DINAMIKA FOLIKEL DAN REPEATABILITAS
PERTUMBUHAN GELOMBANG FOLIKEL PADA SAPI
PERANAKAN ONGOLE (PO)
Follicular dynamic and repeatability of follicular wave
development in Peranakan Ongole (PO) Cattle
MUHAMMAD IMRON1, IMAN SUPRIATNA2, AMROZI2, MOHAMAD AGUS SETIADI2
1
Balai Embrio Ternak Cipelang, PO Box 485 Bogor 16004. Email: hibbanie@gmail.com
2
Bagian Reproduksi dan Kebidanan, Departemen Klinik Patologi dan Reproduksi, Fakultas Kedokteran Hewan, Institut Pertanian Bogor
Jln. Agatis, Kampus IPB Dramaga-Bogor 16680. ABSTRAK
MUHAMMAD IMRON, IMAN SUPRIATNA, AMROZI, MOHAMAD AGUS SETIADI. 2016. Dinamika folikel dan repeatabilitas pertumbuhan gelombang folikel pada sapi peranakan ongole (PO). JITV.
Perlakuan superovulasi pada sapi PO (Bos indicus) memiliki respon yang rendah dibandingkan dengan rumpun Bos taurus yang mungkin berhubungan dengan perbedaan dinamika folikel antara kedua rumpun sapi. Penelitian ini dilakukan untuk mengetahui dinamika folikel dan repeatabilitas pola pertumbuhan gelombang folikel pada sapi PO. Pada tahap awal, penelitian menggunakan 9 ekor sapi PO berumur 5 – 7 tahun. Pengamatan dilakukan dengan menggunakan ultrasonografi (USG) setiap hari. Hasil pengamatan menunjukkan sapi PO memiliki pola 3 (66%) dan 4 (34%) gelombang pertumbuhan folikel dalam satu siklus. Gelombang folikel pertama dari pola gelombang 3 dan 4 terlihat berturut-turut hari ke 0,4+0,9 dan 1,4+1,1 relatif terhadap terjadinya ovulasi. Gelombang kedua terjadi berturut-turut hari ke 9,8+1,5 dan 7,4+1,9 pada pola 3 dan 4 gelombang, relatif terhadap terjadinya ovulasi. Pola 3 gelombang memiliki durasi folikel dominan lebih panjang (11,6+1,5 hari) dibandingkan pola 4 gelombang (10+2,9 hari). Kecepatan pertumbuhan folikel dominan tidak berbeda antara pola 3 dan 4 gelombang folikel (0,87+0,23 dan 0,94+0.25 mm/hari). Diameter folikel ovulatori antara pola 3 dan 4 gelombang folikel tidak berbeda yaitu berturut-turut 12,24+0,71 dan 12,30+0,22 mm. Diameter CL juga tidak berbeda antara pola 3 dan 4 gelombang folikel, yaitu 18,94+0,47 dan 19,44+0,87 mm. Pengamatan repeatabilitas pola gelombang yang dilakukan menggunakan 6 ekor menunjukkan sapi PO memiliki repeatabilitas tinggi pada pola gelombang (0,88) dan jumlah folikel yang berkembang (0,91). Penelitian ini menunjukkan data dinamika perkembangan folikel, pola gelombang dan repeatabilitasnya yang diharapkan membantu mendesain protokol perlakuan superovulasi atau teknologi reproduksi yang lain berbasis dinamika folikel sapi PO agar memperoleh respon yang lebih baik.
ABSTRACT
MUHAMMAD IMRON, IMAN SUPRIATNA, AMROZI, MOHAMAD AGUS SETIADI. 2016. Follicular dynamic and repeatability of follicular wave development in Peranakan Ongole (PO) Cattle. JITV
Superovulation treatment on PO cattle (Bos indicus) was less responsive compared to Bos taurus cattle breed may due to the difference of their follicular dynamic. This study was conducted to investigate the follicular dynamics and its repeatability in PO cattle. Follicular dynamics observations conducted on 9 cows trough ultrasound scanning every day. Observations of wave patterns repeatability were performed using 6 cows which already wave pattern known on the next consecutive IOI. The result indicated that PO cattle had 3 (66%) and 4 (34%) waves pattern. The first wave of 3 and 4 wave pattern emerged on day -0,4+0,9 and 1,4+1,1 respectively. The second wave of 3 and 4 wave pattern emerged on day 9,8+1,5 and 7,4+1,9 respectively. The pattern of 3 wave has a longer follicle dominance duration (11,6+1,5 day) in the first wave of estrous cycle, compared with 4 wave pattern (10+2,92 and 7+1.00 day respectively). The growth rate of dominant follicle were not different significantly between 3 and 4 waves patterns (0,87+0,23 and 0,94+0,25 mm/day). Similarly, ovulatory follicle diameter between 3 and 4 waves patterns also not significant different, 12,24+12,34 and 12,30+12,23 mm respectively. Observations of wave patterns repeatability in 6 PO cows indicate that PO cattle had high repeatability in follicular wave pattern (0,88) and number of growing follicle (0,91). This study resulted data for dynamic of follicular development, wave pattern, its repeatability which be expected to redesign the protocol for superovulation treatment or others based on follicular dynamic to improve its result in PO cattle. Keyword: PO cattle, follicular wave, repeatability, interovulatory interval
Pendahuluan
Gelombang folikel dicirikan dengan pertumbuhan sekelompok folikel kecil dengan diameter 3-4 mm (Taylor dan Rajamahendran 1991). Pola pertumbuhan gelombang folikel ini dapat diamati pada periode prapubertal (Melvin et al. 1999), selama periode kebuntingan (Taylor dan Rajamahendran 1991), setelah beranak (Murphy et al. 1990) dan selama siklus berahi (Roche et
al. 1999). Pertumbuhan gelombang folikel didahului dengan peningkatan
konsentrasi serum FSH dan diikuti dengan pertumbuhan 8-41 folikel kecil secara tiba-tiba dalam waktu 2-3 hari (Ginther et al. 1997). Satu folikel akan terpilih untuk terus tumbuh menjadi folikel dominan (FD) sedangkan folikel lain yang
lebih kecil disebut folikel subordinat (FS) dan akan mengalami atresia. Jika terjadi luteolisis pada saat FD dalam fase pertumbuhan, maka folikel akan mengalami proses pematangan final dan dilanjutkan dengan ovulasi. Jika tidak ada luteloisis selama FD sedang tumbuh, maka FD juga akan mengalami atresia (Vasenna et al. 2003, Jaiswal et al. 2004, Adam et al. 2008).
Mayoritas rumpun sapi menunjukkan pola dua dan tiga gelombang folikel dalam satu siklus berahi (Adam et al. 2008) walaupun terkadang dilaporkan juga adanya pola satu, empat dan lima gelombang folikel (Bleach et al. 2004, Viana et
al. 2000). Dinamika folikel merupakan salah satu subyek penelitian yang banyak
dilakukan pada sapi rumpun eropa (B. taurus) namun sangat terbatas informasi yang diperoleh dari sapi zebu (B. indicus). Karakteristik reproduksi sapi B.
indicus juga tidak identik dengan B. taurus antara lain dalam hal panjang siklus
berahi, waktu berahi, tingkah laku berahi, pertumbuhan folikel dominan dan CL, sudah direview oleh Bó et al. (2003).
Sementara itu, ada permasalahan dalam program superovulasi di BET Cipelang yang dilakukan pada donor sapi PO (Bos indicus) yaitu menghasilkan rasio embrio layak transfer per total hasil koleksi embrio yang lebih rendah (1.5 embrio) dibandingkan dengan sapi dari rumpun Bos Taurus antara lain Angus, Simmental dan Limousin yang memiliki rasio berturut turut 4.75, 3.37 dan 2,96 embrio (Anonimous, 2012). Pelaksanaan program penyuntikan hormon dalam perlakuan superovulasi tidak dibedakan antara donor dari rumpun Bos taurus (Simmental, Limousin) dan donor dari rumpun Bos indicus (sapi PO). Bó dan Mapletoft (2014) menyatakan respons superovulasi akan menjadi optimal jika perlakuan superstimulasi folikel dimulai pada saat awal terjadinya gelombang
pertumbuhan folikel. Perlakuan gonadotropin yang dimulai lebih cepat atau lebih lambat satu hari dari terjadinya awal gelombang folikel, akan mengurangi respons superovulasi dibandingkan dengan memulai perlakuan tepat pada saat terjadinya awal gelombang folikel (Bó et al., 2008). Diduga ada kaitan antara perbedaan karakteristik reproduksi antara kedua rumpun sapi ini terutama dalam hal pola gelombang folikel dengan kejadian kendala program superovulasi pada sapi PO. Karena itu informasi karakteristik dinamika ovarium dan repeatabilitasnya diperlukan agar program superovulasi pada sapi PO menjadi lebih optimal.
Penelitian ini dilakukan untuk mengetahui karakteristik dinamika ovarium selama siklus berahi dan tingkat repeatabilitas pola pertumbuhan gelombang folikel pada sapi Peranakan Ongol (PO).
Materi dan Metoda
Materi Penelitian
Penelitian ini menggunakan sapi PO betina dewasa sebanyak 9 ekor yang berumur antara 5 sampai 7 tahun dengan berat badan antara 375 sampai 450 kg. Semua sapi yang digunakan memiliki Nilai Kondisi Tubuh (NKT) antara 2,7 sampai 3,2 dalam skala penilaian 1 sampai 5, dengan siklus birahi yang normal. Pakan diberikan sebanyak 30 sampai 40 kg rumput per ekor per hari dan ditambah dengan 2 sampai 3 kg pakan konsentrat per ekor per hari.
Sinkronisasi Ovulasi
Penelitian ini dimulai dengan sinkronisasi ovulasi untuk menyeragamkam awal pengamatan dinamika ovarium melalui pemasangan preparat progesteron intravaginal (Cuemate®, mengandung 1.56 mg progesteron dalam dua pod silicon, Bioniche Animal Health (A/Asia) Pty.Ltd., Australia) disertai penyuntikan 100 µg
GnRH (Fertagyl®, Intervet Schering-Plough Animal Health, Jerman) pada hari pertama. Cuemate® dimasukkan kedalam vagina selama 7 hari menggunakan aplikator khusus yang diberi pelumas gel isotonik. Penyuntikan PGF2α (Prostavet® C, 5 mg of etiproston per 2 ml of solution; Virbac Animal Health, Prancis) dilakukan secara intramuskular bersamaan dengan pelepasan Cuemate® dari dalam vagina dan dua hari kemudian diberikan 100 µg GnRH (Fertagyl®, Intervet Schering-Plough Animal Health, Jerman) secara intramuskular untuk menginduksi sinkronisasi ovulasi.
Pengamatan dinamika folikel
Pengamatan dinamika folikel dilakukan satu hari sekali dengan waktu dan operator yang sama mengunakan ultrasound portabel (EasyScan Lite, Inggris) yang dilengkapi dengan monitor goggle dan probe dengan frekuensi dinamis 4 – 8 Mhz. Unit utama ultrasound dihubungkan dengan komputer dan data pengamatan direkam dalam format video. Pengukuran diameter folikel dan CL dilakukan menggunakan Microsoft image tool. Pengamatan harian menggunakan ultrasound dimulai sejak sapi PO diberikan suntikan PGF2α. Satu interovulatory interval (IOI) didefinisikan sebagai periode waktu antara satu ovulasi dengan ovulasi berikutnya. Ovulasi pertama yang diamati setelah perlakuan sinkronisasi ovulasi dianggap sebagai hari ke-0 dalam satu IOI.
Pada tahap awal, pengamatan dilakukan untuk mendapatkan karakteristik dinamika folikel dan memetakan pola gelombang dalam satu IOI. Sapi yang digunakan dalam penelitian tahap awal sebanyak 9 ekor. Sedangkan untuk mendapatkan data repeatabilitas pertumbuhan gelombang folikel, pengamatan dinamika folikel dilanjutkan pada satu IOI berikutnya tanpa jeda menggunakan 6
ekor sapi yang telah diketahui pola jumlah gelombangnya, terdiri dari 3 ekor pola 3 gelombang dan 3 ekor pola 4 gelombang.
Prosedur Analisa Data
Satu gelombang folikel didefinisikan sebagai durasi pertumbuhan folikel mulai diameter sekitar 4 mm sampai folikel dominan kembali mengecil keukuran semula. Folikel yang mencapai ukuran diameter terbesar dari gelombang yang sama dianggap sebagai folikel dominan (FD), sedangkan folikel yang lebih kecil sebagai folikel subordinat (FS). Fase pertumbuhan folikel adalah periode folikel terdeteksi pada diameter 4 mm sampai hari saat diameter folikel tersebut tidak bertambah besar. Fase regresi dimulai pada saat folikel terdeteksi mengalami pengurangan diameter sampai folikel mulai sulit terdeteksi secara individual. Fase statis didefinisikan sebagai periode antara hari terakhir fase pertumbuhan dan hari pertama fase regresi. Data kemudian dikelompokkan berdasarkan pola jumlah gelombang folikel yang teramati dalam satu IOI, yaitu kelompok pola 3 dan 4 gelombang folikel dan diuji dengan Independent-Sample T Test. Repeatabilitas (nilai antara 0 sampai 1) didefinisikan sebagai proporsi ragam total yang dapat berkontribusi terhadap ragam individu, dihitung dengan rumus σ2 individu / (σ2 individu + σ2
galat) (Burns et al. 2005). Komponen dari nilai ragam untuk repeatabilitas diestimasi menggunakan ANOVA dengan faktor tunggal. Persentase terjadinya ovulasi folikel merupakan jumlah kejadian ovulasi pada ovarium kiri atau kanan dibagi dengan total kejadian ovulasi folikel yang teramati. Persentase terjadinya gelombang folikel pada ovarium merupakan jumlah
gelombang folikel yang terjadi pada ovarium kanan atau kiri dibagi dengan jumlah total gelombang folikel yang teramati.
Hasil dan Pembahasan
Berdasarkan pengamatan harian dinamika folikel menggunakan ultrasound terhadap 9 ekor sapi PO, 6 ekor (66%) menunjukkan pola 3 gelombang folikel dan 3 ekor (33%) menunjukkan pola 4 gelombang folikel (Gambar 1) dan tidak ada pola 2 gelombang yang teramati dalam penelitian ini. Melia et al. (2014) juga melaporkan bahwa sapi PO menunjukkan pola 3 gelombang folikel pada 6 ekor sapi yang diamati. Pada penelitian lain Viana et al. (2000) melaporkan hanya 1 dari 15 ekor (6%) sapi Gyr (B. indicus) memiliki pola 2 gelombang folikel, sedangkan 60% dan 26.7% berturut-turut memiliki pola 3 dan 4 gelombang folikel.
Jumlah gelombang dalam satu siklus dilaporkan tidak dipengaruhi oleh rumpun sapi (Adam et al. 2008), namun Bó et al. (2003) melaporkan terdapat peningkatan pola 3 gelombang pada sapi dengan kualitas nutrisi yang buruk dan stress karena panas. Pada sapi perah tidak ada perbedaan terkait proporsi pola gelombang antara sapi muda dan dewasa (Wolfenson et al. 2004). Tetapi dalam penelitian lain Adam et al. (2008) menyatakan mayoritas sapi dara (65%) pada sapi Nellor (B. indicus) menunjukkan pola 3 gelombang sedangkan mayoritas sapi induk (83%) menunjukkan pola 2 gelombang. Selanjutnya Satheshkumar et al. (2015) melaporkan pola 3 gelombang lebih banyak terjadi pada musim dingin dibandingkan pola 2 gelombang, dan sebaliknya terjadi pada musim panas.
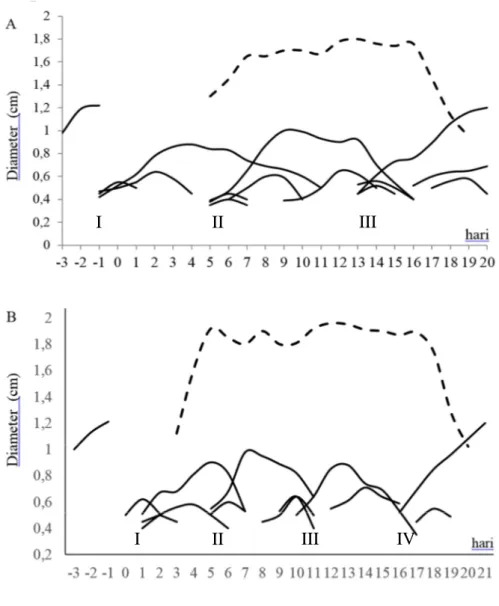
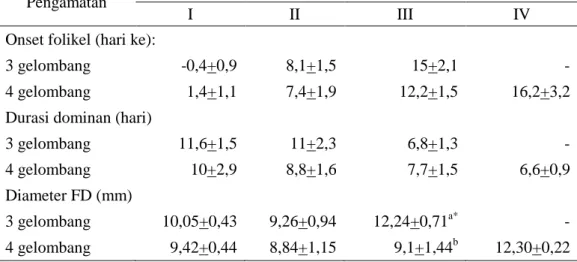
Gambar 1. Pola perkembangan gelombang folikel dan CL pada sapi PO dengan 3 gelombang folikel (A) dan 4 gelombang folikel (B). Keterangan: garis putus-putus adalah pola pertumbuhan CL, garis solid adalah pola pertumbuhan folikel
Pertumbuhan gelombang folikel pertama mulai terjadi hari ke -0,4+0,9 dan 1,4+1,1 relatif terhadap terjadinya ovulasi pada sapi yang memiliki pola gelombang 3 dan 4. Gelombang folikel berikutnya mulai tumbuh pada 8,1+1,5 dan 7,4+1,9 untuk pola gelombang 3 dan 4 (Tabel 1). Pola 3 gelombang memiliki durasi dominan yang paling panjang (11,6+1,5 hari) pada gelombang pertama dan lebih pendek pada pola 4 gelombang yaitu 10,0+2,9. Hasil ini sesuai dengan Adam et al. (2008) yang menyatakan terdapat korelasi yang tinggi antara durasi dominan dengan pola jumlah gelombangnya, yaitu semakin pendek durasi
I II III
dominan akan menambah jumlah gelombang dalam satu IOI. Dalam penelitian lain Jaiswal et al. (2009) menyatakan durasi dominan pada gelombang pertama dalam IOI dapat digunakan untuk memprediksi pola jumlah gelombang.
Tabel 1. Karakteristik per gelombang folikel dominan pada sapi PO dengan pola 3 dan 4 gelombang folikel pada satu IOI.
Pengamatan Gelombang Folikel
I II III IV
Onset folikel (hari ke):
3 gelombang -0,4+0,9 8,1+1,5 15+2,1 -
4 gelombang 1,4+1,1 7,4+1,9 12,2+1,5 16,2+3,2
Durasi dominan (hari)
3 gelombang 11,6+1,5 11+2,3 6,8+1,3 -
4 gelombang 10+2,9 8,8+1,6 7,7+1,5 6,6+0,9
Diameter FD (mm)
3 gelombang 10,05+0,43 9,26+0,94 12,24+0,71a* -
4 gelombang 9,42+0,44 8,84+1,15 9,1+1,44b 12,30+0,22
Huruf superscript yang berbeda pada kolom dan kriteria yang sama, berbeda nyata pada P<0.05. *: Diameter folikel ovulatori .
Diameter folikel dominan (FD) masing-masing gelombang folikel pada pola 3 dan 4 tidak berbeda nyata walaupun terdapat kecenderungan diameter folikel dominan semakin mengecil dengan bertambahnya jumlah gelombang dalam satu IOI (Tabel 1). Viana et al. (2000) menyatakan penurunan diameter folikel dominan dan durasi dominannya diasosiasikan dengan peningkatan jumlah gelombang dalam satu siklus berahi. Dalam penelitian lain Bó et al. (2003) melaporkan banyaknya jumlah sapi B. indicus yang memiliki pola 4 gelombang (16%) dibandingkan dengan B. taurus (0%) diduga merupakan konsekuensi diameter folikel dominan yang lebih kecil dan durasi dominan yang lebih pendek pada B. indicus. Estradiol dengan konsentrasi tinggi menekan impuls LH yang menginduksi sintesa progesteron dari sel-sel luteal (Goodman et al. 1981, William
et al. 1978), sehingga diduga dapat mempengaruhi fungsi luteal dan masa
hidupnya (Jaiswal et al. 2009).
Namun pada gelombang ketiga dari pola 3 gelombang yang merupakan folikel ovulatori memiliki diameter nyata lebih besar dibandingkan dengan gelombang ketiga dari pola 4 gelombang (Tabel 1). Demikian pula diameter folikel ovulatori lebih lebih besar dibandingkan folikel non ovulatori pada pola gelombang yang sama. Progesteron yang dihasilkan oleh CL akan menekan konsentrasi estradiol dan kecepatan pertumbuhan folikel dominan (Ramana et al. 2013, Junior et al. 2010), sehingga mengakibatkan folikel yang tumbuh selama fase luteal memiliki diameter yang lebih kecil dibandingkan dengan folikel ovulatori yang tumbuh pada saat terjadi luteolisis.
Siklus satu IOI dalam penelitian ini juga cenderung lebih panjang dengan bertambahnya pola gelombang yaitu 22+1,6 dan 22,8+2,9 hari berturut-turut pada pola 3 dan 4 (Tabel 2), yang mungkin juga berhubungan dengan ukuran diameter folikel pada masing-masing pola gelombang. Goff et al. (2004) menyatakan estradiol pada gelombang pertama IOI meregulasi waktu munculnya reseptor oxytosin dalam endometrium yang akhirnya mengatur waktu produksi prostaglandin untuk memicu luteolisis. Sehingga ada dugaan bahwa ukuran diameter folikel dominan pada gelombang pertama IOI mempengaruhi pola jumlah gelombang folikel yang terbentuk (Adam et al. 2008, Jaiswal et al. 2009). Namun Boer et al. (2011) menyatakan mekanisme terbentuknya pola gelombang masih belum jelas dan diduga melibatkan regulasi pertumbuhan folikel yang lebih komplek.
Tabel 2. Karakteristik dinamika folikel dominan, folikel sub dominan, dan corpus luteum (CL) pada sapi PO yang memiliki 3 dan 4 gelombang folikel.
Karakteristik 3 gelombang 4 gelombang
Folikel Dominan
Jumlah IOI 7 5
Panjang IOI (hari) 22,0+1,6 22,8+2,9
Kecepatan pertumbuhan (mm/hari) 0,87+0,23 0,94+0,25
Fase statis (hari) 3,7+2,0 2,7+1,9
Kecepatan atresia (mm/hari) 0,83+0,21 O,91+0,31
Pertumbuhan folikel sampai ovulasi (hari) 7,6+0,6 7,4+1,1
Diameter folikel ovulatori (mm) 12,24+0,71 12,30+0,22
Diameter saat deviasi (mm) 6,25+0,37 6,44+0,44
Kecepatan pertumbuhan folikel FS (mm/hari) 0,83+0,25 0,72+0,33
Diameter CL hari ke-10 (mm) 18,94+0,47 19,44+0,87
Dalam penelitian ini, kecepatan pertumbuhan antara folikel dominan (FD) tidak berbeda nyata antara pola 3 dan 4 gelombang (0,87+0,23, 0,94+0,25 mm/hari) (Tabel 2). Demikian pula kecepatan pertumbuhan FD tidak berbeda nyata dengan folikel subdominannya (FS) sampai terjadi deviasi folikel. Hasil ini sesuai dengan Adam et al. (2008) yang menyatakan bahwa sekelompok folikel pada awal pertumbuhan folikel memiliki kecepatan pertumbuhan yang sama sampai terpilih satu folikel untuk terus tumbuh dan menjadi FD sedangkan yang lain mengalami atresia.
Awal dari perbedaan pertumbuhan antar dua folikel terbesar dinamakan deviasi folikel (Ginther et al. 2003). Dalam penelitian ini deviasi terjadi setelah FD terpilih mencapai diameter 6,25+0,37 dan 6,44+0,44 mm pada pola 3 dan 4 gelombang (Tabel 2), lebih kecil dibandingkan dengan deviasi folikel pada sapi B.
taurus yang mencapai diameter 8,5-9,0 mm (Ginther dan Hoffman et al. 2014,
melaporkan sapi Nellor (B. indicus) memiliki diameter folikel saat deviasi dan ovulatori lebih kecil dibandingkan B. taurus. Sapi rumpun B. indicus memiliki diameter folikel dominan yang lebih kecil dibandingkan sapi B. Taurus sehingga diameter folikel pada saat deviasi folikel juga menjadi lebih kecil pada B. indicus (Bó et al. 2003).
Pada saat FD menjadi tergantung pada LH, maka hanya ada dua kemungkinan yang terjadi yaitu ovulasi atau mengalami regresi (Lucy 2007). FD yang tidak terpapar LH surge akan tetap bertahan pada periode waktu tertentu (fase statis) dan kemudian akan mengalami regresi (Valdez et al. 2005). Selama periode ini, FD akan tergantung pada growth factor yang mendukung transisi fase G1 ke fase S dari siklus sel dan mencegah apoptosis dalam sel-sel granulosa (Quirk et al. 2004). Pada saat tertentu dalam fase statis, FD akan kehilangan fungsi dominannya walaupun folikel ini secara morfologi masih dominan, yaitu memiliki ukuran paling besar (Ireland et al. 2000). Dalam penelitian ini sapi dengan pola 3 gelombang memiliki fase statis yang lebih panjang yaitu 3,7+1,9 hari dibandingkan dengan 4 gelombang (1,9+1,4 hari).
Folikel ovulatori antara pola 3 dan 4 gelombang memiliki diameter yang tidak berbeda nyata, yaitu berturut-turut 12,24+0,34 dan 12,30+0,23 mm (Tabel 2). Lama pertumbuhan folikel sampai ovulasi juga tidak berbeda antara pola 3 dan 4 gelombang yaitu 7,6+0,5 dan 7,4+1,1 hari (Tabel 2). Demikian pula diameter CL hari ke-10 setelah ovulasi juga tidak berbeda nyata antara pola 3 dan 4 gelombang yaitu 18,94+0,47 dan 19,44+0,87 mm.
Pola jumlah gelombang dalam penelitian ini memiliki nilai repeatabilitas yang tinggi yaitu 0,88 pada individu yang sama (Tabel 3). Hasil ini sesuai dengan
penelitian Jaiswal et al. (2009) yang menyatakan pola gelombang memiliki nilai repeatabilitas tinggi pada sapi persilangan Hereford (B. Taurus). Namun pada penelitian lain Sichtar et al. (2010) melaporkan proporsi yang hampir sama antara yang mengalami perubahan pola jumlah gelombang maupun yang tidak dalam individu yang sama pada sapi perah.
Jumlah folikel yang berkembang dalam penelitian ini juga memiliki nilai repeatabilitas yang tinggi pada individu yang sama yaitu 0,91. Beberapa penelitian yang lain juga melaporkan bahwa jumlah folikel yang direkrut kedalam satu gelombang memiliki repeatabilitas yang tinggi pada individu yang sama, tapi sangat bervariasi pada individu yang berbeda (Santos et al. 2014, Ireland et al. 2007, David et al. 2005). Pada penelitian yang lain Singh et al. (2004) melaporkan bahwa jumlah folikel antral yang tinggi memiliki korelasi positif terhadap perlakuan gonadotropin selama superovulasi dan menghasilkan jumlah oosit dan embrio layak transfer yang lebih banyak. Dengan demikian karakteristik repeatabilitas jumlah folikel yang tinggi pada sapi PO mungkin dapat digunakan sebagai parameter untuk seleksi donor dalam rangka meningkatkan efektifitas program superovulasi.
Tabel 3. Nilai beberapa karakteristik pengamatan dinamika folikel pada sapi PO
No Pengamatan Nilai
1 Repeatabilitas pola gelombang folikel (3 dan 4) berturut-turut
dalam dua IOI pada individu yang sama 0,88
2 Repeatabilitas jumlah folikel yang berkembang pada individu
yang sama 0,91
3 Rata-rata jumlah folikel pada awal pertumbuhan gelombang
folikel 27,3+9,4
4 Persentase terjadinya ovulasi pada ovarium kanan dan kiri 72% dan
28%
5 Persentase terjadinya gelombang folikel pada ovarium kanan
dan kiri
52% dan 48%
6 Repeatabilitas terjadinya gelombang folikel pada ovarium
kanan dan kiri
0,14 dan 0,10
Pada penelitian ini, kejadian ovulasi pada ovarium kanan lebih besar (72%) dibandingkan dengan kiri (28%) (Tabel 3), konsisten dengan penelitian yang melaporkan bahwa ovulasi lebih banyak terjadi pada ovarium kanan dibandingkan dengan yang kiri (Vasenna et al. 2003, Nation et al. 1999). Tetapi pada penelitian lain Purwantara et al. (2006) dan Ginther et al. (1989) melaporkan kejadian ovulasi yang berimbang pada ovarium kanan atau kiri. Dilain pihak, persentase jumlah gelombang folikel yang terbentuk antara ovarium kanan dan kiri hampir seimbang, yaitu 52% versus 48 % yang menunjukkan bahwa sebenarnya ovarium kanan dan kiri memiliki keaktifan yang sama dalam proses folikulogenesis, walaupun ovulasi lebih banyak terjadi pada ovarium kanan.
Selain itu repeatabilitas terjadinya gelombang folikel pada ovarium yang sama sangat kecil, baik pada ovarium kanan dan kiri (0,14 vs 0,10) yang menunjukkan proses terbentuknya gelombang folikel terjadi secara acak pada ovarium kanan maupun kiri. Hasil ini berbeda dengan Vasenna et al. (2003) yang melaporkan aktifitas ovarium kanan lebih tinggi dibandingkan yang kiri. Ginther
et al. (1989) menyatakan FD mempengaruhi pertumbuhan folikel subordinatnya
lebih dengan mekanisme sistemik endokrin baik pada ovarium kanan dan kiri. Hanya satu folikel dari sepasang ovarium yang akan terseleksi menjadi FD, baik pada posisi ipsilateral maupun kontralateral terhadap folikel subordinat terbesar.
Penelitian ini memberikan data dasar dinamika folikel dan repeatabilitasnya pada sapi PO berbasis pengamatan dengan USG. Pemahaman yang lebih baik tentang dinamika folikel sangat membantu dalam pemanfaatannya untuk keperluan yang lebih spesifik manajemen reproduksi seperti dalam program superovulasi. Beberapa percobaan di lapangan telah dilakukan untuk
menerapkan program superovulasi pada sapi rumpun lokal yang lain, antara lain sapi Bali, sapi Madura dan sapi Aceh dengan protokol superovulasi standar seperti yang diterapkan pada sapi rumpun Bos taurus. Namun sampai saat ini, respon superovulasi yang dihasilkan belum sesuai dengan harapan. Hal ini mengindikasikan urgensi pemahaman terhadap karakteristik dinamika folikel pada masing-masing rumpun sapi lokal yang sampai saat ini belum banyak diungkapkan, agar dapat digunakan sebagai data dasar dalam acuan penerapan teknologi reproduksi, khususnya program superovulasi.
Simpulan dan Saran
Penelitian ini menunjukkan karakteristik dinamika folikel pada sapi PO yang didominasi oleh pola 3 gelombang folikel dalam satu siklus berahi. Repeatabilitas pola gelombang dan jumlah folikel memiliki nilai tinggi pada individu sama sehingga karakteristik ini mungkin dapat digunakan untuk manajemen reproduksi sapi donor dalam rangka meningkatkan efektifitas program superovulasi berbasis informasi individu.
Daftar Pustaka
Adams GP. 1999. Comparative patterns of follicle development and selection in ruminants. J Reprod Fertil. (Suppl). 54:17–32.
Adams GP, Jaiswal R, Singh J Malhi P. 2008. Progress in understanding ovarian follicular dynamics in cattle. Theriogenology 69:72–80
Anonimous. 2012. Laporan Tahunan: Evaluasi produksi dan transfer embrio. Balai Embrio Ternak. 31-36
Bleach, ECL, RG. Glencross, PG. Knight. 2004. Association between ovarian follicle development and pregnancy rates in dairy cows undergoing spontaneous oestrous cycles. Reproduction. 127:621–629.
Boer HMT, Röblitz L, Stötzel C, Veerkamp RV, Kemp B, Woelders H. 2011. Mechanisms regulating follicle wave patterns in the bovine estrous cycle investigated with a mathematical model. J Dairy Sci. 94:5987–6000
Bó GA, Baruselli PS, Mart MF. 2003. Pattern and manipulation of follicular development in Bos indicus cattle. Anim Reprod Sci.78:307–326
Beg MA, Bergfelt DR, Kot K, Ginther OJ. 2002. Follicle selection in cattle: dynamics of follicular fluid factors during development of follicle dominance.
Biol Reprod. 66: 120-126
Burns DS, Jimenez-Krassel FJ, Ireland JJ. 2005. Numbers of antral follicles during follicular waves in cattle: evidence for high variation among animals, very high repeatability in individuals, and an inverse association with serum follicle-stimulating hormone concentrations. Biol Reprod 73:54–62.
David SB, Jimenez-Krassel F, Ireland JLH, Knight PG, James J. 2005. Numbers of Antral Follicles During Follicular Waves in Cattle: Evidence for High Variation Among Animals, Very High Repeatability in Individuals, and an Inverse Association with Serum Follicle-Stimulating Hormone Concentrations.
Biology of Reproduction. 73: 54–62.
Ereno RL, Pupulim AG, Loureiro B, Favoreto MG, Castilho ACS, Buratini J, Barros CM. 2013. The effect of nutrition onovarian follicle population and plasma anti-Mullerian hormone con-centration in Aberdeen Angus heifers (abstract). Reprod Fertil Dev. 25, 236.
Fair T, Hulshof SCJ, Hyttel P, Greve T, Boland M. 1997. Oocyte ultrastructure in bovine primordial to early tertiary follicles. Anat Embryo 195: 327–336.
Fortune JE, Cushman RA, Wahl CM, Kito S. 2000. The primordial to primary follicle transition. Mol. Cell Endocrinol. 163: 53-60
Ginther OJ, Knopf L, Kastelic JP. 1989. Temporal associations among ovarian events in cattle during oestrous cycles with two and three follicular waves. J Reprod Fertil. 87:223–30.
Ginther, O.J., Kot, K. Kulick, L.J., Wiltbank, M.C. 1997. Emergence and deviation of follicles during the development of follicular waves in cattle.
Theriogenology. 48(1): 75-87.
Ginther OJ, Hoffman MM. 2014. Intraovarian effect of dominant follicle and corpus luteum on number of follicles during a follicular wave in heifers.
Theriogenology 82 :169–175
Ginther OJ, Beg MA, Donadeu FX dan Bergfelt DR. 2003. Mechanism of follicle deviation in monovular farm species. Anim Reprod Sci. 78: 239 – 257.
Goodman RL, Bittman EL, Foster DL, Karsch FJ. 1981. The endocrine basis for the synergistic suppression of luteinizing hormone by estradiol and progesterone. Endocrinology. 109: 1414–7.
Goff AK. 2004. Steroid hormone regulation of prostaglandin secretion in the ruminant endometrium during the estrous cycle. Biol Reprod. 71:11–6.
Ireland JJ, M Mihm, E Austin, MG Diskin, JF Roche. 2000. Historical perspective of turnover of dominant follicles during the bovine estrous cycle: key concepts, studies, advancements, and terms: J Dairy sci. 83 1648-1658 Ireland JJ, Ward F, Jimenez-Krassel F, J Ireland JLH, Smith GW, Lonergan P dan
Evans ACO. 2007. Follicle numbers are highly repeatable within individual animals but are inversely correlated with FSH concentrations and the proportion of good-quality embryos after ovarian stimulation in cattle. Human
Reproduction. 22 (6): 1687–1695
Jaiswal RS, J. Singh, L. Marshal, GP Adam. 2009. Repeatability of 2-wave and 3-wave patterns of ovarian folicular development during the bovine estous cycle. Theriogenology. 71(1):81-90.
Junior CI, Sa Filho OG, RFG Perez, FHS Aono, ML Day, JLM Vasconcelos. 2010. Reproductive Performance of prepubertal Bos indicus herifer after progesteron-based treatments. Theriogenology. 74:903-911.
Kanitz W. 2003. Follicular dynamic and ovulation in cattle – a review. Arch
Tierz. 46(2): 187-198
Melia J, Amrozi, Tumbelaka LI. 2014. Dinamika ovarium sapi endometritis yang diterapi dengan gentamicine, flumequine dan analog prostaglandin F2 alpha (PGF2α) secara intra uterus. J kedokteran hewan. 8(2): 111-115.
Melvin EJ, Lindsey BR, Quintal-franco J, Zanella, Fike KE, Van Tassell CP, Kinder JE. 1999. Circulating concentrations of estradiol, luteinizing hormone, and follicle-stimulating hormone during waves of ovarian follicular development in prepubertal cattle. Biol Reprod 60: 405-412
Mihm, M., P.J. Baker, J.L.H. Ireland, G.W. Smith, P.M. Coussens, A.C.O. Evans and J.J. Ireland, 2006. Molecular evidence that growth of dominant follicles involves a reduction in follicle stimulating hormone dependence and an increase in luteinizing hormone dependence in cattle. Biol Reprod. 74: 1051 – 1059
Monniaux D, Barbey S, Rico C, Fabre S, Gallard Y, Larroque H, 2010. Anti-Müllerian hormone: a predictive marker of embryo productionin cattle? Reprod
Fertil Dev. 22: 1083–1091.
Mossa F, Walsh SW, Butler ST, Berry DP, Carter F, Lonergan P, Smith GW, Ireland JJ, Evans ACO. 2012. Low numbers of ovar-ian follicles ≥3 mm in diameter are associated with low fertility indairy cows. J Dairy Sci. 95: 2355– 2361.
Murphy MG, Boland MP, Roche JF. 1990. Pattern of follicular growth and resumption of ovarian activity in post-partum beef suckler cows. J Reprod
Fertil. 90: 523-533
Nation DP, Burke CR, Rhodes FM, Macmillan KL. 1999. The inter-ovarian distribution of dominant follicles is influenced by the location of the corpus luteum of pregnancy. Anim Reprod Sci. 56:169–176
Lucy MC. 2007. The bovine dominant ovarian follicle. J Anim Sci. 85:89-99. Lussier JG, Matton P, Dufour JJ. 1987. Growth rates of follicles in the ovary of
the cow. J Reprod Fertil. 81:301–307
Parrott JA, Skinner MK. 2000. Kit ligand actions on ovarian stromal cells: effects on theca cell recruitment and steroid production. Mol Reprod Dev. 55:55-64 Purwantara B, Hoier R, Schmidt M, Greve T. 2006. Preovulatori changes and
ovulation in cattle undergoing spontaneous or clopprostenol-induced luteolyis.
Biotropia. 13(2):75-84
Quirk SM, Cowan RG, Harman RM, Hu CL, Porter DA. 2004. Ovarian follicular growth and atresia: The relationship between cell proliferation and survival. J
Anim Sci. 82(Suppl): 40–52.
Ramana KV, Rao KV, Supriya K, Rajanna N. 2013. Postpartum ovarian follicular dynamics and estrus activity in lactating Ongole cows. J Res Angrau. 41(1): 51-55
Roche JF, Austin EJ. Ryan M, O'rourke M, Mihm M, Diskin MG. 1999. Regulation of follicle waves to maximize fertility in cattle. J Reprod Fertil. (Suppl) 54:61-71
Santos KC, Santos GMG, Siloto LS, Morotti F, Marcantonio TN, Seneda MM, 2014. Comparison of antral and preantral ovar-ian follicle populations between
Bos indicus and Bos taurus cows with high or low antral follicles counts. Reprod Dom Anim. 49,48–51.
Sartori R, Fricke PM, Ferreira JCP, Ginther OJ, Wiltbank MC. 2001. Follicular deviation and acquisition of ovulatory capacity in bovine follicles. Biol Reprod. 65: 1403–1409
Sartorelli ES, Carvalho LM, Bergfelt DR, Ginther OJ, Barros CM. 2005. Morphological characterization of follicle deviation in Nelore (Bos indicus) heifers and cows. Theriogenology. 63:2382–94.
Satheshkumar S, K. Brindha, A. Roya, T.G. Devanathan, D. Kathiresan, K. Kumanan. 2015. Natural influence of season on follicular, luteal, and endocrinological turnover in Indian crossbred cows. Theriogenology. 84:19–23 Sichtar, J., R. Tolman, R. Rajmon, P. Klabanová, P. Berka, and J. Volek. 2010. A
comparison of the follicular dynamics in heifers of the Czech Fleckvieh and Holstein breeds. Czech J Anim Sci. 55:234–242.
Silva-Santos KC, Santos GMG, Siloto LS, Morotti F, Marcantonio TN, Seneda MM, 2014a. Comparison of antral and preantral ovarian follicle populations between Bos indicus and Bos indicus-taurus cows with high or low antral follicles counts. Reprod Dom Anim. 49:48–51.
Siddiqui MAR, Ginther OJ. 2014. Switching of largest follicle from dominant to subordinate status when follicle and CL are in same ovary in heifers.
Theriogenology. 82: 259–265
Taylor C, Rajamahendran R. 1991. Follicular dynamics, corpus luteum growth and regression in lactating dairy cattle. Canadian Journal of Animal Science. 71(1): 61-68.
Valdez K E, Cuneo SP, Turzillo AM. 2005. Regulation of apoptosis in the atresia of dominant bovine follicles of the first follicular wave following ovulation. Reproduction 130:71–81.
Viana JHM, Ademir MF, Wanderlei FS, Almeida LS, Camargo. 2000. Follicular dynamics in zebu cattle. Pesq agropec bras. 35(12): 2501-2509
Williams MT, Marsh JM. 1978. Estradiol inhibition of luteinizing hormone-stimulated progesterone synthesis on isolated bovine luteal cells. Endo. 103:1611–8.
Wolfenson D, Inbar G, Roth Z, Kaim M, Bloch A,Braw R. 2004. Follicular dynamics and concentrations of steroids and gonadotropins in lactating cows and nulliparous heifers. Theriogenology. 62:1042–1055.
Vasenna R, Mapletoft RJ, Allodi S, Singh J, Adams GP. 2003. Morphology and developmental competence of bovine oocytes relative to follicular status.