan informasi hasil penelitian tentang pemuliaan dan pengembangan budidaya tanaman hotong masih sangat minim (Diptan, 2003).
Pulau Buru termasuk daerah rawan kekeringan dengan jumlah curah hujan tahunan relatif rendah yaitu berkisar antara 1000-1400 mm/thn. Curah hujan rata-rata bagi pertumbuhan tanaman hotong berkisar antara 200 - 300 mm/bln atau lebih sedikit dari curah hujan musiman (Dekany, 1999). Ketersediaan air yang rendah akibat rendahnya curah hujan di daerah tersebut terutama pada masa awal pertumbuhan tanaman dapat mengakibatkan cekaman kekeringan (drought stress).
508
Kekeringan merupakan faktor lingkungan utama yang dapat menghambat pertumbuhan tanaman dan menurunkan produksi bergantung pada besarnya tingkat cekaman yang dialami dan fase pertumbuhan tanaman ketika mendapat cekaman kekeringan (Levitt 1980). Pada periode cekaman kekeringan yang panjang akan mempengaruhi seluruh proses metabolisme di dalam sel dan mengakibatkan penurunan produksi tanaman (Bohnert, 1995).Hamim (2004) menyatakan bahwa pada tahap awal, kekeringan menyebabkan berkurangnya pembukaan stomata untuk meminimalisir kehilangan air di bawah kondisi cahaya berlebihan. Peristiwa ini mengakibatkan terjadinya penurunan konsentrasi CO2 intrasel,
sehingga tanaman mengalami overreduksi pada transfer elektron fotosintesis (Zlatev & Yordanov 2004). Hal ini dapat mengakibatkan terbentuknya reactive oxygen species (ROS) yang diawali dengan pengikatan elektron pada transfer elektron fotosintesis oleh oksigen (Asada, 2006).
Sintesis senyawa antioksidan seperti asam askorbat (Sgherri et al., 2000) yang berfungsi sebagai agen reduksi dalam menetralisir radikal oksigen dilaporkan meningkat pada beberapa spesies tanaman saat mengalami cekaman kekeringan. Selain itu tanaman juga mengakumulasi senyawa prolin yang berfungsi untuk pengaturan derajat osmotik sel (osmotic adjustment). Akumulasi prolin dapat menurunkan potensial osmotik sehingga menurunkan potensial air dalam sel tanpa membatasi fungsi enzim dan menjaga turgor sel (Hamim, 1995). Sintesis senyawa-senyawa ini mengindikasi toleransi tanaman terhadap cekaman kekeringan (Tuasamu, 2009).
Hotong umumnya tumbuh pada daerah kering dan toleran terhadap suhu tinggi, akan tetapi tidak toleran terhadap periode cekaman kekeringan yang panjang (Leder & Monda 2004). Tanaman hotong yang mengalami cekaman kekeringan panjang pada fase pertumbuhannya sukar mengalami recovery setelah dilakukan pengairan kembali (rewatering) (Karyudi & Fletcher, 2003). Antisipasi yang dapat dilakukan untuk mengurangi tingkat penurunan produksi akibat cekaman kekeringan adalah dengan menanam varietas unggul tanaman hotong yang toleran kekeringan. Dalam upaya pengembangan varietas unggul tersebut diperlukan sejumlah informasi mengenai mekanisme toleransi tanaman tersebut terhadap cekaman kekeringan, sehingga proses seleksi bisa berjalan secara efisien dan efektif.
Kemampuan penetrasi akar untuk mencapai zona yang lebih dalam pada tanaman ini merupakan suatu mekanisme penghindaran
terhadap cekaman kekeringan untuk meningkatkan kemampuan penyerapan air. Selain itu, akumulasi senyawa prolin pada daun selama periode cekaman kekeringan akan membantu dalam pelindungan jaringan tanaman dari kerusakan akibat suhu tinggi (Tuasamu, 2009).
Pada dasarnya, tanaman memiliki mekanisme tertentu untuk mempertahankan diri terhadap kondisi kekeringan dan juga cekaman lain yang ditimbulkan oleh cekaman kekeringan yaitu melalui adaptasi morfologi, fisiologi dan anatomi (Fahn, 1990). Mekanisme toleransi tanaman hotong dalam menghadapi kondisi kekeringan belum banyak diteliti. Dengan demikian menarik untuk dikaji mekanisme toleransinya melalui penelitian dengan mempelajari respon anatominya.
METODE PENELITIAN
Bahan Tanaman dan Media Tanam
S. italica yang digunakan dalam penelitian ini terdiri atas tiga aksesi hotong (A1, A2, dan A3), aksesi 1 dan 2 merupakan aksesi lokal yang diperoleh dari Dinas Pertanian Kabupaten Buru propinsi Maluku dan aksesi 3 merupakan aksesi hasil pemuliaan dari Departemen Agronomi dan Hortikultura IPB. Media tanam merupakan campuran tanah dan pasir (1:1 v/v) masing-masing 3 kg.
Perlakuan Kekeringan
Perlakuan kekeringan dilakukan saat tanaman berumur 6 minggu yang terdiri atas dua taraf yaitu perlakuan disiram setiap hari (C0), dan tanpa disiram selama 14 hari (CK).
Parameter Anatomi yang Diukur
Kerapatan dan indeks stomata pada lapisan epidermis daun bagian atas dan bagian bawah yang diukur berdasarkan persamaan Wilmer (1983) melalui preparasi sayatan paradermal berdasarkan metode Sass yang dimodifikasi (Sass, 1951), diameter xilem berdasarkan metode Nakamura (1995) melalui sayatan transversal daun, batang dan akar pada pada tanaman kontrol dan tanaman perlakuan yakni pada 14 hari setelah perlakuan (HSP) cekaman kekeringan.
HASIL DAN PEMBAHASAN
Karakteristik Anatomi Daun, Batang, dan Akar
Perlakuan cekaman kekeringan dengan menunda waktu penyiraman selama 14 hari
509
pada ketiga aksesi hotong secara nyata menyebabkan perubahan stabilitas struktur anatomi daun, batang, dan akar (Tabel1& 2).perubahan terbesar terjadi pada anatomi daun baik secara kuantitatif maupun kualitatif (Gambar 1A, B, G & H).Secara umum penurunankandungan air relatif (KAR) daun (publikasi artikel 1)pada ketiga aksesi hingga akhir periode cekaman (14 HSP) berkorelasi positif terhadap kerapatan stomata terutama pada lapisan epidermis daun bagian bawah (0.74 µm) (data penelitian penulis).Penurunan KAR daun selama periode cekaman kekeringan menyebabkan hilangnya turgiditas sel, sehingga mempengaruhi stabilitas struktur anatomi pada ketiga aksesi hotong yang ditandai dengan penururunan kerapatan stomata pada lapisan epidermis daun bagian atas maupun bagian bawah terutama pada aksesi 2 (Tabel 1).Kerapatan stomata pada aksesi 1 dan 3 cenderung lebih stabil dibanding aksesi 2.Hal ini menunjukkan bahwa aksesi 1 dan 3 lebih tahan terhadap perlakuan cekaman kekeringan dibanding aksesi 2.Penurunan kerapatan stomata pada akesi 2 diduga berpengaruh terhadap terhadap laju fotosintesis yang ditunjukkan melalui penurunan parameter fotosintesis yang berhubungan dengan penurunan produksinya dibanding aksesi 1 dan 3 (Publikasi artikel 1).
Morfologi dan anatomi daun termasuk kerapatan dan indeks stomata, diduga berpengaruh dan berhubungan dengan pertukaran gas dan faktor lingkungan seperti cahaya, status air media dan daun serta konsentrasi CO2 atmosfer.Penurunan KAM dan
KAR daun pada aksesi 2 lebih rendah dibanding aksesi 1 dan 3 (publikasi artikel 1), hal ini menunjukkan pemanfaatan air pada aksesi 2 untuk aktivitas fisiologi (transpirasi) dan proses metabolisme cukup efiesien. Efisiensi pemanfaaatan air pada aksesi 2 diduga berkaitan dengan penurunan jumlah stomata karena kurangnya air yang dibutuhkan untuk pembelahan dan pemelaran sel. Selain itu, respon aksesi 2 terhadap penyerapan cahaya mungkin lebih tinggi dari kedua aksesi lainnya, dan ini menunjukkan bahwa peningkatan laju fotosintesisnya mungkin menghambat pembentukan dan perkembangan stomata (Roelfsema et al., 2007). Keseimbangan antara suplai karbon untuk fotosintesis dan transpirasi melalui stomata mungkin secara spontan mempengaruhi efisiensi penggunaan air pada aksesi 2 sehingga mempengaruhi perkembangan selnya termasuk perkembangan stomata. Penurunan kerapatan stomata pada aksesi 2 menunjukkan bahwa keseimbangan perkembangan luas daun dan jaringannya mungkin berkaitan dengan perkembangan sel
penjaga pada kondisi status air yang rendah (Galmes et al., 2007).
Zhang et al., (2006), menyatakan bahwa kerapatan stomata daun berkorelasi positif dengan konduktasi stomata terhadap laju transpirasi dan laju fotosinteisis neto, sebaliknya laju transpirasi meningkat dengan meningkatnya jumlah stomata, akan tetapi tidak signifikan. Lebih lanjut dijelaskan oleh Zhou dan Xu (2008), bahwa respon efisiensi penggunaan air (fotosintesis dan transpirasi) pada kondisi cekaman kekeringan menunjukkan korelasi positif antara kerapatan stomata dengan efisiensi penggunaan air. Berdasarkan hal ini, diduga kestabilan kerapatan dan indeks stomata pada aksesi 1 dan 3 berkaitan dengan peningkatan laju transpirasi dan fotosintesis, sehingga KAM dan KAR daun pada kedua aksesi ini lebih rendah dibanding aksesi 2 (publikasi artikel 1).
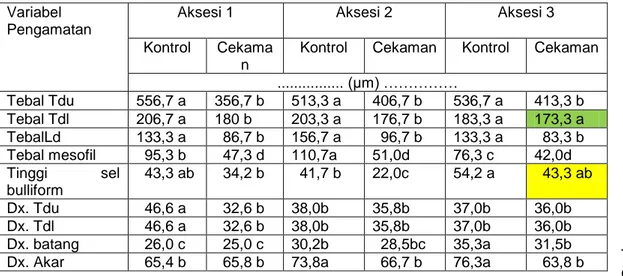
Perubahan stabilitas struktur anatomi tanaman hotong juga ditandai dengan penurunan tebal tulang daun utama, tebal tulang daun pada lamina, tebal lamina daun, tebal mesofil, dan diameter xilem pada tulang daun utama maupun tulang daun pada lamina (Tabel 2).Perubahan terbesar terjadi pada aksesi 1 dan 2. Hal ini disebabkan oleh defisit air akibat penurunan KAM dan KAR daun (aksesi 1), efisiensi penggunaan air (aksesi 2) untuk aktivitas fisiologi dan metabolisme dasar lainnya mempengaruhi penurunan tekanan turgor sel. Penurunan tekanan turgor sel ini selanjutnya akan menghambat proses pemanjangan dan pemelaran sel (Salisbury & Ross, 1992), akibatnya terjadi pengkerutan sel yang tidak dapat balik (Sakurai & Kuraishi, 1988). Fenomena ini juga menyebabkan penurunan jumlah stomata, reduksi area permukaan daun, dan penggulungan daun yang berbeda nyata pada aksesi 2 (publikasi artikel 1). Wise dan Naylor (1987) menyatakan bahwa, cekaman kekeringan menyebabkan perubahan struktur anatomi da ultrastruktur sel antara lain menurunkan tebal daun, tebal mesofil, jumlah kloroplas, perubahan orientasi tilakoid dan peningkatan tebal dinding sel.
Penurunan ukuran diameter xilem pada daun tanaman perlakuan lebih besar dibanding pada batang dan akar.Perubahan ini diduga disebabkan oleh tekanan akar (tekanan akar lebih negative dibanding tajuk), sehingga mempengaruhi ukuran diameter xilem pada daun yang terlihat agak pipih (Gambar 1B). Hal ini menunjukkan bahwa konduktivitas hidrolik xilem lebih baikpada batang dan akar masih cukup baik. Menurut Radwan (2007), xilem dengan diameter yang lebih besar memiliki konduktivitas air di dalam xylem lebih baik dan cepat prosesnya dibanding xylem yang
510
berdiameter kecil. Perubahan karakteristik anatomi daun ini di pengaruhi oleh besarnya tingkat cekaman yang dialami pada fase pertumbuhan tanaman.Secara kualitatif, perubahan struktur anatomi tanaman hotong dapat diamati melalui sayatan transversal daun pada ketiga aksesi (Gambar 1 A – H). Susunan struktur anatomi daun pada ketiga aksesi hotong menunjukkan susunan struktur yang sama yaitu dari permukaan daun bagian atas berturut-turut adalah lapisan epidermis atas (adaxialepidermis), sel bulliform (sel kipas) yang menyatu dengan sel epidermis atas, sel-sel mesofil, bundle sheat (sel seludang pembuluh) dan jaringan pembuluh serta lapisan epidermis bawah (abaxial epidermis) (Gambar 1A).pengamatan secara kualitatif menunjukkan perubahan yang nyata antara daun tanaman kontrol dengan perlakuan kekeringan (Gambar 1A, B, G, & H). sedangkan pada batang dan akar tanaman perlakuan tidak mengalami perubahan yang nyata seperti pada struktur anatomi daun (Gambar 1C – F). Gambar 1Bb menunjukkan terjadinya pengkerutan sel bulliform.Sel ini tidak atau sedikit mengandung kutikula, sehingga hilangnya air melalui transpirasi berlangsung lebih cepat dibanding sel epidermis.Perubahan ini merupakan respon tanaman terhadap cekaman kekeringan untuk mengurangi laju transpirasi dan mempertahankan KAR daun (Salisbury & Ross, 1992).
Penurunan KAR daun selama periode cekaman (publikasi artikel 1) diduga mempengaruhi pemelaran atau pembesaran sel pada ketiga aksesi terutama pada daun (Gambar 1A, B, G & H). Penghambatan pemelaran sel terlihat pada kecilnya ukuran diameter xilem terutama pada daun tanaman perlakuan sehingga menghambat proses transportasi air ke daun yang mengakibatkan terjadinya penurunan tekanan turgor sel dan pelayuan (dokumentasi peneliti). Proses pelayuan dan penggulungan daun tanaman hotong merupakan salah satu respon tanaman terhadap kondisi rawan air akibat cekaman kekeringan dan secara visual mulai tampak pada 8 hari setelah perlakuan (HSP) hingga akhir periode cekaman (dokumentasi peneleti). Penggulungan daun terjadi Karena proses pengkerutan sel bulliform (Gambar 4Bb) yang berfungsi untuk melindungi jaringan dibawahnya agar tidak mengalami kerusakan akibat kehilangan air yang lebih besar.
Secara umum penurunan tebal lapisan mesofil pada ketiga aksesi pada 14 HSP kekeringan berkorelasi positif dengan parameter fotosintesis terutama qP (0,83 m-2s-1) dan qY (0,84 m-2s-1) (publikasi artikel 1). Pengecilan
ukuran sel mesofil (Gambar 1Bc) diduga mempengaruhi tahanan mesofil terhadap difusi CO2, sehingga mempengaruhi perubahan laju
fotosintesis terutama pada aksesi 1 dan 2 (publikasi artikel 1).Perubahan struktur anatomi daun terbesar terjadi pada aksesi 2 dibanding aksesi 1 dan 3 (Tabel 1 dan 2), sehingga menurunkan laju fotosintesisnya dan tidak mampu mempertahankan produksi setara dengan tanaman kontrol (publikasi artikel 1).Hal ini menunjukkan bahwa aksesi 2 peka terhadap perlakuan cekaman kekeringan disbanding aksesi 1 dan 3.
Pengamatan pada epidermis daun menunjukkan bahwa ketiga aksesi hotong memiliki stomata dengan sel penjaga memanjang berbentuk halter dan tersusun dalam deretan sejajar (Gambar 1 G & H). Ujung sel penjaga berdinding tipis dan bagian tengah memanjang berdinding tebal dengan lumen sel menyempit. Bentuk stomata ini berperan sebagai unit fisiologi tunggal yaitu sel penjaga dengan segera akan merapat ketika terjadi perubahan tekanan turgor sel akibat penurunan kadar air media (KAM). Selain itu, stomata berada pada kedua lapisan epidermis (atas dan bawah) yang disebut dengan amfistomatik dan diduga berkaitan dengan peningkatan laju transpirasi dan asimilasi CO2.
Cekaman kekeringan selama 14 hari juga menyebabkan ukuran stomata dan sel penjaga pada tanama perlakuan terlihat lebih kecil dan tipis disbanding tanaman control (Gambar 1 Hg & h).Pengecilan ukuran stomata dan sel penjaga ini disebabkan oleh pengkerutan sel yang merupakan respon tanaman terhadap cekaman kekeringan untuk meminimalisir kehilangan air melalui transpirasi dan meningkatkan efisiensi asimilasi CO2 (Zhou
& Xu, 2008)..
KESIMPULAN
Aksesi 3 (A3) merupakan aksesi hotong yang tahan terhadap cekaman kekeringan yang ditunjukkan dengan respon yang berbeda dibanding aksesi yang peka (A2) dalam menghadapi cekaman kekeringan jika dilihat berdasarkan kestabilan struktur anatomi daun terutama kerapatan stomata, tebal daun dan lapisan mesofil yang relatif sama dengan tanaman kontrol.
SARAN
511
yang baik untuk dibudidaya pada lahan-lahan kering dengan curah hujan berkisar antara 1000–1400 mm/thn dan perlu penelitian lebih lanjut untuk skala yang lebih luas (penelitian lapang) dan isolasi serta karakterisasi ultrastruktur anatomiuntuk memperoleh informasi dasar ke arah molekuler.DAFTAR PUSTAKA
Andrawulan. 2003. Analisis Kadar Gizi Hotong. Jurusan Teknologi Pangan dan Gizi. Fakultas Teknologi Pertanian. Bogor: Institut Pertanian Bogor.
Asada K. 2006. Production and scavenging of reactive oxygen species in chloroplast and their function. Plant Physiol 143: 391-396. Bohnert HJ, Nelson DE, Jensen RG. 1995.
Adaptation to environmental stress.Plant Cell. 7: 1099-1111.
[Diptan] Dinas Pertanian Tanaman Pangan. 2003. Pengembangan Usaha Tani Tanaman Pangan Lahan Kering dan Pelaksanaan Program Transmigrasi di Kabupaten Buru [laporan hasil penelitian].Kerjasama Fakultas Pertanian Universitas Pattimura dengan Pemerintah Kabupaten Buru.
Fahn A. 1990. Plant Anatomy.4th Ed. New York: Pergamon Press.
Galmes J, Flexas J, Save R, Medrano H. 2007. Water relations and stomatal chracteristics of mediterranean plants with different growth forms and leaf habits: responses to water stress and recovery. Plant and Soil. 290: 139-155.
Hamim. 1995. Toleransi Kedelai Terhadap Cekaman Kekeringan; Pendekatan Morfologi dan Fisiologi. [tesis] Program Pascasarjana. Bogor: Institut Pertanian Bogor.
. 2004. Underlying drought stress effects on plant: Inhibition of photosynthesis [ulasan]. Hayati 11(4): 164-169.
Karyudi, Fletcher RJ. 1999. Osmoregulative capacity in Setaria italica L. (Foxtail millet) and Panicum miliacum L. (Proso millet) in respon to water stress after heading. Proceedings. 11th Australian Plant Breeders’ Conference, Adelaide.
. 2003. Osmoregulation in birdseed millet under conditions of water stress II. Variation in F3 lines of Setaria italica and its relationship to plant morphology and yield. Euphytica 132: 191-197.
Levitt J. 1980. Responses of Plant to Environmental Stresses Water. Radiation Salt and Other Stresses. New York: Academic Press.
Leder I, Monda S. 2004. The sorghum and millet and their use.In Hongarian.
Elemezesi ipar 39 (3): 98-104.
Nakamura TN. 1995. Method for Cells and Tissues Observation.In Hinata K, Hashiba T, Ed.A Manual Experiment for Plant Biology. Tokyo: Soft Science Publication. Hlm 15-21.
RadwanAAU. 2007. Plant water relations, stomatal behavior, photosynthetic pigments and anatomical characteristics of Solanostemma arghel (Del). Hayne under
hyper environmental
conditions.SciResearch 2(2): 80-92.
RoelfsemaMRG, Konrad K, Marten H, Psaras G, Hartung W, Hedrich R.2006. Guard cells in albino leaf patcehes do not respond to photosynthetically active radiation, but are sensitive to blue light, CO2 and abscisic acid. Plant Cell and Envoronment. 29: 1595-1605.
Sakurai N, Kuraishi S. 1998. Water potential and mechanical properties of the cell wall of hypocotyles of dark-grown squash (Cucurbita maxima Duch) under water stress conditions. Plant and Cell Physiol. 29: 1337-1343.
Salisbury FB, Ross CW. 1992. Plant Physiology. California: 4th Ed Wadsworth Publishing Company.
Sass JE. 1951. Botanical Microtechnique. Iowa: The Iowa State Collage Press.
Sgherri C, Maffei LM, Navari-Izzo F. 2000. Antioxidative enzymes in wheat subjected to increasing water deficit and rewatering. Plant Physiol 157: 273-279.
Tuasamu Y. 2009. Toleransi Hotong (Setaria italica L. Beauv) pada Berbagai Cekaman Kekeringan: Pendekatan anatomi dan Fisiologi. [tesis]. Sekolah Pasca Sarjana. Institut Pertanian Bogor. Bogor.
512
Tuasamu Y. 2010. Respon Fisiologi Hotong(Setaria italica L. Beauv) Terhadap Berbagai Cekaman Kekeringan. [Artikel 1]. Jurnal Bimafika FKIP Unidar. Universitas Darussalam. Ambon.
Sgherri C, Maffei LM, Navari-Izzo F. 2000. Antioxidative enzymes in wheat subjected to increasing water deficit and rewatering. Plant Physiol 157: 273-279.
Wilmer CM. 1983. Stomata. Department of Biology University of Stirling.Longman Inc. Wise Nailor AW. 1987. Chilling-enhanced
photo-oxidation. The peroxidative destruction of lipids during chilling injury to photosynthesis and ultrastucture. Acta Phytoecologica Sinica. 27:133-136.
Zhang XY, Wang HM, Hou ZD, Wang GX. 2003. Stomatal dencity and distributions of spring wheat leaves under different planting
densities and soil moisture levels. Plant Physiol. 83:278-282.
Zhang YP, Wang ZM, Wu YC, Zhang X. 2006. Stomatal characteristics of different planting densities green organs in wheat under different irrigation. Acta Agronomica Sinica. 32:70-75.
Zhou G, Xu Z. 2008. Responses of leaf stomatal dencity to water status and its relationship with photosynthesis in a grass . Exp Bot. 59 (12):3317-3325.
Zlatev ZS, Yordanov IT. 2004. Effects of soil drought on photosynthesis and chlorophyll fluorescence in bean plant. Plant Physiol. 30(3-4): 3-18.
LAMPIRAN
Tabel 1. Pengaruh cekaman kekeringan terhadap kerapatan dan indeks stomata lapisan epidermis daun bagian atas dan bagian bawah pada ketiga aksesi hotong pada 14 hari setelah perlakuan cekaman kekeringan.
Variabel Pengamatan
Aksesi 1 Aksesi 2 Aksesi 3
Kontrol Cekaman Kontrol Cekaman Kontrol Cekaman KSEA (mm2) 120,6 ab 118,4 ab 140,4 a 109,6 b 140,4 a 138,2 ab KSEB (mm2) 157,9 a 120,6 ab 155,7 a 107,5 b 155,7 a 129,4 ab ISEA (%) 15,0 b 19,4 a 19,0 a 17,2 ab 17,2 ab 20,6 a ISEB (%) 18,9 a 17,8 a 18,7 a 19,4 a 17,6 a 17,9 a KSEA &KSEB : kerapatan stomata pada lapisan epidermis daun bagian atas dan bawah, ISEA & ISEB : indeks stomata pada lapisan epidermis daun bagian atas dan bawah. Angka yang diikuti oleh huruf yang samapada masing-masing baris tidak berbeda nyata antara tanaman kontrol dan perlakuan pada taraf uji 5%(DMRT).
513
Tabel 2. Pengaruh cekaman kekeringan terhadap struktur anatomi daun, batang, dan akar ketiga aksesi hotong pada 14 hari setelah perlakuan cekaman kekeringan.
T d u :tulang daun utama, Tdl : tulang daun pada lamina. Ld : lamina daun. Dx : diameter xilem. Angka yang diikuti oleh huruf yang samapada masing-masing baris tidak berbeda nyata antara tanaman kontrol dan perlakuan pada taraf uji 5%(DMRT).
A B C D
E F G H
Gambar 1.Sayatan transversal daun, batang, & akar serta sayatan paradermal daun ketiga aksesi hotong pada tanaman kontol & perlakuan pada 14 hari setelah perlakuan cekaman kekeringan.Struktur anatomi daun tanaman kontrol (A) & cemakan (B), xilem tanaman kontrol (C) & cemakan (F),xilem batang tanaman kontrol (G) & cemakan (H).Sel epidermis atas (a) sel bulliform (b),sel mesofil (c) bundle sheat (d) xilem (e),floem (f),stomata (g), & sel panjang (h).Tanda panah (xyilem akar & batang).Tanda bintang (floem akar & batang).Daun & akar pembesaran 400x.Batang pembesaran 1000x.
Variabel Pengamatan
Aksesi 1 Aksesi 2 Aksesi 3 Kontrol Cekama
n
Kontrol Cekaman Kontrol Cekaman ... (µm) ………
Tebal Tdu 556,7 a 356,7 b 513,3 a 406,7 b 536,7 a 413,3 b Tebal Tdl 206,7 a 180 b 203,3 a 176,7 b 183,3 a 173,3 a TebalLd 133,3 a 86,7 b 156,7 a 96,7 b 133,3 a 83,3 b Tebal mesofil 95,3 b 47,3 d 110,7a 51,0d 76,3 c 42,0d Tinggi sel bulliform 43,3 ab 34,2 b 41,7 b 22,0c 54,2 a 43,3 ab Dx. Tdu 46,6 a 32,6 b 38,0b 35,8b 37,0b 36,0b Dx. Tdl 46,6 a 32,6 b 38,0b 35,8b 37,0b 36,0b Dx. batang 26,0 c 25,0 c 30,2b 28,5bc 35,3a 31,5b Dx. Akar 65,4 b 65,8 b 73,8a 66,7 b 76,3a 63,8 b