TIDAK. 21, Picea sitchensis (Bong.) Makalah Konsensus Biologi Carr. Sitka Spruce) (2002) Dokumen Konsensus Tanaman Brassica (Brassica spp). No.64, Dokumen Konsensus Biologi Salmon Atlantik (Salmo salar) (2017) No.57, Dokumen Konsensus Biologi Singkong (Manihot esculenta Crantz) (2014).
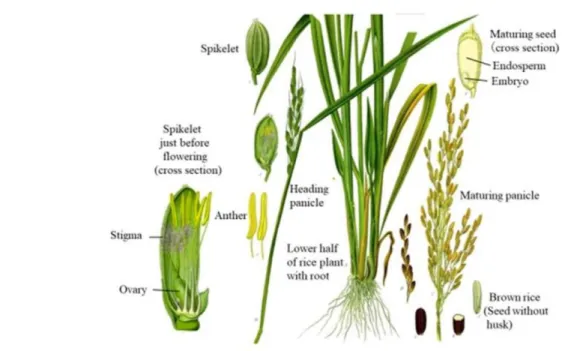
MORFOLOGI
BAGIAN 1. URAIAN UMUM
TERMASUK TAKSONOMI DAN
Klasifikasi dan nomenklatur
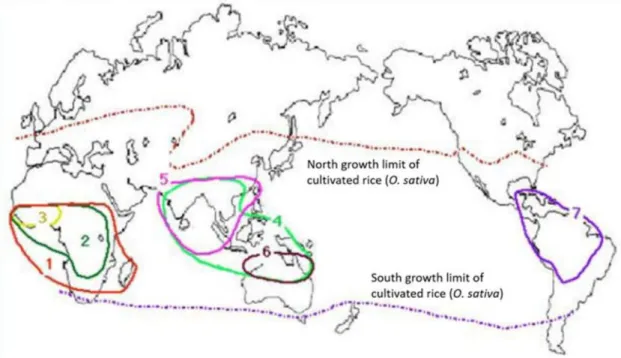
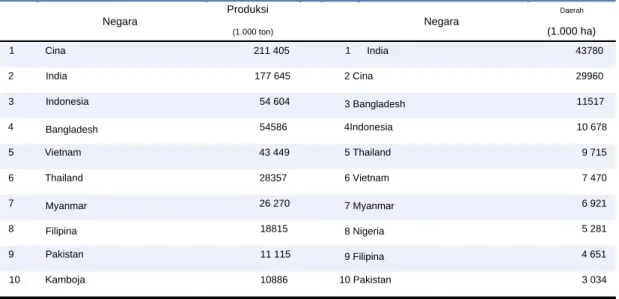
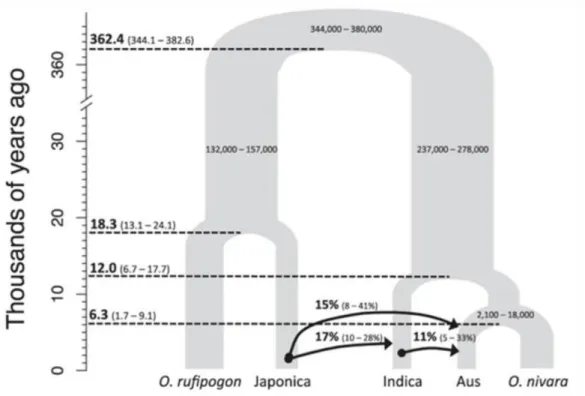
Padi yang ditanam di Asia diduga berasal dari lembah hilir Sungai Yangtze, Republik Rakyat Tiongkok, berdasarkan jejak geografis, arkeologi dan cerita rakyat (Fuller et al., 2009). Siklus hidup dan ciri-ciri morfologi yang dijelaskan di atas merupakan ciri khas rata-rata tanaman padi yang ditanam di seluruh dunia, namun sangat berfluktuasi, bahkan untuk kultivar yang sama, bergantung pada kondisi tanah dan cuaca.
BAGIAN 2. PUSAT ASAL, DISTRIBUSI GEOGRAFIS
DAN PRAKTIK AGRONOMI
Ekosistem agronomi dan ekosistem lainnya yang dikelola secara intensif di mana spesies tersebut tumbuh atau tumbuh sendiri, termasuk praktik pengelolaannya
Ekosistem dan habitat tempat spesies tersebut hidup secara asli, dan tempat spesies tersebut dinaturalisasi
Padi gogo tadah hujan mencakup 32% dari total luas sawah di Afrika, namun hanya 6% di Asia. Beberapa metode penyemaian langsung yang berbeda, seperti penyemaian, pencelupan, dan pengeboran, digunakan di daerah dataran tinggi tadah hujan di Asia dan Afrika.
BAGIAN 3. BIOLOGI REPRODUKSI
- Waktu dan durasi generasi dalam keadaan alami, dan ketika ditanam atau dikelola
- Reproduksi (produksi bunga, biji, dan bibit vegetatif)
Hilangnya viabilitas serbuk sari diperkirakan disebabkan oleh kekeringan (Nakayama, 1934; Koga et al., 1971; Khatun dan Flowers, 1995). Kedalaman tanam yaitu kedalaman tanah tempat benih mulai berkecambah mempengaruhi perkecambahan benih padi (Ohno et al., 2018).
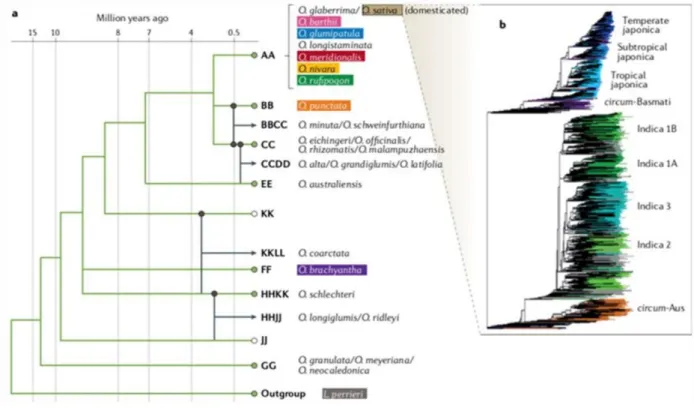
BAGIAN 4. GENETIKA
- Informasi genetik terperinci yang relevan tentang spesies tersebut
- Pendekatan pemuliaan
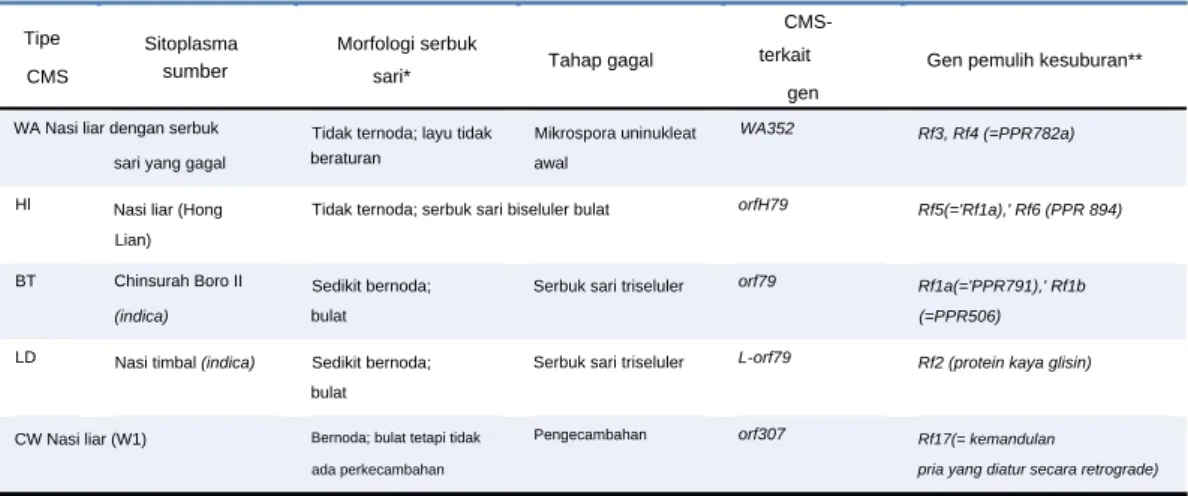
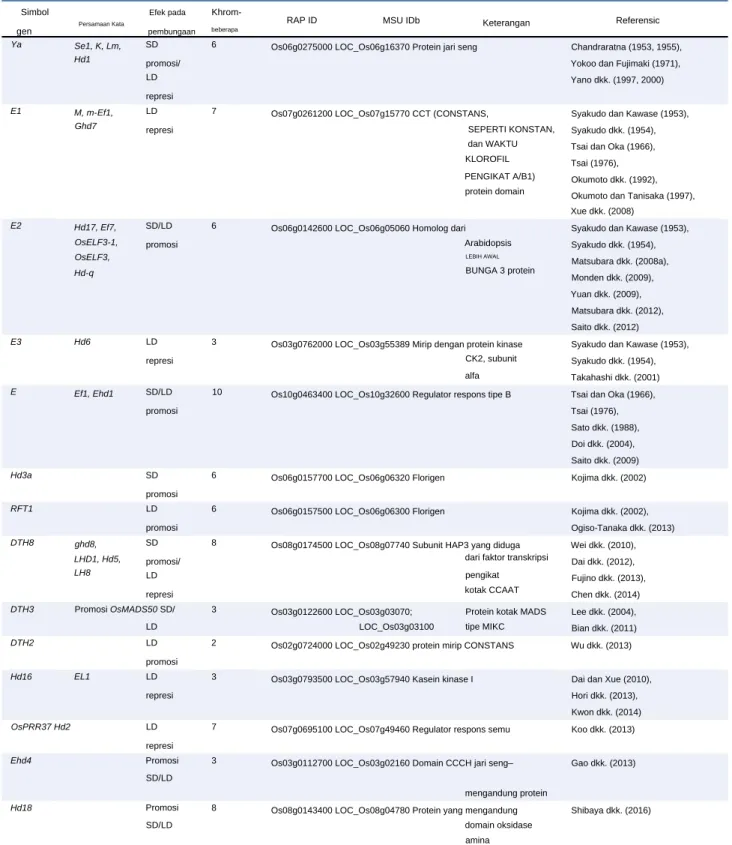
Gen Hd1 (pos tanggal 1) berperan sebagai promotor bunga pada kondisi hari pendek namun berperan sebagai penekan bunga pada kondisi hari panjang (Yano et al., 2000). Untuk ekotipe tropis dan ekotipe Aus japonica yang dibudidayakan di daerah tropis dan subtropis, alel cacat Hd1 (hd1) dominan (Fujino et al., 2010). CMS tipe WA paling umum digunakan untuk tetua betina dalam pemuliaan padi hibrida (diulas dalam Huang et al., 2014).
Jenis CMS lain yang digunakan untuk pemuliaan padi hibrida antara lain CMS tipe BT dan CMS tipe HL (Honglian) (diulas dalam Huang et al., 2014). Terdapat beberapa gen Rf lain yang diketahui bertanggung jawab atas lemahnya pemulihan kesuburan (diulas dalam Huang et al., 2014). Gen dominan laki-laki steril Pingxiang ditetapkan sebagai Ms-p dan dipetakan ke kromosom 10 (Huang et al., 2007).
Alel netral banyak digunakan dalam program persilangan dan pemuliaan hibrida F1 di beberapa negara Asia (Chen et al., 2008). Terungkap bahwa gen Sd1 mengkode GA20 oksidase (Os20ox2) dan fenotip gambar pendek disebabkan oleh hilangnya fungsi sd1 (Sasaki et al., 2002).
BAGIAN 5. HIBRIDISASI DAN INTROGRESI
- Persilangan luar dan aliran gen pada padi
- Produksi eksperimental hibrida interspesifik
- Informasi dan data introgresi alam
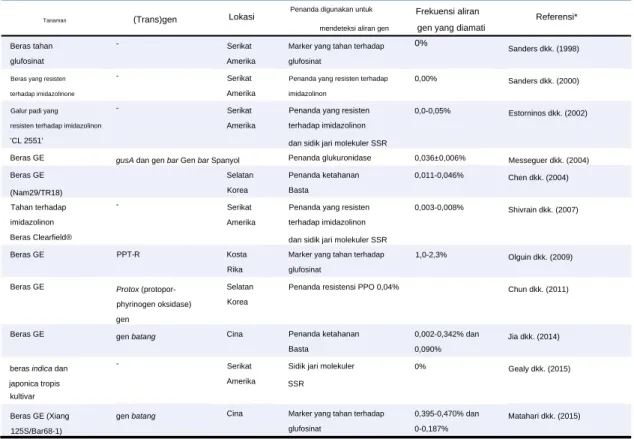
Namun, melalui aliran gen yang dimediasi serbuk sari, transgen dapat berpindah dari padi budidaya ke padi liar (O. sativa f. spontanea) atau salah satu dari enam spesies liar (O. Eksperimen lapangan untuk mendeteksi frekuensi aliran yang dimediasi serbuk sari ( trans ) gen dari padi budidaya ke padi liar, sehingga transgen dapat diekspresikan pada padi liar dan spesies liar dan berpotensi mengubah kebugaran liar/gulma serta dinamika populasi liar.
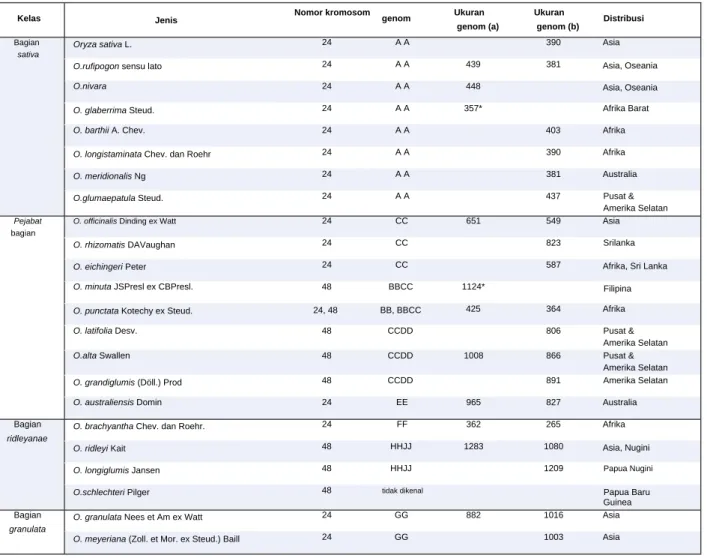
Beberapa hambatan persilangan membatasi aliran gen dari spesies liar ke tanaman padi (Sitch, 1990; Khush dan Brar, 1992). Berbeda dengan spesies liar dengan genom AA, hibrida tidak dapat dihasilkan melalui persilangan langsung antara padi dan spesies liar yang termasuk dalam kompleks officinalis (genom BB, CC, BBCC, CCDD, EE) tanpa penyelamatan embrio di F1. Jenis hibridisasi silang di habitat alami ini biasa terjadi pada padi, padi kurus, dan spesies liar dengan genom AA.
Gen lain yang berguna dari spesies liar seperti Xa21, Xa23 dan Xa38 untuk ketahanan terhadap BB telah diperkenalkan ke dalam padi. Selain itu, banyak galur introgresi yang telah dikembangkan yang mengandung variasi sifat komponen hasil dari lima spesies liar berbeda dengan genom AA (Bhatia et al., 2017).
BAGIAN 6. BERBAGAI INTERAKSI
DENGAN ORGANISME LAIN (EKOLOGI)
Interaksi dalam ekosistem alami dan agroekosistem
- Interaksi alelopati
Pengenalan kultivar modern ke wilayah geografis yang berbeda juga mengakibatkan padi kurus (Kawasaki et al., 2009) akibat hibridisasi indica-japonica. Potensi alelopati mungkin merupakan suatu sifat poligenik dan korelasinya dengan sifat-sifat padi lainnya masih kontroversial (Dilday et al., 1991). Ebana dan Okuno melaporkan analisis QTL dengan beras alelopati (Ebana et al., 2001, Okuno dan Ebana, 2003).
Methanogen yang terdiri dari domain archaea, Methanosaeta, Methanocella dan Methanobacterium merupakan komponen utamanya (Imchen et al., 2019). Diketahui bahwa sebagian besar metana yang dihasilkan dioksidasi di rhizosfer oleh metanotrof (Kögel-Knabner et al., 2010). Banyak jenis mikroba pengikat nitrogen telah ditemukan (59 genera) (Wang et al. 2019), dan beberapa di antaranya diyakini berperan bermanfaat dalam pertumbuhan padi (Banik, Mukhopadhaya, & Dangar, 2016).
Di bawah AOA, Nitrosocaldus (Imchen et al., 2019) dan/atau Nitrososphaera (Chen et al., 2008) dilaporkan melimpah. Dalam kasus AOB, Nitrospira paling umum terjadi di Jepang (Bowatte et al., 2006) dan Cina (Chen et al., 2008).
Pengaruh beras pada organisme yang biasa melakukan kontak dekat
Kelas imunitas lain disebut imunitas yang dipicu PAMP (PTI), di mana komponen sel patogen, PAMP (Pola Molekuler Terkait Patogen), menginduksi respons pertahanan pada padi (Liu et al., 2013). ETI), sangat spesifik pada ras. Protein kecil pengikat GTP, OsRac1, telah terbukti terlibat secara kritis dalam ETI dan PTI melalui siklus aktivasi/inaktivasinya (Liu et al., 2013). Infeksi ini menyebabkan pertumbuhan kerdil yang ekstrim, penggelapan daun dan pengeritingan bagian distal daun muda pada tanaman padi (Wu et al., 2020).
Virus ini berkembang biak di dalam tubuh hama dan dapat ditularkan secara transovarial ke keturunannya (Honda et al., 2007). Namun, beberapa sumber melaporkan bahwa resistensi yang diinduksilah yang berkontribusi terhadap resistensi varietas (Kaloshian dan Walling, 2016; Ling, Ang dan Weilin, 2019; Du et al., 2020). Berdasarkan informasi genom, beberapa gen resistensi BPH telah diisolasi dan dikarakterisasi dari varietas padi tahan BPH dan herba padi liar (Ling et al., 2019;.
Protein BPH14 diduga membentuk homokompleks yang berinteraksi dengan faktor transkripsi WRKY46 dan WRKY72 (Hu et al., 2017). BPH6 mengkodekan protein yang sebelumnya tidak dikarakterisasi yang terlokalisasi pada eksokista dan berinteraksi dengan subunit eksokista OsEXO70E1 (Guo et al., 2018).
Reymond, (2019), “Interaksi Molekuler Antara Tumbuhan dan Serangga Herbivor”, Annual Review of Plant Biology, Vol. 1991), “Aktivitas alelopati padi (Oryza sativa L.) terhadap duckweed (Heteranthera limosa (Sw))”. Yano (2016), “Kontrol genetik waktu berbunga pada padi: integrasi genetika dan genom Mendel”, Genetika Teoritis dan Terapan, Vol. 2015), "Pentatricopeptida mengulangi fungsi protein keluarga RF6 dengan heksokinase 6 untuk menyelamatkan kemandulan sitoplasma jantan pada beras", Prosiding National Academy of Sciences, Vol. 2010), "Peraturan OsSPL14 oleh OsmiR156 mendefinisikan arsitektur tanaman ideal pada padi", Nature Genetics, Vol. 2012), "Endonuklease DNA berpemandu RNA ganda yang dapat diprogram dalam imunitas bakteri adaptif".
Monograf Genetika Teoritis dan Terapan, Vol. 2009), "Konstitusi molekuler padi kurus (Oryza sativa L.) ditemukan di prefektur Okayama, Jepang", Joshi, RC, AT Barrion dan LS Sebastian (ed Rice Black Insects: Taxonomy, Ecology, Biology and Management. Yoshimura (2005) , "Epistasis yang mendasari kemandulan betina terdeteksi pada cacat hibrida pada persilangan padi Japonica-Indica (Oryza sativa L.)", Theoretical and Applied Genetics, Vol. 2010), "Persepsi oligosakarida kitin berkontribusi terhadap ketahanan penyakit terhadap jamur ledakan Magnaporthe oryzae di rice", The Plant Journal, Vol. Reuter and Tytthus chinensis (Steel), dan keberadaannya di sawah”, Kyushu Plant Protection Research, Vol. 1989), Weeds, dilaporkan pada padi di Asia Selatan dan Tenggara, International Rice Research Institute .
Zhang (2018), "Revolusi genom beras: dari biji-bijian kuno menjadi Beras Super Hijau", Nature Review Genetics, Vol. 2007), “Interaksi dua gen resesif, hbd2 dan hbd3, menyebabkan cacat hibrida pada padi”, Theoretical and Applied Genetics, Vol. Pemetaan molekuler Sc, gen sterilitas serbuk sari F1 pada padi budidaya", Euphytica, Vol. 1999) "Pemetaan molekuler lokus.
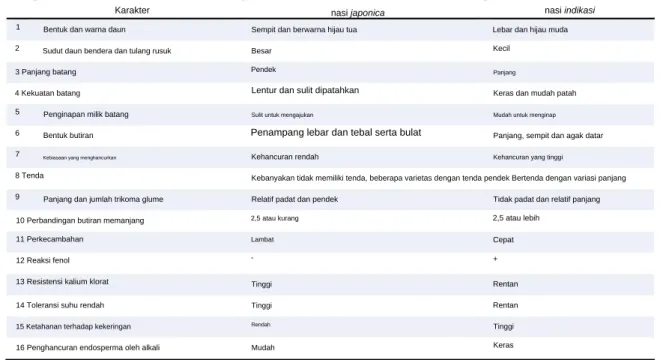
Ciri-ciri tipe ekologi padi
Penyakit jamur dan oomycete
Arthropoda
Arthropoda (lanjutan)
Moluska Nematoda
Beras transgenik
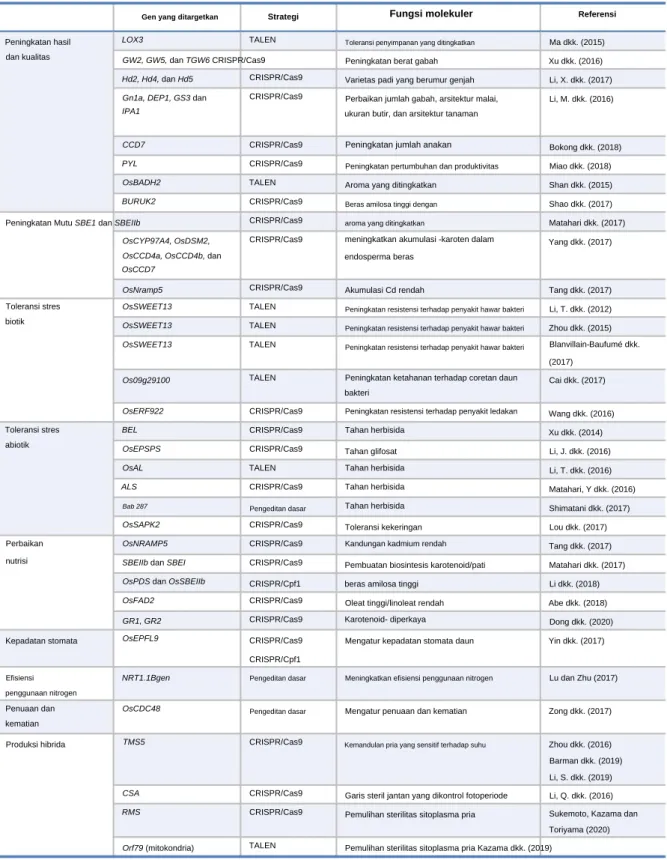
Selain itu, keamanan dan stabilitas vaksin oral berbahan dasar beras yang disebut MucoRice-CTB juga telah terbukti (Azegami et al., 2015). Selain itu, gen Cry (pengkodean Cry1A, Cry1B, Cry1C, Cry1Ab, dan Cry9B) telah diperkenalkan untuk memberikan resistensi terhadap hama Lepidopteran, dan GNA, yang memberikan resistensi terhadap hama Hemipteran termasuk wereng (Shabir et al., 2015). Toleransi herbisida: Mutasi diperkenalkan pada gen ALS untuk membiakkan padi yang toleran herbisida (Li, T. et al., 2016; Sun, Y. et al., 2016).
Pengeditan genom telah berhasil dilakukan pada beras menggunakan kombinasi pengenalan PAM karakter tunggal dan editor dasar (Endo et al., 2019). Selain itu, SpCas9 dirancang untuk mengenali karakter tunggal (-NG) dan dikembangkan untuk menghilangkan batasan rangkaian PAM yang terdiri dari banyak basis (Nishimasu et al., 2018). Resistensi terhadap penyakit: Protein pengikat bakteri di wilayah promotor OsSWEET14 dihancurkan untuk menghasilkan resistensi terhadap hawar daun (Li, T. et al., 2012).
Demikian pula, OsSWEET13 telah dimodifikasi oleh CRISPR/Cas9 untuk mencegah efek Transcription Activator-Like (TAL) (Zhou et al., 2015). Upaya telah dilakukan untuk mengurangi infektivitas rice tungro-spherical virus (RTSV) dan rice tungrobacilliform virus (RTBV) dengan mengganggu urutan terjadinya infeksi ( Macovei et al., 2018 ).
2015), "Mutagenesis LOX3 lipoksigenase berbasis TALEN meningkatkan toleransi penyimpanan benih padi (Oryza sativa), PLoS ONE., Vol. 2016), “Pengembangan galur padi japonica steril jantan genetik peka cahaya dengan penyuntingan kepala sari yang kekurangan karbon (diakses pada 30 Oktober 2020). Jurnal Genetika dan Genomik, Vol. 2007), "Kemajuan dalam bioteknologi padi transgenik", Tinjauan Kritis dalam Ilmu Tanaman, Vol. 2018), “Mutasi pada subfamili gen reseptor asam absisat meningkatkan pertumbuhan dan produktivitas padi”, Prosiding National Academy of Sciences Amerika Serikat, Vol. 2002), “Ekspresi laktoferin manusia dalam butiran beras transgenik untuk aplikasi susu formula”, Plant Science, Vol. 2020), "Percepatan domestikasi beras lokal Afrika yang dimediasi CRISPR", PLoS One, Vol. 2017), "OsSAPK2 menganugerahkan sensitivitas asam absisat dan toleransi terhadap cekaman kekeringan pada padi", Frontiers in Plant Science, Vol. Gheysen (2014), "Infeksi nematoda parasit tanaman pada padi: wawasan molekuler dan seluler", Tinjauan Tahunan Fitopatologi, Vol. menggunakan CRISPR/Cas9”, Jurnal Genetika dan Genomik, Vol. 1989), Gulma, Dilaporkan mengenai padi di Asia Selatan dan Tenggara, IRRI, Filipina.
Zhu (2017), "Pengeditan Basis Target Tepat pada Genom Padi Menggunakan Sistem CRISPR/Cas9 yang Dimodifikasi", Nature, Vol. 2012), “Pengeditan gen berbasis TALEN dengan efisiensi tinggi menghasilkan beras yang tahan penyakit”, Nature Biotechnology, Vol. 2003), “Peningkatan kadar metionin dan sistein pada benih padi transgenik melalui akumulasi albumin 2S wijen”, Biosains, Bioteknologi dan Biokimia, Vol. Akumulasi triptofan pada beras transgenik yang mengekspresikan mutan OASA1 yang tidak peka terhadap umpan balik”, Plant Physiology, Vol. 1989), "Tanaman padi transgenik subur yang diregenerasi dari protoplas yang diubah", Nature, Vol. Toriyama (2020), "Restorasi Kemandulan Jantan Sitoplasma pada Padi Liar Tipe Liar Tiongkok dengan Pengeditan Genom yang Dimediasi CRISPR/Cas9 dari STERILITAS PRIA TERORGANISASI RETROGRADE Berkode Nuklir", Bioteknologi Tanaman, Vol. 2014) “Pengembangan padi tahan penyakit menggunakan komponen pengatur ketahanan penyakit yang diinduksi dengan knockout OsNramp5 menggunakan sistem CRISPR/Cas9 menghasilkan beras indica dengan akumulasi Cd rendah tanpa penurunan hasil”, Laporan Ilmiah, Vol. 2016), “Rekayasa tanaman padi tahan herbisida dengan rekombinasi homolog yang dimediasi CRISPR/Cas9.
Matsumoto (eds.), Asia-Pacific Weed Science Society, Hyderabad, p Pengeditan nukleotida yang ditargetkan menggunakan sistem kekebalan adaptif prokariotik dan vertebrata hibrida Penciptaan beras dengan kadar amilosa tinggi melalui CRISPR/pati mutagenesis yang dimediasi enzim percabangan yang ditargetkan. Cas9,” Frontiers dalam Ilmu Tanaman , vol. 2018), “Rekayasa nuklease CRISPR-Cas9 dengan cakupan penargetan yang diperluas”, Science, vol. 2015), “Beras Transgenik: Kemajuan dan Prestasi”, Kemajuan Rekayasa Genetika,. 2017) "Pengeditan basa yang ditargetkan pada beras dan tomat menggunakan fusi cytidine deaminase CRISPR-Cas9", Alam. 2005), “Meningkatkan nilai gizi Beras Emas dengan meningkatkan kandungan pro-vitamin A”, Nature Biotechnology, Vol. 2015), "Pembuatan beras aromatik dengan knockout gen OsBADH2 yang ditargetkan menggunakan teknologi TALEN", Plant. 2007), "Beras WRKY45 memainkan peran penting dalam resistensi ledakan yang diinduksi benzothiadiazole", The Plant Cell,. dari asetolaktat sintase,” Pabrik Molekuler, Vol. 2006), "Infeksi awal jaringan scutellum dengan Agrobacterium memungkinkan transformasi padi dengan kecepatan tinggi", The Plant Journal, Vol. 2005), "Benih padi yang dimodifikasi secara genetik yang mengakumulasi analog GLP-1 merangsang sekresi insulin melalui https://www.cabi.org/isc/abstract CRISPR/CAS9 yang dimediasi pengeditan gen aroma Badh2 dalam beras", Chinese Journal of Ilmu Padi,.