TINJAUAN PUSTAKA
Tinjauan Umum Sapi Perah FH di Indonesia
Sapi perah merupakan hasil domestikasi dari Bos taurus primigenius sekitar 2000 tahun yang lalu (Anderson & Kiser 1966; Mason 1984; Gillespie 1992). Sapi perah FH berkembang dari sapi perah di Eropa. Sapi perah termasuk dalam famili Bovidae. Salah satu ciri dari famili ini mempunyai sepasang tanduk yang tidak bercabang (Mason 1984). Sapi perah yang berkembang di Eropa adalah jenis Bos taurus (Gillespie 1992). Ciri khas dari Bos taurus yaitu tidak mempunyai punuk.
Sapi perah masuk ke Indonesia sejak jaman penjajahan Belanda (Sudono 1999). Pemerintah Belanda mendatangkan sapi perah FH untuk memenuhi kebutuhan susu bagi karyawan Belanda (Ibrahim et al. 1992). Pada tahun 1891– 1893 telah dilakukan pemuliaan bibit sapi perah di Indonesia dengan mengimpor pejantan FH dari Belanda (Sudono 1999). Setelah Indonesia merdeka, pertumbuhan populasi sapi perah terus meningkat. Hal ini ditandai dengan adanya pembangunan usaha-usaha sapi perah oleh swasta (Yusdja 2005). Kebijakan importasi sapi perah dalam jumlah besar dan teratur, cukup memberikan kontribusi dalam perkembangan sapi perah (Anggraeni & Iskandar 2008). Menurut Ibrahim et al. (1992), selama tahun 1979-1990 pemerintah Indonesia mengimpor sapi perah FH dara sekitar 100.000 ekor. Sapi-sapi tersebut terutama diimpor dari Australia dan Selandia Baru serta dalam jumlah kecil dari Amerika Serikat (Diwyanto et al. 2006). Jumlah populasi sapi perah di Indonesia sampai dengan tahun 2007 mencapai 377.772 ekor. Penyebaran sapi perah FH tersebut paling banyak di pulau Jawa. Populasi sapi perah FH di Jawa Timur diperkirakan sebanyak 138.988 ekor (36.79%), Jawa Tengah 115.377 ekor (30.54%) dan Jawa Barat 102.724 ekor (27.19%) (Ditjen Peternakan 2007).
Sapi perah FH mempunyai sifat jinak dan lebih mudah menyesuaikan diri dengan lingkungan (Gillespie 1992). Menurut Damron (2006) sapi perah FH betina menghasilkan susu dengan jumlah paling tinggi dibanding dengan bangsa sapi perah lainnya. Rata-rata produksi susu sapi perah FH di Amerika Serikat mencapai 21.671 pound (9.751.95 kg) dalam satu masa laktasi. Susu sapi perah
FH di Amerika Serikat mengandung kadar lemak relatif rendah yaitu 3.6%. Menurut Diwyanto et al. (2006), produksi susu sapi perah FH di Indonesia khususnya di Jawa Tengah dan Jawa Barat mencapai 3.500-4.750 kg dalam satu masa laktasi.
Perkawinan sapi perah FH di Indonesia, sebagian besar menggunakan sistem IB (Diwyanto et al. 2006). Semen beku yang digunakan dalam IB sebagian besar diproduksi oleh sapi perah FH pada Balai Besar Inseminasi Buatan (BBIB) dan Balai Inseminasi Buatan (BIB). Sebagian semen beku yang lain diimpor dari Amerika, Kanada, Perancis, Australia, Selandia Baru dan Jepang (Diwyanto et al. 2006). Penyebaran semen beku di wilayah sentra sapi perah sebagian besar dilakukan oleh BBIB Singosari dan BIB Lembang (Diwyanto et al. 2006). Diwyanto dan Herliantien (2006) melaporkan bahwa penggunaan IB telah mencapai lebih dari 90% populasi sapi perah betina.
Keunggulan penggunaan perkawinan IB diantaranya dapat mempercepat penyebaran sifat-sifat unggul dalam populasi (Foote 2002). Perkawinan IB lebih efektif dan efisien dibanding dengan perkawinan alami pada sapi perah (Diwyanto & Herliantien 2006). Sebagai ilustrasi, perkawinan alami satu jantan hanya mampu melayani 20-30 betina pertahun. Pada inseminasi buatan satu jantan dalam sekali ejakulasi dapat diproduksi sekitar 200 straw atau lebih. Dengan demikian jika sperma satu pejantan ditampung dua kali dalam satu minggu, maka dalam satu tahun dapat diambil 20.800 straw. Jumlah tersebut dapat dipakai untuk membuahi 10.000 sapi betina (Diwyanto & Herliantien 2006).
Kelainan Genetik pada Sapi Perah
Kelainan genetik adalah kelainan bersifat menurun yang menyebabkan kelainan fisik atau fungsi tubuh (Čitek et al. 2006). Salah satu dampak yang ditimbulkan oleh kelainan genetik yaitu kematian pada sapi dengan kelainan genetik (letal) (Maciejowski & Zieba 1982). Penyebaran kelainan genetik dapat terjadi melalui proses perkawinan (Basrur & King 2005). Kelainan ini sebagian besar disebabkan oleh peristiwa mutasi pada tingkat gen maupun kromosom (Maciejowski & Zieba 1982).
Kelainan genetik pada sapi sebagian besar terjadi pada individu bergenotipe resesif homozigot. Resesif homozigot dapat muncul dari perkawinan antar individu karier atau heterozigot (Healy 1996). Individu dengan genotipe heterozigot tampak seperti individu normal (Basrur & King 2005). Hal ini terjadi karena sifat resesif tertutupi oleh sifat dominan pasangannya (Maciejowski & Zieba 1982). Keadaan ini memberi peluang sifat resesif tetap berada dalam populasi (Čitek & Bláhová 2004).
Dua di antara beberapa kelainan genetik pada sapi perah yang telah diidentifikasi adalah: sitrulinemia (Robinson et al. 1993; Padeeri et al. 1999; Nassiry et al. 2005; Patel et al. 2006; Čitek et al. 2006 ) dan DUMPS (Shanks et al. 1987; Čitek et al. 2006; Patel et al. 2006; Rahimi et al. 2006; Vătăşescu et al. 2006). Kelainan genetik tersebut menyebabkan kelainan pada metabolisme tubuh. Kelainan metabolisme tubuh kadangkala mengakibatkan kematian pada sapi dengan kelainan genetik (Dennis et al. 1989).
Sitrulinemia
Sitrulinemia pertama kali ditemukan pada manusia tahun 1962 (Healy et al. 1996). Sekitar tahun 1989 kelainan ini ditemukan juga pada sapi perah FH (Harper et al. 1989). Menurut Dennis et al. (1989) sitrulinemia merupakan defisiensi enzim argininosuccinate synthase (ASS) yang bersifat menurun. Enzim ASS merupakan salah satu enzim yang berperan penting dalam siklus urea terutama dalam pembentukan urea. ASS berperan dalam mengubah sitrulin dan aspartat menjadi arginin suksinat. Perubahan tersebut dilakukan dengan cara menghidrolisis adenosine triphosphate (ATP) menjadi adenosine monophosphate (AMP) dan pyrophosphate (Pi) pada siklus urea yang terjadi di dalam hati (Jenkinson et al. 1996). Jika terjadi defisiensi ASS maka akan terjadi penumpukan sitrulin yang menyebabkan pembentukan urea di dalam hati terhenti.
Sitrulinemia pada sapi perah FH disebabkan perubahan basa pada kodon 86 berupa transisi sitosin (CGA/arginin) menjadi timin (TGA/stop kodon). Mutasi tersebut terjadi pada ekson 5 gen penyandi enzim arginin suksinat sintase kromosom 11 (Padeeri et al. 1999). Sapi dengan defisiensi ASS mempunyai jumlah asam amino lebih sedikit dibanding sapi normal. Jumlah asam amino sapi
defisiensi ASS sebanyak 85 sedangkan sapi normal sebanyak 412 (Dennis.et.al..1989).
Konsentrasi enzim ASS dalam leukosit pada sapi heterozigot sebesar 0.28 ± 0.10 miliunit/mg protein. Konsentrasi enzim ASS dalam leukosit pada sapi normal homozigot sebesar 0.40 ± 0.25 miliunit/mg protein. Konsentrasi enzim ASS pada sapi heterozigot lebih kecil dibanding pada sapi normal homozigot. Meskipun begitu, konsentrasi enzim ASS pada sapi heterozigot masih dalam batas normal.
Tanda-tanda klinis sapi resesif homozigot (penderita sitrulinemia) yaitu: mengalami ataxia (lemah otot), kebutaan, sitrulin dalam darah meningkat hingga mencapai 200 kali dibanding normal. Konsentrasi enzim arginin suksinat sintase menurun secara ekstrim di hati, dan biasanya mengakibatkan kematian setelah kelahiran (Dennis et al. 1989).
Healy (1996) melaporkan kejadian sitrulinemia di Australia dengan frekuensi karier sebesar 13% dari 98 sapi FH yang diuji. Robinson et al. (1993) melaporkan kejadian sitrulinemia di Amerika sebesar 0.27% dengan jumlah sapi yang diuji sebanyak 367 ekor. Kejadian sitrulinemia di Australia diketahui berasal dari salah satu pejantan yang diimpor dari Amerika. Pembawa kelainan sitrulinemia di Australia diperkirakan berasal dari keturunan sapi Holstein Amerika yaitu Linmack Kriss King (LMKK) (Healy 1996). Berdasarkan kejadian tersebut, beberapa negara pengimpor sapi perah FH melakukan deteksi kelainan sitrulinemia. Beberapa negara pengimpor sapi perah FH di antaranya adalah India (Patel et al. 2006); Iran (Nassiry et al. 2005) dan Chekoslovakia (Čitek et al. 2006). Hasil identifikasi dari India, Iran dan Chekoslovakia menyatakan tidak ada kejadian sitrulinemia pada sapi perah FH mereka.
Deficiency of Uridine Monophosphate Synthase
Deficiency of uridine monophosphate synthase pertama kali ditemukan pada sapi perah tahun 1985 (Smith et al. 1985). Pada tahun 1987 sapi perah FH di Amerika terdeteksi mengalami kelainan tersebut (Shanks et al. 1987). Deficiency of uridine monophosphate synthase merupakan kelainan genetik yang disebabkan oleh defisiensi enzim uridin 5-monophosphate synthase (UMPS). Enzim UMPS
adalah salah satu enzim yang berperan dalam biosintesis pirimidin (Robinson et al. 1983). Akibat dari kelainan ini dapat terjadi gangguan pada biosintesis pirimidin, yaitu ketidakmampuan uridin 5-monophosphate synthase (UMPS) untuk mengkatalisis asam urotik menjadi UMP (Shanks & Robinson 1989).
Kelainan genetik DUMPS pada sapi perah FH disebabkan perubahan basa pada kodon 405. Perubahan basa itu berupa transisi sitosin (CGA/arginin) menjadi timin (TGA/stop kodon). Mutasi tersebut terjadi pada ekson 5 gen penyandi enzim UMPS kromosom nomor 1 (Schwenger et al. 1994). Menurut Robinson et al. (1983) UMPS pada sapi dalam keadaan resesif homozigot mengandung asam amino yang lebih sedikit daripada UMPS sapi normal.
Ciri-ciri sapi heterozigot atau karier DUMPS, selama laktasi mengalami penumpukan asam orotat pada susu, urin dan darah (Shanks & Robinson 1990). Menurut Robinson et al. (1983) konsentrasi orotat di dalam susu bervariasi tergantung kondisi ternak seperti pada saat melahirkan, masa laktasi, dan jumlah laktasi. Rata-rata hewan ternak mempunyai orotat ± 15.8–81.1 μg/ml. Sapi karier mempunyai orotat lebih dari 300 μg/ml bahkan ada yang mencapai 1.000 μg/ml. Dampak kandungan orotat yang tinggi pada sapi karier belum diketahui. Ciri lain dari sapi heterozigot yaitu interval melahirkan lebih lama dari sapi normal setelah kelahiran anak pertama (Shank et al. 1987).
Defisiensi UMPS pada sapi Friesian-Holstein dalam keadaan resesif homozigot menyebabkan embrio mati pada saat implantasi di dalam uterus (Rahimi et al. 2006). Perkembangan zigot hanya sampai pembelahan blastula saja dan biasanya gugur tidak lama setelah implantasi (Schwenger et al. 1994). Sapi resesif homozigot tidak pernah dijumpai dalam keadaan dewasa.
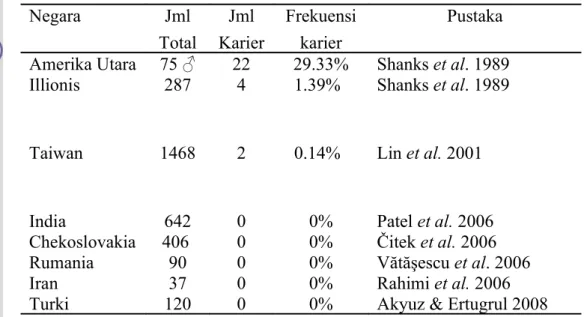
Kasus penyebaran DUMPS telah diidentifikasi di beberapa negara. Salah satu di antaranya di pusat inseminasi buatan sapi perah FH Amerika Serikat dengan frekuensi kejadian sebesar 29.33%, sedangkan pada peternakan rakyat sebesar 1.39% (Shanks et al.1989). Kelainan ini juga ditemukan di Taiwan dengan frekuensi kejadian sebesar 0.14% (Lin et al. 2001) (Tabel 1).
Penyebaran kelainan genetik DUMPS terjadi karena pemakaian pejantan pembawa kelainan genetik dalam IB (Robinson et al. 1983). Beberapa negara seperti India (Patel et al. 2006), Chekoslovakia (Čitek et al. 2006), Rumania (Vătăşescu et al. 2006), Iran (Rahimi et al. 2006) dan Turki (Akyuz & Ertugrul 2008) melakukan identifikasi terhadap kelainan genetik DUMPS. Hasil identifikasi tersebut ternyata tidak ditemukan karier DUMPS. Menurut Čitek dan Bláhová (2004), penyebaran kelainan genetik dapat dipercepat melalui
penggunaan inseminasi buatan. Dengan demikian, identifikasi kelainan genetik DUMPS diperlukan untuk mengantisipasi kerugian yang ditimbulkan pada sapi perah (Kamiński et al. 2005).
Tabel 1 Identifikasi kelainan DUMPS di beberapa negara
Negara Jml Total Jml Karier Frekuensi karier Pustaka Amerika Utara 75 ♂ 22 29.33% Shanks et al. 1989 Illionis 287 4 1.39% Shanks et al. 1989
Taiwan 1468 2 0.14% Lin et al. 2001
India 642 0 0% Patel et al. 2006
Chekoslovakia 406 0 0% Čitek et al. 2006
Rumania 90 0 0% Vătăşescu et al. 2006
Iran 37 0 0% Rahimi et al. 2006
Turki 120 0 0% Akyuz & Ertugrul 2008
Metode Polymerase Chain Reaction-Restriction Fragment Length Polymorphism (PCR-RFLP)
Polymerase chain reaction (PCR) merupakan metode enzimatis untuk memperbanyak fragmen DNA secara eksponensial dengan cara in vitro (Mullis & Faloona 1989). Dengan menggunakan metode PCR, dapat diperoleh fragmen DNA (110 bp, 5 X 10-19 mol) sebesar 200 000 kali setelah dilakukan 20 siklus reaksi selama 220 menit (Mullis & Faloona 1989).
Teknik PCR dipengaruhi oleh empat komponen utama yaitu: DNA cetakan, oligonukleotida primer, deoksiribonukleotida trifosfat (dNTP) dan enzim DNA polymerase (Sambrook & Russell 2001). Lebih lanjut Sambrook dan Russell (2001) serta Naqvi (2007) secara garis besar memaparkan prinsip kerja PCR yaitu denaturasi, penempelan primer (annealing) dan pemanjangan (elongasi). Proses dari denaturasi, penempelan dan pemanjangan disebut sebagai satu siklus. Proses ini biasanya berlangsung 25–40 siklus. Suhu denaturasi dan ekstensi bersifat permanen, masing-masing pada 95 °C dan 72 °C, sedangkan suhu penempelan bergantung pada panjang–pendeknya primer.
Metode PCR-RFLP merupakan teknik PCR yang menggunakan enzim restriksi (Viljoen et al. 2005). Enzim restriksi ini bersifat sangat spesifik dalam memotong urutan nukleotida yang dikenalinya (Naqvi 2007). Enzim tersebut mengenali urutan nukleotida tertentu dalam urutan nukleotida suatu gen.
Urutan nukleotida yang mampu dikenali oleh enzim restriksi disebut situs restriksi (Naqvi 2007). Panjang situs restriksi ini terdiri dari empat sampai enam nukleotida (Dowling et al. 1996). Jika situs restriksi mengalami mutasi (meskipun pada satu basa) maka enzim restriksi tidak mampu mengenalinya.
Menurut Vătăşescu et al. (2006) metode PCR-RFLP mampu mendeteksi individu karier sitrulinemia dan DUMPS dengan baik. Metode tersebut mempunyai keuntungan yaitu dapat dilakukan dengan mudah dan cepat. Enzim restriksi yang digunakan untuk deteksi kelainan sitrulinemia (enzim Ava II) (Patel et al. 2006) dan DUMPS (enzim Ava I) (Rahimi et al. 2006) tidak dapat mengenali alel mutan. Situs restriksi pada alel mutan kedua kelainan genetik tersebut tidak dikenali oleh enzim restriksi.
Hasil kerja enzim restriksi dapat dilihat pada perbedaan jumlah potongan pita DNA yang diperoleh setelah proses visualisasi pada gel poliakrilamid (Palumbi 1996). Selain gel poliakrilamid, proses visualisasi dapat pula menggunakan gel agarosa. Pita DNA yang diperoleh dari teknik tersebut selanjutnya dianalisis untuk mengetahui frekuensi alel normal dan alel mutan.
Panjang fragmen DNA produk PCR gen ASS adalah 176 pb. Produk PCR gen ASS setelah dipotong dengan Ava II pada individu normal akan dihasilkan dua pita DNA yang berukuran 98 pb dan 78 pb. Pada individu karier akan dihasilkan tiga pita DNA masing-masing berukuran 176 pb, 98 pb, dan 78 pb. Secara teori individu resesif homozigot akan menghasilkan satu pita berukuran 176 pb (Robinson et al. 1993). Ava II mengenali situs ggacc pada alel normal dan memotong antara g dengan g. Pada alel mutan tidak ada situs ggacc sehingga Ava II tidak mengenali situs tersebut.
Panjang fragmen DNA produk PCR gen UMPS adalah 108 pb. Produk PCR gen UMPS setelah dipotong dengan Ava I pada individu normal akan dihasilkan tiga pita DNA yang berukuran 53 pb, 36 pb, dan 19 pb. Pada individu karier akan dihasilkan empat pita DNA masing-masing berukuran 89 pb, 53 pb, 36 pb, dan 19 pb. Secara teori individu resesif homozigot akan menghasilkan dua pita berukuran
89 pb dan 19.pb (Schwenger et al. 1993). Ava I mengenali situs c(c/t)cg(a/g)g pada alel normal dan memotong antara c dengan c/t. Pada alel mutan tidak ada situs c(c/t)cg(a/g)g sehingga Ava I tidak mengenali situs tersebut dan tidak terjadi pemotongan.
Sampel Pool DNA
Sampel Pool DNA adalah metode pengelompokan DNA hasil ekstraksi dari sampel yang digunakan dalam kegiatan riset ke dalam satu tempat (pool) (Sham et al. 2002). Analisis sampel pool DNA dilakukan untuk mengetahui keberadaan alel normal maupun alel mutan. Jika dalam sampel pool DNA ditemui alel mutan, maka semua anggota pool tersebut diperiksa satu persatu.
Metode pool DNA merupakan cara yang praktis dalam proses deteksi terhadap alel penyebab berbagai kelainan genetik. Metode ini secara efisien dapat mengurangi pembiayaan untuk keperluan pereaksi PCR. Dengan demikian proses penentuan genotipe dapat dilakukan dengan murah dalam skala besar, dibanding bila tiap sampel diuji satu persatu (Sham et al. 2002). Selain itu, metode sampel pool DNA dapat mengefisiensikan peralatan (Yang et al. 2003) dan waktu (Churchill et al. 1993) yang dibutuhkan dalam proses deteksi.
Berbagai uji terhadap metode sampel pool DNA yang telah dilakukan (Churchill et al. 1993; Zhou et al. 2001; Sham et al. 2002; Yang et al. 2003; Meaburn et al. 2006; Szyda et al. 2008) menunjukkan bahwa metode ini dapat menjadi metode penentuan genotipe alternatif yang cukup akurat, efisien dan praktis. Menurut Szyda et al. (2008) satu sampel pool DNA dapat digunakan antara 20-40 sampel DNA. Yang et al. (2003) merekomendasikan isi satu sampel pool DNA sampai 30 sampel. Dengan demikian, waktu deteksi lebih singkat, hasil deteksi lebih cepat diketahui dan akurat.