TINJAUAN PUSTAKA
Struktur Anatomi Otot Rangka
Otot rangka (skeletal muscle) bertanggung jawab atas pergerakan tubuh secara sadar. Otot rangka disebut juga otot lurik (striated muscle) karena pengaturan filamennya yang tumpang tindih, sehingga memberikan sel-sel itu penampakan berlurik atau bergaris di bawah mikroskop. Sebuah otot terdiri atas berkas serat otot (sel-sel) bernukleus majemuk, yang masing-masing merupakan berkas miofibril (Campbell et al. 2004).
Masing-masing miofibril terdiri atas beberapa tipe protein, yaitu; miosin, aktin, troponin, tropomiosin, titin, dan nebulin. Miosin merupakan protein motor dari miofibril. Pada otot lurik, sekitar 250 molekul miosin bersatu membentuk sebuah filamen tebal. Sementara aktin merupakan molekul protein yang membentuk filamen tipis. Filamen tebal dan tipis tersebut diatur dalam suatu unit kontraktil yang disebut sarkomer. Satuan fungsional yang disebut sarkomer ini memiliki beberapa elemen di dalamnya, yaitu suatu pita A yang gelap dan pita I yang terang yang tersusun selang-seling beraturan. Pusat pita A disebut zona H, kurang padat dibandingkan bagian lain dari pita. Sebuah pita A dibagi dua oleh garis M, sedangkan pita I dibagi dua oleh garis Z yang sangat sempit. Filamen tipis selain mengandung aktin juga mengandung tropomiosin dan kompleks troponin, keduanya merupakan protein yang mengatur kejadian kontraksi (the regulatory proteins). Kemudian titin yang membentang dari garis Z sampai ke daerah yang berdekatan dengan garis M, serta nebulin yang berada di sepanjang tepi filamen tipis dan menempel pada garis Z (Silverthorn 2009).
Miofibril dikelilingi oleh struktur yang dibentuk membrana yang tampak dalam foto mikograf elektron sebagai vesikel dan tubulus. Struktur ini membentuk susunan sarkotubulus, yang dibentuk dari sistem T dan suatu retikulum sarkoplasma. Sistem T tubulus transversa yang bersambung dengan membrana serabut otot membentuk suatu kisi yang diperforasi oleh miofibril tersendiri. Ruang diantara dua lapisan sistem T merupakan suatu perluasan ruang ekstra sel. Retikulum sarkoplasma yang membentuk suatu tirai tidak teratur di sekeliling tiap fibril mempunyai sisterna terminalis yang membesar dalam kontak erat dengan
sistem T pada sambungan antara pita A dan I. Pada titik kontak ini, susunan sistem T sentral dengan suatu sisterna retikulum sarkoplasma pada sisi manapun telah membawa ke penggunaan istilah trias untuk menggambarkan sistem ini. Sistem T berfungsi sebagai hantaran cepat potensial aksi dari membrana sel ke semua fibril dalam otot (Ganong 1995). Secara keseluruhan struktur anatomi otot rangka dapat dilihat pada Gambar 1 berikut;
Gambar 1 Anatomi otot rangka (Silverthorn 2009)
mitokondria retikulum sarkoplasma nukleus sistem T sarkolema filamen tebal filamen tipis miofibril sarkomer pita A Garis Z Garis Z pita I Garis M Zona H miofibril Garis Z Garis Z Garis M Garis M filamen tebal filamen tipis
Dasar Molekuler Mekanisme Kontraksi Otot Rangka
Ketika sebuah otot berkontraksi, panjang masing-masing sarkomer menjadi berkurang, yaitu; jarak antara garis Z ke garis Z berikutnya menjadi lebih pendek. Pada sarkomer yang berkontraksi, pita A tidak berubah panjangnya. Akan tetapi pita I memendek dan zona H menjadi hilang. Peristiwa ini dapat dijelaskan dengan model filamen luncur (sliding-filament model) pada kontraksi otot. Menurut model ini, bukan filamen tipis dan bukan juga filamen tebal yang berubah panjangnya ketika otot berkontraksi, melainkan filamen tersebut meluncur di atas satu sama lain secara longitudinal. Sehingga derajat tumpang-tindih filamen tipis dan tebal meningkat. Jika daerah tumpang-tumpang-tindih itu meningkat, baik bagian yang hanya ditempati oleh filamen tipis (pita I) maupun bagian yang hanya ditempati oleh filamen tebal (zona H) harus berkurang panjangnya (Campbell et al. 2004).
Peluncuran filamen tersebut didasarkan pada interaksi molekul aktin dan miosin yang menyusun filamen tipis dan filamen tebal. Kepala miosin merupakan pusat reaksi bioenergetik yang memberi energi untuk kontraksi otot. Kepala miosin dapat mengikat ATP dan menghidrolisisnya menjadi ADP serta fosfat anorganik. Energi yang dibebaskan dari pemecahan ATP dipindahkan ke miosin, sehingga menjadi konfigurasi penambah energi tinggi. Miosin yang sudah bertenaga ini berikatan dengan tempat spesifik pada aktin, membentuk suatu titian silang (cross-bridge) (Campbell et al. 2004).
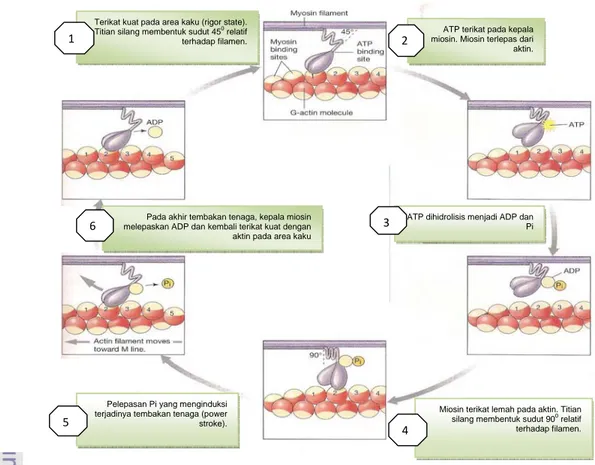
Hal tersebut secara lebih terperinci ditunjukkan oleh Silverthorne (2009) serta dapat dilihat juga pada Gambar 2, dalam enam tahapan berikut ini;
1. Area kaku (the rigor state)
Kepala miosin berikatan kuat dengan tempat spesifik pada aktin (Globular protein) membentuk suatu titian silang (cross-bridge). Pada area ini tidak ada nukleotida, baik ATP maupun ADP yang menempati sisi perlekatan dari kepala miosin. Pada otot aktif, the rigor state ini hanya terjadi untuk periode/waktu yang sangat singkat sekali.
2. Pengikatan ATP dan pelepasan miosin
Sebuah molekul ATP terikat pada kepala miosin. Hal ini merubah afinitas perlekatan aktin terhadap miosin, dan miosin pun memisahkan diri dari aktin.
3. Hidrolisis ATP
Kepala miosin yang mengikat ATP kemudian menghidrolisis ATP menjadi ADP serta fosfat anorganik (Pi).
4. Miosin terikat lemah pada aktin
Energi yang dibebaskan dari pemecahan ATP menyebabkan kepala miosin terikat lemah pada aktin, terutama molekul aktin yang baru. Pada titik ini, miosin memiliki energi yang potensial, sehingga menjadi konfigurasi energi tinggi.
5. Pelepasan fosfat anorganik
Pelepasan fosfat anorganik dari sisi perlekatan miosin menginduksi terjadinya sebuah tembakan tenaga (the power stroke). Kepala miosin kemudian berotasi lalu mendorong filamen aktin bergerak menuju garis M dan melewatinya.
6. Pelepasan ADP
Miosin melepaskan ADP. Pada titik ini, kepala miosin kembali terikat kuat dengan aktin pada area kaku (the rigor state). Siklus berulang ketika sebuah molekul ATP baru terikat pada kepala miosin.
Gambar 2 Siklus dasar molekuler kontraksi otot rangka (Silverthorn 2009)
Terikat kuat pada area kaku (rigor state). Titian silang membentuk sudut 450
relatif terhadap filamen.
ATP terikat pada kepala miosin. Miosin terlepas dari aktin.
ATP dihidrolisis menjadi ADP dan Pi
Miosin terikat lemah pada aktin. Titian silang membentuk sudut 900
relatif terhadap filamen. Pelepasan Pi yang menginduksi
terjadinya tembakan tenaga (power stroke).
1 2
5
4
Pada akhir tembakan tenaga, kepala miosin melepaskan ADP dan kembali terikat kuat dengan aktin pada area kaku
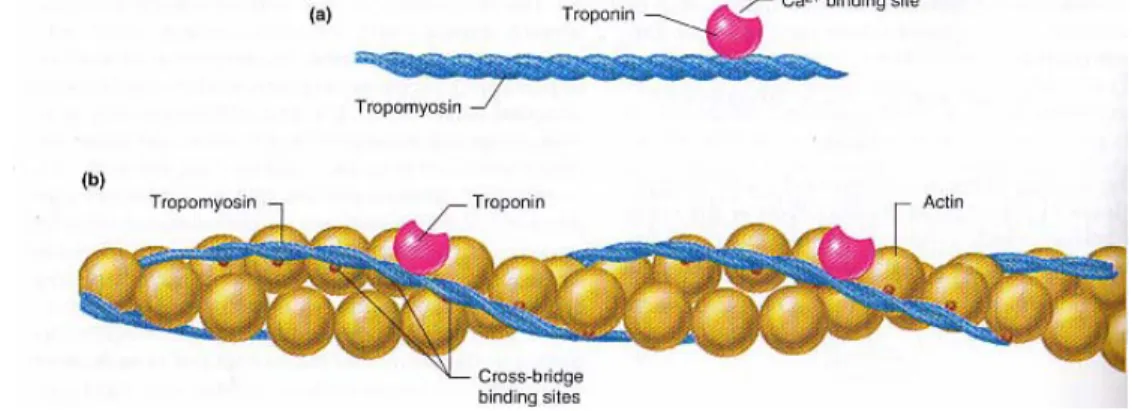
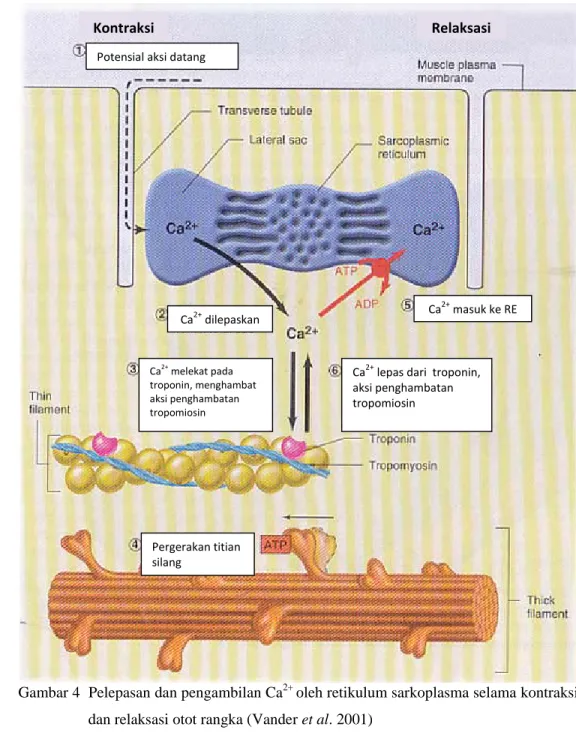
Agar siklus kontraksi dapat terus berlangsung, efek penghambatan oleh kompleks troponin-tropomiosin pada bagian aktif dari filamen aktin normal otot yang sedang relaksasi harus dihambat, karena menyebabkan area ini tidak dapat melekat pada kepala filamen miosin untuk menimbulkan kontraksi. Kondisi ini memunculkan peran ion Ca2+ . Dengan adanya ion Ca2+ dalam jumlah besar, efek penghambatan oleh kompleks troponin-tropomiosin terhadap filamen itu sendiri dapat dihambat (Guyton dan Hall 1997). Perlekatan ion Ca2+ pada sisi perlekatan spesifik troponin membuat bentuk troponin berubah, dan berakibat juga pada terlepasnya tropomiosin dari sisi perlekatan miosin pada tiap molekul aktin. Sehingga kondisi sebaliknya, yakni ketiadaan ion Ca2+, akan menyebabkan aktivitas kontraktil terhenti (Vander et al. 2001).
Ion-ion Ca2+ tersebut disimpan di dalam retikulum sarkoplasma. Membrana retikulum sarkoplasma mengandung protein transport aktif primer, yaitu Ca-ATPase, yang akan memompa ion Ca2+ dari sitosol kembali ke dalam lumen retikulum. Pemompaan ion Ca2+ kembali ke dalam lumen retikulum membutuhkan waktu yang lebih lama daripada pelepasannya. Oleh karena itu, konsentrasi dari sisa-sisa ion Ca2+ dalam sitosol ditingkatkan, dan kontraksi akan berlanjut sampai beberapa waktu setelah mendapat sebuah potensial aksi (Vander et al. 2001).
Gambar 3 Kedudukan molekul troponin terhadap molekul tropomiosin dan sisi perlekatan mekanisme titian silang (Vander et al. 2001)
Gambar 4 Pelepasan dan pengambilan Ca2+ oleh retikulum sarkoplasma selama kontraksi dan relaksasi otot rangka (Vander et al. 2001)
Sel otot secara khas hanya menyimpan cukup ATP untuk beberapa kontraksi saja. Sel-sel otot juga menyimpan glikogen diantara miofibril, akan tetapi sebagian besar energi yang diperlukan untuk kontraksi otot yang berulang disimpan dalam bahan yang disebut fosfagen. Keratin fosfat, yang merupakan fosfagen vertebrata, menyediakan gugus fosfat ke ADP untuk membentuk ATP (Campbell et al. 2004). Kontraksi Relaksasi Potensial aksi datang Ca2+dilepaskan Ca2+ melekat pada troponin, menghambat aksi penghambatan tropomiosin Pergerakan titian silang Ca2+ lepas dari troponin, aksi penghambatan tropomiosin Ca2+masuk ke RE
Metabolisme Energi pada Aktivitas Otot Rangka
Mader (2001) menyebutkan bahwa kontraksi otot membutuhkan pasokan ATP yang berlimpah. Ada tiga jalur yang dapat memasok ATP tambahan selama kontraksi otot;
1. Sel-sel otot memiliki mitokondria yang dapat membentuk ATP melalui mekanisme respirasi selular aerobik.
2. Sel-sel otot mengandung kreatin fosfat, yang digunakan sebagai penyimpan pasokan fosfat berenergi tinggi.
Kreatin fosfat tidak berpartisipasi secara langsung dalam kontraksi otot. Sebaliknya, ia digunakan untuk membentuk ATP melalui reaksi berikut;
Kreatin fosfat + ADP ATP + Kreatin
3. Ketika pasokan kreatin fosfat habis, sel otot masih mampu memproduksi ATP secara anaerob.
Respirasi anaerob terjadi ketika sel tidak cepat mendapatkan pasokan oksigen yang cukup untuk melakukan respirasi aerobik. Hal ini dapat terjadi misalnya selama melakukan olahraga yang berat. Respirasi anaerob hanya dapat memasok ATP untuk waktu yang sangat singkat, karena mekanisme ini menghasilkan asam laktat yang dapat menyebabkan kesakitan otot dan kelelahan.
Mekanisme Pembentukan ATP melalui Respirasi Aerobik
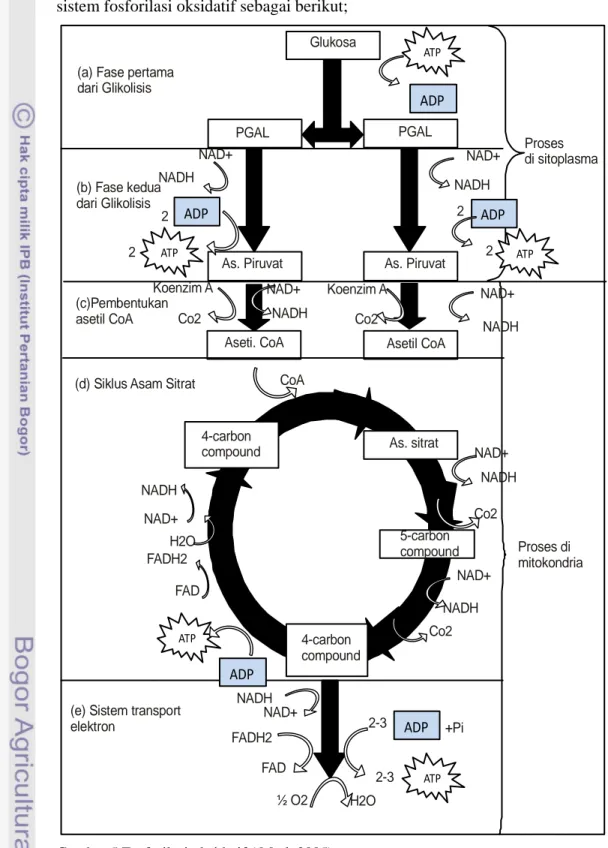
Mekanisme respirasi aerobik merupakan sintesis ATP menggunakan sistem transport elektron, dan disebut juga sebagai proses fosforilasi oksidatif, karena membutuhkan oksigen untuk bertindak sebagai penerima akhir dari elektron dan H+. Proses ini hanya akan terjadi apabila tersedia cukup oksigen. Pergerakan dari elektron melintasi sistem transport elektron ini dideskripsikan dengan sebuah model yang disebut teori kemiosmotik. Berdasarkan teori ini, sepasang elektron berenergi tinggi berjalan dari kompleks ke kompleks sepanjang jalur sistem transport, kemudian sebagian energi dilepaskan dan digunakan untuk memompa H+ dari matrik mitokondria keluar menuju ruang antar membran. Pergerakan H+ menuju ruang antar membran menyebabkan peningkatan konsentrasi H+. Selama H+ kembali melintasi membran (menurunkan tigkat konsentrasinya) masuk ke
dalam matrik mitokondria, beberapa energi kinetik yang potensial ditangkap sebagai ATP (Silverthorne 2009). Mark (2005) memberikan gambaran lengkap sistem fosforilasi oksidatif sebagai berikut;
Gambar 5 Fosforilasi oksidatif ( Mark 2005) ATP ADP ATP ATP ATP ATP ADP ADP ADP ADP Glukosa PGAL PGAL 2 2 2 2 NAD+ NADH NAD+ NADH Koenzim A Co2 Koenzim A Co2 NAD+ NADH NAD+ NADH CoA
As. Piruvat As. Piruvat
Aseti. CoA Asetil CoA
4-carbon
compound As. sitrat
5-carbon compound 4-carbon compound NAD+ NADH Co2 NAD+ NADH Co2 FAD FADH2 H2O NAD+ NADH 2-3 2-3 +Pi NADH NAD+ FADH2 FAD ½ O2 H2O Proses di sitoplasma Proses di mitokondria (a) Fase pertama
dari Glikolisis
(b) Fase kedua dari Glikolisis
(c)Pembentukan asetil CoA
(d) Siklus Asam Sitrat
(e) Sistem transport elektron
Agar proses fosforilasi oksidatif terus berjalan, otot-otot yang sedang aktif bergantung pada pasokan oksigen dan nutrien yang adekuat melalui sistem sirkulasi untuk mempertahankan aktivitasnya. Pada metabolisme ini, Pengubahan ADP menjadi ATP terjadi berkaitan dengan molekul protein yang disebut ATPase. Konsentrasi ion H+ yang tinggi di bagian luar bilik dan perbedaan potensial listrik yang besar melalui membran bagian dalam menyebabkan ion H+ mengalir ke dalam matriks mitokondria melalui zat dari molekul ATPase. Sewaktu melakukan hal tersebut, energi yang dihasilkan dari aliran ion H+ ini digunakan oleh ATPase untuk mengubah ADP menjadi ATP. Untuk tiap dua elektron yang berjalan melalui rantai transpor elektron (mewakili ionisasi dari 2 atom hidrogen), dapat disintesis sampai 3 molekul ATP (Guyton dan Hall 1997).
Mekanisme Pembentukan ATP melalui Pemecahan Kreatin Fosfat
Kreatin (Cr) merupakan jenis asam amino yang tersimpan di dalam otot sebagai sumber energi. Di dalam otot, bentuk kreatin yang sudah terfosforilasi menjadi kreatin fosfat (PCr) akan memiliki peranan penting dalam proses metabolisme energi secara anaerobik di dalam otot untuk menghasilkan energi. Saat istirahat, sejumlah ATP di dalam mitokondria memindahkan fosfatnya ke kreatin, sehingga terbentuk simpanan kreatin fosfat. Enzim kreatin kinase membantu pemecahan kreatin fosfat menjadi fosfat anorganik dan kreatin, proses ini juga disertai dengan pelepasan energi sebesar 43 kJ (10,3 kkal) untuk tiap 1 mol PCr. Fosfat anorganik yang dihasilkan dari proses pemecahan PCr ini melalui proses fosforilasi dapat mengikat molekul ADP untuk kembali membentuk molekul ATP, sehingga memungkinkan kontraksi berlanjut (Irawan 2007; Ganong 1995).
Kreatin fosfat (PCr) membawa ikatan fosfat berenergi tinggi yang serupa dengan ATP. Ikatan fosfat berenergi tinggi dari kreatin fosfat membawa memiliki jumlah energi bebas yang sedikit lebih tinggi daripada yang dimiliki oleh ikatan ATP. Jumlahnya tiga sampai delapan kali lebih banyak. Disamping itu, ikatan energi tinggi kreatin fosfat mengandung kira-kira 8.500 kalori tiap mol pada keadaan standar, dan sebanyak 13.000 kalori tiap mol pada keadaan di dalam tubuh (380C dan konsentrasi reaktan rendah). Hasil ini sedikit lebih besar
daripada 12.000 kalori tiap mol dalam setiap dua ikatan fosfat berenergi tinggi dari ATP. Kombinasi energi dari ATP cadangan dan kreatin fosfat di dalam otot masih dapat menimbulkan kontraksi otot maksimal hanya untuk 5 sampai 8 detik (Guyton dan Hall 1997).
Mekanisme Pembentukan ATP melalui Respirasi Anaerobik
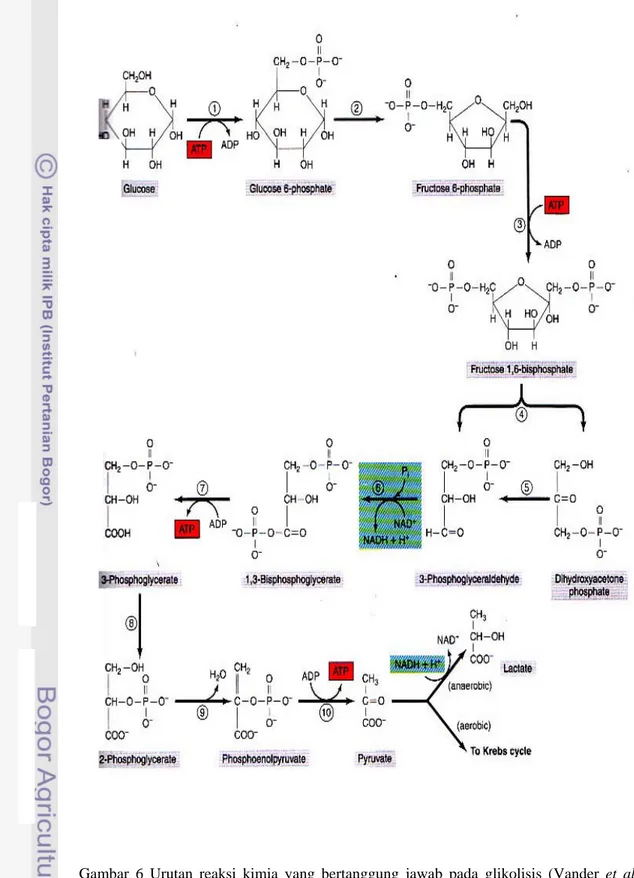
Mekanisme pembentukan ATP melalui respirasi anaerobik terjadi dalam jalur glikolisis. Glikolisis berarti memecahkan molekul glukosa untuk membentuk dua molekul asam piruvat. Proses metabolisme energi ini menggunakan simpanan glukosa yang sebagian besar diperoleh dari glikogen otot atau dari glukosa yang terdapat di dalam aliran darah untuk menghasilkan ATP. Walaupun terdapat banyak reaksi kimia dalam rangkaian glikolitik, hanya sebagian kecil energi bebas dalam molekul glukosa yang dibebaskan dari setiap langkah. Akan tetapi diantara tahap 1,3-asam difosfogliserat dan 3-asam fosfogliserat, dan sekali lagi diantara tahap asam fosfoenol piruvat dan asam piruvat, jumlah energi yang dibebaskan lebih dari 12.000 kalori per mol, jumlah yang dibutuhkan untuk membentuk ATP. Sehingga terdapat 4 molekul ATP yang dibentuk dari masing-masing molekul fruktosa 1,6 fosfat yang dipecah menjadi asam piruvat. Namun 2 molekul ATP masih dibutuhkan untuk fosforilasi glukosa asal untuk membentuk fruktosa 1,6 fosfat sebelum glokolisis dapat dimulai. Oleh karena itu, hasil akhir dari seluruh proses glikolitik hanya 2 molekul ATP untuk masing-masing molekul glukosa yang dipakai (Guyton dan Hall 1997).
Sistem glikolitik dapat membentuk molekul ATP kira-kira 2,5 kali lebih cepat daripada mekanisme fosforilasi oksidatif di mitokondria. Oleh karena itu, bila sejumlah besar ATP dibutuhkan untuk kontraksi otot dalam waktu singkat sampai sedang, mekanisme glikolisis anaerob ini dapat digunakan sebagai sumber energi cepat. Sistem ini kira-kira setengah cepatnya dari sistem fosfagen (Kreatin fosfat). Di bawah kondisi optimal, sistem glikolitik dapat menyediakan aktivitas otot yang maksimal selama 1,3 sampai 1,6 menit sebagai tambahan dari yang disediakan oleh sistem fosfagen (Farhan 2009).
Berikut 10 langkah reaksi kimia yang berurutan dalam proses glikolisis, seperti yang ditunjukkan oleh Vander et al. (2001);
Gambar 6 Urutan reaksi kimia yang bertanggung jawab pada glikolisis (Vander et al.
Kelelahan Otot Rangka
Kelelahan adalah suatu mekanisme perlindungan tubuh agar tubuh terhindar dari kerusakan lebih lanjut sehingga terjadi pemulihan setelah istirahat. Istilah kelelahan biasanya menunjukkan kondisi yang berbeda-beda dari setiap individu, tetapi semuanya bermuara kepada kehilangan efisiensi dan penurunan kapasitas kerja serta ketahanan tubuh. Kelelahan otot adalah tremor/kejang pada otot atau perasaan nyeri pada otot. Kelelahan otot dapat dipengaruhi oleh sistem saraf pusat, cadangan glikogen otot, dan keadaan ion fosfat maupun kalium dalam otot. Kelelahan itu dikarenakan sumber energi yang dimiliki oleh tubuh menurun atau habis, asam laktat meningkat, keseimbangan cairan dan elektrolit terganggu. Akibatnya menimbulkan rasa lemah, lesu, dan penurunan konsentrasi (Anonim 2004).
Sementara Sherwood (2002) mendefinisikan kelelahan sebagai kondisi penurunan kapasitas dan atau berkurangnya kemampuan kerja, dan menurut Foss dan Keteyian (1998), kelelahan adalah ketidakmampuan tubuh membentuk tenaga dan atau kecepatan, yang merupakan akibat dari aktivitas otot. Istilah kelelahan menjadi sangat bervariasi karena banyaknya penyebab dengan penentu utama yang berbeda bergantung kepada intensitas dan durasi aktivitas, serta kondisi lingkungan (Tirtayasa 2001).
Berbagai jenis faktor yang dapat menyebabkan kelelahan kemudian dapat diklasifikasikan ke dalam dua mekanisme besar, yaitu; mekanisme central fatigue di sistem saraf pusat, dan mekanisme peripheral fatigue dimanapun diantara neuromuscular juntion dan elemen-elemen kontraktil dari otot itu sendiri (Silverthorne 2009). Pada mekanisme central fatigue, kelelahan sentral terjadi jika sistem saraf pusat tidak dapat mengaktifkan neuron motorik yang mempersarafi otot yang bekerja secara adekuat. Individu memperlambat atau menghentikan olahraganya walaupun otot-ototnya masih mampu bekerja. Pada aktivitas olahraga ringan-sedang, kelelahan sentral mungkin menyebabkan penurunan kerja fisik berkaitan dengan kejenuhan (Wilmore dan Costil 1994).