SKRIPSI
OLEH :
FRISYLIA NAINGGOLAN 160301222
AGROTEKNOLOGI – PEMULIAAN TANAMAN
PROGRAM STUDI AGROTEKNOLOGI FAKULTAS PERTANIAN
UNIVERSITAS SUMATERA UTARA 2021
i SKRIPSI
OLEH :
FRISYLIA NAINGGOLAN 160301222
AGROTEKNOLOGI – PEMULIAAN TANAMAN
Skripsi Sebagai Salah Satu Syarat untuk dapat Memperoleh Gelar Sarjana di Program Studi Agroteknologi Fakultas Pertanian
Universitas Sumatera Utara
PROGRAM STUDI AGROTEKNOLOGI FAKULTAS PERTANIAN
UNIVERSITAS SUMATERA UTARA 2021
ii Nama : Frisylia Nainggolan
NIM : 160301222
Program Studi : Agroteknologi Minat : Pemuliaan Tanaman
Diketahui Oleh, Komisi Pembimbing
Prof. Dr. Ir. Rosmayati, MS Ir. Revandy I.M Damanik, M.Si., M.Sc., Ph.D Ketua Anggota
Diketahui Oleh :
Dr. Nini Rahmawati SP., M.Si.
Ketua Program Studi Agroteknologi
i
Varietas Kedelai (Glycine max (L.) Merill) terhadap Kondisi Tergenang pada Umur 8 MST. dibimbing oleh Prof. Dr. Ir. Rosmayati, MS dan Ir. Revandy Iskandar M.
Damanik, M.Si., M.Sc., Ph.D. Penelitian ini dilaksanakan di Lahan sawah irigas i Sememe Kecamatan Deli Tua, Sumatera Utara, Medan dan Laboratorium Kultur Jaringan Universitas Sumatera Utara, Medan pada bulan Juni 2020 hingga April 2021, menggunakan Rancangan Acak Kelompok (RAK) satu faktor yaitu: varietas (Anjasmoro ,Grobogan, Deja 1 dan Devon 1). Penelitian ini bertujuan untuk mengetahui karakteristik fisiologis beberapa varietas kedelai (Glycine max (L.) Merill) pada umur 8 MST saat kondisi tergenang. Parameter yang diamati adalah total klorofil, enzim peroksidase, dan kandungan prolin. Varietas Anjasmoro memiliki kandungan enzim peroksidase dan kandungan prolin yang lebih tinggi saat kondisi tergenang, dibandingkan dengan varietas Grobogan, varietas Deja 1 dan varietas Devon 1. Varietas Devon 1 memiliki kandungan total klorofil yang lebih tinggi saat kondisi tergenang dibandingkan dengan varietas Anjasmoro, varietas Grobogan dan varietas Deja 1.Varietas Anjasmoro mampu beradaptasi pada keadaan tergenang, karena memiliki kandungan POD yang lebih tinggi dibandingkan tiga varietas yg lain. Empat varietas yang digenangi selama 96 jam memberikan pengaruh yang nyata terhadap sifat fisiologis pada kandungan klorofil, enzim peroksidase, dan kandungan prolin, pada umur 8 MST.
Kata kunci : genangan, generatif, karakter fisiologis, kedelai, varietas
ii
of Soybean (Glycine Max (L.) Merill) Varieties at 8 WAP Against Inundation Conditions guiding by Prof. Dr. Ir. Rosmayati, and MS Ir. Revandy I.M Damanik, M.Si., M.Sc., Ph.D,. This research was conducted in Sememe irrigated rice fields Deli Tua District, North Sumatra, Medan and Laboratory of Tissue Culture, Universitas Sumatera Utara, Medan from June 2020 to April 2021, using a non- factorial Randomized Blok Design (RAD), namely: varieties (Anjasmoro, Grobogan, Deja 1 and Devon 1). This study aims to determine the physiological characters of several varieties of soybean (Glycine max (L.) Merill) in growth phase 8 weeks after planting to inundation conditions. The parameters observed were total chlorophyll, peroxidase enzymes and proline. The Anjasmoro variety contains peroxidase enzyme and higher proline content in inundation conditions than the other 3 varieties. The Devon 1 variety had a higher total chlorophyll content and total protein content than the other 3 varieties in inundation conditions. The Anjasmoro variety is able to adapt to flooding conditions, because it has a higher POD content than the other three varieties.Four varieties that were flooded for 96 hours had a significant effect on the physiological properties of the chlorophyll content, peroxidase enzyme, and proline content, in the generative phase.
Keywords: generative, Inundation soybean, physiological character, variety
.
iii
Ayahanda Alm. Karianto Nainggolan dan Ibunda Lementeria Silitonga. Penulis merupakan anak keempat dari lima bersaudara.
Pendidikan formal yang pernah ditempuh adalah SD Negeri 173165 Sipahutar lulus pada tahun 2010, SMP Negeri 1 Sipahutar lulus tahun 2013 dan tahun 2016 penulis lulus dari SMA Negeri 2 Balige dan pada tahun yang sama lulus seleksi penerimaan mahasiswa baru melalui jalur UMB-PT pada program studi Agroekoteknologi, Fakultas Pertanian Universitas Sumatera Utara, Medan.
Selama mengikuti perkuliahan, penulis menjadi anggota di Paduan Suara Transeamus FP USU. Penulis juga menjadi anggota organisasi kemahasiswaa n yaitu Himagrotek Fakultas Pertanian Universitas Sumatera Utara, Medan.
Penulis melaksanakan praktek kerja lapangan (PKL) di PTPN III Kebun Aek Nabara Selatan Kabupaten Labuhan Batu Selatan Provinsi Sumatera Utara pada bulan Juli-Agustus 2019 dan melaksanakan KKN di Desa Brohol, Kec. Sei Suka, Batu Bara Provinsi Sumatera Utara pada bulan Juli-Agustus 2019.
iv
dan karunia-Nya sehingga penulis dapat menyelesaikan skripsi ini.
Adapun judul dari skripsi ini adalah “ Respon Karakter Fisiologi pada Beberapa Varietas Kedelai (Glycine max (L.) Merill) terhadap Kondis i Tergenang pada Umur 8 MST ” yang merupakan salah satu syarat untuk mendapat gelar sarjana di Fakultas Pertanian Universitas Sumatera Utara, Medan dan sebagai bahan informasi bagi pihak yang membutuhkan.
Penulis menyadari bahwa banyak pihak yang telah membantu penulis selama penyusunan skripsi ini. Oleh karena itu, pada kesempatan ini penulis ingin mengucapkan terima kasih kepada, keluarga tercinta, Alm. Bapak, Mama, Abang Gene, Kakak Gene, Abang Anda, Kakak Duma, Abang Andi, Adel, Genesis, serta semua keluarga. Terima kasih atas doa, kasih, cinta, semangat, motivasi yang selalu diberikan.
Ibu Prof. Dr. Ir. Rosmayati, MS selaku ketua komisi pembimb ing.
Bapak Ir. Revandy I.M Damanik, M.Si., M.Sc., Ph.D selaku anggota komisi pembimbing.yang telah memberikan saran, koreksi, dukungan dan motivasi selama penyusunan skripsi ini.
Ibu Dr. Nini Rahmawati SP., M.Si, beserta Ibu Dr. Diana Sofiah Hanafia h SP., MP., yang sudah bersedia menjadi penguji penulis dalam pelaksanaan sidang meja hijau. Semua tenaga pendidik dan karyawan Program Studi Agroteknolo gi, terimakasih untuk motivasi, serta bantuan administrasi yang telah diberikan selama perkuliahan dan penyusunan skripsi ini.
v
diberikan, untuk sahabat terkasih Kak Anggi, Kak Rences, Adriana, Tiur, Audrin, Fio, terima kasih untuk doa dan dukungannya, untuk Tim Genangan, Dina, Tara, Ester yang menjadi teman berjuang selama penelitian. Kepada abang, kakak, teman- teman, adik-adik Paduan Suara Transeamus penulis sangat berterimakasih untuk semangat, doa dan kebersamaan kekeluargaan yang diberikan. Ucapan terima kasih untuk seluruh teman teman seangkatan 2016, abang, kakak, serta adik - adik yang berada di lingkungan Program Studi Agroteknologi beserta semua yang terlibat di dalam proses penyelesaian pembuatan dan penulisan skripsi ini.
Terima kasih kepada EXO, NCT U, NCT 127, NCT Dream, WayV, Jerome Polin beserta Waseda Boys, yang memberikan motivasi, dukungan, dan menjadi salah satu alasan penulis untuk selalu semangat selama penulis menyelesa ika n skripsi ini, serta kepada seluruh pihak yang telah bertanya “ kapan lulus?”, “kapan sidang”, dan sejenisnya, yang menjadi salah satu alasan penulis untuk segera menyelesaikan skripsi ini.
Penulis berharap semoga skripsi ini bermanfaat bagi pihak yang membutuhkan.
Medan, Desember 2021
Penulis
vi
ABSTRACT ... ii
RIWAYAT HIDUP ... iii
KATA PENGAN TAR ... iv
DAFTAR ISI ... vi
DAFTAR TABEL... viii
DAFTAR GAMBAR ... ix
DAFTAR LAMPIRAN ... x
PENDAHULUAN Latar Belakang ... 1
Tujuan Penelitian... 3
Hipotesa Penelitian... 3
Kegunaan Penulisan ... 3
TINJAUAN PUSTAKA Botani Tanaman Kedelai (Glycine max (L.) Merill) ... 4
Syarat Tumbuh Tanaman Kedelai (Glycine max (L.) Merill) ... 6
Iklim ... 6
Tanah ... 7
Fase Pertumbuhan Kedelai ... 7
Varietas... 9
Cekaman Genangan Terhadap Pertumbuhan Kedelai... 10
Klorofil ... 13
Enzim Peroksidase Dismutase (POD)... 14
Prolin ... 15
BAHAN DAN METODE Tempat dan Waktu Penelitian ... 16
Bahan dan Alat Penelitian ... 16
Metode Penelitian ... 17
PELAKSANAAN PENELITIAN Persiapan Lahan ... 18
Penanaman ... 18
Pemupukan ... 18
Pemeliharaan Tanaman ... 18
Penyulaman ... 18
Penyiangan ... 19
vii
Uji K lorofil ... 20
Pengukuran Total Protein ... 20
Analisis Enzim Peroksida Dismute ... 21
Uji Proline ... 21
HASIL DAN PEMBAHASAN Hasil ... 23
Analisis Total K lorofil ... 23
Analisis Enzim Peroksida Dismute (POD) ... 25
Analisis Proline ... 27
KESIMPULAN DAN SARAN Kesimpulan ... 29
Saran ... 29 DAFTAR PUSTAKA
LAMPIRAN
viii
No. Hal
1. Karakteristik fase tumbuh vegetatif pada tanaman kedelai... 8
2. Karakteristik fase tumbuh generatif pada tanaman kedelai ... 8
3. Rataan Total K lorofil ... 23
4. Rataan Prolin ... 27
ix
1. Tanaman Kedelai yang digenangi ... 20 2. Histogram Enzim Peroksidase ... 25
x
1. Deskripsi Varietas Anjasmoro ... 36
2. Deskripsi Varietas Grobogan ... 37
3. Deskripsi Varietas Deja 1 ... 38
4. Deskripsi Varietas Devon 1 ... 39
5. Bagan Penelitian di Lapangan ... 40
6. Jadwal Kegiatan Pelaksanaan Penelitian ... 41
7. Data Analisis Klorofil ... 42
8. Sidik ragam analisis klorofil... 42
9. Data Analisis Prolin ... 42
10. Sidik ragam Analisis Prolin ... 42
11. Data Analisis Peroksida Oksidase (POD) ... 43
12. Data Analisis Klorofil... 43
PENDAHULUAN Latar Belakang
Kedelai merupakan salah satu komoditi pangan yang utama dalam upaya pemerintah untuk mencapai swasembada pangan. Dikarenakan kedelai sangat diperlukan sebagai sumber pangan murah dan bergizi, pakan ternak serta bahan baku industri. Berkaitan dengan hal tersebut kebutuhan terhadap komoditi kedelai terus meningkat dari tahun ketahun sejalan dengan meningkatnya laju pertumbuha n
penduduk dan meningkatnya kesadaran masyarakat akan gizi makanan ( Kuncahyo, 2018 )
Upaya untuk meningkatkan produksi kedelai nasional dapat ditempuh dengan tiga pendekatan yaitu: 1) peningkatan produktivitas, 2) peningkatan intensitas tanam, dan 3) perluasan areal tanam. Produktivitas kedelai nasional saat ini masih rendah yaitu rata-rata 1,29 ton/ha ( Rachman et al, 2016 ). Peningkatan produksi kedelai dalam negeri dapat dilakukan dengan cara ekstensifikasi dan intesifikas i.
Intensifikasi adalah usaha meningkatkan hasil pertanian dengan teknologi yang tepat seperti penggunaan kultivar unggul, sedangkan ekstensifikasi atau perluasan areal
pada berbagai agroekosistem memerlukan kultivar yang beragam ( Dwiputra et al, 2015 ).
Adanya Inovasi pengembangan kedelai toleran sangat berpotensi dalam meningkatkan jumlah produksi kedelai. Selain itu, genangan tidak hanya bermanfaat bagi pengembangan kedelai di lahan sawah, tetapi juga prospektif bagi wilayah yang sering mengalami cekaman genangan seperti lahan pasang surut untuk menunja ng hasil produksi pasokan kedelai untuk memenuhi kebutuhan sehari-hari.
(Suriadikarta dan Sutriadi, 2007).
Genangan merupakan masalah utama dibanyak daerah pertanian di dunia dan kedelai, merupakan tanaman yang peka terhadap genangan. Di Indonesia, kedelai dapat diusahakan di lahan sawah setelah padi. Kondisi tanah yang tergenang (jenuh air) akibat air sisa penanaman padi atau air hujan sering menjadi salah satu penyebab rendahnya produktivitas kedelai di lahan sawah. Genangan atau kondisi jenuh air disebabkan oleh kandungan lengas tanah yang berada di atas kapasitas lapang (Rohmah dan Saputro, 2016).
Kondisi genangan merupakan kondisi rendahnya kadar oksigen dari batas normal yang terjadi di akibat presipitasi tinggi, kualitas tanah yang buruk, dan over- irigasi diikuti oleh drainase yang lambat. Beberapa jenis tumbuhan memilik i kapasitas yang luar biasa untuk bertahan dalam kondisi oksigen rendah, meski terjadi perubahan dalam sifat morfologi dan fisiologis pada tingkat antar dan intraspesifik, memiliki dampak yang kuat pada kelimpahan spesies dan distribusi ekosistem di Indonesia yang rawan banjir (Pompeianoa, et al., 2017).
Menurut Hapsari dan Adie (2010) genangan menyebabkan tanaman mengalami penuaan dini daun mengalami klorosis, nekrosis, dan gugur serta pertum- buhan terhambat, dan akhirnya menurunkan hasil. Umumnya kehilangan hasil pada fase vegetatif relatif lebih kecil dibandingkan pada fase reproduktif,yaitu 1743%
pada fase vegetatif dan 5056% pada fase reproduktif. Besar-nya penurunan hasil bergantung pada varietas yang digunakan, fase pertumbuhan tanaman, lamanya tergenang,tekstur tanah, dan kehadiran penyakit.
Menurut Linkemer et al., 1998, tanaman kedelai akan mengala mi kehilangan hasil sebanyak 93% apabila mengalami penggenangan selama 7 hari pada fase perkembangan R3 (tanaman mulai membentuk polong), demikian juga
pada stadia perkembangan R1 (tanaman mulai berbunga) dan R5 (tanaman mula i membentuk biji) mencapai 63% kehilangan hasil. Sedangkan pada fase pertumbuhan V2 (tanaman membentuk buku kedua) dapat mengurangi hasil sebanyak 30%.
Hasil penelitian ini diharapkan dapat digunakan untuk memberikan informas i kondisi fisiologis beberapa varietas tanaman kedelai pada kondisi tergenang. Dimana informasi ini penting untuk pengembangan varietas kedelai tahan genangan.
Berdasarkan uraian diatas maka penulis tertarik untuk melakukan penelit ia n yaitu respon karakter fisiologi beberapa varietas Kedelai (Glycine Max (L.) Merril) terhadap penggenangan pada umur 8 minggu setelah tanam (MST).
Tujuan Penelitian
Untuk mengetahui karakter fisiologi beberapa varietas kedelai (Glycine max (L.) Merill) saat kondisi tergenang pada umur 8 minggu setelah tanam
(MST)
Hipotesa Penelitian
Ada pengaruh beberapa varietas kedelai (Glycine max (L.) Merill) kondisi tergenang pada umur 8 minggu setelah tanam (MST) terhadap karakter fisiolo gis tanaman.
Kegunaan Penulisan
Sebagai salah satu syarat untuk mendapatkan gelar sarjana di Program Studi Agroteknologi Fakultas Pertanian Universitas Sumatera Utara, Medan dan sebagai bahan informasi bagi pihak yang membutuhkan.
TINJAUAN PUSTAKA Botani Tanaman
Kedelai dikenal dengan beberapa nama, yaitu Glycine soja atau Soja max.
Tahun 1984 telah disepakati bahwa nama botani yang dapat diterima dalam istila h ilmiah yaitu Glycine max (L.) Merril. Klasifikasi tanaman kedelai sebagai berikut : Kingdom: Plantae Sub-kingdom: Tracheobionta Super Divisi: Spermatophyta Divisi: Magnoliophyta Kelas: Magnoliopsida Sub Kelas: Rosidae Ordo: Fabales
Famili: Fabaceae Genus: Glycine Spesies: Glycine max (L.) Merr.
(Adisarwanto, 2005).
Sistem perakaran tanaman kedelai terdiri dari akar tunggang, akar sekunder (serabut) yang tumbuh dari akar tunggang, serta akar cabang yang tumbuh dari akar sekunder. Akar tunggang merupakan perkembangan dari akar radikal yang sudah mulai muncul sejak masa perkecambahan. Akar ini mempunyai akar-akar cabang yang lurus. Akar serabut merupakan akar yang tumbuh ke bawah sepanjang 20 cm.
Tanaman ini juga memiliki akar-akar lateral (cabang) yang tumbuh ke samping sepanjang 5-25 cm. Pada akar lateral terdapat akar serabut, fungsinya untuk menghisap air dan unsur hara, pada akar ini juga terdapat bintil akar (nodule) yang mengandung bakteri Rhizobium, kegunaannya sebagai pengikat zat nitrogen dari udara ( Departemen Pertanian, 2006 ).
Kedelai berbatang semak, dengan tinggi batang antara 30 - 100 cm. Setiap batang dapat bentuk 3 - 6 cabang sedangkan apabila jarak antara tanaman dalam barisan rapat, cabang menjadi berkurang atau tidak bercabang sama sekali. Batang dapat dibedakan menjadi dua yaitu bagian batang di bawah keping biji yang belum lepas disebut hypocotyl, sedangkan bagian di atas keping biji disebut epycotyl.
Batang kedelai tersebut berwarna ungu atau hijau. Tipe pertumbuhan dapat dibedakan menjadi 3 macam yakni tipe ujung batang melilit (indeterminate), tipe batang tegak (determinate), dan tipe semi determinit (Suprapto, 1991).
Daun kedelai ada dua bentuk, yaitu bulat (oval) dan lancip (lanceolat e).
Bentuk daun tersebut dipengaruhi oleh faktor genetik. Daerah yang mempunya i tingkat kesuburan tanah yang tinggi sangat cocok untuk varietas kedelai yang mempunyai bentuk daun yang lebar. Daun mempunyai stomata yang berjumla h antara 190-320 buah/m2 (Adisarwanto, 2005).
Tanaman kedelai memiliki bunga sempurna, yaitu dalam satu bunga terdapat alat kelamin jantan (benang sari) dan alat kelamin betina (putik). Warna bunga putih bersih atau ungu muda. Bunga tumbuh pada ketiak daun dan biasanya terdapat 3 – 15 kuntum bunga, namun sebagian besar bunga rontok, hanya beberapa dapat membentuk polong (Sugeng, 2000).
Penyerbukan bunga terjadi pada saat bunga masih tertutup sehingga kemungkinan penyerbukan silang sangat kecil, yaitu hanya 0,1%, warna bunga kedelai ada yang ungu dan putih. Potensi jumlah bunga yang terbentuk bervariasi, tergantung dari varietas kedelai, tetapi umumnya berkisar antara 40–200 bunga pertanaman. Hanya saja, umumnya di tengah masa pertumbuhannya, tanaman kedelai kerap kali mengalami kerontokan bunga hal ini masi di kategorikan wajar bila kerontokan yang terjadi berada pada kisaran 20–40% (Adisarwanto, 2008).
Polong pertama kali tampak sekitar 7-10 hari setelah munculnya bunga pertama. Pembentukan polong berlanjut sama cepatnya seperti pembentukan bunga dan dalam keadaan normal memakan waktu kurang lebih 21 hari. Panjang
polong muda sekitar 1 cm. Jumlah polong yang terbentuk pada setiap ketiak tangkai daun sangat beragam antara 1-10 buah dalam setiap kelompok (Hidayat, 1985) Syarat Tumbuh
Iklim
Pengembangan kedelai dapat dilakukan di lahan sawah maupun dilahan kering, bergantung kepada iklim dan kebutuhan petani setempat. Tanaman Kedelai dapat tumbuh pada berbagai jenis tanah asal drainase (tata air)danaerasi(tata udara) tanah cukup baik, curah hujan 100-400 mm/bulan,suhu udara 23-30°C,kelembaban 60-70%, pH tanah 5,8-7 dan ketinggian kurang dari 600 m dpl ( BPTP, 2009 )
Tanaman kedelai sebagian besar tumbuh di daerah yang ber iklim tropis dan subtropis. Sebagai barometer iklim yang cocok bagi kedelai adalah bila cocok bagi tanaman jagung.Bahkan daya tahan kedela i lebih baik daripada jagung.Iklim kering lebih disukai tanaman kedelai dibandingkan iklim lembab.Tanaman kedelai dapat tumbuh baikdi daerah yang memiliki curah hujan sekitar 100-400 mm/bula n.
Sedangkan untuk mendapatkan hasil optimal, tanaman kedelai membutuhkan curah hujan antara100-200 mm/bulan. Suhu yang dikehendaki tanaman kedelai antara 21- 34ᵒC, akan tetapi suhu optimum bagi pertumbuhan tanaman kedelai 23-27ᵒC. Pada proses perkecambahan benih kedelai memerlukan suhu yang cocok sekitar 30ᵒC. Saat panen kedelai yang jatuh pada musim kemarau akan lebih baik dari pada musim hujan, karena berpengaruh terhadap waktu pemasakan biji dan pengeringan hasil (Irwan, 2006).
Kedelai dapat tumbuh baik di tempat pada daerah berhawa panas, di tempat terbuka dengan curah hujan 100–400 mm3 per bulan. Oleh karena itu, kedelai kebanyakan ditanam di daerah yang terletak kurang dari 400 m di atas permukaan
laut. Jadi tanaman kedelai akan tumbuh baik, jika ditanam di daerah beriklim kering (Andrianto dan Indarto, 2004).
Tanah
Kedelai umumnya dapat beradaptasi terhadap berbagai jenis tanah, dan menyukai tanah yang bertekstur ringan hingga sedang, dan berdrainase baik, akan tetapi peka terhadap salinitas (Rubatzky dan Yamaguchi, 1998).
Tanaman kedelai menghendaki tanah yang subur, gembur, dan kaya akan humus atau bahan organik. Nilai pH ideal bagi pertumbuhan kedelai dan bakteri Rhizobium adalah 6,0-6,8. Apabila pH diatas 7,0 tanaman kedelai akan mengala m i
klorosis sehingga tanaman menjadi kerdil dan daunnya menguning (Waisimon, 2012).
Fase Pertumbuhan Kedelai
Stadia pertumbuhan kedelai secara garis besar dapat dibedakan atas pertumbuhan vegetatif dan generatif. Stadia pertumbuhan vegetatif dihitung sejak tanaman mulai muncul ke permukaan tanah sampai saat mulai berbunga. Stadia perkecambahan dicirikan dengan adanya kotiledon, sedangkan penandaan stadia pertumbuhan vegetatif dihitung dari jumlah buku yang terbentuk pada batang utama. Menurut Kadarwati (2006), nitrogen merupakan unsur hara makro yang paling banyak dibutuhkan tanaman, unsur nitrogen sangat berperan dalam fase vegetatif tanaman. Stadia vegetatif umumnya dimulai pada buku ketiga. Tanda V dimaksudkan untuk menandakan stadia vegetatif yag diikuti oleh angka unt uk menunjukkan jumlah buku (Tabel 1)
Stadia pertumbuhan reproduktif (generatif) dihitung sejak tanaman kedelai mulai berbunga sampai pembentukan polong, perkembangan biji dan pemasakan
biji. Pada fase ini sangat memerlukan unsur P dan K dalam jumlah yang lebih banyak (Kadarwati, 2006), Penandaan setiap stadia pada periode generatif yaitu tanda R (reproduktif) dan diikuti dengan penulisan angka 1-8. ( Tabel 2 )
Tabel 1. Penandaan stadia pertumbuhan vegetatif kedelai.
Sumber: Sumarno dkk., (1999)
Tabel 2. Penandaan stadia pertumbuhan reproduktif tanaman kedelai.
Singkatan Stadia Tingkatan Stadia Keterangan
R1 Mulai berbunga Munculnya bunga pertama pada buku mana pun pada batang R2 Berbunga penuh Bunga terbuka penuh pada satu
atau dua buku paling atas pada batang utama dengan daun yang
telah terbuka
R3 Mulai berpolong Polong telah terbentuk dengan panjang 0,5 cm pada salah satu
buku batang utama R4 Berpolong penuh Polong telah mempunyai
panjang 2 cm pada salah satu buku teratas pada batang utama
R5 Mulai pembentukan
biji
Ukuran biji dalam polong mencapai 3 mm pada salah satu
buku batang utama Singkatan Stadia Tingkatan Stadia Keterangan
VE Stadia Pemunculan Kotiledon muncul ke permukaan tanah
VC Stadia Kotiledon Daun unifololioliate
berkembang
V1 Stadia Buku Pertama Daun terbuka penuh pada buku unifololioliate
V2 Stadia Buku Kedua
Daun trifololioliate terbuka penuh pada buku kedua diatas
buku unifololioliate
V3 Stadia Buku Ketiga
Pada buku ketiga, batang utama terdapat daun yang
terbuka penuh
Vn Stadia Buku ke – n
Pada buku ke-n, batang utama telah terdapat daun yang
terbuka penuh
R6 Berbiji penuh Setiap polong pada batang utama telah berisi biji satu atau
dua
R7 Mulai Masak Salah satu warna polong pada batang utama telah berubah menjadi cokelat kekuningan atau
warna masak
R8 Masak Penuh 95% jumlah polong telah
mencapai warna masak Sumber: Fehr dkk., 1971
Varietas
Tiga varietas kedelai sudah ditanam di Indonesia pada mulanya berasal dari diantaranya Jepang, Taiwan, Amerika Serikat, dan sebagainya. Kriteria varietas unggul kedelai adalah berproduksi tinggi, berumur pendek, tahan (resisten) terhadap penyakit berbahaya, mempunyai daya adaptasi luas terhadap berbagai keadaan lingkungan tumbuh (Hanum, 2008).
Varietas unggul yang memiliki produktivitas tinggi dan mempunyai sifat ketahanan terhadap cekaman biotik dan abiotik serta karakteristik yang sesuai dengan permintaan pasar merupakan modal utama dalam upaya meningkatkan produksi dan pendapatan petani. Hasil penelitian Puslitbangtan untuk lahan sawah dianjurka n penggunaan varietas berumur sedang (85-90 hari) seperti Willis, Kerinci, Tampomas, Krakatau dan Jayawijaya (Atman, 2006).
Varietas merupakan salah satu teknologi utama yang mampu meningka tka n produktivitas kedelai dan pendapatan petani. Tersedianya beberapa varietas kedelai, kini petani dapat memilih varietas yang sesuai dengan kondisi lingkungan setempat, berdaya hasil dan bernilai jual tinggi. Oleh karena itu uji adaptasi varietas di suatu tempat perlu terus dilakukan oleh instansi terkait dalam upaya mendapatkan varietas yang sesuai di suatu tempat (Yusuf dan Harnowo, 2012).
Varietas Anjasmoro memiliki potensi hasil 2,25 ton ha-1, tahan rebah, polong tidak mudah rebah pecah, agak tahan terhadap penyakit karat daun, ukuran biji besar (16 g/100 biji),umur panen 83-93 hari, dengan kemampuan adaptasi yang tinggi, varietas Anjasmoro juga biasanya ditanaman di lahan sawah. Varietas Grobogan mempunyai keunggulan yakni umur pendek (76 hari), ukuran polong besar, produksi tinggi, kandungan protein tinggi yakni mencapai 43,9 persen dan daun rontok saat jelang panen (Balai Penelitian Tanaman Aneka Kacang dan Umbi, 2005).
Kedelai varietas Devon-1 dilepas pada tahun 2015. Umur matang Devon-1 83 hari, tinggi batang 58 cm, batang agak tahan rebah, dan umur berbunga 34 hari, dan potensi hasil dari varietas Devon-1 yaitu 3,09 ton/ha. Deja 1 memiliki umur masak genjah (79 hari), berukuran biji sedang (12,9 g/100 biji), agak tahan hama ulat grayak, tahan hama penggerek polong dan pengisap polong, serta agak tahan penyakit karat daun, dengan kandungan protein 39,6% dan lemak 17,3%. Varietas Deja 1 berpotensi hasil tinggi dan toleran terhadap cekaman jenuh air mulai umur 14 hari (fase V2) hingga fase masak (fase R7). Pada kondisi tercekam kondisi tanah jenuh air, DEJA 1 mampu memberikan hasil biji rata-rata 2,39 t/ha dan 2,38 t/ha, dengan potensi hasil masing- masing 2,87 t/ha dan 2,75 t/ha (Balitkabi, 2016).
Cekaman Genangan Terhadap Pertumbuhan Kedelai
Genangan dapat terjadi pada lahan asah alami maupun lahan basah buatan.
Notohadiprawiro (1989) mendeskripsikan lahan basah alami sebagai lahan yang karena drainase yang buruk, bersifat basah sementara atau sepanjang waktu. Keadaan ini terjadi karena iklim basah dan berkaitan dengan kedudukan lahan yang berenergi potensial rendah (daerah berketinggian rendah) atau karena bentuk lahan yang berupa cekungan tambat (retention basin). Lahan basah buatan yakni lahan yang bentuknya
sengaja dibuat sedemikian rupa sehingga dapat menambat banyak air untuk membuat tanah jenuh air atau mempertahankan genangan air pada permukaan tanah selama waktu tertentu.
VanToai et al. (2001) membagi genangan berdasarkan kondisi pertanaman menjadi dua, yaitu: 1) kondisi jenuh air (waterlogging) di mana hanya akar tanaman yang tergenang air, dan 2) kondisi bagian tanaman sepenuhnya tergenang air (complete submergence). Kekurangan oksigen dalam tanah akibat genangan merupakan faktor pembatas pertumbuhan dan produktivitas tanaman. Kekurangan oksigen menggeser metabolisme energi dari aerob menjadi anaerob sehingga berpengaruh kurang baik terhadap serapan nutrisi dan air. Akibatnya, tanaman menunjukkan gejala kelayuan walaupun tersedia banyak air (Sairam et al. 2009).
Genangan dapat menurunkan pertukaran gas dalam tanah dan di udara sehingga mengurangi ketersediaan O2 bagi akar dan menghambat pasokan O2 bagi akar dan mikroorganisme (Riche 2004). Pada kondisi tergenang, volume pori tanah yang berisi udara kurang dari 10% sehingga menghambat pertumbuhan akar.
Kedelai merespons kondisi genangan dengan mengaktifkan metabolisme atau melakukan pemulihan secara cepat setelah terjadi cekaman genangan diikuti dengan aklimatisasi (penyesuaian diri). Waktu yang dibutuhkan untuk pemulihan lebih cepat
2−4 hari pada kedelai yang toleran dibandingkan dengan yang peka (Hapsari dan Addie, 2010).
Tanaman yang tergenang dalam waktu singkat akan mengalami kondisi hipoksia (kekurangan O2). Hipoksia biasanya terjadi jika hanya bagian akar tanaman yang tergenang (bagian tajuk tidak tergenang) atau tanaman tergenang dalam periode yang panjang tetapi akar berada dekat permukaan tanah. Jika tanaman tergenang
seluruhnya, akar tanaman berada jauh di dalam permukaan tanah dan mengala mi penggenangan lebih panjang sehingga tanaman berada pada kondisi anoksia (keadaan lingkungan tanpa O2). Kondisi anoksia tercapai 68 jam setelah penggenangan, karena O2 terdesak oleh air dan sisa O2 dimanfaatkan oleh mikroorganisme. Pada kondisi tergenang, kandungan O2 yang tersisa dalam tanah lebih cepat habis bila terdapat tanaman karena laju difusi O2 di tanah basah 10.000 kali lebih lambat dibandingkan dengan di udara (Amstrong 1979 dalam Dennis et al.
2000). Kondisi hipoksia atau anoksia tidak hanya menghalangi fiksasi N, tetapi juga distribusi N dan mineral lain sehingga menghambat pertumbuhan akar dan nodulasi.
Akibat transportasi N dan mineral ke bagian tajuk tidak mencukupi, daun akan menguning yang akan diikuti oleh pengguguran daun. Scott et al. (1989) melaporkan, pengaruh penggenangan ditunjukkan oleh daun yang menguning, pengguguran daun pada buku terbawah, kerdil, serta berkurangnya berat kering dan hasil tanaman lebih lambat (Hapsari dan Addie, 2010).
Penggenangan yang dilakukan pada fase pembungaan hingga pengisia n polong, mengakibatkan kerusakan akar serta diikuti dengan kematian bintil akar, penggenangan pada fase ini akan meningkatkan gugurnya bunga dan polong muda (Whigham and Minor, 1978). Menurut Tampubolon et al ( 1989) Penggenangan pada umur 8-10 MST daun-daun akan menguning dan mulai gugur, sehingga mengga nggu proses fotosintesis.
Genangan berdasarkan kondisi pertanaman dibagi menjadi dua, yaitu: (1) Kondisi jenuh air (waterlogging) dimana hanya akar tanaman yang tergenang air, dan
(2) Kondisi bagian tanaman sepenuhnya tergenang air (complete submergenc e) ( Van Toai et al, 2001 ).
Berdasarkan hasil penelitian Fatimah dan Triono ( 2016 ) menyatakan bahwa perlakuan genangan terhadap tanaman Kedelai mengakibatkan berkurangnya produktifitas dari kedelai tersebut sekitar 90 – 95 %. Hal ini terjadi akibat kurangnya suply O2 pada akar tanaman kedelai yang mengalami perendaman.
Klorofil
Klorofil merupakan komponen kloroplas yang utama dan kandungan klorofil ini relatif berkorelasi positif dengan laju fotosintesis ( li, et.al, 2006). Sintesis klorofil dipengaruhi oleh berbagai faktor seperti cahaya, gula atau karbohidrat, air, temperatur, faktor genetik, unsur-unsur hara seperti N, Mg, Fe, Mn, Cu, Zn, S dan O (Hendriyani dan Setiari, 2009).
Selama genangan air terjadi, stomata daun menutup, terjadi degradasi klorofil, penuaan daun, dan menguningnya mengurangi kemampuan daun untuk menangkap cahaya dan pada akhirnya menyebabkan penurunan laju fotosintes is.
Tanaman dapat mempertahankan produksi energi sampai batas tertentu selama hipoksia yang disebabkan oleh genangan air, melalui glikolisis dan fermentas i etanol. Namun, durasi genangan air dan respirasi anaerob yang berkepanjangan pada akhirnya menyebabkan akumulasi metabolit beracun seperti asam laktat, etanol, dan aldehida, dikombinasikan dengan peningkatan spesies oksigen reaktif (ROS), terutama hidrogen peroksida, sehingga akhirnya menyebabkan kematian sel dan penuaan tanaman (Kuai, et. al., 2014)
Fotosintesis merupakan proses perubahan senyawa anorganik (CO2 dan H2O) menjadi senyawa organik (karbohidrat) dan O2 dengan bantuan cahaya matahari. Sifat kimia klorofil, antara lain (1) tidak larut dalam air, melainkan larut dalam pelarut organik yang lebih polar, seperti etanol dan kloroform; (2) inti Mg
akan tergeser oleh 2 atom H bila dalam suasana asam, sehingga membentuk suatu persenyawaan yang disebut feofitin yang berwarna coklat (Dwidjoseputro, 1994).
Enzim Peroksidase Dismutase (POD)
Enzim peroksidase merupakan salah satu enzim tanaman yang mempunya i hubungan dengan proses ketahanan. Untuk mengetahui kepekaan dan ketahanan tanaman terhadap serangan penyakit dipergunakan pendekatan mengenai pengaruh stres lingkungan terhadap proses fisiologi tanaman. Cekaman lingkungan dapat mempengaruhi aktivitas gen dan menentukan kapan, bagaimana dan berapa banyak
suatu enzim/protein dapat diproduksi dalam organ atau jaringan tanaman (Imelda et al., 2001).
Peroksidase menyebar luar dalam jaringan tanaman. Enzim ini berperan dalam perkembangan dan senesens jaringan tumbuhan. Beberapa peran tersebut antara lain dalam proses biogenesis etilena, peroksidase mengatur pematangan, penguraian klorofil dan oksidasi asam indol-3-asetat. Beberapa jenis peroksidase konvesional yang berasal dari tanaman antara lain pada tanaman lobak (horseradis h), kedelai, cengkeh dan bonggol jagung (Ilmi dan Kuswytasari, 2013).
Peroksidase mendorong oksidasi berbagai senyawa peroksida, terutama H2O2 (Hidrogen Peroksida) yang direduksi, membentuk air, enzim yang memilik i sifat aktif, pada tanaman yang tercekam dapat mengurangi kerusakan yang disebabkan oleh kerusakan lingkungan maupun penyakit seperti cendawan busuk pangkal batang. Diperkirakan bahwa peroksidase menghapus ROS, dan membant u mencegah kerusakan. Peroksidase merupakan senyawa oksireduktase dimana 2 elektron dan atom hidrogen dipindahkan dari alkohol menjadi aldehid. 2 elektron
pada ikatan karbon-hidrogen alkohol dipindah ke NAD+.NAD+ merupakan kofaktor yang menjembatani berbagai reaksi oksidasi reduksi (Yanti, 2011).
Prolin
Salah satu senyawa yang berperan dalam penyesuaian osmotikal sel adalah prolin. Yoshiba et al., (1997) mengatakan bahwa prolin adalah salah satu osmolitik kompatibel paling umum pada stres air tanaman yang berfungsi dalam pengaturan tekanan osmosis dan akumulasinya di dalam sel yang berperan dalam menjaga kesetimbangan air.
Prolin berfungsi sebagai pengatur tekanan di dalam sel, prolin juga berperan dalam mengurangi radikal bebas di dalam sel sehingga kerusakan yang disebabkan cekaman oksidatif dapat dicegah (Hong et al,. 2000). Menurut Pritchard et al. (2000), tanaman yang mendapat cekaman akan melakukan perlindungan dengan membentuk enzim senyawa antioksidan untuk mengura ngi tingkat detoksifikasi Reactive Oxygen Species (ROS) secara langsung maupun tidak langsung di dalam sel tanaman. Enzim-enzim yang terbentuk inilah yang akan dilindungi oleh prolin dari denaturasi, sehingga upaya pencegahan kerusakan dapat berlangsung secara optimal (Konstantinova et al,. 2002).
Dengan meningkatkan aku mulasi prolin di daun untuk menjaga keseimbangan potensial osmotik pada tanaman. Hal ini merupakan respon fisiolo gi yang cukup penting pada tanaman untuk mempertahankan tekanan turgor dengan menurunkan potensial osmotik sebagai mekanisme toleransi terhadap cekaman yang diberikan (Hamim et al,. 1996).
BAHAN DAN METODE Tempat dan Waktu Penelitian
Penelitian ini dilaksanakan di Lahan sawah irigasi Sememe Deli Tua Sumatera Utara, Medan, dan di Laboratorium Kultur Jaringan Universitas Sumatera Utara, Medan pada bulan Juni 2020 hingga April 2021
Bahan dan Alat Penelitian
Bahan yang digunakan dalam penelitian ini adalah benih kedelai varietas ( Anjasmoro,Grobogan, Deja 1, Devon 1 ), tanah top soil, kompos, GA3, Asam Salisilat, air, pupuk : Urea, TSP, KCL, aquades, insektisida, fungisid a, kertas label, aluminium foil, micro test tube ukuran 1,5 ml, nitrogen cair, PVP (Polyvinylpyrrolidone), EDTA (Ethylenediamineteraacetic acid), Monokalium fosfat (KH2PO4), Dikalium fosfat (K2HPO4), L- Methionin, NBT (Nitro Blue Tetrazolium), Kalsium Klorida (CaCl2), fenol, 4 Dimethylaminoantipyrine, riboflavin, etanol 95%, asam fosfor, aseton 85%, PVP (Polyvinylpyrrolidone), CBB G-250 (Coomasie Brillian Blue G-250), Asam Nihidrin.
Adapun alat yang digunakan adalah spektrofotometri, pipet tetes, erlenmeyer, cawan petri, batang pengaduk, gelas ukur, tabung reaksi, tube, micropipet, mortal dan alu, pinset, scalpel, gunting, Laminar Air Flow Cabinet, spatula, waterbath, kuvet, Centrifuge, spritus, hot plate, cangkul, handsprayer, saringan buchner, timbangan analitik, dan kamera.
Metode Penelitian
Penelitian ini menggunakan Rancangan Acak Kelompok ( RAK ) Satu Faktor yang terdiri dari :
V1 : Varietas Anjasmoro dengan lama penggenangan 96 jam V2 : Varietas Grobogan dengan lama penggenangan 96 jam V3 : Varietas Deja 1 dengan lama penggenangan 96 jam V4 : Varietas Devon 1 dengan lama penggenangan 96 jam
Jumlah Ulangan : 6
Jumlah plot : 24
Ukuran Plot : 1200 cm x 100 cm Jumlah tanaman per plot : 6 tanaman
Jumlah sampel per plot : 3 tanaman Jumlah sampel seluruhnya : 72 tanaman Jumlah Tanaman seluruhnya : 144 tanaman
Penelitian dilakukan dengan menggunakan Rancangan Acak Kelompok Satu Faktor dengan model matematis sebagai berikut :
Yij = µ + αi + βj + εij Dimana :
Yij = respon atau nilai pengamatan dari perlakuan ke-i dan ulangan ke-j µ = nilai tengah umum
αi = pengaruh perlakuan ke–i βj = pengaruh blok ke-j
εij = pengaruh galat percobaan dari pelakuan ke i dan ulangan ke-j.
PELAKSANAAN PENELITIAN Persiapan lahan
Areal yang dibutuhkan untuk penelitian terlebih dahulu diukur sesuai dengan kebutuhan yaitu 3,6 m x 3,7 m.
Persiapan Benih
Benih direndam selama 15 menit dengan fungisida terlebih dahulu sebelum penanaman agar terhindar dari jamur.
Penanaman
Penanaman dilakukan dengan membuat lubang tanam pada areal tanam dengan kedalaman ± 2 cm, kemudian dimasukkan 2 benih per lubang tanam dan kemudian ditutup kembali.
Pemupukan
Pemupukan dilakukan sesuai dengan dosis anjuran kebutuhan pupuk kedelai di lahan sawah dari Badan Penelitian dan Pengembangan Pertanian (2008) yaitu 75 kg urea/ha (9,6 g/plot), 100 kg SP-36/ha (12,8 g/plot), dan 100 kg KCl/ha (12,8 g/plot). Dilakukan sebanyak 2 kali, dilakukan pada awal pertumbuhan yaitu saat tanaman berumur 2 MST dan awal pembentukan polong atau pada saat tanaman berumur 4 MST
Pemeliharaan Tanaman Penyulaman
Penyulaman dilakukan untuk menggantikan tanaman yang mati dengan tanaman cadangan yang masih hidup. Penyulaman dilakukan pada saat varietas Grobogan dan Devon 1 berumur 2 MST, sedangkan penyulaman pada varietas
Anjasmoro dan varietas Deja 1 pada saat tanaman berumur 3 MST, dan dilakukan pada sore hari, untuk mencegah agar tidak terjadi proses pelayuan.
Penyiangan
Penyiangan dilakukan setiap minggu, sesuai dengan kondisi lapangan.
Penyiangan dilakukan dengan tujuan untuk menghindari persaingan hara antara gulma dengan tanaman. Penyiangan dilakukan secara manual untuk membersihka n gulma yang ada disekitar tanaman.
Aplikasi zat pengatur tumbuh (ZPT) GA3 dan Asam Salisilat
ZPT GA3 dan Asam Salisilat diaplikasikan dengan sistem penyemprotan pada semua tanaman dengan dosis 150 ppm untuk GA3 dan 100 ppm untuk Asam Salisila t pada saat tanaman memasuki umur 2 MST sampai sebelum pembungaan, namun dikarenakan adanya perbedaan umur berbunga setiap varietas aplikasi ZPT juga berbeda, pada varietas Anjasmoro dan varietas Deja 1 dilakukan aplikasi pada saat umur 2 MST sampai umur 4 MST, sedangkan pada varietas Grobogan dan Varietas Deja 1 dilakukan aplikasi ZPT pada umur 2 MST sampai umur 3 MST. ZPT GA3
dan Asam Salisilat di aplikasikan secara bergantian pada tanaman tiap minggunya.
Pada varietas Anjasmoro dan varietas Deja 1, aplikasi ZPT GA3 dilakukan sebanyak 2 kali, dan aplikasi ZPT Asam Salisilat sebanyak 1 kali, sedangkan pada varietas Grobogan dan Deja 1, aplikasi ZPT GA3 dilakukan sebanyak 1 kali, dan Aplikasi ZPT Asam Salisilat sebanyak 1 kali
Penggenangan
Penggenangan dilakukan dengan membuka saluran irigasi menuju plot tanaman dan diberi air ketinggiannya setengah dari tinggi tanaman. Penggenanga n
dilakukan saat tanaman memasuki umur 8 minggu setelah tanam (MST), digenangi selama 96 jam. (Gambar 1).
Peubah Amatan Pengukuran Klorofil
Analisis klorofil dilakukan di laboratorium kultur jaringan Fakultas Pertanian berdasarkan metode Arnon (1949) yaitu dengan mengambil 1 gram sampel daun tanaman yang telah dikompositkan kemudian dipotong kecil-kecil, digerus dengan menggunakan mortal dan alu sampai halus, dilarutkan kedalam aceton 85%, kemudian disentrifuse pada kecepatan 10000 rpm dengan waktu 5 menit, dilakukan pembacaan dengan menggunakan spektofotometer, diukur dengan panjang gelombang 649 nm dan 665 nm, jumlah klorofil a (g/ml), jumlah klorofil b (g/ml), jumlah klorofil total (g/ml) dihitung dengan menggunakan rumus OD (Optical Density) dengan rumus :
Klorofil a = {(13,7 x A665) – (5,76 x A649)} /10 Klorofil b = {(25,8 x A649) – (7,60 x A665)} / 10 Total klorofil = {(20,0 x A649) + (6,10 x A665)} / 10 Pengukuran Total Protein
Analisis protein dilakukan di laboratorium kultur jaringan Fakultas Pertanian Universitas Sumatera Utara. Sampel daun yang digunakan adalah daun yang telah berkembang sempurna. Analisis protein dinyatakan dalam satuan mg dan diamati berdasarkan metode yang dilakukan oleh Bradford (1976).
Gambar 1. Tanaman kedelai yang sedang digenangi.
Tahap analisis dimulai dengan dibuat reagen Bradford dengan menimba ng 0,02 g Coomasie B Briliant Blue (CBB) dan melarutkannya ke dalam 10 ml etanol 95% dan 20 ml asam fosfor. Diaduk dalam kondisi gelap dan disaring dengan kertas saring, lalu larutan dicampur 150 ml akuades. Pada tahap analisis dimulai dengan dimasukkan buffer ekstrak 1 ml ke dalam tube. Kemudian ditimbang daun kedelai 0,1 g lalu digerus dengan N2 cair. Ditambahkan PVP 1% sebanyak 0,1 g dan digerus kembali sampai menjadi serbuk, kemudian dimasukkan ke dalam tube yang berisi BE dan disimpan dalam kondisi dingin lalu disentrifuse dengan kecepatan 10.000 rpm dan suhu 40C selama 10 menit. Disiapkan tabung reaksi dan dimasukkan sampel 50 μL, ditambahkan bradford 2,5 ml dan diinkubasi selama 10-60 menit dalam kondisi gelap. Dibuat larutan blanko yang terbuat dari larutan bradford + BE 50 μL dan kemudian dibaca dengan spektrofotometer dengan panjang gelombang 59,5 nm.
Enzim Peroksida Dismutase (POD) (mg/g)
Analisis enzim peroksida dismutase dilakukan di Laboratorium Kultur Jaringan Fakultas Pertanian Universitas Sumatera Utara. Analisis POD diamati berdasarkan metode yang dilakukan oleh Standart Operating Procedures (1994).
Aktivitas POD dinyatakan dalam satuan unit/mg protein. Selanjutnya dihitung dengan rumus:
Aktivitas POD = Af − Ai mg protein Keterangan:
Af = pembacaan peroksidase akhir Ai = pembacaan peroksidase awal Uji Prolin
Pengukuran akumulasi prolin dilakukan dengan metode Ninhidr in (Bates et al. dalam Umebese et al., 2009).
Tahapan analisis dilakukan dengan disiapkan alat dan bahan yang diperlukan, lalu ditimbang daun sebanyak 10 gr/sampel setelah itu digerus daun yang sudah ditambahkan Aquadest dengan mortal dan alu sampai halus seperti bubur, hasil gerusan di saring dengan menggunakan kertas saring. Ditambahka n 25 ml alkohol 96% Hasil gerusan dibaca dengan menggunakan spektofotometer dan dihitung menggunakan rumus:
=( Prolin/ml x ml Toluena )( g Sampel ) ( 115,5 µg/mol)(5)
HASIL DAN PEMBAHASAN Hasil dan Pembahasan
Hasil penelitian menunjukkan bahwa analisis total klorofil, analis is peroksidae oksidase, dan analisis prolin pada saat dilakukan penggenangan pada parameter analisis total klorofil, analisis total protein, analisis peroksidae oksidase, dan analisis prolin berpengaruh nyata antara keempat varietas yang digunaka n.
Untuk lebih jelas mengenai hasil pada masing masing parameter dapat dilihat pada paragraf berikut ini :
Analisis Total Klorofil
Pengukuran klorofil total dilakukan dengan penggunaan spektrofotometer dengan panjang gelombang 649 nm dan 665 nm. Hasil pengukuran klorofil dapat dilihat pada lampiran 7 dan tabel 3 sebagai berikut:
Tabel 3. Analisis Total Klorofil Kondisi Tergenang
Keterangan : angka yang diikuti huruf yang sama pada kolom yang sama menunj uka n tidak berpengaruh nyata pada uji rataan berganda duncan’s multip le range test (DMRT) pada taraf 5 %
Berdasarkan hasil pengamatan klorofil total yang di tunjukkan pada tabel 2 menunjukkan bahwa pada kandungan klorofil terjadi penurunan pada setiap varietas, sehingga kandungan klorofil tertinggi berada pada varietas Devon 1 ( 1,96 mg/g) dan terendah tetap berada di varietas Anjasmoro (0,46 mg/g).
Varietas Kondisi Tergenang
Anjasmoro 0,46a
Grobogan 1,96c
Deja 1 1,30b
Devon 1 2,00d
Berdasarkan hasil analisis menunjukkan bahwa kandungan klorofil setiap varietas terhadap penggenangan pada umur 8 minggu setelah tanaman (MST) menunjukkan adanya perbedaan yang nyata antara varietas Anjasmoro, varietas Grobogan, varietas Devon 1 dan varietas Deja 1.
Dari hasil penelitian juga menunjukan bahwa varietas Devon 1 memilik i kandungan klorofil yang paling tinggi dibandingkan tiga varietas lainnya, namun memiliki kandungan klorofil yang tidak berbeda jauh dengan varietas Grobogan, hal ini dikarenakan kondisi fisik dari varietas Devon 1 dan varietas Grobogan yang lebih stabil saat dilakukan penggenangan.
Hasil penelitian menunjukkan bahwa varietas Anjasmoro memilik i kandungan klorofil yang paling rendah dibandingkan tiga varietas lainnya. Hal ini disebabkan karena cekaman air langsung dapat menyebabkan penurunan turgor tanaman. Tekanan turgor sangat berperan dalam menentukan ukuran tanaman, berpengaruh terhadap pembesaran dan perbanyakan sel tanaman, membuka dan menutupnya stomata, perkembangan daun, pembentukan dan perkembangan bunga.
Secara tidak langsung hal tersebut berpengaruh terhadap proses fisiologis seperti fotosintesis. Hal ini sejalan dengan pernyataan Tampubolon et al ( 1989) Penggenangan pada umur 8-10 MST daun-daun akan menguning dan mulai gugur, sehingga mengganggu proses fotosintesis.
Saat dilakukan penggenangan tanaman akan terlebih dahulu beradaptasi untuk mempertahankan kondisi tanaman, agar tidak mengalami pelayuan saat dilakukan penggenangan. Menurut Kosova, et.al (2011) genangan pada tanah menyebabkan akar tanaman mengalami gangguan dalam respirasi, penyerapan unsur hara dan metabolisme tanaman secara keseluruhan. Unsur hara yang kurang pada
tanaman menyebabkan pembentukan klorofil terganggu dan kadar klorofil pada daun menjadi turun. Selain itu, menurut Syafi (2008) telah disebutkan sebelumnya bahwa sintesis klorofil dipengaruhi oleh berbagai faktor salah satunya adalah unsur N dan Mg. Genangan menyebabkan pH media cenderung menurun (masam) sehingga menyebabkan serapan N dan Mg menurun dan aktivitas mikroorganisme tanah Rhizobium terganggu.
Analisis Aktivitas Enzim Peroksidase (POD)
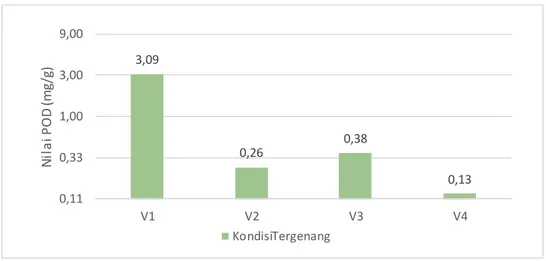
Parameter analisis enzim peroksidase (POD) dapat dilihat pada lampiran 12 dan pada gambar. 2 sebagai berikut :
Gambar 3. Histogram rataan aktivitas enzim peroksidase (pod) 4 (empat) varietas kedelai pada kondisi tergenang.
Berdasarkan hasil analisis enzim peroksidase (POD) yang ditunjukkan pada gambar 2 dapat dilihat varietas kedelai saat kondisi tergenang menunjukkan bahwa kandungan enzim peroksidase tertinggi terdapat pada varietas Anjasmoro (3,09 mg/g) dan kandungan enzim peroksidase yang terendah terdapat pada varietas Devon 1 (0.13 mg/g). Berdasarkan hasil analisis menunjukkan bahwa kandungan protein setiap varietas terhadap penggenangan pada umur 8 minggu setelah tanaman (MST) menunjukkan pola yang berbeda-beda pada setiap varietas
3,09
0,26 0,38
0,13 0,11
0,33 1,00 3,00 9,00
V1 V2 V3 V4
Nilai POD (mg/g)
KondisiTergenang
Hal ini menunjukkan bahwa varietas Anjasmoro toleran terhadap genangan, karena nilai dari enzim peroksidase yang tinggi, penggenangan yang dilakukan pada umur 8 MST varietas Anjasmoro sudah memasuki fase pengisisan polong, dimana pada fase ini merupakan kondisi tanaman yang sensitif, sehingga peningkata n aktivitas POD untuk mempertahankan kondisi tanaman kedelai saat terjadi genangan.
Hal ini sesuai dengan pernyataan Sembiring, et al (2016) yang menyatakan bahwa, semakin tinggi aktifitas enzim peroksidase maka semakin tahan tanaman atau varietas itu terhadap stress genangan. Hal ini juga sejalan dengan pernyataan Ali dan Alqurainy (2016) yang menyatakan bahwa pada tanaman yang toleran terhadap genangan dalam jangka pendek akan terjadi peningkatan aktivitas enzim peroksidase, sedangkan dalam waktu yang berkepanjangan akan menurunkan aktivitas enzim peroksidase.
Berdasarkan pernyataan Abdullah dan Titik (2012) Kebutuhan air tanaman kedelai pada fase generatif lebih tinggi dibandingkan pada fase vegetatif, namun saat kondisi tanaman mengalami kelebihan kandungan air akan mengga nggu metabolisme pertumbuhan tanaman, sehingga untuk mempertahankan kondisi tanaman kedelai dari cekaman genangan, maka akan terjadi peningkatan kandungan enzim peroksidase dismute (POD). Hal ini sejalan dengan pernyataan Manurung (2017) yang menyatakan bahwa tanaman yang mengalami cekaman genangan akan memproduksi molekul reaktif seperti singlet oksigen (O2), hidrogen-peroksida (H2
O2), superoksida (O2–) dan radikal hidroksil (OH-) yang dapat merusak sel dan kerja enzim. Salah satu cara tanaman untuk melindungi sel dari efek cekaman ini adalah dengan memproduksi enzim antioksidan yaitu enzim POD. Semakin tinggi nila i enzim POD maka tanaman akan semakin tahan pada cekaman genangan.
Analisis Prolin
Parameter analisis prolin dapat dilihat pada lampiran 11 dan pada tabel 5 sebagai berikut :
Tabel 4. Analisis Prolin Kondisi Tergenang
Keterangan : angka yang diikuti huruf yang sama pada kolom yang sama menunj uka n tidak berpengaruh nyata pada uji rataan berganda duncan’s multip le range test (dmrt) pada taraf 5 %
Berdasarkan hasil analisis kandungan prolin pada tabel 5, menunjukan bahwa kandungan prolin pada saat kondisi tergenang menunjukkan adanya peningkata n pada setiap varietas, dan kandungan prolin tertinggi terdapat pada varietas Anjasmoro (2,36 µmol/gr ) sedangkan kandungan yang terendah terdapat pada varietas Devon 1 (0,73 µmol/gr ).
Berdasarkan hasil analisis menunjukkan bahwa kandungan prolin setiap varietas terhadap penggenangan pada umur 8 MST menunjukkan adanya perbedaan yang nyata antara varietas Anjasmoro, varietas Grobogan, varietas Devon 1 dan varietas Deja 1.
Pada dasarnya kandungan prolin yang terkandung pada tanaman kedelai dipengaruhi oleh ketersediaan air. Hal ini menyatakan bahwa varietas Anjasmoro merupakan varietas yang toleran terhadap genangan, karena semakin tinggi akumulasi prolin yang dihasilkan, semakin baik resistensinya, dimana prolin merupakan senyawa metabolit yang pada dasarnya sudah berada di dalam tanama n.
Varietas Setelah
Penggenangan
Anjasmoro 2,36d
Grobogan 1,07b
Deja 1 1,57c
Devon 1 0,73a
Terjadi peningkatan konsentrasi prolin, dikarenakan prolin berfungsi untuk melindungi tanaman tersebut dari stress atau cekaman, Hal ini sesuai dengan pendapat Yoshiba et al., (1997) mengatakan bahwa prolin adalah salah satu osmolitik kompatibel paling umum pada stres air tanaman yang berfungsi dalam pengaturan tekanan osmosis dan akumulasinya di dalam sel yang berperan dalam menjaga kesetimbangan air.
Naiknya nilai dari kandungan prolin pada varietas Anjasmoro akibat dilakukan penggenangan juga mempengaruhi produksi varietas Anjasmoro dimana berdasarkan penelitian Manullang (2021) bobot 100 biji dan jumlah polong berisi yang mengalami cekaman genangan 96 jam pada varietas Anjasmoro memiliki nila i yang paling tinggi dibandingkan tiga varietas lainnya, hal ini menunjukkan bahwa kenaikan kandungan prolin dapat mempertahankan kestabilan kondisi tanaman kedelai, agar hasil produksinya tidak terganggu.
KESIMPULAN DAN SARAN Kesimpulan
1. Varietas Anjasmoro kondisi tergenang memiliki kandungan enzim peroksidase dan kandungan prolin yang lebih tinggi dibandingkan dengan varietas Grobogan, varietas Deja 1 dan varietas Devon 1
2. Varietas Devon 1 kondisi tergenang memiliki kandungan total klorofil dan kandungan total protein yang lebih tinggi, dibandingkan dengan varietas Anjasmoro, varietas Grobogan dan varietas Deja 1
3. Empat varietas kedelai yang digenangi selama 96 jam pada umur 8 MST memberikan pengaruh yang nyata terhadap sifat fisiologis pada kandungan klorofil, enzim peroksidase, dan kandungan prolin.
Saran
Saran dari peneliti sebaiknya dilakukan pengujian dengan menggunaka n varietas terbaik dari penelitian ini yaitu varietas Anjasmoro dan varietas Devon 1 untuk melihat karakter generatif secara fisiologi pada saat kondisi tergenang.
DAFTAR PUSTAKA
Adisarwanto, T. 2005. Kedelai. Jakarta : Penebar Swadaya.
Adisarwanto, T. 2008. Budidaya Kedelai Tropika. Penebar Swadaya. Jakarta.
Ali, A., dan F. Alqurainy. 2016. Activities of Antioxidant in Plants Under Environmental Stress. Faculty of Science.King Saud University. Saudi Arabia.
Andrianto, T. T., dan N. Indarto. 2004. Budidaya dan Analisis Usaha Tani Kedelai, Kacang Hijau dan Kacang Panjang. Absolut, Yogyakarta.
Armstrong W. 1979.Aeration in higher plants. Advances in Botanical Research7:
225-332.
Atman. 2006. Budidaya Kedelai di Lahan Sawah Sumatera Barat. Jurnal Ilmiah Tambua 5(3).
Baharsyah, J.S., Suardi, D., Las, I. 1985. Hubungan Iklim dan Pertumbuhan Kedelai, hal 87-102. dalam Somaatmadja, S, Ismunadji, M, Sumarno, Syam, M, Manurung, SO, Yuswadi (Eds). Kedelai. Balai Penelitian dan Pengembanga n Pertanian. Pusat Penelitian Tanaman Pangan. Bogor
Balai Penelitian Tanaman Aneka Kacang dan Umbi. 2005. Deskripsi Varietas Unggul Kacang-Kacangan dan Umbi- Umbian. Malang
Balitkabi. 2016. Deskripsi Varietas Unggul Kedelai 1918 – 2016. Malang
Beauchamp C., and I. Fridovich. 1971. Superoxide Dismutase: Improved Assays and an Assay Applicable to Acrylamide Gels. Analytical Biochemistry 44, 276-287.
Blokhina, O., Virolainen, E. dan Fagerstedt, K. V. 2003. Antioxidants, Oxidative Damage and Oxygen Deprivation Stress: A Review. An-nals of Botany 91:179–194.
BPTP NAD. 2009. Badan Ketahanan Pangan dan Penyuluh Pertanian Aceh, Aceh Bradford, M. 1976. Rapid and Sensitive Method For The Quantitation Of
Microgram Quantities Of Protein Utilizing The Principle Dye Binding.
Analytical of Biochemistry., 72, 248-254.
Dennis, ES, R. Dolferus, M. Ellis, M. Rahman, Y. Wu, F.U. Hoeren, A. Grover, K.P. Ismond, A.G. Good, and W.J. Peacock. 2000. Molecular strategies for improving waterlogging tolerance in plants. J. Exp. Bot. 51(342):89-97
Dwidjoseputro, 1994.Pengantar Fisiologi Tumbuhan. Jakarta: PT Gramedia Pustaka Utama.
Dwiputra, Adhytia, Erwin Didik, dan Eka Tawarca Susila, 2015. Hubunga n Komponen hasil dan Tiga Belas Kultivar Kedelai (Glycine max (L.) Merr.).
Fakultas Pertanian Universitas Gadjah Mada, Yogyakarta. Vol. 4
Fatimah, V., S., dan T., B., Saputro. 2016. Respon Karakter Fisiologi Kedelai (Glycine max L. Merr.) Varietas Grobogan terhadap Cekaman Genangan.
Jurnal Sains dan Seni ITS Vol. 5, No. 2.
Fehr. W. R., C. E. Cavieness, D. T. Burmood, and J. S. Pennington. 1971. Stage of Development Description for Soybean Glycine max (L.) Merril. Crop Sci.
Madison. USA. 11 hlm.
Girousse, C., R. Bournoville, and J.L. Bonnemain. 1996. Water Deficit Induced Changes In Concentrations of Proline and Some Other Amino Acids In Phloem Sap Alfalfa. Plant Physiol. 111 : 109 – 113.
Hamim, D. Sopandie, M. Jusup. 1996. Beberapa karakteristik morfologi dan fisiolo gi kedelai toleran dan peka cekaman kekeringan. Hayati. 3:3
Hanum, C. 2008. Teknik Budidaya Tanaman Jilid 2. Direktorat Pembinaan Sekolah Menengah Kejuruan. Departemen Pendidikan Nasional.
Hapsari, R. T. dan M. M. Adie. 2010. Peluang perakitan dan pengembangan kedelai toleran genangan. Balai Penelitian dan Pengembangan Tanaman Kacang- kacangan dan Umbi-umbian. Bogor. Jurnal Litbang Pertanian, 29(2).
Hidayat, O. D. 1985. Morfologi Tanaman Kedelai. Hal 73-86. Dalam S.
Somaatmadja et al. (Eds.). Puslitbangtan. Bogor.
Hong Z, Lakkineni K, Zhang Z, Verma DPS. 2000. Removal of Feedback Inhibit io n of D1 Pyrroline-5- Carboxylate Synthetase Results in Increased Proline Accumulation and Protection of Plants from Osmotic Stress1. Plant Physiology, 122 : 1129-1136.
Ilmi, I. M., Kuswytasari, N. D. 2013. Aktifitas Enzim Lignin Peroksidase oleh Gliomastix sp. T3.7 pada Limbah Bonggol Jagung dengan Berbagai pH dan Suhu. Jurnal Sains Dan Seni Pomits Vol. 2, No.1.
Imelda, M., Estiati, A., Hartati, N. S. 2001. Induction of Mutation through Gamma Irradition in three Cultivars of Banana J. Annalaes Bogorienses7(2):75-82.
Irwan, A. W. 2006. Budidaya Tanaman Kedelai (Glycine max (L.) Merril).
Universitas Padjajaran. Bandung.
Kadarwati, T.F. 2006. Pemupukan Rasional dalam Upaya Peningkatan Produktivitas Kapas. Malang : Balai Penelitian Tanaman Tembakau dan Serat. Jurnal Perspektif. Volume 5 (2) hlm: 59-70.
Konstantinova T, Parvanova D, Attanassov A, Djilianov D. 2002. Freezing Tolerant Tobacco, Transformed To Accumulate Osmoprotectants. Plant Science 163 : 157-164.
Kosova, K., Vıtamvas, P., Prasil, I.T., Renaut, J. 2011. Plant Proteome Changes under Abiotic Stress-Contribution of Proteomics Studies to Understanding Plant Stress Response. J Proteom, 74: 1301-1302
Kuai, J., Liu, Z., Wang, Y., Meng, Y., Chen, B., Zhao, W. 2014. Waterlogging during flowering and boll forming stages affects sucrose metabolism in the leaves subtending the cotton boll and its relationship with boll weight. Plant Sci. 223, 79–98. doi: 10.1016/j.plantsci.2014.03.010
Kuncahyo, Aditya. 2018. Studi Pertumbuhan, Produksi, dan Mutu Benih Sembila n Varietas Kedelai (Glycine max [L.] Merril) yang Ditanam Dilahan Sawah Musim Kemarau. Universitas Lampung, Lampung.
Kurniawan, S., Aslim Rasyad, dan Wardati. 2013. Pengaruh Pemberian Pupuk Posfor Terhadap Pertumbuhan Beberapa Varietas Kedelai (Glycine max (L)Merril ).
Jurnal Online. Universitas Riau
Linkemer, G., Board, J.E. and Musgrave, M.E. 1998. Waterlogging effects on growth and yield components in late-planted soybean. Crop Sci. 38:1576- 1584.
Manullang, Ester.,S. 2021. Respon Karakter Morfologi Beberapa Varietas Tanaman Kedelai (Glycine max L. Merril) pada Fase R5 terhadap Kondisi Tergenang.
Universitas Sumatera Utara. Medan
Manurung, Ida Rumia. 2017. Respon Fisiologi Kedelai ( Glycine max (L) Merill.) Generasi Keempat Tahan Salin dengan Pemberian Beberapa Jenis Antioksidan. Tesis. Universitas Sumatera Utara. Medan.
Mathius, N.T., G. Wijana, E. Guharja, H. Aswindinnoor, Y. Sudirman, dan Subronto.
2001. Respon Tanaman Kelapa Sawit (Elaeis guineensis Jacq.) terhadap Cekaman Kekeringan. Menara Perkebunan 69 : 29 - 45.
N. Hendriyani, I.S., dan Setiari, “Kandungan Klorofil Dan Pertumbuhan Kacang Panjang (Vignasinensis) Pada Tingkat Penyediaan Air Yang Berbeda,” J.
Sains Mat, vol. 17, no. 3, p. 150, 2009
Notohadiprawiro, T. 1989. Pola kebijakan pemanfaatan sumber daya lahan basah, rawa, dan pantai. Seminar Ilmiah Dies Natalis ke - 25 Universitas Jember 14- 15 Juli 1989. <http://soil.faperta.ugm.ac.id/> [20 January 2009].
Pompeianoa, A., T. H., Reyesb, T. M. Molesb, M. Villanib, M. Volterranib, L.
Guglielminettib, dan A. Scartazzac. 2017. Inter- and intraspecific variabilit y in physiological traits and post-anoxia recovery of photosynthetic efficie nc y in grasses under oxygen deprivation. Physiologia Plantarum 161: 385–399.
2017, ISSN 0031-9317.
Pritchard SG, Ju Z, Santen EV, Qiu J, Weaver DB, Prior SA, Roger H. 2000. The Influenc of elevated CO2 on the activities of antioxidative enzymes in two soybean genotypes. Aust J Plant Physiol 27 : 1061- 1068.
Rachman, Ahmad, IGM, Subiksa,dan Wahyunto. 2016. Perluasan Areal Tanaman Kedelai ke Lahan Suboptimal. Balai Penelitian Tanah. Bogor.
Riche, C.J. 2004. Identification of soybean cultivars tolerance to waterloggi ng through analyses of leaf nitrogen concentration. Lousiana State Univers it y Electronic Thesis and Dissertation Collection. http://etd. ls u.
edu/docs/available/etd-04132004-154236/unrestricted/Riche_thesis.pdf.
[20 January 2020].
Rohmah ,Eka Afiyanti dan Triono Bagus Saputro ,2016 Analisis Pertumbuha n Tanaman Kedelai (Glycine max L.) Varietas Grobogan Pada Kondisi Cekaman Genangan . jurnal FMIPA , institut teknologi sepuluh november.
Rubatzky, V. E., dan M. Yamaguchi. 1998. Sayuran Dunia, Prinsip, Produksi, dan Gizi. Edisi Kedua. Penerjemah C. Herison. ITB Press, Bandung.
Sabehat A, D Weiss, S Lurie .1998. Heatshock proteins and cross-tolerance in plants.
Physiol Plant. 103: 437-441.
Sairam, R.K., D. Kumutha, and K. Ezhilmathi. 2009. Waterlogging tolerance:
nonsymbiotic haemoglobin-nitric oxide homeostatis and antioxida nt s.
Curr. Sci. 96(5): 674-682
Scott, H. D., J. De Angulo, M. B. Daniels, and L. S. Wood. 1989. Flood duration effect on soybean growth and yield. Agronomy 81:631-636.
Sembiring, Meilina J, Revandy I. M. Damanik dan Luthfi A. M. Siregar. 2016 Respon Pertumbuhan Beberapa Varietas Kedelai (Glycine max L. Merrill) Pada Keadaan Tergenang Terhadap Pemberian GA3. Universitas Sumatera Utara. Medan
Sugeng. 2000. Bercocok Tanam Sayuran. Aneka Ilmu, Semarang.
Sumarno, dkk. 1999. Taksonomi, Botani, Teknik Produksi dan pengembanga n Kedelai. Pusat Penelitian dan Pengembangan Tanaman Pangan. Bogor Sunardi, O., SA Adimihardja, dan Y Mulyaningsih. 2013. Pengaruh Tingkat
Pemberian ZPT Gibberellin ( GA3 ) terhadap Pertumbuhan Vegetatif