POLIMORFISME GEN SRY PADA POPULASI MONYET EKOR
PANJANG DI KAWASAN JAWA TIMUR
I Nengah Wandia1,2), I GA. Arta Putra1,3) I Gede Soma1,2) 1
Pusat Penelitian Satwa Primata LPPM Unud, Kampus Bukit Jimbaran Bali, E-mail: [email protected]
2
Fakultas Kedokteran Hewan, Universitas Udayana, Jl. PB Sudirman, Denpasar
3
Fakultas Peternakan Universitas Udayana, Kampus Bukit Jimbaran Bali
Abstract
A gene polymorphism is a genetic resource within a species or population, that its existence is not yet broadly explored in Indonesia.This research aimed to examine the spatial distribution of the variation of SRY gene in long tailed macaque populations in the Region of East Java. The total of 21 male macaque blood samples was collected from 2 populations (Baluran and Alas Purwo) and extracted using QIAamp® Blood Mini Kits to find out the total DNA. The SRY gene was amplified by PCR technique using a specific primer, and then, sequenced by dideoxynucleotide chain-termination method at public company of Macrogen Inc., Korea. The research found that the length of segment of SRY gene was 733 nucleotide with 1 variable site (T182A). Two haplotypes were found out in total populations, which were distributed to one haplotype in Alas Purwo population and two haplotypes in Baluran population.The result of the researh indicated that male macaques migrated in one direction, that was, from Alas Purwo popuatian to Baluran population. Phylogenetic reconstruction analysis showed that all haplotypes were seperated to be two haplogroups, namely the clade of Baluran, which is composed of sry_baluran haplotype and the clade of Alas Purwo which is composed of sry_alas_purwo haplotype. It can be concluded that both haplotypes can be rendered as a marker of identity for each population.
Key words:long tailed macaque, locus polymorphism, SRY gene, haplotype, East Java
Abstrak
Polimorfisme suatu gen adalah sumber daya genetik di dalam spesies/populasi, yang keberadaannya belum banyak diungkapkan di Indonesia. Penelitian ini bertujuan untuk mengungkapkan sebaran spasial dari variasi gen SRY pada populasi monyet ekor panjang di Kawasan Jawa Timur. Sejumlah 21 sampel darah monyet jantan dikoleksi dari dua populasi (Baluran dan Alas Purwo) dan diekstraksi menggunakan QIAamp® Blood Mini Kits untuk mendapatkan DNA total. Gen SRY diamplifikasi menggunakan satu set primer khusus melalui teknik PCR dan selanjutnya, disekuen dengan metode dideoxynucleotide chain-termination pada perusahaan public Macrogen Inc., Korea. Hasil penelitian mendapatkan bahwa panjang segmen gen SRY 733 nukleotida dengan 1 situs variatif (T182A). Sebanyak 2 haplotipe ditemukan di seluruh populasi, dengan distribusi 1 haplotipe di populasi Alas Purwo dan 2 haplotipe di Populasi Baluran. Hasil penelitian mengindikasikan bahwa monyet jantan bermigrasi dalam satu arah yaitu dari populasi Alas Purwo ke populasi Baluran. Analisis rekonstruksi pohon filogeni menunjukkan bahwa seluruh haplotipe terbagi dalam 2 haplogroup yaitu clade Baluran yang tersusun atas haplotipe syr_baluran dan clade Alas Purwo yang tersusun atas haplotipe sry_alas_purwo. Berdasarkan pada hasil penelitian, dapat disimpulkan bahwa kedua jenis haplotipe ini dapat dijadikan marka identitas untuk masing-masing populasi.
2
PENDAHULUAN
Polimorfisme genetik populasi merupakan biodiversitas pada tingkat yang paling dasar.
Kerakteristik genetik ini merupakan refleksi dari kisah kehidupan yang telah dilaluinya dimasa
lampau dan masa kini, serta dapat digunakan untuk memprediksi kondisi mendatang yang akan
dialaminya (Nozawa et al., 1996; Hartl dan Clark, 1997; Frankham et al., 2004). Oleh karena itu,
data biodiversitas pada tingkat genetik suatu spesies atau populasi bukan saja dapat menerangkan
sejarahnya, tetapi juga dapat digunakan sebagai dasar pertimbangan strategi konservasi dan bahan
referensi sehubungan dengan molekuler forensik.
Monyet ekor panjang (Macaca fascicularis) adalah primata non human yang hidup dalam
sebuah kelompok sosial multi-male multi-female group dan besifat filopatri betina (Jolly, 1985).
Dalam satu kelompok sosial terdapat beberapa jantan dan betina dewasa serta anak-anaknya
(Napier dan Napier, 1985; Mitchell dan Erwin, 1986; Bennett et al., 1995; Rowe 1996). Monyet
betina membentuk inti permanen dari kelompok sosial karena tetap tinggal pada kelompok
kelahirannya, sementara, monyet jantan sering bermigrasi ke kelompok sosial lainnya (Napier dan
Napier, 1985; Mitchell dan Erwin, 1986).
Sejarah penyebaran monyet ekor panjang di Kepulauan Selatan Indonesia masih belum
jelas. Akhli primata meyakini bahwa penyebarannya berjalan dari barat ke timur dengan Jawa
sebagai sumber awal populasi (Wandia, 2007). Migrasi monyet ekor panjang dari Pulau Jawa ke
Pulau Bali berlangsung beberapa kali dan migrasi terakhir terjadi ± 18 ribu tahun yang lalu (Eudey,
1980; Fooden, 1995). Kini, monyet ekor panjang hidup sebagai populasi-populasi lokal (habitat
terbatas) sebagai akibat dari fragmentasi habitat yang ditimbulkan oleh peningkatan penggunakan
lahan untuk pertanian, industri, dan area kependudukan di dalam suatu pulau (Kawamoto et al.,
1984; Wandia, 2003; Wandia et al., 2004; Brotcorne et al., 2014; Brotcorne et al., 2015). Namun,
bagaimana proses pemisahan populasi di dalam suatu pulau dan hubungan genetik antar populasi
masih belum banyak diungkapkan.
Struktur genetik populasi dapat diungkapkan dengan menerapkan berbagai sumber marka
molekuler. Kawamoto et al. (1984) mengungkapkan variasi genetik monyet ekor panjang di
Indonesia dengan marka protein darah. Beliau menemukan bahwa ada ketidakselarasan antara
keragaman genetik bersanding dengan letak geografi kelompok monyet ekor panjang Jawa, Bali,
dan Lombok. Keragaman genetik populasi di Pulau Bali seyogyanya lebih tinggi daripada yang
ditemukan di Pulau Lombok sebagai akibat dari efek founder, tetapi hasil penelitian Kawamoto et
al. (1984) menunjukkan hal sebaliknya. Wandia (2007) yang meneliti mengenai variasi genetik
monyet ekor panjang Jawa, Bali, dan Lombok dengan menggunakan marka molekuler DNA
mikrosatelit kromosom somatik menemukan bahwa variasi genetik monyet Bali lebih tinggi
daripada yang ditemukan di Lombok. Hasil tersebut sejalan dengan pola penyebarannya dari barat
Bali menggunakan 3 lokus mikrosatelit pada kromosom Y. Beliau menemukan bahwa diversitas
haplotipe populasi sangat rendah karena ketiga marka molekuler bersifat monomorfik, dan ketiga
marka molekuler tidak mampu membedakan karakteristik genetik antar populasi.
Untuk itu, penelitian pendahuluan kali ini dilakukan dengan menerapkan gen sex
determining region kromosom Y sebagai marka molekuler untuk mengungkapkan struktur genetik
populasi yang ditelusuri dari garis jantan. Penelitian dilakukan pada dua populasi monyet ekor
panjang di Kawasan jawa Timur . Gen sex determining region (SRY) merupakan gen yang sangat
penting untuk mengontrol perkembangan seksual jantan, yang terletak di antara pseudosomatic
regions (PARs) dan eukromatin pada lengan pendek kromosom Y (Bachtrog dan Charlesworth
2001). Gen ini tidak ditemukan pada kromosom X sehingga sering digunakan sebagai uji
penentuan jenis kelamin (Erdal dan Barlas 2000; Drobnic 2006; Rovie-Ryan et al. 2013). SRY
mengkode produk testis determinating factor (TDF) yang merangsang jaringan gonad embrio
yang belum terdiferensiasi untuk membentuk testis. SRY atau versi lainnya yang berhubungan
ditemukan pada seluruh mammalia (Klug dan Cummings, 2005).
MATERI DAN METODE PENELITIAN
Lokasi Sampling
Penelitian dilakukan pada dua populasi monyet ekor panjang yaitu populasi Alas Purwo
(Kabupaten Banyuwangi, Jawa Timur) dan Baluran (Kabupaten Situbondo, Jawa Timur). Kedua
populasi terletak di ujung timur Pulau Jawa, yang berpotensi besar bermigrasi ke Pulau Bali. Jarak
antar populasi lebih kurang 100 km (diukur sepanjang garis pantai dengan bantuan fasilitas google
map (https://www.google.co.id/maps )
Sampel Darah
Sampel darah monyet ekor panjang jantan berhasil dikoleksi sebanyak 12 sampel dari
populasi Alas Purwo dan 10 sampel dari populasi Baluran. Sampel diambil saat monyet dalam
kondisi terbius (dibius menggunakan Ketamin HCl dosis 10 mg/kg bobot badan dikombinasi
xylazil dengan perbandingan 5:1). Darah sebanyak 5 ml diambil dengan menggunakan alat suntik
10 ml yang telah diisi 0,1 ml EDTA 10% sebagai antikoagulan dari vena femoralis. Sampel darah
telah tersedia dan disimpan di Laboratorium Genetika dan Kultur Jaringan- Pusat Penelitian Satwa
Primata, LPPM Unud Kampus Bukit-Jimbaran.
Ekstraksi DNA Total dan Perancangan Primer Gen SRY
Total DNA diekstraksi menggunakan QIAamp DNA Blood Kits produksi Qiagen yang
prosedurnya sesuai dengan protokol yang direkomendasikan oleh perusahaan (Qiagen, 2007).
4
SRY referensi AF284304 (GenBank). Perancangan dilakukan in silico menggunakan software
primer3 secara online (http://www.ncbi.nlm.nih.gov/genbank/). Tiga pasang primer dipilih yaitu:
1. SRY F 5’-AGGGGGTAGCCTGGTTGGGC-3’
SRY R 5’-AGTGGCTGTAGCGGTCCCGT- 3’
2. SRY F 5’-GGTGTTGGGGGCGGAGAAATG-3’
SRY R 5’-TGGCTGTAGCGGTCCCGTTG-3’
3. SRY F 5’-TTGGGCGGAGTTGAGAGGGG-3’
SRY R 5’-TGATGGGCGGTAAGTGGCCT-3’
Ketiga pasang primer diujicobakan dan PCR yang menghasilkan pita tunggal dipilih untuk
penelitian lebih lanjut. Volume PCR dibuat 25 µL yang setiap reaksinya mengandung adalah 1x
bufer PCR, 3 mM MgCl2, 0,2 mM DNTP (mix), 0,2 mM masing-masing primer, 0,75 U DNA
polimerase, dan template dengan volume 1 µL. PCR dilakukan 3 tahapan yaitu Pre PCR: 94 oC 3
menit; PCR (diulang 35 siklus) : 94 oC 40 detik, 58 oC 40 detik, 72 oC 40 detik; dan Post PCR: 72
o
C 5 menit. Berdasarkan pada hasil PCR, primer 2 menghasillkan produk PCR tunggal (pita
tunggal) sekitar 700 nukleotia dan dipilih untuk penelitian selanjutnya.
Amplifikasi Gen SRYdan Sekuensing
Amplifikasi gen SRY menggunakan sepasang primer khusus (Primer 2). Satu unit PCR
mengandung 1x buffer PCR, 4 mM MgCl2, 0,2mM DNTP (mix), 0,2 µM masing-masing Primer,
dan 1 U enzim DNA polymerase. Sebanyak 2 µl template dan aquades bebas ion ditambahkan
untuk mendapatkan volume akhir 50 µL. PCR terdiri atas 3 tahap yaitu Pre PCR dengan suhu
danaturasi 94oC selama 3 menit; PCR 40 siklus dengan urutan denaturasi 94oC selama 40 detik,
annealing 58oC selama 40 detik, dan elongasi 72oC selama 40 detik; Post PCR dengan suhu
elongasi 72oC selama 5 menit. PCR menggunakan mesin Applied Biosystems 2720 Thermal
Cycler. Hasil amplifikasi dipisahkan secara elektroforesis dengan gel agarose 1,5% yang
dimigrasikan pada 50 V selama 30 menit. Fragmen teramplifikasi dimunculkan dengan pewarnaan
etidium bromida dan ukuran panjang basa disetarakan dengan menggunakan marker standard 100
bp DNA ladder (Gibco BRL, Life Technologies). Produk PCR, selanjutnya, disekuen dengan
metode dideoxynucleotide chain-termination pada perusahaan publik Macrogen Inc. Korea.
Analisis Data
Pelurusan dan pengeditan urutan nukleotida menggunakan software MEGA 6 (Tamura et
al., 2013). Analisis terhadap jenis haplotipe dan distribusinya serta diversitas genetik dalam
populasi (tingkat nukleotida dan hapotipe) menggunakan software dnaSPver. 5.10.01 (Rozas et al.,
dan metode maximum likelihood dikerjakan dengan bantuan software MEGA 6 (Tamura et al.,
2013).
HASIL DAN PEMBAHASAN
Distribusi Haplotipe Gen SRY
Dari total 22 sampel, 21 sampel menghasilkan produk PCR sedangkan 1 sample lainnya
tidak menghasilkan produk PCR. Selanjutnya, sekuensing dilakukan terhadap 21 sampel. Analisis
urutan nukleotida menghasilkan panjang gen SRY 733 nukleotida dengan komposisi 22,1% (T/U),
23,1% (C), 30,7% (A), dan 24,1% (G). Satu 1 situs variatif (parsimony informative site) ditemukan
dari keseluruhan panjang segmen. Mutasi titik T/A terjadi pada situs 182 (tipe transversi). Analisis
urutan penyandi protein pada gen SRY memperlihatkan substitusi T182A tidak mengubah translasi
asam amino (silent mutatian).
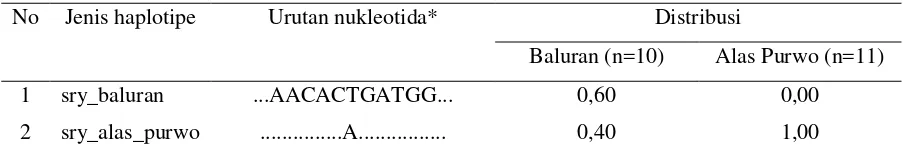
Sebanyak 2 jenis haplotipe gen SRY ditemukan dari seluruh sampel (Tabel 1). Pada
Populasi Alas Purwo ditemukan 1 jenis haplotipe (sry_alas_purwo), sementara kedua haplotipe
(sry_baluran dan sry_alas_purwo) ditemukan di populasi Baluran . Rekonstruksi filogeni seluruh
haplotipe membentuk dua kelompok (haplogroup) yaitu clade Alas Purwo dan clade Baluran
(haplogroup dinamai secara arbitrari). Masing-masing haplogroup memiliki jenis haplotipe yang
unik (Gambar 1).
Adanya shared haplotipe mencerminkan adanya gene flow atau migrasi antar populasi.
Keunikan haplotipe pada masing-masing populasi terlebih haplotipe dominan dapat digunakan
sebagai haplotipe penanda/haplotipe identitas untuk masing-masing populasi. Sejarah evolusi dan
perkembangan populasi dimasa mendatang dapat diikuti melalui pemantauan dinamika haplotipe
yang bersifat unik tersebut.
Tabel 1. Haplotipe Gen SRY dan Distribusi pada Populasi Monyet Ekor Panjang di Kawasan Jawa Timur
No Jenis haplotipe Urutan nukleotida* Distribusi
Baluran (n=10) Alas Purwo (n=11)
1 sry_baluran ...AACACTGATGG... 0,60 0,00
2 sry_alas_purwo ...A... 0,40 1,00
* Nukleotida yang ditampilkan adalah nukleotida 177-187. Nukleotida variatif pembeda haplotipe: nukleotida 182.
Diversitas Genetik Dalam Populasi dan Diferensiasi Genetik Antar Populasi
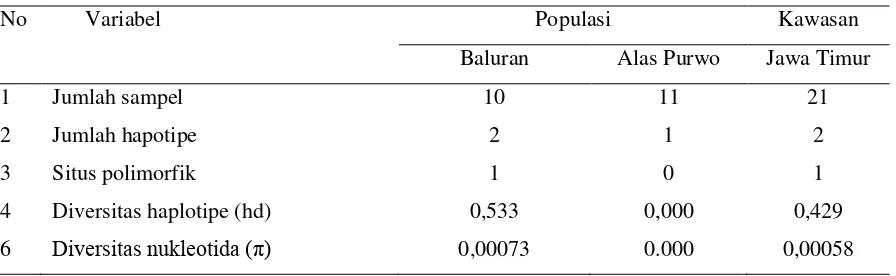
Diversitas genetik dalam populasi yaitu diversitas haplotipe dan diversitas nukleotida
ditampilkan pada Tabel 2. Diversitas haplotipe (hd) menyatakan peluang dua haplotipe untuk
berbeda saat diambil secara bersamaan, sedangkan diversitas nukelotida mewakili peluang
6
Populasi Baluran memiliki hd yang tinggi (hd=0,533) sedangkan populasi Alas Purwo memiliki hd
yang rendah (hd=0,000). Hal ini berkaitan dengan adanya 2 haplotipe di populasi Baluran dengan
frekeuensi relatif sama. Sebaliknya, pada populasi Alas Purwo ditemukan hanya satu jenis
hapotipe. Rendahnya diversitas genetik dalam populasi mencerminkan sangat rendah atau tidak
adanya gene flow ke dalam populasi, atau populasi mengalami bottle neck, dan atau merupakan
efek founder (Avise, 1994; Nozawa et al., 1996; Hartl dan Clark, 1997).
Tabel 2. Diversitas Genetik Dalam Populasi
No Variabel Populasi Kawasan
Baluran Alas Purwo Jawa Timur
1 Jumlah sampel 10 11 21
2 Jumlah hapotipe 2 1 2
3 Situs polimorfik 1 0 1
4 Diversitas haplotipe (hd) 0,533 0,000 0,429
6 Diversitas nukleotida (π) 0,00073 0.000 0,00058
Diferenssai genetik antar populasi dengan menggunakan tiga ukuran ditampilkan pada
Tabel 3. Nilai Nst dengan Fst sama, dengan memberikan nilai gene flow (Nm)= 0,8 ekor/generasi,
sementara dengan ukuran Gst memberikan nilai Nm= 1,53 ekor/generasi. Nm mencerminkan
jumlah individu yang bermigrasi antar populasi per generasi. Dari hasil analisis dapat dinyatakan
bahwa migrasi antar populasi 1-2 ekor per generasi dengan pola migrasi satu arah (dari Alas Puwo
ke Baluran sesuai jenis haplotipe yang ditemukan (Tabel 1).
Tabel 3. Diferensiasi genetik antar populasi Baluran dan Alas Purwo Jawa Timur.
Ukuran diferensiasi
genetik
Nilai diferensiasi gentik Gene flow (Nm)
(ekor/generasi)
Referensi
GST 0,39572 1,53 Nei 1973
Nst 0,55556 0,80 Lynch and Crease 1990
Fst 0,55556 0,80 Hudson, Slatkin and Maddison
1992
Jarak Genetik Antar populasi dan Rekonstruksi Filogeni
Jarak genetik antar populasi mengukur rata-rata divergensi nukeotida dua haplotipe antar
populsi (Excoffier et al., 2005). Jarak genetik populasi Baluran dengan Alas Purwo sangat rendah
antar populasi, seperti ditemukannya haplotipe sry_alas_purwo pada populasi Baluran (shared
haplotipe).
Tabel 4. Jarak Genetik Netto Antar Populasi di Kawasan Jawa Timur
Populasi Baluran Alas Purwo
Baluran 0,0004
Alas Purwo 0,0005
Keterangan: Angka di bawah diagonal menyatakan jarak genetik (metode Kimura 2 parameter . Angka di atas diagonal menyatakan standar eror (bootstrap 1000)
Rekonstruksi pohon filogeni menghasilkan 2 kelompok haplotipe (haplogroup) yaitu clade
Alas Purwo dan clade Baluran (Gambar 1). Empat individu yang berasal dari Baluran (BL M8, BL
M9, BL M10, dan BL M12) masuk ke dalam haplogroup Alas Purwo. Hal ini menunjukkan bahwa
migrasi monyet jantan telah terjadi dari populasi Alas Porwo ke populasi Baluran, namun, arah
migrasi balik tidak terjadi (tidak ditemukan jenis hapotipe sry-baluran di populasi Alas Purwo
(Tabel 1).
KESIMPULAN
Pada Populasi Baluran ditemukan dua jenis haplotipe (sry_Alas-Purwo dan sry_baluran)
dengan diversitas haplotipe (hd) sebesar 0,533 dan diversitas nukleotida (π) sebesar 0,00073,
sedangkan pada populasi Alas Purwo hanya ditemukan satu jenis haplotipe (sry_alas_purwo).
Gambar 1. Rekonstruksi filogeni monyet ekor panjang di Jawa Timur. Model
substutusi Kimura 2 parameter metode maximum likelihood
8
Diferensiasi genetik antar populasi cukup tinggi (Gst=0,396). Haplotipe gen SRY membentuk dua
haplogroup yaitu clade Alas Purwo yang tersusun oleh haplotipe sry_alas_puwo, dan clade
Baluran yang tersusun oleh haplotipe sry_baluran. Tipe haplotipe pada masing-masing clade dapat
dijadikan marka identitas untuk masing-masing populasi.
UCAPAN TERIMA KASIH
Ucapan terima kasih Kami sampaikan kepada Bapak Rektor Universitas Udayana atas
biaya penelitian yang diberikan melalui skim penelitian Unggulan Udayana dengan Surat
Penugasan Penelitian Nomor 246-326/UN14.2/PNL.01.03.00/2015 Tanggal 21 April 2015.
DAFTAR PUSTAKA
Avise JC. 1994. Molecular Markers, Natural History, and Evolution. Chapman and Hall Inc. New York.
Bachtrog D &Charlesworth B. 2001.Towards a complete sequence of the human Y chromosome.
Genome Biology 2001, 2(5):reviews1016.1–1016.5
Bennett BT, Abee CR, Hendrickson R. 1995. Nonhuman Primates in Biomedical Research. Biology and Management. Academic Press Inc. San Diego, California.
Drobnic K. 2006. A new primer set in a SRY gene for sex identification. International Congress Series 1288 (2006) 268–270.
Erdal ME & Barlas IO. 2000. Detection of the SRY Gene in a 46,XY Phenotypic Female by the PCR-SSCP Method. Turk J Med Sci 30 (2000) 501-503.
Eudey AA. 1980. Pleistocene glacial phenomena and the evolution of Asian macaques. In The
Macaques. Studies in Ecology, Behavior and Evolution. Edited by D.G. Lindburg. :52-83.
Excoffier L, Laval G, and Schneider S. 2005. Arlequin ver. 3.0: An integrated software package for population genetics data analysis. Evolutionary Bioinformatics Online 1:47-50.
Brotcorne F, Fuentes A, Wandia I N, Roseline C, Beudels-Jamar, Huynen M-C. Changes in Activity Patterns and Intergroup Relationships After a Significant Mortality Event in Commensal Long-Tailed Macaques (Macaca Fascicularis) in Bali Indonesia. Int J Primatol
(2015) 36:548 566. DOI 10.1007/s10764-015-9841-5
Brotcorne F, Maslarov C, Wandia I N, Fuentes A,. Beudels‐ Jamar RC, and Huynen M-C. 2014. The Role of Anthropic, Ecological, and Social Factors in Sleeping Site Choice by Long‐Tailed Macaques (Macaca fascicularis). American Journal of Primatology. DOI: 10.1002/ajp.22299. Published online XX Month Year in Wiley Online Library (wileyonlinelibrary.com).
Frankham R, Ballou JD, Briscoe DA. 2004. A Primier of Conservation Genetics. Cambridge University Press. Cambridge
Fooden J. 1995. FIELDIANA. Zoology. New Series No. 81. Systematic Review of Southeast Asian Longtail Macaques, Macaca fascicularis (Raffles, [1821]). Published by Field Museum of Natural History. USA.
Hartl DL & Clark AG. 1997. Principles of Population Genetics. Third ed. Snauer Associates, Inc. Publishers. Sunderland, Massachusetts
Jolly A. 1985. The Evolution of Primate Behavior. 2nd Ed. Macmillan Publishing Company. New York.
Kawamoto Y, Ischak TM, Supriatna J. 1984. Genetic variation within and between troops of the crab-eating macaque (Macaca fascicularis) on Sumatra, Jawa, Bali, Lombok and Sumbawa, Indonesia. Primates, 25(2):131-159.
DNA and Geographical Distribution. PLoS ONE 8(3): e59660. doi:10.1371/journal.pone.0059660.
Klug WS & Cummings MR. 2005. Essentials of Genetics. International Ed. 5th Ed. Pearson Education, Inc. USA.
Mitchell G & Erwin J. 1986. Behavior, Cognition, and Motivation. Comparative Primate Biology. Volume 2, Part A. Alan R. Liss, Inc., New York.
Napier JR & Napier PH. 1985. The Natural History of the Primates. The British Museum (Natural History). Cromwell, London.
Nozawa K, Shotake T, Minezawa M, Kawamoto Y, Kawamoto K, Kawamoto S. 1996. Population-genetic studies of the Javanese macaque, Macaca fuscata. In: Variations in the Asian
Macaques. T. Shotake and K. Wada (eds.). Tokai University Press. Tokyo, Japan.: 1-36
Qiagen 2007. QIAamp DNA Mini and Blood Mini Hanbook. 2nd Eds. November 2007. Pp 27-29. Rovie-Ryan JJ, Abdullah MT, Sitam FT, Abidin ZZ, and Tan SG. 2013.
Y-chromosomal gene flow of Macaca fascicularis (Cercopithecidae) between the insular and mainland peninsula of Penang state, Malaysia. Journal of Science and Technology in the Tropics (2013) 9: 113-126.
Rowe N. 1996. The Pictorial Guide to the Living Primates. Pogonias Press. New York.
Rozas, J dan Librado P. Librado (2009). DnaSP v5: a Software for comprehensive analysis for DNA polymorphism data. Bioimformatics 25: 1451-1452.
Tamura K, Stecher G, Peterson D, Filipski A, and Kumar S (2013) MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Molecular Biology and Evolution:30 2725-2729.
Wandia I N. 2003. Mikrosatelit sebagai penanda molekul untuk mengukur polimorfisme genetik monyet ekor panjang di Sangeh, Bali. J. Vet. 4(3):93-100.
Wandia I N, Supraptini-Mansjoer S, Suryobroto B. 2004. Polimorfisme genetik monyet ekor panjang di daerah pariwisata Uluwatu, Bali. J. Vet. 5(2):67-76.
Wandia I N. 2007. Struktur dan Keragaman Genetik Populasi Lokal Monyet Ekor Panjang (Macaca fascicularis) di Tawa Timur, Bali, dan Lombok. Disertasi. Sekolah Pascasarjana IPB, Bogor. Wandia I N, Arta Putra I GA, Soma I G. 2014. Polymorphism of Microsatellite Loci on Y
Chromosome in Long-Tailed Macaque Populations in Bali Island, Indonesia. Jurnal Ilmu