commit to user
ANALISIS MOLEKULER REGIO PRE-S1, PRE-S2, DAN S ISOLAT
VIRUS HEPATITIS B 09IDSKAB-3
SKRIPSI
Untuk Memenuhi Persyaratan
Memperoleh Gelar Sarjana Kedokteran
ANGGA DWI PRASETYO
G.0009014
FAKULTAS KEDOKTERAN
UNIVERSITAS SEBELAS MARET
Surakarta
ABSTRAK
Angga Dwi Prasetyo, G0009014, 2012, Analisis Molekuler Regio S1, Pre-S2, dan S Isolat Virus Hepatitis B 09IDSKAB-3. Skripsi, Fakultas Kedokteran, Universitas Sebelas Maret, Surakarta.
Latar Belakang: Virus Hepatitis B (VHB) diketahui sebagai salah satu penyakit menular paling berbahaya di dunia. VHB merupakan jenis virus untai ganda sirkuler Deoxyribonucleic acid (DNA) yang bereplikasi dengan transkripsi balik. Mutasi genetik regio pre-S1, pre-S2, dan S VHB sering terjadi selama proses trankripsi balik tersebut. Studi ini bertujuan untuk menganalisis genotipe, subtipe, dan variasi genetik pada regio pre-S1, pre-S2, dan S isolat VHB 09IDSKAB-3 yang diperoleh dari komunitas Man Who Have Sex With Man di Surakarta.
Metode: Produk ekstraksi DNA VHB 09IDSKAB-3 digunakan sebagai cetakan untuk amplifikasi regio pre-S1, pre-S2, dan S. Sekuens yang diperoleh selanjutnya disejajarkan dengan metode Clustal W dengan reference sequences (genotipe A-H) yang dilaporkan di GenBank/DDBJ/EMNL. Genotipe VHB ditentukan dengan analisis phylogenetic. Subtipe VHB ditentukan berdasarkan beberapa asam amino yang berada pada sekuens HBsAg. Variasi genetik di regio pre-S1, pre-S2, dan S diidentifikasi dengan MEGA 4.0.
Hasil: Berdasarkan hasil BLAST dari GenBank, isolat VHB 09IDSKAB-3 diklasifikasikan ke dalam genotipe B3. Phylogenetic tree menunjukkan bahwa isolat VHB 09IDSKAB-3 diklasifikasikan ke dalam genotipe B3. Berdasarkan beberapa asam amino yang berada pada sekuens HBsAg, isolat VHB 09IDSKAB-3 diklasifikasikan ke dalam subtipe adw2. Variasi D27E ditemukan pada regio pre-S1 dan tidak ditemukan adanya variasi genetik pada regio pre-S2 maupun S.
Simpulan: Secara keseluruhan, isolat VHB 09IDSKAB-3 termasuk ke dalam genotipe B3 dan subtipe adw2. Di dalam studi ini hanya ditemukan variasi D27E. Variasi tersebut diindikasikan dapat merubah proses replikasi VHB, oleh karena itu diperlukan penelitian lebih lanjut untuk mengetahui pengaruh variasi tersebut.
commit to user
v ABSTRACTAngga Dwi Prasetyo, G0009014, 2012, Molecular Analysis of Pre-S1, Pre-S2, and S Region of 09IDSKAB-3 Hepatitis B Virus Isolate. Mini Thesis, Faculty of Medicine, Sebelas Maret University, Surakarta.
Background: Hepatitis B Virus (HBV) is known as one of the most dangerous infectious disease in the world. HBV is a partially double stranded DNA which replicates through reverse transcription. Genetic mutation frequently found in pre-S1, pre-S2, and S region during such the reverse transcription process. The aim of this study is to analyze genotype, subtype, and genetic variation in pre-S1, pre-S2, and S region of 09IDSKAB-3 HBV isolate from Man Who Have Sex With Man community in Surakarta.
Methods: 09IDSKAB-3 HBV DNA extraction was used as a template for amplication of pre-S1, pre-S2, and S region. The sequence results was then aligned by Clustal W with all of reference sequences (genotype A-H) reported in GenBank/DDBJ/EMNL. HBV genotype was identified by phylogenetic analysis. HBV subtype was deduced on the basis of the predicted amino acid sequences of HBsAg. Genetic variations in pre-S1, pre-S2, and S region were identified using MEGA 4.0.
Results: Based on BLAST search in GenBank, 09IDSKAB-3 HBV isolate was classified into genotype B3. Phylogenetic tree showed that 09IDSKAB-3 HBV isolate was classified into genotype B3. Based on the basis of predicted amino acid sequences of HBsAg, 09IDSKAB-3 HBV isolate was classified into subtype adw2. D27E variation was found in pre-S1 region, and there were not genetic variations in pre-S2 and S region.
Conclusion: Overall, 09IDSKAB-3 HBV isolate was classified into genoytpe B3 and subtype adw2. In this study, only D27E variation was found. The amino acid variation may have relevant changes the HBV replication efficiency, therefore, the amino acid variation found in the present report need further study.
DAFTAR ISI
E. Rancangan Penelitian... 13
F. Instrumen Penelitian ... 13
1. Alat Penelitian ... 13
2. Bahan Penelitian ... 14
G. Cara Kerja... 14
1. Ekstraksi DNA VHB ... 14
commit to user
viii3. Sekuensing dan Analisis data... 17
BAB IV HAS IL PENELITIAN
A. Isolasi DNA VHB ... 18
B. Amplifikasi PCR ... 18
C. Sekuensing Regio Pre-S1, Pre-S2, dan S Isolat VHB
09IDSKAB-3... 18
D. Proses Analisis Data Molekuler Regio Pre-S1, Pre-S2, dan S
Isolat VHB 09IDSKAB-3 ... 19
E. Hasil Analisis Data dari Isolat VHB 09IDSKAB-3 ... 24
BAB V PEMBAHASAN ... 25
BAB VI PENUTUP
A. SIMPULAN ... 33
B. SARAN... 33
DAFTAR PUSTAKA ... 35
BAB I
PENDAHULUAN
A. Latar Belakang Masalah
Virus Hepatitis B (VHB) adalah salah satu penyakit menular paling
berbahaya di dunia. Lebih dari dua miliar orang di dunia yang terinfeksi
VHB, 400 juta di antaranya berlanjut menjadi kronis, dan 600 ribu orang di
antaranya meninggal setiap tahun akibat VHB terkait sirosis dan atau
karsinoma hepar (WHO, 2009; Hepatitis B Foundation, 2009). Sedangkan
penderita hepatitis B di Indonesia, diperkirakan ada 13 juta orang yang
terinfeksi hepatitis B (Depkes RI, 2011). Jumlah tersebut termasuk dalam
jumlah terbanyak ketiga di dunia setelah Cina dan India. Oleh karena itu
Indonesia digolongkan termasuk ke dalam negara endemis VHB di dunia
dengan kategori Intermediate to High (Hwang dan Cheung, 2011).
Virus Hepatitis B (VHB) merupakan jenis virus untai ganda sirkuler
Deoxyribonucleic acid (DNA) dengan gap untai tunggalnya antara 600-2100 nukleotida (Rodes et al., 2007). Saat VHB bereplikasi melalui transkripsi
balik sangat memungkinkan untuk terjadinya mutasi genetik. Mutasi genetik
pada proses transkripsi balik tersebut dapat terjadi dengan perkiraan 1,4 s.d
3,2 x 10-5 basa/nukleotida/tahun (Ie et al., 2010). Selain akibat proses transkripsi balik tersebut, respon kekebalan pejamu dan terapi antiviral dapat
juga meningkatkan risiko terjadinya mutasi genetik (Sheldon dan Soriano,
commit to user
Salah satu mutasi genetik VHB yang paling sering ditemukan adalah
mutasi pada regio pre-S1, pre-S2, dan S. Mutasi yang terjadi akibat terjadinya
delesi pada regio pre-S1 dan pre-S2 merupakan salah satu faktor yang
meningkatkan risiko terjadinya karsinoma hepatoseluler bagi penderita VHB
(Liu et al., 2009). Sedangkan pada regio S, mutasi yang sering terjadi adalah
adanya subtitusi G145R pada determinan ‘a’ VHB, yang dikaitkan dengan
resistensi terhadap vaksin bagi penderita VHB (Sch iff et al., 2007). Selain itu
regio pre-S1, pre-S2, dan S juga sangat menentukan genotipe, subgenotipe,
maupun subtipe VHB sehingga apabila terjadi mutasi pada regio tersebut
sangat memungkinkan untuk ditemukannya genotipe, subgenotipe, maupun
subtipe VHB baru (Schaefer, 2007).
Sampai saat ini penelitian mengenai mutasi pada regio pre-S1, pre-S2,
dan S VHB terus berkembang karena analisis molekuler mengenai regio
pre-S1, pre-S2, dan S masih sangat diperlukan untuk memperoleh data strategi
replikasi, patogenesis, diagnosis, terapi, dan vaksin VHB. Sampai bulan Juni
2012, jumlah publikasi ilmiah mengenai regio pre-S1, pre-S2, dan S VHB
yang dipublikasikan di PubMed (http://www.ncbi.nlm.nih.gov/pubmed/)
sudah sebanyak 570 publikasi ilmiah. Sedangkan di Indonesia, sampai bulan
Juni 2012 baru ada 15 publikasi ilm iah di PubMed yang membahas regio
pre-S1, pre-S2, dan S VHB. Oleh karena itu peluang mengembangkan penelitian
mengenai regio pre-S1, pre-S2, dan S untuk pengembangan ilmu pengetahuan
3
penulis sangat tertarik untuk melakukan penelitian mengenai analisis
molekuler regio pre-S1, pre-S2, dan S VHB tersebut.
Sejak tahun 2009, grup riset Blood Borne Virus Universitas Sebelas Maret telah melakukan studi epidemiologi molekuler mengenai virus yang
menular melalui darah di antaranya yaitu VHB. Dari kesempatan yang ada
ini, penulis tertarik bergabung dalam penelitian tersebut dengan melakukan
analisis molekuler regio pre-S1, pre-S2, dan S isolat VHB 09IDSKAB-3 di
Laboratorium Biomedik FK UNS.
B. Perumusan Masalah
Bagaimanakah analisis molekuler regio pre-S1, pre-S2, dan S iso lat
Virus Hepatitis B 09IDSKAB-3?
C. Tujuan Penelitian
Penelitian ini bertujuan melakukan analisis molekuler regio S1,
pre-S2, dan S iso lat Virus Hepatitis B 09IDSKAB-3.
D. Manfaat Penelitian
1. Aspek teoritis
Penelitian ini dapat memberikan informasi ilmiah mengenai analisis
molekuler regio pre-S1, pre-S2, dan S isolat Virus Hepatitis B
commit to user
2. Aspek aplikatifAnalisis molekuler yang didapatkan melalui penelitian ini dapat
digunakan untuk penelitian lanjutan mengenai strategi pembuatan vaksin
VHB yang lebih adekuat dari pada vaksin yang sudah ada sekarang. Selain
itu penelitian ini juga dapat dijadikan bahan rujukan penelitian VHB
BAB II
LANDASAN TEORI
A. Tinjauan Pustaka
1. Virus Hepatitis B(VHB)
Virus Hepatitis B (VHB) termasuk dalam famili Hepadnaviridae, dan memiliki dua genera, yaitu genus Avihepadnavirus dan Orthohepadnavirus. Genus Avihepadnavirus ditemukan pada burung, sedangkan genus Orthohepadnavirus ditemukan pada hewan pengerat, woolly monkey, dan manusia (Schaefer, 2007). VHB kemudian digolongkan menjadi delapan genotipe (A-H) berdasarkan perbedaan
intergenotipe paling minimal 8% pada urutan nukleotida lengkap atau
lebih dari 4% pada gen S (Rodes et al., 2007).
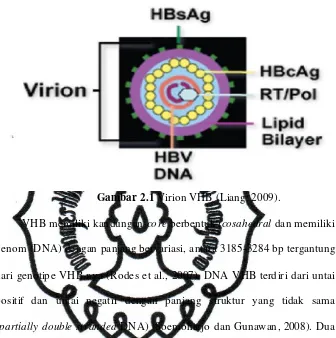
Virion VHB (partikel dane) terdiri dari mantel dan nukleokapsidnya.
Mantel VHB dahulu dikenal dengan nama Australia antigen yang sekarang
lebih dikenal dengan nama hepatitis B surface antigen (HBsAg) (Soemoharjo dan Gunawan, 2008). Sedangkan nukleokapsid VHB terdiri
dari subunit protein hepatitis B core antigen (HBcAg) dan hepatitis B ‘e’ antigen (HBeAg) dengan diameter 27 nm (Gambar 2.1) (Kuntz dan Kuntz,
commit to user
Gambar 2.1 Virion VHB (Liang, 2009).
VHB memiliki kandungan core berbentuk icosahedral dan memiliki genom (DNA)dengan panjang bervariasi, antara 3185-3284 bp tergantung
dari genotipe VHB-nya (Rodes et al., 2007). DNA VHB terd iri dari untai
positif dan untai negatif dengan panjang struktur yang tidak sama
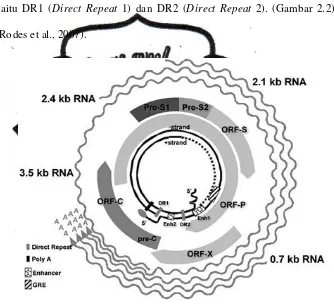
(partially double stranded DNA) (Soemoharjo dan Gunawan, 2008). Dua untai tersebut sebenarnya berbentuk lurus, tetapi karena terjad i overlaps 234 basa pada posisi 5’ terminal DNA VHB menjadikan DNA VHB berbentuk sirkuler (Gambar 2.2) (Rodes et al., 2007).
7
dikontrol oleh empat promotor (promotor pre-S1, pre-S2, core, dan x) dan
dua enhancer (Enh1 dan Enh2), yang berfungsi dalam pengaturan transkripsi RNA dan sinyal poliadenilasi yang dibutuhkan untuk replikasi
DNA. Ada dua regio penting lain yang juga berperan dalam replikasi VHB
yaitu DR1 (Direct Repeat 1) dan DR2 (Direct Repeat 2). (Gambar 2.2) (Rodes et al., 2007).
Gambar 2.2 Genom VHB (Liang, 2009).
DNA VHB merupakan virus yang melakukan replikasi dengan cara
commit to user
mRNA akan menghasilkan HBcAg, HBeAg, dan enzim polimerare,
sedangkan translasi mRNA LHBsAg, MHBsAg, dan SHBsAg akan
menghasilkan komponen protein HBsAg yaitu large (L) HBsAg, middle (M) HBsAg, dan small (S) HBsAg. Dalam proses selanjutnya pgRNA akan dimaturasi dengan proses transkripsi balik dengan dibantu enzim
polimerase. Proses ini dimulai dengan proses priming sintesis untai DNA (-) yang terjadi bersamaan dengan degradasi pgRNA, dan selanjutnya
berlangsung sintesis DNA (+) sehingga diperoleh DNA VHB secara utuh
(Soemoharjo dan Gunawan, 2008).
2. Hepatitis B Surface Antigen (HBsAg)
Hepatitis B surface Antigen (HBsAg) ada dalam tiga bentuk, yaitu selubung luar virion dan partikel HBsAg lepas yang berbentuk sferis
(bulat) dan tubuler (filamen) (Soemoharjo dan Gunawan, 2008). Dalam
perjalanan infeksi VHB, ada saat-saat dimana ketiga bentuk partikel
tersebut dapat ditemukan dalam darah secara bersamaan (Liang, 2009).
Semua partikel HBsAg selalu memiliki antigen ‘a’ yang sering
dinamakan determinan ‘a’. Di samping itu ada dua pasang subdeterminan
lain pada HBsAg, yaitu subdeterminan ‘d’ atau ‘y’, dan subdeterminan ‘w’
atau ‘r’. Atas dasar determinan ‘a’ dan subdeterminan tersebut didapatkan
subtipe HBsAg yaitu adw, adr, ayw, dan ayr (Rodes et al., 2007). Subdeterminan ini ditentukan oleh jenis asam amino yang menduduki
9
subtipe yaitu adw, adr, ayw, ayr ada tambahan subtipe baru, yaitu ayw 1-ayw4, adw2, adw4, adrq-, dan adrq+ (Soemoharjo dan Gunawan, 2008).
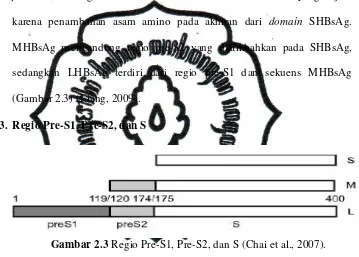
Ada tiga jenis protein dari HBsAg yang diterjemahkan oleh ORF S
yaitu large protein (LHBsAg), middle protein (MHBsAg), dan small protein (SHBsAg). Protein ini dibedakan berdasarkan ukuran yang terjadi karena penambahan asam amino pada akhiran dari domain SHBsAg. MHBsAg mengandung regio pre-S2 yang ditambahkan pada SHBsAg,
sedangkan LHBsAg terdiri dari regio pre-S1 dan sekuens MHBsAg
(Gambar 2.3) (Liang, 2009).
3. Regio Pre-S1, Pre-S2, dan S
Gambar 2.3 Regio Pre-S1, Pre-S2, dan S (Chai et al., 2007).
Regio pre-S dan S VHB merupakan regio yang memiliki jumlah
asam amino sekitar 400 aa (Gambar 2.3) (Hung et al., 2011). Regio pre-S
kemudian dapat dibagi menjadi pre-S1 dan pre-S2. Regio pre-S1 memiliki
panjang kira-kira 2848-3215 nukleotida dan regio pre-S2 memiliki panjang
kira-kira 1-154 nukleotida. Sedangkan untuk regio S memiliki panjang
sekitar 155-835 nukleotida (Soemoharjo dan Gunawan, 2008). Regio pre-S
merupakan regio yang sangat menentukan infektivitas VHB. Sifat dari
commit to user
Sedangkan, regio S bersifat hidrofobik pada bagian yang menghadap ke
dalam membran dan hidrofilik pada bagian yang menghadap luar, serta
sangat resisten tehadap protease (Shen et al., 2009).
Regio pre-S dan S merupakan regio yang sangat rentan terjadi
mutasi. Mutasi yang terjadi pada regio pre-S dan S terjadi di beberapa
tempat. Pada regio pre-S dapat terjadi akibat delesi pada 3’ terminal regio pre-S1, delesi pada start codon dari pre-S2, delesi pada 5’ terminal regio pre-S2, dan mutasi titik pada start codon regio pre-S2. Semua mutasi ini dikaitkan dengan risiko terjadinya karsinoma hepatoseluler (Liu et al.,
2009). Mutasi yang paling sering terjadi pada regio pre-S adalah pada
regio pre-S2, yaitu terjadinya delesi 2-55 nukleotida pada start codon regio pre-S2 (Fang et al., 2008; Hung et al., 2002).
Pada regio S, mutasi dapat terjadi akibat hilangnya determinan ‘a’
(escape mutant). Mutasi yang paling sering terjadi pada regio S terjadi akibat subtitusi Glisin menjadi Arginin pada kodon 145 (G145R) dan
Aspartat menjadi Alanin pada kodon 144 (D144A) di determinan ‘a’.
Mutasi ini ditemukan setelah pasien yang terinfeksi VHB diberikan vaksin
11
B. Kerangka Pemikiran
Kandidat Diagnostik
Isolat VHB 09IDSKAB-3
Sekuensing
Kloning Gen Penyandi Regio Pre-S1, Pre-S2, dan S
Analisis Regio Pre-S1, Pre-S2, dan S
Vaksin Kandidat
Diagnostik
Ekspresi Mutasi pada Regio S
Fokus Penelitian
Ekspresi Mutasi pada Regio Pre-S1 dan Pre-S2
commit to user
12 BAB IIIMETODE PENELITIAN
A. Jenis Penelitian
Penelitian ini bersifat eksploratif.
B. Lokasi Penelitian
Penelitian dilakukan di Laboratorium Biomedik Fakultas Kedokteran
Universitas Sebelas Maret.
C. Subjek Penelitian
Sampel darah yang diperoleh dari komunitas Man Who Have Sex With Man.
D. Objek Penelitian
13
E. Rancangan Penelitian
F. Instrumen Penelitian
1. Alat Penelitian:
a. centrifuge (Eppendorf, Hamburg, Jerman); b. thermocycler (Eppendorf, Hamburg, Jerman); c. micropipet (P1000, P200, P10) (Gilson, Wisconsin, USA); d. vortex (Thermo Fisher Scientific, Massachusetts, USA); e. microwave (Panasonic, Osaka, Jepang); f. deepfreezer (New Brunswick Scientific, New Jersey, USA ); g. tube rack; l. apparatus
Sampel Darah dari Komunitas Man Who Have Sex With Man
Uji Serologi HBsAg
+ -
PCR
Sekuensing
Analisis Regio Pre-S1, Pre-S2, dan S
Isolasi DNA
commit to user
USA); h. gel documentation (BioRad, California, USA); i. autoclave (Hirayama, Saitama, Jepang); j. refrigerator (Sharp, Osaka, Jepang); k. class II safety cabinet (ESCO, Oregon, USA); l. digital scale (Mettler Toledo, Greifensee, Switzerland); m. magnetic stirrer (Cimarex, Colorado, USA); n. Glove; o.Masker; p. Tissue.
DNA VHB diekstraksi dengan menggunakan PureLink™ Viral RNA/DNA Mini Kit, protokol dalam menggunakannya sebagai berikut: a. Lysate disiapkan.
b. Binding DNA:
15
2) Lysate (~640 µl) dengan lysis/binding buffer dan etanol dimasukkan ke dalam PureLinkTM Spin Column.
3) Column dipusingkan dengan 10.000 x g selama satu menit.
4) Collection tube dibuang dan spin column ditempatkan dalam collection tube baru.
c. Pembilasan DNA
1) Column dibilas dengan 500 µl wash buffer 1 yang disiapkan dengan etanol.
2) Column dipusingkan dengan 1000 x g selama satu menit pada suhu ruangan. Kemudian collection tube dibuang dan ditempatkan pada collection tube baru.
3) Column dicuci dengan menggunakan 500 µl wash buffer 2 yang disiapkan dengan etanol.
4) Column dipusingkan pada kecepatan maksimum selama tiga menit pada suhu ruangan. Kemudian collection tube dibuang.
d. Eluting DNA:
1) Spin column ditempatkan ke dalam 1,5 ml microsentrifuge tube steril.
2) DNA dielusikan menggunakan 25-200 µl PureLinkTM Genomic elution buffer.
commit to user
2. Amplifikasi dan Sekuensi PCRProses amplifikasi DNA VHB regio pre-S1, pre-S2, dan S
menggunakan kit GoTaq® Green Master Mix dan primer 12 dan KL-33 (Okamoto et al., 1990), dengan protokol sebagai berikut:
a. GoTaq® Green Master Mix dicairkan dalam temperatur ruangan, kemudian dipusingkan sebentar dengan microsentrifuge.
b. Komponen berikut ditambahkan ke dalam PCR tube on ice:
Komponen Jumlah
GoTaq® Green Master Mix 12,5 ml
upstream primer 10 µM (KL 12) 1 ml
downstream primer 10 µM (KL 33) 1 ml
DNA template 5 µl
Nuclease-Free Water 25 ml
c. Kemudian dilakukan proses preheated pada suhu 94 oC selama 5 menit, selanjutnya dilakukan 40 kali siklus PCR yang terdiri dari:
Denaturation 94°C 1 menit
Annealing 55 °C 1 menit
Extension 72 °C 2 menit
d. Setelah siklus selesai suhu diatur agar dalam kondisi 4 oC, kemudian
produk PCR dapat disimpan pada suhu –20 oC hingga digunakan.
17
H. Sekuensing dan Analisis Data
Sekuens nukleotida dari hasil amplifikasi ditentukan menggunakan
BigDye deoxy Terminator v1.1 cycle sequencing kit (Applied Biosystems) dan ABI Prism 310 genetic analyzer (Perkin Elmer). Data sekuens yang diperoleh kemudian dibandingkan dengan Bank data VHB di GenBank/DDBJ/EMBL dan dianalisis dengan menggunakan aplikasi MEGA 4.0 (Tamura et al.,
commit to user
18 BAB IVHASIL PENELITIAN
A. Isolasi DNA VHB
Sampel darah yang diperoleh dari komunitas Man Who Have Sex With Man yang positif dengan uji HBsAg selanjutnya dilakukan ekstraksi DNA dengan menggunakan PureLink TM Viral RNA/DNA Mini Kit sehingga diperoleh hasil isolat DNA VHB 09IDSKAB-3.
B. Amplifikasi PCR
Produk isolasi DNA VHB yang telah diperoleh selanjutnya digunakan
sebagai cetakan untuk amplifikasi regio pre-S1, pre-S2, dan S dengan
menggunakan GoTaq® Green Master Mix. Dalam amplifikasi dengan PCR tersebut, pasangan primer yang digunakan adalah KL-12 dan KL-33
(Okamoto et al., 1990). Hasil amplifikasi menunjukkan isolat VHB
09IDSKAB-3 positif dengan primer KL-12 dan KL-33.
C. Sekuensing Regio Pre-S1, Pre-S2, dan S VHB
Isolat VHB 09IDSKAB-3 selanjutnya dilakukan penentuan sekuens
nukleotida dengan menggunakan BigDye deoxy Terminator v1.1 cycle
19
digunakan untuk mencari isolat dengan kemiripan sekuens tertinggi pada
suatu daerah menggunakan aplikasi Basic Local Alignment Search Tool (BLAST) (Lampiran 3). Dari hasil BLAST tersebut didapatkan data isolat
yang memiliki skor BLAST terbesar dengan isolat VHB 09IDSKAB-3 yaitu
isolat VHB AB554070 (Lampiran 4).
D. Proses Analisis Data Molekuler Regio Pre-S1, Pre-S2, dan S Isolat VHB
09IDSKAB-3
Dalam usaha mencari informasi dan data pada tingkat molekuler dari
isolat VHB 09IDSKAB-3 dengan isolat di dunia, data yang diperoleh dari
hasil BLAST tersebut selanjutnya dijadikan pembanding kemiripan dan
kedekatan isolat VHB 09IDSKAB-3 dengan isolat VHB yang ada di dunia.
Pada proses selanjutnya penulis mencari reference sequences yang digunakan sebagai data pembanding untuk melihat phylogenetic tree dari isolat VHB 09IDSKAB-3. Data dari hasil tersebut diperoleh kedekatan
commit to user
Tabel 4.1 Daftar Reference Sequences yang Digunakan dalam Analisis Regio Pre-S1, Pre-S2, dan S Isolat VHB 09IDSKAB-3.
21
Seluruh sekuens tersebut sebelumnya disajikan menggunakan program
CLC Sequence Viewer, yang selanjutnya dilakukan phylogenetic analysis menggunakan aplikasi MEGA 4.0 (Tamura et al., 2007) (Lampiran 5). Data
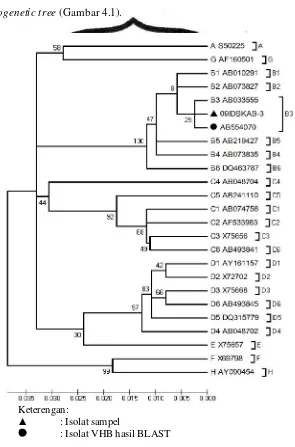
dari hasil phylogenetic analysis tersebut kemudian disajikkan dalam bentuk phylogenetic tree (Gambar 4.1).
Keterengan:
: Isolat sampel
: Isolat VHB hasil BLAST
commit to user
Analisis penentuan subtipe dari isolat VHB 09IDSKAB-3 dilakukan
dengan menggunakan aplikasi MEGA 4.0. Data yang diambil dalam analisis
ini diperoleh dari multiple alignment asam amino dari sekuens HBsAg VHB isolat VHB 09IDSKAB-3 pada posisi asam amino 116-183. Seluruh
reference sequences maupun data hasil BLAST dicocokkan, kemudian dilihat pada deteminan ‘a’ dan asam amino pada posisi 122, 127, 134, 159, 160, dan
177 untuk menentukan subtipe dari isolat VHB 09IDSKAB-3 (Gambar 4.2).
Gambar 4.2 Penyejajaran Asam Amino Posisi 116-183 Regio S (determinan ‘a’) dari Isolat VHB 09IDSKAB-3 dengan Reference Sequences Subtipe VHB.
Selain menganalisis genotipe, subgenotipe, dan subtipe dari isolat VHB
09IDSKAB-3. Analisis regio pre-S1, pre-S2, dan S VHB pada penelitian ini
juga dilakukan pada tingkat asam amino dan nukleotida dari isolat VHB
23
pre-S2, dan S dari isolat tersebut. Hasil dari multiple alignment pada regio pre-S1, pre-S2 dan S dapat dilihat pada gambar 4.3-4.5.
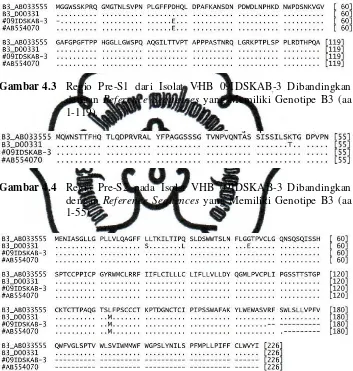
Gambar 4.3 Regio Pre-S1 dari Isolat VHB 09IDSKAB-3 Dibandingkan dengan Reference Sequences yang Memiliki Genotipe B3 (aa 1-119).
Gambar 4.4 Regio Pre-S2 pada Isolat VHB 09IDSKAB-3 Dibandingkan dengan Reference Sequences yang Memiliki Genotipe B3 (aa 1-55).
commit to user
E. Hasil Analisis Data dari Isolat VHB 09IDSKAB-3 (Accession Number:
JQ965941)
Hasil phylogenetic analysis dari isolat VHB 09IDSKAB-3 menunjukkan bahwa isolat tersebut termasuk ke dalam genotipe B3 (Gambar
4.1). Data dari hasil BLAST pada isolat ini menunjukkan bahwa isolat ini
memiliki skor BLAST tertinggi dengan isolat VHB AB554070 (Lampiran 3).
Isolat VHB AB554070 merupakan isolat yang berasal dari Papua (Indonesia)
dengan genotipe yang sama dengan isolat VHB 09IDSKAB-3.
Data pada gambar 4.2 yang merupakan determinan ‘a’ dari VHB
memperlihatkan bahwa isolat VHB 09IDSKAB-3 dan AB554070 merupakan
isolat VHB yang tergolong dalam subtipe adw2. Pada tingkat regio, data pada regio pre-S1 isolat VHB 09IDSKAB-3 maupun AB554070 tampak adanya
perbedaan asam amino dengan reference sequences yang memiliki genotipe B3 yaitu variasi D27E (Aspartic Acid menjadi Glutamic Acid) (Gambar 4.3). Sedangkan pada regio pre-S2 maupun regio S tidak ditemukan adanya variasi
genetik pada sekuens asam amino isolat VHB 09IDSKAB-3 (Gambar 4.4 dan
BAB V
PEMBAHASAN
A. Analisis Genotipe dan Subtipe Isolat VHB 09IDSKAB-3
Sugauchi et al. (2002) mengklasifikasikan VHB/B ke dalam dua grup
yaitu Bj (“j” singkatan dari Jepang) karena paling banyak ditemukan di
Jepang dan Ba (“a” singkatan dari Asia), karena ditemukan di seluruh Asia.
Subgrup dari Ba kemudian dibagi lagi menjadi empat subgenotipe yaitu
VHB/B2-VHB/B5. Dari keempat jenis subgenotipe VHB tersebut, jenis
VHB/B3 merupakan jenis VHB yang paling banyak ditemukan di Indonesia.
Sedangkan untuk beberapa jenis genotipe dan subgenotipe lainnya yang ada
di Indonesia, dapat ditemukan genotipe VHB/A di wilayah Balikpapan dan
Kupang, VHB/D berada di wilayah Papua (VHB/D6) dan Maluku (VHB/D1
dan VHB/D3) (Mulyanto et al., 2009; Utama et al., 2009).
Berdasarkan phylogenetic tree dari isolat VHB 09IDSKAB-3 menunjukkan bahwa isolat tersebut termasuk ke dalam genotipe B3 (Gambar
4.1). Hasil ini dapat dikatakan sesuai dengan teori karena dari studi
sebelumnya menyatakan bahwa di Indonesia genotipe yang dominan adalah
B3 (Utama et al., 2009). Studi ini juga menunjukkan bahwa isolat VHB
09IDSKAB-3 termasuk dalam subtipe adw2 (Gambar 4.2). Hasil ini
menunjukkan kesesuaian dengan studi yang ada sebelumnya bahwa subtipe
commit to user
di wilayah Sumatra, Jawa, Kalimantan Selatan, Bali, Lombok, Ternate, dan
Morotai (Lusida et al., 2003; Mulyanto et al., 1997).
B. Regio pre-S1 Isolat VHB 09IDSKAB-3
Regio pre-S1 dari LHBsAg diketahui sebagai regio yang sangat penting
bagi siklus hidup dan infektifitas VHB (Salisse dan Sureau, 2009). Di dalam
regio pre-S1 terdapat beberapa sub-elemen penentu infektifitas VHB, yaitu
pada asam amino 2-75. Pada area tersebut terdapat myristoyl yang berikatan dengan Gly2, receptor binding site (aa 2-48), dan daerah aa 49-75 (Duff et al., 2009). Salah satu sub-elemen tersebut, yaitu (hepatosite) receptor binding site, memiliki sebuah struktur yang cukup berperan dalam siklus hidup VHB, struktur tersebut adalah N-terminal dari residu 75 asam amino pada regio pre-S1. N-terminal regio pre-S1 akan mengirimkan sebuah signal berupa myristylation signal, signal ini merupakan signal yang akan membuat N-terminal dari regio pre-S1 dapat menempel pada permukaan sel sehingga protein VHB dapat disekresikan (Chai et al., 2007). Pada proses selanjutnya
ketika ikatan dengan membran hepatosit sudah terjadi maka receptor binding site (aa 2-48) akan melengkapi ikatan dan sisa asam amino yang lainnya (aa 49-75) akan menstabilkan ikatan yang terjadi (Duff et al., 2009).
Mutasi genetik pada aa 2-75 dilaporkan dapat menyebabkan perubahan
pada proses replikasi dan infektifitas VHB. Beberapa laporan mutasi genetik
27
G2A menjadi mutasi yang menyebabkan perubahan secara signifikan pada
proses pengiriman myristylation signal sehingga terjadi gangguan pada proses replikasi dan infektifitas VHB (Yeung et al., 2011). Di dalam sekuens dari
isolat VHB 09IDSKAB-3, tidak menunjukkan adanya substitusi G2A
maupun delesi pada receptor binding site dan aa 49-75. Data dari isolat VHB 09IDSKAB-3 hanya ditemukan adanya variasi D27E (Gambar 4.3). Namun,
studi yang menyatakan variasi yang terjadi pada isolat VHB 09IDSKAB-3
memang belum ada sehingga pengaruhnya terhadap infektifitas VHB juga
belum diketahui secara pasti.
Selain sub-elemen aa 2-75 pada regio pre-S1, di dalam regio pre-S1
juga terdapat beberapa area lain yang memiliki fungsi tertentu, di antaranya
adalah promotor S (aa 67-111) dan kotak CCAAT (nt 3137-3141) (Yeung et
al., 2011). Promotor S berperan cukup penting dalam menjaga keseimbangan
dari sintesis tiga protein HBsAg (large, middle, dan small). Di dalam studi sebelumnya yang dilakukan oleh Yeung et al. (2011), delesi pada promotor S
dilaporkan dapat merubah keseimbangn sintesis protein dari HBsAg dimana
akan berakibat pada peningkatan stres pada Retikulum Endoplasma (RE).
Stres tersebut kemudian dikaitkan dengan berkembangnya penyakit
karsinoma hepatoseluler. Di dalam studi ini, isolat VHB 09IDSKAB-3
diketahui bahwa promotor S terkonservasi dengan baik sehingga tidak
ditemukan indikasi adanya stres dari RE yang diakibatkan mutasi promotor S.
Mutasi yang terjadi pada kotak CCAAT dilaporkan dapat meningkatkan
commit to user
Namun, fungsi dari CCAAT dalam perjalanan penyakit VHB masih
dipertanyakan (Yeung et al., 2011). Isolat VHB 09IDSKAB-3 tidak
ditemukan adanya mutasi pada kotak CCAAT sehingga dapat dikatakan
peningkatan stres pada RE yang diakibatkan o leh mutasi pada kotak CCAAT
tidak ditemukan.
Pada studi sebelumnya yang dilakukan dengan matched nested case-control sudy menyebutkan terdapat keterkaitan antara delesi yang terjadi pada regio pre-S1 dengan meningkatnya risiko terjadinya karsinoma
hepatoseluler (Fang et al., 2008). Yeung et al. (2011) juga menyebutkan
bahwa adanya delesi pada regio pre-S1 dapat meningkatkan risiko
berkembangnya karsinoma hepatoseluler pada penyakit hepatitis B kronis.
Delesi yang terjadi pada regio pre-S1 tersebut adalah pada start codon dan daerah 3’ terminal dari regio pre-S1. Kejadian delesi yang paling berpengaruh terhadap berkembangnya karsinoma hepatoseluler adalah delesi pada daerah
3’ terminal regio pre-S1. Di dalam sekuens asam amino isolat VHB 09IDSKAB-3 tidak ditemukan adanya delesi start codon dan 3’ terminal regio pre-S1 sehingga indikasi adanya peningkakan risiko berkembangnya
karsinoma hepatoseluler tidak ditemukan.
C. Regio pre-S2 Isolat VHB 09IDSKAB-3
Regio pre-S2 merupakan regio hidrofilik yang memiliki beberapa
29
asparagine112 (N112) diketahui berkaitan penting dengan mekanisme N-glycosylation yang berperan dalam proses replikasi dan maturasi VHB (Rodes et al., 2007). N-glycosilation diketahui berperan penting pada berbagai protein, perannya antara lain sebagai protein folding, kontrol kualitas protein, sekresi protein, dan modulasi dari respon imun. Secara lebih
khusus, N-glycosylation memiliki fungsi berdasarkan ukuran protein HBsAg-nya. Pada LHBsAg dan SHBsAg, N-glycosilation berperan dalam replikasi VHB, sedangkan pada MHBsAg diketahui bahwa N-glycosilation berperan dalam maturasi VHB (Lambert dan Prange, 2007). Mutasi genetik N112
dilaporkan dapat menyebabkan terhambatnya replikasi dan maturasi VHB.
Pada isolat VHB 09IDSKAB-3, N112 diketahui terkonservasi dengan baik
sehingga indikasi terhambatnya maturasi VHB tidak ditemukan.
Area human serum albumin receptor belum diketahui secara jelas fungsinya, namun dipercaya berperan penting bagi infektifitas VHB. Mutasi
genetik yang terjadi pada area ini juga belum diketahui secara jelas
pengaruhnya terhadap perjalanan penyakit (Yeung et al., 2011).
Mutasi pada regio pre-S2 dilaporkan dapat menyebabkan produksi
protein di dalam RE secara berlebihan sehingga dapat meningkatkan stres
dari RE. Stres RE ini dapat berakibat pada kerusakan DNA dan
ketidakstabilan genom yang akhirnya memicu karsinoma hepar (Hsieh et al.,
2004). Hsieh et al. (2007) melaporkan bahwa mutasi pada regio pre-S2 dapat
commit to user
menyebabkan siklus sel tidak terkontrol sehingga menimbulkan mekanisme
onkogenik.
Hasil studi meta analysis dan a match nested case-control study menunjukkan delesi pada daerah 5’ terminal regio pre-S2 menjadi mutasi paling banyak ditemukan pada orang yang terkena karsinoma hepar (Fang et
al., 2008; Liu et al., 2009). Di dalam studi lain juga dijelaskan selain delesi
pada daerah 5’ terminal regio pre-S2, mutasi juga dapat ditemukan berupa delesi dan mutasi titik pada start codon regio pre-S2 (Yeung et al., 2011). Di dalam isolat VHB 09IDSKAB-3, tidak ditemukan adanya mutasi tersebut,
maka dapat dikatakan peningkatan risiko untuk perkembangnya karsinoma
hepatoseluler pada isolat VHB 09IDSKAB-3 tidak ditemukan jika dilihat dari
faktor regio pre-S2.
D. Regio S Isolat VHB 09IDSKAB-3
Di dalam regio S terdapat suatu regio hidrofilik yang dikenal dengan
Major Hidrophilic Region (MHR) (aa 101-172). MHR merupakan area yang memiliki penentu antigenitas VHB yang disebut determinan ‘a’. Di dalam
MHR terdapat polipeptida yang akan mendefinisikan determinan ‘a’ pada
reseptor. Polipeptida ini memiliki struktur berbentuk setengah lingkaran dan
disebut dengan Antigen Loop (AGL). Peran AGL antara lain menginisiasi perlekatan ke sel dan pelekat spesifik pada reseptor. Letak AGL pada
C-31
terminal yang bersifat hidrofobik (aa 173-226) (Duff et al., 2009; Salisse dan Sureau, 2009).
Secara setruktur dan fungsi, di dalam AGL terdapat asam amino yang
sangat berpengaruh terhadap infektifitas VHB yaitu C121, C124, C137,
C139, C147, C149, dan N146. Keenam asam amino cysteine (C) sangat terkonservasi dengan baik dan memiliki peranan penting karena memiliki
ikatan disulfida yang sangat diperlukan dalam menjaga struktur dari
determinan ‘a’, sedangkan asparagine146 (N146) memiliki peranan dalam perlekatan virus ke sel karena berperan penting dalam mekanisme N-glycosylation (Rodes et al., 2007; Yong-lin et al., 2012).
Perubahan genetik pada AGL diketahui sangat mempengaruhi
infektifitas VHB. Mutasi genetik terutama yang terjadi pada asam amino
yang berperan penting bagi infektifitas VHB dapat merubah kemampuan
VHB dalam menginfeksi sel. Duff et al. (2009) melaporkan substitusi
C139S, C147S, dan C149S dapat menurunkan infektifitas VHB terutama
substitusi pada C147S (Duff et al., 2009). Namun demikian, ketiga macam
substitusi tersebut tidak ditemukan pada isolat VHB 09IDSKAB-3 sehingga
tidak di-temukan indikasi terjadinya perubahan infektifitas VHB dalam isolat
VHB 09IDSKAB-3.
Mutasi yang terjadi pada N146 dilaporkan sangat mempengaruhi proses
perlekatan virus pada permukaan sel, karena asam amino ini sangat berkaitan
commit to user
genetik N146 dilaporkan dapat menyebabkan proses pembentukan protein
(replikasi) menjadi terganggu (Lambert dan Prange, 2007; Purdy MA, 2007).
Data dari isolat VHB 09IDSKAB-3 menunjukkan N146 terkonservasi dengan
baik dan tidak mengalam i mutasi sehingga proses pembentukan protein VHB
dapat dikatakan berlangsung dengan baik.
Mutasi genetik yang pertama kali dan paling banyak ditemukan pada
VHB adalah substitusi G145R. Substitusi ini dikaitkan dengan resistensi
terhadap vaksin karena substitusi ini sering terjadi pada pasien yang telah
menggunakan vaksin VHB (Schiff et al., 2007). Substitusi tersebut dikaitkan
dengan perubahan yang dapat mengakibatkan terganggunya proses
pengikatan antibodi tetapi tidak mengubah struktur dari AGL (Salisse dan
Sureau, 2009). Namun demikian, di dalam studi ini substitusi G145R tidak
BAB VI
PENUTUP
A. Simpulan
1. Isolat VHB 09IDSKAB-3 termasuk ke dalam genotipe B3 dan subtipe
adw2.
2. Hasil penyejajaran isolat VHB 09IDSKAB-3 dengan reference sequences genotipe B3 menunjukkan data bahwa:
a. Pada regio pre-S1 ditemukan variasi D27E yang masih belum diketahui
implikasi perubahan tersebut terhadap infektifitas VHB.
b. Pada regio pre-S2 dan S tidak ditemukan adanya variasi genetik baik di
tingkat asam amino maupun nukleotida.
B. Saran
1. Untuk melihat genotipe dan subtipe dari isolat VHB 09IDSKAB-3 dengan
lebih teliti diperlukan analisis molekuler pada seluruh sekuens isolat VHB
09IDSKAB-3, tidak hanya pada regio pre-S1, pre-S2, dan S saja.
2. Adanya variasi asam amino pada regio pre-S1, pre-S2, dan S isolat VHB
09IDSKAB-3 perlu diteliti lebih lanjut terutama hubungannya dengan
virulensi, patogenesis, replikasi, dan keberhasilan terapi.
3. Perlu dilakukan analisis molekuler lebih lanjut dengan menggunakan data
aligmentasi yang didapat dari penelitian in i, terutama untuk mencari
commit to user
patogenesis maupun terapi yang belum dikonfirmasi keberadaannya dalam