Analisis Lintasan Genotipik dan Fenotipik Karakter Sekunder Jagung
pada Fase Pembungaan dengan Pemupukan Takaran Rendah
Sutoro
Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian Jl. Tentara Pelajar 3A Bogor, Jawa Barat
ABSTRACT. Path Analysis of the Genotypic and Phenotypic Secondary Characters of Maize Plant at Flowering Stage under Low Fertilizer Input. High yielding potential of corn
varieties which adaptive under low fertilizer input is needed in part of corn breeding program, because few farmers usually apply fertilizer less than the optimal dosage. Plant selection on primary character (grain yield) could be intensified based on selection of secondary plant characters, namely plant traits which contributed to grain yield as direct effect or indirect effect. Study was conducted to identify the contribution of direct and indirect effect of secondary plant characters to grain yield of corn under low fertilizer application. The contributions of traits were identified by using path analysis, which partitions the correlation among plant characters into direct and indirect effects. Genetic and phenotypic correlations were studied using material developed by North Caroline Design II mating. The progenies consisted of 27 sets, namely each of 6 S1 parent
lines (3 as females and 3 as males). The progenies (243 full-sib families) were evaluated under low fertilizer dosage, at Cikeumeuh, experiment station Bogor in 2002, using incomplete randomized block design, 3 replications. Direct and indirect effects of secondary characters (anthesis silking interval (ASI), chlorophyll, ear position on the stem, leaf number above ear, leaf area index and leaf number at flowering stage, and seed initiation rate) to grain yield showed that genetic and phenotypic correlation between ASI, seed initiation rate and grain yield resulted in the same direction, and most of their correlation contributed by direct effect. Therefore, ASI and seed initiation rate could be considered as selection criteria for grain yield of corn.
Keywords: Corn, path analysis, secondary characters
ABSTRAK. Kegiatan pemuliaan tanaman perlu mendapatkan varietas
yang berdaya hasil tinggi dan adaptif pada kondisi pemupukan takaran rendah, mengingat seringnya petani memberkan pupuk dalam jumlah yang kurang optimum. Untuk meningkatkan efisiensi seleksi karakter primer (bobot biji) dalam pemuliaan jagung dapat digunakan karakter sekunder. Tujuan penelitian ini adalah untuk mendapatkan informasi tentang kontribusi pengaruh langsung dan tidak langsung dari karakter sekunder secara genetik dan fenotipik antara bobot biji jagung dengan karakter sekunder pada kondisi pemupukan takaran rendah. Untuk mendapatkan koefisien korelasi genetik fenotipik dibuat persilangan antargalur S1 yang berasal dari populasi Bisma menurut rancangan persilangan NCD II. Biji hasil persilangan antargalur S1 terdiri atas 27 set persilangan,
masing-masing set terdiri atas enam galur S1 (tiga galur sebagai tetua jantan dan tiga galur sebagai tetua betina). Selanjutnya biji hasil persilangan (243 famili full-sib) dievaluasi dalam rancangan acak kelompok tidak lengkap (Incomplete randomized block design) di Kebun Percobaan Cikeumeuh, Bogor, pada tahun 2002. Pengaruh langsung dan tidak langsung yang menjelaskan hubungan antar karakter sekunder dengan bobot biji dipelajari melalui analisis lintasan. Hasil penelitian menunjukkan bahwa korelasi fenotipik dan genotipik antara bobot biji dengan ASI dan laju pengisian biji memberikan arah yang sama dan sebagian besar diperoleh dari kontribusi pengaruh langsung.
Oleh karena itu, karakter ASI dan laju pengisian biji dapat dipertimbangkan dalam seleksi hasil biji jagung.
Kata kunci: Jagung, analisis lintasan, karakter sekunder
P
hara yang tersedia dalam tanah. Kemampuanroduktivitas tanaman antara lain dipengaruhi oleh petani dalam memberikan hara dalam bentuk pupuk pada tanaman semakin terbatas, dengan semakin mahalnya harga pupuk. Dengan demikian pupuk yang diberikan tidak dalam jumlah yang optimum. Oleh karena itu, kegiatan pemuliaan tanaman perlu mendapatkan varietas yang berdaya hasil tinggi dan adaptif pada kondisi pemupukan rendah. Varietas yang adaptif pada kondisi pemupukan takaran rendah dapat diperoleh melalui seleksi populasi tanaman terhadap karakter primer maupun sekunder.Untuk meningkatkan efisiensi seleksi karakter primer (bobot biji) dapat digunakan karakter sekunder (Banziger and Lafitte 1997; Chapman and Edmeades 1999). Pengaruh langsung dan tidak langsung karakter sekunder terhadap bobot biji pada pemupukan takaran rendah dapat diduga melalui analisis lintasan (path analysis) berdasarkan korelasi genotipik dan fenotipik. Dalam analisis lintasan pada dasarnya korelasi dua karaker terdiri atas dua komponen lintasan, langsung dan tidak langsung melalui karakter lain. Berdasarkan analisis lintasan maka bila korelasi genetik antar karakter primer (berat biji) dengan karakter sekunder hampir sama dengan pengaruh langsungnya, maka korelasi yang dihasilkan menunjukkan hubungan yang sebenarnya dari kedua karakter tersebut, sehingga seleksi langsung pada karakter sekunder tersebut akan efektif. Jika koefisien korelasi berlawanan tanda dengan koefisien lintasan pengaruh langsung maka korelasi genetik atau fenotipik disebabkan oleh pengaruh tidak langsung. Dalam situasi ini maka pengaruh seleksi tidak langsung melalui karakter sekunder yang lain perlu dipertimbangkan (Singh dan Chaudhary 1979).
Hasil penelitian sebelumnya menunjukkan bahwa karakter sekunder seperti anthesis silking interval (ASI), klorofil, jumlah daun, dan LAI saat berbunga memiliki korelasi genetik yang nyata dengan bobot biji (Sutoro et al. 2007). Namun belum diketahui besarnya pengaruh
langsung maupun tidak langsung karakter sekunder tersebut. Jika korelasi fenotipik lebih besar daripada korelasi genetik dan memiliki tanda (+/-) yang sama berarti pengaruh lingkungan dan atau gen nonaditif berperan dalam arah yang sama (Sidwell et al. 1976). Jika pengaruh langsung fenotipik dan genotipik karakter sekunder hampir sama besarannya maka karakter tersebut dipengaruhi oleh gen aditif (Kang et al. 1983) atau sedikit dipengaruhi oleh pengaruh gen nonaditif dan atau lingkungan (Sidwell et al. 1976).
Tujuan penelitian ini adalah untuk mendapatkan informasi tentang kontribusi pengaruh langsung dan tidak langsung dari karakter sekunder secara genetik dan fenotipik antara bobot biji jagung dengan karakter sekunder pada kondisi pemupukan takaran rendah.
BAHAN DAN METODE
Untuk mendapatkan koefisien korelasi genetik dan fenotipik, dibuat persilangan antargalur S1 yang berasal dari populasi Bisma menurut rancangan persilangan NCD II. Biji hasil persilangan antargalur S1 terdiri atas 27 set persilangan, masing-masing set terdiri atas enam galur S1 (tiga galur sebagai tetua jantan dan tiga galur sebagai tetua betina). Selanjutnya, biji hasil persilangan dievaluasi dalam rancangan acak kelompok tidak
lengkap (Incomplete randomized block design) dengan tiga ulangan pada pemupukan takaran rendah (75 kg urea dan 25 kg SP36/ha) di Kebun Percobaan Cikeumeuh, Bogor, pada tahun 2002. Masing-masing progeni ditanam sebanyak 20 tanaman tiap plot. Karakter sekunder tanaman jagung yang diamati adalah ASI, kadar klorofil yang diukur dengan Chlorophylmeter Minolta SPAD-502, tinggi letak tongkol, jumlah daun, dan LAI (leaf area index), jumlah daun di atas tongkol pada saat pembungaan, dan laju pengisian biji yang dihitung dengan bobot biji pada saat panen dibagi dengan selisih umur panen dan umur berbunga betina (silking).
Besaran ragam aditif 2
A dan dominan 2
D dihitung
menurut Subandi et al. (1972), dan rasio 2 D/
2 A
digunakan untuk menduga derajat dominansi karakter menurut Robinson et al. (1955).
Korelasi genetik antardua karakter i dan j (Falconer and Mackay 1996), yaitu
Covz (ij) rG (ij) = ————————, (2 z(i) 2 z(j)) 1/2
dan korelasi fenotipik antar dua karakter i dan j yaitu Covz (ij) + Cove (ij) /r
rP (ij) = ——————————————, [(2 z(i) + 2 e(i) /r)( 2 z(j) + 2 e(j) /r] 1/2 di mana
Covz (ij)= peragam genotipik antara karakter i dan j, Cove (ij)= peragam galat antara karakter i dan j, 2z(i) = ragam genotipik karakter i dan j, 2e(i) = ragam galat karakter i,
r = banyaknya ulangan.
Besaran 2
z(i) dan Covz (ij) serta 2
e(i) diduga dari tabel
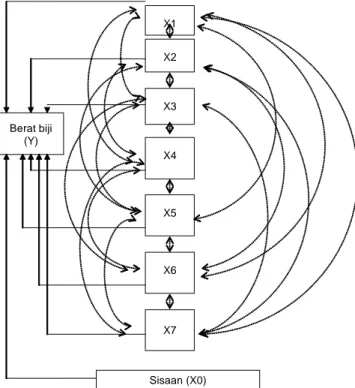
analisis ragam peragam berdasarkan rancangan persilangan NCDII. Pengaruh langsung dan tidak langsung yang menjelaskan hubungan antarkarakter sekunder dengan bobot biji diasumsikan seperti disajikan pada diagram analisis lintasan (Gambar 1). Koefisien pengaruh langsung dan tidak langsung diduga menurut arahan Singh dan Chaudhar y (1979). Pendugaan derajat dominansi dari setiap karakter digunakan pendugaan menurut Robinson et al. (1955). Rasio ragam dominan dan ragam aditif untuk satu lokus 2
D/ 2
A = 4q
2 (1 – q)2 a2 / 2q(1-q){1 + (1 – 2q)a}2,
sedangkan q = frekuensi alel favorable, a = derajat dominansi. Berat biji (Y) X1 X2 X3 X4 X5 X6 X7 Sisaan (X0)
HASIL DAN PEMBAHASAN
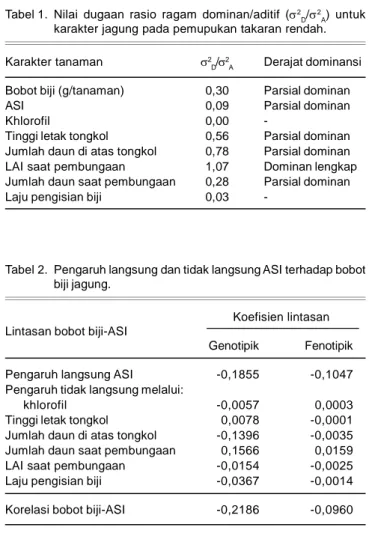
Pendugaan Derajat Dominansi Karakter Nilai dugaan rasio ragam dominan/aditif dari karakter tanaman jagung varietas Bisma pada pemupukan takaran rendah disajikan pada Tabel 1. Derajat dominansi apabila bernilai 1 merupakan dominan lengkap, >1 sebagai over dominan dan < 1 sebagai parsial dominan (Kearsy and Pooni 1996).
Dengan asumsi semua lokus memiliki derajat dominansi yang sama, dan frekuensi alel favorable untuk semua karakter pada pemupukan takaran rendah 0,7<q<0,9. Nilai q dipilih dengan nilai tersebut dengan pertimbangan pada pemupukan optimal frekuensi alel favorable >90% (q>0,9) karena varietas Bisma ter-bentuk setelah mengalami seleksi pada kondisi pemupukan optimal sehingga alel favorable tinggi. Pada pemupukan takaran rendah dipilih nilai q lebih kecil karena varietas Bisma tidak diseleksi langsung pada lingkungan pemupukan yang lebih rendah sehingga alel favorable-nya diduga lebih kecil.
Berdasarkan pertimbangan tersebut maka derajat dominansi bobot biji pada pemupukan takaran rendah adalah parsial dominan, karena rasio ragam dominan/ aditif sebesar 0,3 dan nilai derajat dominansi 0,6<a<0,7 (Robinson et al. 1955). Hallauer dan Miranda (1985) menyatakan aksi gen bobot biji kebanyakan meng-ekspresikan secara parsial dominan daripada dominan lengkap maupun overdominan. Karakter sekunder ASI, klorofil dan laju pengisian biji memiliki dominansi parsial yang sangat kecil pada kondisi pemupukan rendah (Tabel 1). Tinggi letak tongkol dan jumlah daun di atas tongkol memiliki derajat dominansi parsial. L AI berbunga memiliki dominan lengkap, sedangkan karakter jumlah daun saat pembungaan memiliki derajat dominansi parsial.
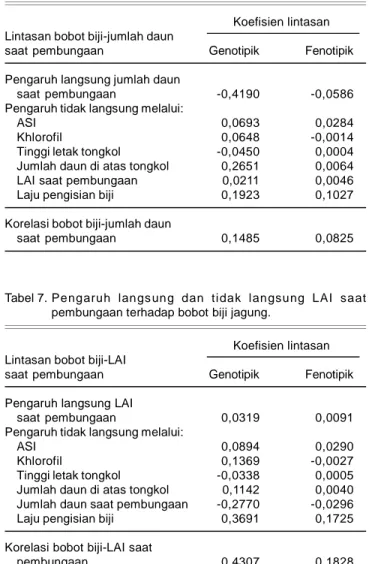
Pengaruh Lintasan Bobot Biji-ASI
ASI merupakan perbedaan antara umur keluarnya bunga jantan dengan bunga betina. Makin besar nilai ASI, kemungkinan terjadinya penyerbukan secara sempurna menjadi semakin kecil dan akibatnya tongkol tidak berbiji penuh. Korelasi fenotipik dan genotipik antara bobot biji dengan ASI memberikan arah yang sama, tetapi besarannya berbeda (Tabel 2). Korelasi genotipik dan fenotipik hampir sama dengan pengaruh langsungnya. Korelasi tersebut sebagian besar diperoleh dari kontribusi pengaruh langsung ASI secara genotipik dan fenotipik. Hal ini menunjukkan bahwa seleksi langsung terhadap ASI dapat memberikan respon terkorelasi terhadap hasil biji. Perbedaan korelasi genetik dan fenotipik nampaknya tidak disebabkan oleh gen
yang bersifat dominan. Hal ini menunjukkan sebagian besar dipengaruhi oleh gen nonaditif dan atau ling-kungan. Penduga derajat dominansi ASI tidak memiliki derajat dominan. Dengan demikian kemungkinan dipengaruhi oleh faktor lingkungan dan gen nonaditif selain dominan.
Pengaruh Lintasan Bobot Biji-Khlorofil Khlorofil daun mempengaruhi laju fotosintesis tanaman, semakin hijau daun diharapkan semakin efisien proses fotosintesis. Korelasi bobot biji dengan khlorofil daun antara fenotipik dan genotipik memberikan arah dan besaran yang hampir sama (Tabel 3). Hal ini menunjuk-kan adanya pengaruh gen aditif, tetapi bumenunjuk-kan karena pengaruh langsung.
Pengaruh tidak langsung secara genotipik adalah karakter tinggi letak tongkol, jumlah daun saat pembungaan, dan laju pengisian biji berkontribusi cukup besar, dalam hubungan ini yang direduksi oleh pengaruh langsung khlorofil. Pengaruh tidak langsung dari laju pengisian biji terhadap fenotipik bobot biji cukup besar, pengaruh langsungnya sangat kecil.
Tabel 1. Nilai dugaan rasio ragam dominan/aditif (2D/ 2
A) untuk
karakter jagung pada pemupukan takaran rendah. Karakter tanaman 2D/
2
A Derajat dominansi
Bobot biji (g/tanaman) 0,30 Parsial dominan
ASI 0,09 Parsial dominan
Khlorofil 0,00
-Tinggi letak tongkol 0,56 Parsial dominan Jumlah daun di atas tongkol 0,78 Parsial dominan LAI saat pembungaan 1,07 Dominan lengkap Jumlah daun saat pembungaan 0,28 Parsial dominan
Laju pengisian biji 0,03
-Tabel 2. Pengaruh langsung dan tidak langsung ASI terhadap bobot biji jagung.
Koefisien lintasan Lintasan bobot biji-ASI
Genotipik Fenotipik Pengaruh langsung ASI -0,1855 -0,1047 Pengaruh tidak langsung melalui:
khlorofil -0,0057 0,0003
Tinggi letak tongkol 0,0078 -0,0001
Jumlah daun di atas tongkol -0,1396 -0,0035 Jumlah daun saat pembungaan 0,1566 0,0159
LAI saat pembungaan -0,0154 -0,0025
Laju pengisian biji -0,0367 -0,0014
Pengaruh langsung secara fenotipik dan genotipik berbeda dan berlawanan arah. Hal ini menunjukkan adanya pengaruh gen nonaditif dan atau lingkungan. Hasil pendugaan derajat dominansi dari khlorofil menunjukkan tidak ada dominansi. Oleh karena itu, hal tersebut kemungkinan disebabkan oleh faktor lingkungan. Nampaknya khlorofil daun lebih besar pengaruhnya sebagai indikator lingkungan daripada indikator nilai pemuliaan suatu genotipe (Banzinger and Laffite 1997). Dengan demikian karakter khlorofil daun kurang memberikan harapan sebagai karakter sekunder untuk seleksi.
Pengaruh Lintasan Bobot Biji-Tinggi Keberadaan Tongkol
Tinggi letak tongkol diduga berkontribusi terhadap bobot biji, karena berkaitan dengan jarak antara source dengan sink (tongkol). Korelasi genotipik dan fenotipik bobot biji dengan tinggi letak tongkol menunjukkan
besaran yang berbeda tetapi arahnya sama. Hal ini mengindikasikan gen aditif berperan dalam hubungan tersebut. Tetapi tinggi letak tongkol tidak berpengaruh langsung terhadap bobot biji secara genotipik dan fenotipik (Tabel 4). Besaran dan arah pengaruh langsung secara genotipik dan fenotipik dari tinggi letak tongkol berbeda. Hal ini menunjukkan adanya pengaruh gen nonaditif tinggi letak tongkol dan atau lingkungan yang cukup besar kontribusinya. Dari penduga derajat dominansi karakter tinggi letak tongkol, pengaruh gen nonaditif kemungkinan besar berasal dari aksi gen parsial dominan. Korelasi fenotipik sebagian besar berasal dari kontribusi pengaruh tidak langsung dari laju pengisian biji.
Pengaruh Lintasan Bobot Biji-Jumlah Daun di Atas Tongkol
Jumlah daun di atas tongkol sebagai indikator banyak-nya daun jagung yang menerima langsung sebagian besar sinar matahari dalam melakukan fotosintesis mempengaruhi bobot biji jagung. Seperti pada pengaruh lintasan bobot biji-tinggi letak tongkol, korelasi genetik dan fenotipik jumlah daun di atas tongkol, dengan bobot biji memberikan arah yang sama tetapi dengan besaran yang berbeda (Tabel 5). Hal ini menunjukkan adanya pengaruh gen aditif. Pengaruh langsung secara genotipik jumlah daun di atas tongkol terhadap bobot biji sangat besar kontribusinya, tetapi secara fenotipik tidak menunjukkan hal itu. Hal ini terjadi karena adanya pengaruh gen nonaditif dan atau lingkungan. Berdasarkan aksi gen, jumlah daun di atas tongkol sebagai parsial dominan. Secara fenotipik, korelasi jumlah daun di atas tongkol dengan bobot biji terutama disebabkan oleh adanya pengaruh tidak langsung dari laju pengisian biji.
Tabel 4. Pengaruh langsung dan tidak langsung tinggi letak tongkol terhadap bobot biji jagung.
Koefisien lintasan Lintasan bobot biji-tinggi
letak tongkol Genotipik Fenotipik
Pengaruh langsung tinggi tongkol -0,2280 0,0022 Pengaruh tidak langsung melalui:
ASI 0,0064 0,0034
Khlorofil 0,0915 -0,0024
Jumlah daun di atas tongkol 0,1062 0,0030 Jumlah daun saat pembungaan -0,0827 -0,0107
LAI saat pembungaan 0,0047 0,0022
Laju pengisian biji -0,3657 -0,1988 Korelasi bobot biji-tinggi
letak tongkoI -0,4676 -0,2011
Tabel 5. Pengaruh langsung dan tidak langsung jumlah daun di atas tongkol terhadap bobot biji jagung.
Koefisien lintasan Lintasan bobot biji-jumlah daun
di atas tongkol Genotipik Fenotipik
Pengaruh langsung jumlah daun
di atas tongkol 0,3591 0,0125
Pengaruh tidak langsung melalui:
ASI 0,0721 0,0292
Khlorofil 0,0173 0,0002
Tinggi keberadaan tongkol -0,0674 0,0005 Jumlah daun saat pembungaan -0,3093 -0,0300
LAI saat pembungaan 0,0101 0,0029
Laju pengisian biji 0,2789 0,1765
Korelasi bobot biji-jumlah daun
di atas tongkol 0,3609 0,1917
Tabel 3. Pengaruh langsung dan tidak langsung khlorofil terhadap bobot biji jagung.
Koefisien lintasan Lintasan bobot biji-klorofil
Genotipik Fenotipik Pengaruh langsung klorofil -0,1733 0,0110 Pengaruh tidak langsung melalui:
ASI -0,0061 -0,0025
Tinggi letak tongkol 0,1204 -0,0005
Jumlah daun di atas tongkol -0,0358 0,0002 Jumlah daun saat pembungaan 0,1566 0,0077
LAI saat pembungaan -0,0252 -0,0022
Laju pengisian biji 0,1103 0,1409
Pengaruh Lintasan Bobot Biji-Jumlah dan LAI saat Pembungaan
Jumlah daun dan L AI saat pembungaan sebagai indikator kapasitas source yang dimiliki tanaman. Produktivitas tanaman dipengaruhi oleh kapasitas source tanaman. Pengaruh langsung secara genotifik dan fenotipik dari jumlah daun saat pembungaan negatif dan berlawanan arah dengan korelasi nya dengan bobot biji (Tabel 6). Pengaruh langsung secara genotipik lebih besar daripada secara fenotipik mengindikasikan adanya hubungan negatif dari pengaruh gen nonaditif dan atau lingkungan (Kang et al. 1983). Berdasarkan derajat dominansi dari jumlah daun saat pembungaan memiliki aksi gen parsial dominan. Laju pengisian biji memiliki pengaruh tidak langsung yang cukup besar dalam mewujudkan hubungan pengaruh jumlah daun saat pembungaan dengan bobot biji.
Pengaruh langsung LAI saat pembungaan, baik secara fenotipik maupun genotipik, relatif kecil kontribusinya terhadap korelasi fenotipik dan genotipik (Tabel 7). Korelasi genotipik yang lebih besar daripada korelasi fenotipik menunjukkan adanya hubungan negatif aksi gen parsial dominan dan atau lingkungan. Aksi gen LAI saat pembungaan diduga bersifat dominan lengkap.
Pengaruh Lintasan Bobot Biji-Laju Pengisian Biji Korelasi fenotipik (0,9768) dan genotipik (0,9561) antara laju pengisian biji dengan bobot biji memiliki besaran dan arah yang sama (Tabel 8). Korelasi fenotipik dihasilkan oleh pengaruh langsung secara genotipik maupun fenotipik. Berdasarkan penduga derajat dominansi, laju pengisian biji tidak memiliki dominansi, sehingga lebih dipengaruhi oleh gen aditif.
Hasil analisis lintasan memberikan petunjuk karakter sekunder ASI, dan laju pengisian biji dapat diper-timbangkan sebagai karakter untuk seleksi jagung adaptif pada kondisi pemupukan takaran rendah. Pengaruh sisa dari analisis lintasan dengan karakter-karakter sekunder tersebut diperoleh 2,6% untuk genotipik dan 18,5% untuk fenotipik. Hal ini berarti sekitar 97,4% secara genotipik dan 81,5% secara fenotipik dapat diterangkan oleh karakter-karakter sekunder yang diamati dalam peenelitian ini.
KESIMPULAN DAN SARAN
Derajat dominansi bobot biji pada pemupukan takaran rendah dari populasi varietas Bisma adalah parsial dominan dengan rasio ragam dominan/aditif sebesar 0,3.
Tabel 6. Pengaruh langsung dan tidak langsung jumlah daun saat pembungaan terhadap bobot biji jagung.
Koefisien lintasan Lintasan bobot biji-jumlah daun
saat pembungaan Genotipik Fenotipik
Pengaruh langsung jumlah daun
saat pembungaan -0,4190 -0,0586
Pengaruh tidak langsung melalui:
ASI 0,0693 0,0284
Khlorofil 0,0648 -0,0014
Tinggi letak tongkol -0,0450 0,0004 Jumlah daun di atas tongkol 0,2651 0,0064
LAI saat pembungaan 0,0211 0,0046
Laju pengisian biji 0,1923 0,1027
Korelasi bobot biji-jumlah daun
saat pembungaan 0,1485 0,0825
Tabel 7. Pengaruh langs ung dan tidak langsung LAI saat pembungaan terhadap bobot biji jagung.
Koefisien lintasan Lintasan bobot biji-LAI
saat pembungaan Genotipik Fenotipik
Pengaruh langsung LAI
saat pembungaan 0,0319 0,0091
Pengaruh tidak langsung melalui:
ASI 0,0894 0,0290
Khlorofil 0,1369 -0,0027
Tinggi letak tongkol -0,0338 0,0005 Jumlah daun di atas tongkol 0,1142 0,0040 Jumlah daun saat pembungaan -0,2770 -0,0296
Laju pengisian biji 0,3691 0,1725
Korelasi bobot biji-LAI saat
pembungaan 0,4307 0,1828
Tabel 8. Pengaruh langsung dan tidak langsung laju pengisisn biji (LPB) terhadap bobot biji jagung.
Koefisien lintasan Lintasan bobot biji-LPB
Genotipik Fenotipik
Pengaruh langsung LAI 0,8331 0,9778
Pengaruh tak langsung melalui:
ASI 0,0082 0,0002
Khlorofil -0,0229 0,0016
Tinggi letak tongkol 0,1001 -0,0004 Jumlah daun di atas tongkol 0,1202 0,0023 Jumlah daun saat pembungaan -0,0967 -0,0062
LAI saat pembungaan 0,0141 0,0016
Korelasi fenotipik dan genotipik antara bobot biji dengan ASI dan laju pengisian biji memberikan arah yang sama dan sebagian besar diperoleh dari kontribusi pengaruh langsung.
Seleksi awal untuk pertanaman jagung adaptif pada pemupukan takaran rendah dapat dilakukan pada ASI yang lebih cepat dan laju pengisian biji yang tinggi.
Pengamatan laju pengisian biji yang dilakukan sejak saat pembungaan hingga menjelang panen dengan cara memotong tanaman kurang praktis. Karena itu diperlu-kan teknik seleksi yang dapat dilakudiperlu-kan pada fase pertumbuhan awal tanaman seperti melalui marka molekuler.
UCAPAN TERIMA KASIH
Ucapan terima kasih disampaikan kepada Saudara Sunarto, teknisi di Kebun Percobaan Cikeumeuh, Bogor, yang telah membantu penulis dalam pelaksanaan dan pengumpulan data percobaan.
DAFTAR PUSTAKA
Banziger, M. and H.R. Lafitte. 1997. Efficiency of secondary traits for improving maize for low-nitrogen target environments. Crop Sci. 37:1110-1117.
Chapman, S.C. and G.O. Edmeades. 1999. Selection improved drought tolerance in tropical maize population. II. Direct and correlated responses among secondar y traits. Crop Sci. 39:1315-1324.
Falconer, D.S. and T.F. Mackay. 1996. Introduction to quantitative genetics. Longman Group Ltd. Essex.
Hallauer AR and J.B.F.Miranda. 1981. Quantitatve genetic maize breeding. Iowa State Univ. Press. Ames.
Kang, M.S., J.D. Miller, and P.Y.P. Tai. 1983. Genetic and phenotypic analysis and heritability in sugarcane. Crop Sci. 23:643-647. Kearsey, M.J. and H.S. Pooni. 1996. The genetical analysis of
quantitative traits. Chapman & Hall. London.
Robinson, H.F., R.E. Comstock, and P.H. Harvey. 1955. genetic variance in open pollinated varieties of corn. Genetics 40:45-60.
Sidwell, R.J., E.L. Smith, and R.W. McNew. 1976. Inheritance and interrelationship of grain yield and selected yield-related traits in hard red winter wheat cross. Crop Sci. 16:650-654. Singh R.K. and B.D. Chaudar y. 1979. Biometrical methods in
quantitative genetic analysis. Kalyani Publ. New Delhi. Subandi. 1972. Estimate of genetic parameters in an exotic
population of corn (Zea mays L.) grown under different plant population densities. Dissertation. Faculty of the Graduate College Univ. of Nebraska. Lincoln.
Sutoro, A. Bari, Subandi, dan S.Yahya. 2007. Parameter genetik jagung populasi Bisma pada pemupukan yang berbeda. II. ragam dan korelasi genetik karakter sekunder. J. AgroBiogen 3(1):9-14.