ISOLAT SIDRAP, MAMUJU DAN BOGOR
LARA HIKMAHAYATI
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
ISOLAT SIDRAP, MAMUJU DAN BOGOR
LARA HIKMAHAYATI
Skripsi
Sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian pada
Departemen Proteksi Tanaman
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
LARA HIKMAHAYATI. Keragaman Gen Protein Selubung Rice Tungro Bacilliform Badnavirus Isolat Sidrap, Mamuju dan Bogor. Dibimbing oleh ENDANG NURHAYATI.
Tungro merupakan penyakit padi yang disebabkan oleh infeksi kompleks dua virus, rice tungro bacilliform badnavirus (RTBV) dan rice tungro spherical waikavirus (RTSV). Beberapa isolat virus tungro dilaporkan memiliki keragaman genetika pada gen protein selubung RTBV. Penelitian ini bertujuan untuk mempelajari keragaman gen protein selubung RTBV isolat Sidrap, Mamuju dan Bogor, dengan metode PCR-RFLP. Isolat virus tungro diperoleh dari dua daerah endemik tungro di Indonesia yaitu varietas TN 1 dari Sidrap (Sulawesi Selatan), varietas Ciherang dari Mamuju (Sulawesi Barat), serta Galur Harapan (GH) dari Bogor (Jawa Barat). Isolat RTBV Sulawesi diperoleh dari koleksi Loka Penelitian Penyakit Tungro, Lanrang, Sidrap, Sulawesi Selatan. Isolat RTBV Bogor diperoleh dari pertanaman padi milik petani di Ciputih, Kecamatan Darmaga, Kabupaten Bogor, Jawa Barat. Semua isolat dipelihara pada tanaman padi varietas rentan IR64 di rumah kaca dengan ditularkan menggunakan vektor Nephotettix virescens Distant (Hemiptera: Cicadellidae). Tanaman padi yang menunjukkan gejala penyakit tungro dipanen pada 10-15 hari setelah inokulasi. DNA total tanaman padi terinfeksi virus tungro diekstraksi dan gen protein selubung RTBV diamplifikasi menggunakan sepasang primer spesifik RTBV-2L dan RTBV-2R. Hasil amplifikasi dipotong dengan 4 enzim restriksi yaitu EcoRV, NsiI, PstIdan
BclI. Pemotongan amplikon hasil PCR menggunakan enzim restriksi pada isolat Bogor tidak menunjukkan adanya keragaman gen protein selubung RTBV karena tidak terpotong oleh keempat enzim tersebut. Sedangkan isolat Sidrap dan Mamuju mempunyai pola fragmen DNA gen protein selubung RTBV yang mirip karena semua isolat tidak terpotong oleh enzim BclI dan terpotong oleh enzim
EcoRV, Nsi1, dan PstI.
NRP : A34061784
Disetujui Dosen Pembimbing
Dr. Ir. Endang Nurhayati, MS. NIP : 19610430 198603 2 001
Diketahui Ketua Departemen
Dr. Ir. Dadang, M.Sc. NIP: 19640204 199002 1 002
Penulis dilahirkan di Majalengka, Jawa Barat pada tanggal 12 September 1988. Penulis adalah putri keempat dari empat bersaudara dari pasangan Bapak Momo Akma Solihat Allahyarham dan Ibu Lili Ruchijati.
Tahun 2006 penulis menamatkan Sekolah Menengah Umum Negeri 1 Maja, Majalengka. Pada tahun yang sama penulis diterima di Institut Pertanian Bogor jalur Undangan Seleksi Masuk IPB (USMI). Penulis selanjutnya memilih program Studi Hama dan Penyakit Tanaman, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor.
Puji dan syukur penulis panjatkan kehadirat Allah Subhanahuwata’ala
atas segala curahan rahmat dan hidayah-Nya, sehingga penulis dapat menyelesaikan laporan tugas akhir ini yang berjudul keragaman selubung protein
rice tungro bacilliform badnavirus isolat Sidrap, Mamuju dan Bogor. Laporan tugas akhir ini disusun berdasarkan hasil penelitian yang dilakukan selama tujuh bulan sejak bulan Februari sampai Agustus 2010.
Pada kesempatan ini penulis mengucapkan terimakasih kepada Dr. Ir. Endang Nurhayati, MS sebagai dosen pembimbing yang telah memberikan saran, bimbingan, dan pengarahan dalam merencanakan dan melaksanakan penelitian serta menyusun laporan akhir ini. Kepada Dra. Dewi Sartiami, M.Si sebagai dosen penguji tamu, penulis mengucapkan terimakasih atas kritik, saran dan bantuannya demi perbaikan laporan tugas akhir ini. Kepada Ir. Djoko Prijono, MAgrSc penulis juga mengucapkan terimakasih atas saran dan bimbingan serta petunjuk dalam penulisan laporan tugas akhir ini pada mata kuliah Teknik Penyajian Ilmiah. Terimakasih juga diucapkan kepada Ir. Titiek Siti Yuliani, SU selaku dosen pembimbing akademik yang senantiasa memberikan perhatian dan semangat selama ini. Terimakasih juga disampaikan kepada seluruh staf pengajar di Departemen Proteksi Tanaman atas bimbingan yang diberikan selama melaksanakan pendidikan.
Penulis juga mengucapkan terimakasih dan penghargaan yang tak terhingga kepada orang tua penulis, Papa Allahyarham dan Mama tercinta atas kasih sayang, semangat, dan dukungan serta doa yang diberikan selama ini. Ucapan terimakasih juga penulis sampaikan kepada kakak-kakak tersayang (Aminudin, Rizal, Ferry, Isa, Yani, Tami) yang tak pernah lelah memberi dorongan dan semangat selama ini. Terima kasih kepada teman-teman seperjuangan di Departemen Proteksi Tanaman yang selalu memberi semangat dan bantuan serta menjadi motivasi dan inspirasi dalam menyelsaikan skripsi ini terutama untuk Dillah, Ita Sulis, Ita Casillas, Teh Leni Allahyarham, Amel, Candra, Herlie, Anto, Atrie, Moya, Teh Didah, serta rekan-rekan HPT lainnya (42, 43, 44, dan 45) yang namanya tidak dapat disebutkan satu persatu. Terimakasih kepada Mbak Tuti atas segala bantuan selama penulis bekerja di Laboratorium Virologi Tumbuhan, terimakasih juga disampaikan kepada anggota Laboratorium Virologi Tumbuhan; Pak Edi, Ibu Latifah, Pak Irwan, Ibu Asni, Mbak Devi, Mbak Pipit dan Ibu Rita atas saran, masukkan serta bantuan yang diberikan serta Pak Saefudin atas bantuannya di Rumah Kaca Cikabayan. Terimakasih juga disampaikan kepada Ibu Fauziah (Loka Penelitian Penyakit Tungro, Lanrang, Sulawesi Selatan) atas bantuan yang diberikan.
Penulis menyadari bahwa skripsi ini masih banyak kekurangannya, oleh karena itu penulis sangat mengharapkan kritik dan saran demi perbaikan laporan tugas akhir ini. Akhirnya semoga laporan ini dapat memberikan manfaat.
Bogor, Oktober 2010
Halaman
DAFTAR TABEL ... ix
DAFTAR GAMBAR ... x
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan Penelitian ... 2
Manfaat Penelitian ... 2
TINJAUAN PUSTAKA ... 3
Tanaman Padi ... 3
Botani dan Morfologi Padi ... 3
Taksonomi Padi ... 4
Varietas Padi ... 4
Penyakit Tungro ... 7
Arti Penting Penyakit Tungro ... 7
Gejala Penyakit Tungro ... 8
Penularan Penyakit Tungro ... 9
Virus Tungro ... 11
Kergaman Gen Protein Selubung RTBV ... 14
Polymerase Chain Reaction Restriction Fragment Length Polymorphism (PCR-RFLP) ... 14
BAHAN DAN METODE ... 16
Tempat dan Waktu Penelitian ... 16
Persiapan Tanaman Padi ... 16
Perbanyakan Nephotettix virescens ... 16
Pengumpulan Isolat Virus Tungro ... 17
Perbanyakan Virus Tungro ... 17
Uji Keragaman Gen Protein Selubung RTBV Menggunakan Teknik PCR-RFLP ... 18
Ekstraksi DNA Total ... 18
Amplifikasi Gen Protein Selubung RTBV ... 19
Pemotongan DNA RTBV dengan Enzim Restriksi ... 19
Elektroforesis dan Visualisasi ... 20
HASIL DAN PEMBAHASAN ... 21
Nomor Halaman 1 Varietas tahan wereng hijau untuk mengendalikan penyakit
tungro ... 5 2 Pewilayahan kesesuaian varietas tahan virus tungro ... 6 3 Masa inkubasi dan kejadian penyakit pada tanaman padi IR64
diinokulasi virus tungro ... 22 4 Ukuran fragmen yang terbentuk pada pemotongan gen protein
selubung empat strain RTBV dari Philipina menggunakan
enzim EcoRV, PstI, BclI ... 26 5 Hasil pemotongan DNA gen protein selubung isolat RTBV
hasil PCR dengan empat enzim EcoRV, NsiI, PstI, dan
Nomor Halaman 1. Gejala serangan virus tungro pada daun tanaman padi ... 9 2. Gambaran skematik genom RTBV, polyprotein P3 dan gen protein
selubung ... 13 3. Hasil amplifikasi gen protein selubung RTBV
dengan PCR menggunakan primer RTBV-2R dan RTBV-2L ... 23 4. Hasil PCR-RFLP dari DNA hasil amplifikasi gen protein
Latar Belakang
Penyakit tungro adalah salah satu penyakit penting pada tanaman padi di
Asia terutama Asia Selatan dan Asia Tenggara. Penyakit ini disebabkan oleh
infeksi dua virus,
rice tungro bacilliform badnavirus
(RTBV) dan
rice tungro
spherical waikavirus
(RTSV) (Hibino
et al
. 1978). Kedua virus itu ditularkan
secara semipersisten oleh vektor yang mempunyai karakter mobilitas yang tinggi
yaitu wereng hijau
Nephotettix virescens
Distant (Hibino dan Cabunagan 1986).
Luas serangan penyakit tungro cukup tinggi bila dibandingkan penyakit
lainnya yang terdapat pada pertanaman padi. Menurut Soetarto
et al.
(2001),
rata-rata luas serangan penyakit tungro setiap tahun mencapai 12.000 ha. Bahkan pada
saat terjadi ledakan, luas serangan di satu propinsi saja dapat menyamai atau
berkali lipat rata-rata luas serangan tahunan nasional. Di Sulawesi Selatan pada
MT 1972/1973 epidemi penyakit tungro mencapai 43.151 ha yang tersebar di
Kabupaten Pinrang, Sidrap, Luwu, dan Polmas (Halteren dan Sama 1973). Pada
akhir tahun 1995 di Surakarta, Jawa tengah ledakan penyakit tungro menyebabkan
sekitar 12.340 ha sawah puso (gagal panen), dan nilai kehilangan hasil akibat
penyakit tersebut diperkirakan setara dengan Rp 25 milyar. Keberadaan penyakit
tungro tersebut ditemukan pula di beberapa daerah di Jawa Barat seperti
Purwakarta, Cianjur, Sukabumi, dan Bogor (Puslitbangtan 1995). Pada tahun 2004
luas serangan penyakit tungro di Sulawesi Tengah mencapai 217 ha terjadi di
Donggala, Parigi, Banggai dan Tolitoli. Sebelumnya, pada musim tanam 2002
serangan virus tungro terjadi di Kabupaten Donggala, Tolitoli dan Parigi Moutong
(Negara
et al
. 2004).
Isolat virus tungro dari beberapa daerah endemis tungro di Indonesia
menunjukkan adanya variasi virulensi. Perbedaan virulensi tersebut dapat dilihat
dari variasi gen protein selubung
yang dapat diketahui melalui pengujian secara
molekuler menggunakan metode PCR-RFLP (Suprihanto 2005). Arfianis (2006)
juga melaporkan adanya keragaman pada gen protein selubung RTBV dari empat
Penggunaan varietas tahan dan galur harapan (GH) tahan tungro merupakan salah satu bentuk dari pengendalian penyakit ini. Varietas tahan penyakit tungro diklasifikasikan sebagai varietas yang tahan terhadap wereng hijau sebagai penular (vektor) patogen dan tahan terhadap virus yang merupakan patogen penyebab penyakit tungro (Widiarta dan Said 2007). Galur harapan merupakan galur-galur padi yang telah lolos dalam pengujian multilokasi yang kemudian dapat dijadikan sebagai varietas baru (Pakki et al. 2007).
Penelitian mengenai variasi virulensi virus tungro yang dapat diketahui melalui keragaman pada gen protein selubung RTBV, sangat bermanfaat sebagai sumber informasi bagi para peneliti dalam upaya pengendalian virus tungro. Selama ini penelitian mengenai keragaman pada gen protein selubung RTBV sebagian besar dilakukan di wilayah endemik tungro di Jawa Barat, sehingga informasi mengenai variasi virulensi spesifik di wilayah endemik tungro lainnya masih kurang. Selain itu, pengujian terhadap keragaman gen protein selubung RTBV pada galur harapan juga belum ada. Oleh karena itu, penelitian mengenai variasi virulensi virus tungro pada wilayah endemik lain selain Jawa Barat serta pada galur harapan perlu dilakukan.
Tujuan Penelitian
Penelitian ini dilakukan untuk mempelajari keragaman gen protein selubung RTBV Isolat Sidrap, Mamuju, dan Bogor dengan metode PCR-RFLP.
Manfaat Penelitian
Tanaman Padi
Padi merupakan tanaman pangan berupa rumput berumpun yang juga merupakan makanan pokok utama bagi lebih dari sepertiga penduduk dunia. Dari semua padi yang diproduksi dan dikonsumsi lebih dari 90% terpusat di Asia. Sebagai sumber pemberi tenaga, beras merupakan bahan makanan utama untuk ratusan juta umat manusia, terutama bagi umat manusia yang menduduki belahan timur dari benua ini. Oleh karenanya tidaklah mengherankan bahwa tanaman padi yang terluas terdapat di negara-negara Asia yang seluruh penduduknya sebagian besar memperoleh tenaganya dari beras sebagai sumbernya (Siregar 1980).
Tanaman padi diduga berasal dari negara India dan Cina kemudian menyebar ke negara-negara lain. Bukti sejarah memperlihatkan bahwa penanaman padi di Zhejiang, Cina sudah dimulai pada 3.000 tahun SM. Fosil butir padi dan gabah ditemukan di Hastinapur, Uttar Pradesh, India sekitar 100-800 SM. Tanaman padi di Indonesia berasal dari perantau-perantau Malaysia yang membawa tanaman padi sekitar tahun 1.500 SM (Siregar 1980).
Botani dan Morfologi Padi
Tanaman padi (Oryza sativa L.) termasuk golongan tumbuhan Gramineae yaitu tumbuhan yang ditandai dengan batang yang tersusun dari beberapa ruas. Tanaman ini juga termasuk golongan tanaman setahun/semusim (Affandi 1997). Bentuk batangnya bulat dan berongga, daunnya memanjang seperti pita yang berdiri pada ruas-ruas batang dan mempunyai sebuah malai yang terdapat pada ujung batang (Siregar 1980).
lihatkan percabangan dimana cabang yang terpendek menjadi lidah daun (ligula), dan bagian yang terpanjang dan terbesar menjadi kelopak. Di dekat lidah daun dan daun kelopak terdapat dua embel sebelah kiri dan kanan yang disebut auricle. Daun kelopak yang membalut ruas yang paling atas dari batang umumnya disebut daun bendera (flag-leaf). Tepat dimana daun pelepah teratas muncul ruas yang disebut bulir padi. Bulir terdiri dari ruas-ruas pendek. Tiap-tiap ruas sebelah kiri dan kanannya timbul cabang-cabang bulir, dan pada ujung tiap-tiap cabang bulir terdapat bunga padi. Bunga padi terdiri dari tangkai bunga, perhiasan bunga dan daun mahkota yang terdiri dari dua belahan yang tidak sama besarnya (Siregar 1980).
Taksonomi Padi
Padi (Oryza sativa L.) merupakan tumbuhan berbiji tunggal (monokotil), dengan urutan taksonomi (Siregar 1980):
Kingdom : Plantae Subkingdom : Tracheobionta Superdivisi : Spermatophyta Divisi : Magnoliophyta
Class : Liliopsida (Monocotyledons) Subclass : Commelinidae
Family : Poaceae Genus : Oryza Spesies : Oryza sativa L.
Varietas Padi
tahan terhadap tumbangnya pertanaman, mutu beras, dan rasa nasi yang lebih enak (Siregar 1980).
Keunggulan dari varietas tersebut tidak bersifat kekal atau abadi. Predikat unggul yang diberikan pada suatu varietas yang tertentu hanya berlaku sebelum ditemukannya varietas baru yang dapat menandingi varietas terdahuluya dalam sifat-sifatnya (Siregar 1980).
Selain varietas unggul tersebut, padi memiliki beberapa varietas yang resisten, moderat (intermediet), dan rentan terhadap hama dan penyakit. Varietas resisten dan moderat terhadap penyakit tungro diklasifikasikan tahan terhadap wereng hijau sebagai penular (vektor) patogen dan tahan terhadap virus yang merupakan penyebab penyakit tungro.
Varietas tahan wereng hijau dikelompokkan berdasarkan sumber gen tetua tahannya menjadi golongan T1, T2, T3, dan T4 (Tabel 1). Anjuran penggunaan varietas tahan wereng hijau adalah (1) di Jawa Barat dapat ditanam varietas tahan golongan T1, T2, dan T4, (2) Jawa Tengah dapat menanam semua golongan varietas tahan, (3) Yogyakarta dianjurkan menanam varietas tahan dari golongan T2 dan T4, (4) Jawa Timur dan Bali hanya dianjurkan menanam varietas tahan golongan T4, dan (5) NTB dianjurkan menanam varietas tahan virus (Widiarta dan Said 2007).
Tabel 1 Varietas tahan wereng hijau untuk mengendalikan penyakit tungro
Golongan Varietas Gen Tahan
To IR5, Pelita, Atomita, Cisadane, Cikapundung, dan Lusi -
T1 IR20, IR30, IR26, IR46, Citarum, dan Serayu Glh1
T2 IR32, IR38, IR36, IR47, Semeru, Asahan, Ciliwung,
Krueng Aceh dan Bengawan Solo Glh 6
T3 IR50, IR48, IR54, IR52 dan IR64 Glh 5
T4 IR66, IR70, IR72, IR68, Barumun, dan Klara. Glh 4
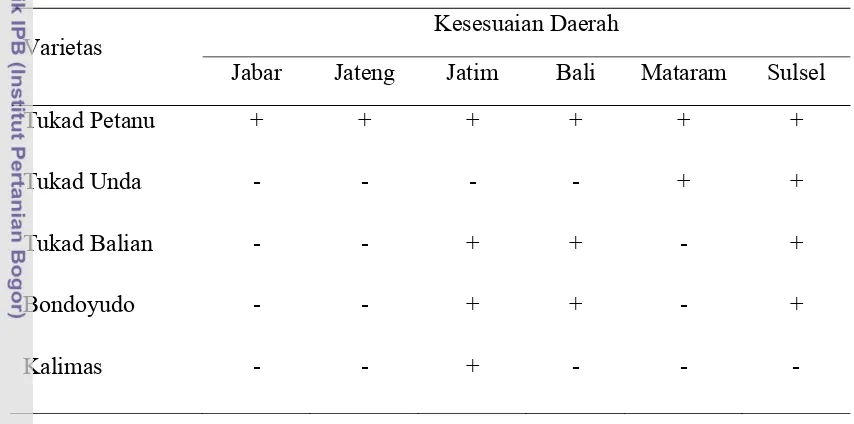
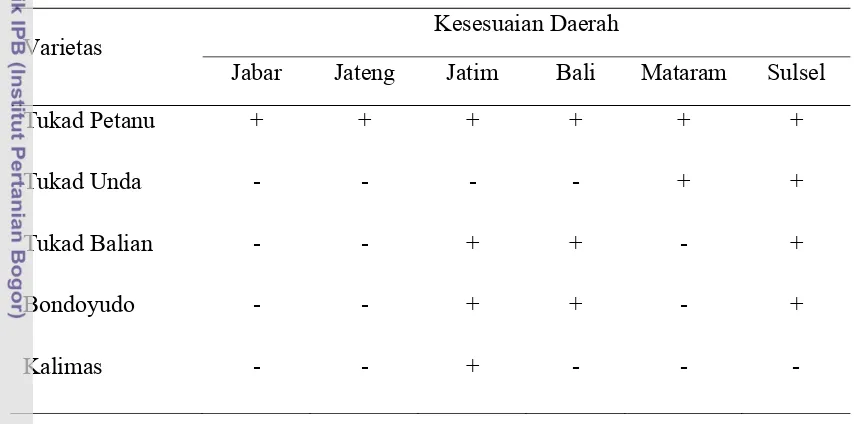
Varietas tahan virus tungro yang telah dilepas antara lain Tukad Petanu, Tukad Unda, Tukad Balian, Kalimas, dan Bondoyudo yang sesuai di setiap daerah (Tabel 2). Varietas Tukad Petanu dapat dianjurkan untuk ditanam di seluruh daerah endemis, sedangkan Tukad Unda dianjurkan ditanam di NTB dan di Sulawesi Selatan. Petani di derah Bali dan Sulawesi Selatan dianjurkan menanam varietas Tukad Balian dan Bondoyudo. Varietas Kalimas dan Bondoyudo diketahui tahan terhadap penyakit tungro di Jawa Timur (Widiarta dan Said 2007).
Tabel 2 Pewilayahan kesesuaian varietas tahan virus tungro
Varietas
Kesesuaian Daerah
Jabar Jateng Jatim Bali Mataram Sulsel
Tukad Petanu + + + + + +
Tukad Unda - - - - + +
Tukad Balian - - + + - +
Bondoyudo - - + + - +
Kalimas - - + - - -
Sumber: Widiarta dan Said 2007 (+): Sesuai (tungro < 50%) (-): Tidak (tungro > 50%)
Dee-Geo-Woo-Gen dan Tsai-Yuan-Chung yang dirakit pada tahun 1949 oleh pemulia tanaman di Taichung District Agricultural Improvement Station, Taiwan. TN1 adalah varietas padi yang rentan terhadap virus tungro dan wereng hijau (Cabautan et al. 1995).
Penyakit Tungro
Arti Penting Penyakit Tungro
Tungro yang berarti ‘pertumbuhan terhambat’, untuk pertama kali ditemukan di Philiphina pada tahun 1963 dan merupakan penyakit yang sangat merugikan. Penyakit ini disebabkan oleh virus yang ditularkan wereng daun, terutama Nephotettix virescens Distant (Semangun 1991).
Di Indonesia, penyakit tungro mula-mula hanya terbatas penyebarannya di daerah tertentu seperti Sulawesi Selatan, Kalimantan Selatan, Nusa Tenggara Barat (NTB), dan Sulawesi Utara. Penyakit ini kemudian menyebar ke Jawa Timur, Jawa Tengah, dan Yogyakarta. Selanjutnya pada awal tahun 1970-an ledakan penyakit tungro dilaporkan terjadi di beberapa daerah sentra produksi padi di Indonesia. Ledakan penyakit tungro terjadi di Bali pada tahun 1980, yang meliputi Kabupaten Badung, Tabanan, dan Gianyar (Rachim 2000).
Daerah endemis penyakit tungro di Indonesia terpusat di daerah sentra produksi padi seperti Sulawesi Selatan, Jawa, Bali dan Nusa Tenggara Barat (Hasanuddin et al. 1995), serta propinsi Sulawesi Tenggara, Papua, dan Sulawesi Tengah terutama di daerah Parigi Moutong (Burhanuddin 2004). Menurut data Balai Informasi Pertanian Palu, luas serangan penyakit tungro di Sulawesi Tengah pada tahun 2004 mencapai 217 ha terjadi di Donggala, Parigi, Banggai dan Tolitoli. Sebelumnya, pada musim tanam 2002 serangan virus tungro terjadi di Kabupaten Donggala, Tolitoli dan Parigi Moutong (Negara et al. 2004).
dalam Agustina 2007). Di Indonesia, kerugian yang dirasakan oleh petani akibat serangan virus tungro pernah terjadi di Sulawesi Selatan tahun 1972, di Bali tahun 1980, dan di Surakarta tahun 1995, dengan kerugian ditaksir milyaran rupiah (Talanca et al. 2007). Pada musim tanam 2005/2006 lalu, virus tungro telah menyerang tanaman padi di NTB dan Manokwari dengan tingkat serangan sedang sampai berat. Bahkan pada musim tanam tahun ini pun produksi padi menurun hingga lebih dari 10% di Bengkulu (Bengkulu Express 2010). Diperkirakan kehilangan hasil akibat serangan virus tungro di seluruh Indonesia rata-rata 12.000 ha/tahun atau kerugiannya senilai Rp 48 miliar/tahun (asumsi harga gabah Rp 1.000/kg). Karenanya penyakit ini perlu diantisipasi, terutama di daerah endemis seperti Sulawesi Selatan, Sulawesi Tengah, NTB, Papua, Jawa Tengah, Jawa Timur, dan Bali (Puslitbangtan 2007).
Gejala Penyakit Tungro
Penyakit tungro disebabkan oleh dua jenis virus yaitu rice tungro bacilliform badnavirus (RTBV) dan rice tungro spherical waikavirus (RTSV). Tanaman padi yang terinfeksi kedua virus tersebut menjadi kerdil dan kelihatan belang serta perubahan warna kuning sampai oranye pada daun. Gejala tungro yang khas dapat disebabkan oleh RTBV dan gejala dapat diperkuat dengan kehadiran RTSV (Agrios 1997).
Gambar 1 Gejala serangan virus tungro pada daun tanaman padi. Warna daun yang menguning (orange), dimulai dari ujung daun dan berkembang kebagian lamina daun bawah ([email protected] ).
Tanaman yang terinfeksi virus tungro tumbuh kerdil, jumlah anakan sedikit, helaian daun dan pelepah daun memendek. Pada bagian bawah helaian daun muda terjepit oleh pelepah daun sehingga daunnya terpuntir atau menggulung sedikit. Malai pendek, gabah tidak terisi sempurna atau kebanyakan hampa dan terdapat bercak-bercak coklat yang menutupi malai. Infeksi virus tungro pada tanaman tua (umur di atas 50 hari setelah tanam) kurang berpengaruh terhadap produksi dan tanaman tidak menampakkan gejala serangan sampai panen (Ling 1972).
Penurunan jumlah anakan sangat tinggi bila infeksi terjadi pada stadium pertumbuhan sangat awal. Jumlah anakan tanaman padi dipengaruhi umur dan mungkin akan meningkat bila infeksi virus tungro setelah tanaman berumur lebih dari satu bulan. Namun jumlah anakan akan tetap sedikit jika selama infeksi terjadi, stadium petumbuhan terhambat (Ling 1972).
Penularan Penyakit Tungro
Virus tungro ditularkan secara semipersisten oleh wereng daun
Serangga penular virus tungro terutama adalah wereng hijau (N. virescens Distant, N. nigropictus (Stal), N. malayanus dan N. parvus). Serangga lain yang dapat juga sebagai penular virus tungro, namun kurang efisien adalah wereng loreng Recilia dorsalis (Motsch). Rentang efisiensi penularan virus oleh populasi
N. virescens antara 35 - 83%, dibandingkan dengan N. nigropictus yang rentang efisiensinya antara 0 - 27%. Spesies wereng hijau lainnya seperti N. malayanus
dan N. parvus memiliki kemampuan menularkan virus berturut-turut 40% dan 7% lebih rendah dari N. virescens (Deptan 1986).
Nephotettix sp. dikenal sebagai wereng hijau. Serangga ini menyerang bagian daun tanaman padi. Serangga dewasa tersebut berukuran 4-6 mm. Telurnya berbentuk bulat panjang atau lonjong berwarna terang (kuning pucat), berukuran 1,3 x 0,30 mm. Telur ini diletakkan berderet sebanyak 5-25 butir. Wereng daun betina mampu bertelur 200-300 butir yang diletakkan di dalam jaringan pelepah daun. Telur tersebut menetas setelah 4-8 hari dan membentuk serangga muda (nimfa). Nimfa ini mengalami 5 kali ganti kulit selama 16-18 hari, kemudian menjadi dewasa setelah 2-3 hari. Terdapat dua jenis Nephotettix sp yang dominan yaitu N. virescens dan N. nigropictus. Spesies N. virescens
berwarna hijau kekuningan dengan ujung kepala meruncing. N. virescens jantan mempunyai ukuran 4 mm dan N. virescens betina 6 mm, sedangkan nimfa
N. virescens berwarna hijau kekuningan hingga hijau tua. Spesies N. nigropictus
berwarna hijau tua dengan ujung kepalanya agak tumpul dan dilengkapi dengan
garis pita hitam yang jelas di atas bagian kepalanya. Serangga jantan
N. nigropictus berukuran 3,6 mm, sedangkan nimfa N. nigropictus berwarna kuning coklat hingga gelap (Deptan 1986).
Cabautan dan Hibino (1984) melaporkan bahwa wereng hijau dapat memindahkan RTSV dari tanaman padi yang hanya terinfeksi RTSV, tetapi tidak mampu memindahkan RTBV dari tanaman yang hanya terinfeksi RTBV. RTBV hanya dapat dipindahkan oleh wereng hijau dari tanaman yang telah terinfeksi RTSV. Dengan demikian RTBV merupakan virus dependent sedangkan RTSV berfungsi sebagai helper. Kedua partikel virus tersebut bersifat noncirculative, yaitu dalam tubuh vektor virus tidak dapat ditularkan dari imago ke telur maupun stadia perkembangan imago (Ling 1966). Disamping itu virus tungro juga tidak dapat ditularkan melalui biji, tanah, air dan secara mekanis (misal pergesekan antara bagian tanaman yang sakit dengan yang sehat). Nimfa wereng hijau juga dapat menularkan virus tungro, tetapi menjadi tidak infektif setelah ganti kulit (Widiarta 2005).
Fluktuasi kepadatan populasi vektor virus tungro sangat mempengaruhi keberadaan tanaman terinfeksi virus tungro bila sumber inokulum virus ini sudah ada di lapang. Persentase tanaman terinfeksi virus tungro yang tinggi pada musim hujan (Desember hingga April) bertepatan dengan kepadatan populasi wereng hijau yang tinggi pada periode yang sama. Sebaliknya pada musim kemarau (Mei sampai November), persentase tanaman terinfeksi virus tungro yang rendah bersamaan dengan kepadatan populasi wereng hijau yang lebih rendah daripada musim hujan (Widiarta 2005).
Virus Tungro
RTSV termasuk kedalam famili Sequiviridae genus Waikavirus. RTSV mempunyai genom poliadenil ssRNA, unipartit, terbungkus partikel isometrik dengan diameter 30 nm (Hibino et al. 1978). Genom RNA RTSV kira-kira 11 kb (kilo base) dan protein selubungnya terbentuk dari dua jenis molekul protein (Agrios 1997).
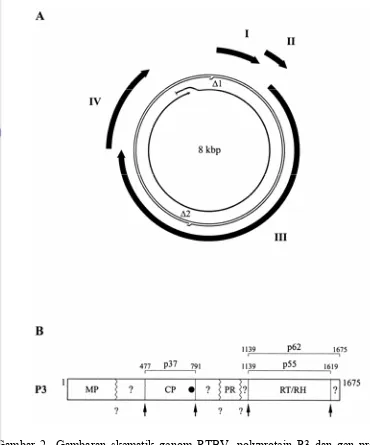
RTBV termasuk famili Caulimoviridae dan genus Badnavirus. Bentuk partikel RTBV adalah bacilliform dengan diameter 30-35 nm dan panjang kira-kira 100-300 nm yang bervariasi antar isolat (Hibino et al. 1978). Asam nukleat RTBV adalah DNA utas ganda dan bulat lebih kurang 8 kb. Asam nukleat tersebut mengandung dua daerah yang tidak bersambung yang merupakan hasil dari proses replikasi oleh reverse transcriptase dan empat open reading frames
Keragaman Gen Protein Selubung RTBV
Pendiagnosisan keragaman virus tungro dengan melihat gejala dan analisis RFLP telah dilakukan pada isolat dari Philippina (Azzam dan Chancellor 2002a) dan Indonesia (Suprihanto 2005). Dari diagnosis tersebut diperoleh pengetahuan bahwa virus tungro memiliki perbedaan pada setiap lokasi. Populasi virus tungro secara geografi dilaporkan hanya stabil pada periode waktu tertentu. Ini menunjukkan bahwa virus tungro memberikan respon yang berbeda terhadap perubahan lingkungan dan inang. Studi lingkungan menunjukkan bahwa pada satu lokasi virus tungro yang memiliki keragaman secara genetik dan biologi dapat hidup berdampingan (Azzam dan Chancellor 2002b).
Cabautan et al. (1995) melaporkan bahwa ada keragaman pada empat strain RTBV (G1, G2, Ic dan L) dari Philippina berdasarkan gejala yang berbeda pada varietas padi FK135 dan TN1. Uji RFLP terhadap genom empat strain RTBV di atas menunjukkan pola RFLP yang beragam. Suprihanto (2005) juga melakukan uji penularan virus tungro pada tanaman diferensial FK 135 dan TN1, dan uji PCR-RFLP terhadap delapan isolat RTBV yang diambil dari daerah endemis tungro di Indonesia. Berdasarkan gejala yang diamati dan pola PCR-RFLP diketahui bahwa delapan isolat RTBV berturut-turut menyebabkan gejala yang berbeda terutama pada warna daun dan keragaman pada gen protein selubungnya. Demikian juga Arfianis (2006) melakukan uji diferensiasi dengan PCR-RFLP empat isolat RTBV yang diambil dari daerah endemis tungro di Jawa Barat. Hasil yang diperoleh pun menunjukkan adanya keragaman pada gen protein selubung RTBV. Pengujian terhadap variasi genetik protein selubung RTBV pada tingkat luasan hamparan padi disuatu wilayah pada varietas juga dilaporkan oleh Agustina (2007). Hasil yang diperoleh yaitu terdapatnya keragaman pada gen protein selubung RTBV.
Polymerase Chain Reaction
Restriction Fragment Length Polymorphism ( PCR-RFLP)
B. Mullis, seorang peneliti di perusahaan CETUS Corporation. Metode PCR sangat sensitif karena dapat digunakan untuk melipatgandakan suatu molekul DNA. Metode ini juga sering digunakan untuk memisahkan gen-gen kanopi tunggal dari sekelompok sekuen genom. PCR mensyaratkan bagian tertentu sekuen DNA yang dilipatgandakan harus diketahui terlebih dahulu sebelum proses pelipatgandaan tersebut dilakukan. Sekuen yang diketahui tersebut penting untuk menyediakan primer, yaitu suatu sekuen oligonukleotida pendek yang berfungsi mengawali sintesis rantai DNA dalam reaksi berantai polimerase (Yuwono 2006 ). Proses PCR pada dasarnya terdiri atas tiga tahap reaksi dengan kondisi suhu yang berbeda secara berulang dalam beberapa siklus tertentu yaitu denaturasi, annealing (penempelan primer) dan ekstensi primer (sintesis DNA). Dengan reaksi amplifikasi DNA secara simultan, maka jumlah sasaran akhir telah dilipatgandakan secara eksponensial (Mc Pherson et al. 1992 dalam Mutaqin 2000). Proses sintesis inilah yang membuat sensitifitas teknik PCR semakin tinggi, karena dari jumlah molekul DNA yang sedikit dapat dikopi menjadi berlipat ganda (Takahashi et al. 1993).
Tempat dan Waktu Penelitian
Penelitian ini dilaksanakan di Rumah Kaca Cikabayan dan Laboratorium Virologi Tumbuhan, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor, dari bulan Februari sampai Agustus 2010.
Persiapan Tanaman Padi
Tanaman padi varietas IR64 disiapkan untuk perbanyakan N. virescens
dan isolat virus tungro. Benih padi IR64 direndam pada wadah yang berisi air selama 1 malam. Kemudian benih tersebut ditabur pada baki yang telah berisi campuran tanah dan pupuk kandang, dengan perbandingan 2:1. Benih IR64 ditabur sebanyak 15-20 butir tiap baki. Bibit yang tumbuh dipelihara dalam rumah kaca yang bebas dari serangga sampai digunakan.
Perbanyakan Masal Nephotettix virescens
Pengumpulan Isolat Virus Tungro
Isolat virus tungro diperoleh dari dua daerah endemik tungro di Indonesia, yaitu varietas TN 1 dari Sidrap, Sulawesi Selatan; varietas Ciherang dari Mamuju, Sulawesi Barat; dan galur harapan (GH) dari Bogor; Jawa Barat. Isolat RTBV Sulawesi diperoleh dari koleksi Loka Penelitian Penyakit Tungro, Lanrang, Sidrap, Sulawesi Selatan. Isolat RTBV Bogor diperoleh dari pertanaman padi milik petani di Ciputih, Kecamatan Darmaga, Kabupaten Bogor, Jawa Barat.
Perbanyakan virus tungro
Perbanyakan inokulum dilakukan pada tanaman padi IR64 dengan cara ditularkan dengan N. virescens mengikuti prosedur dari Azzam et al. (2000).
Uji Keragaman Gen Protein Selubung RTBV Menggunakan Teknik PCR-RFLP
Uji ini dilakukan untuk mengetahui keragaman gen protein selubung virus. Uji ini terdiri dari beberapa tahap, yaitu ekstraksi DNA total, amplifikasi gen protein selubung RTBV dan pemotongan DNA hasil amplifikasi dengan beberapa enzim endonuklease.
Ekstraksi DNA total
Ekstrak DNA RTBV disiapkan dari daun padi varietas rentan IR64 yang terinfeksi tungro menggunakan metode yang dimodifikasi dari Smith et al. (1992). Daun padi sakit sebanyak 0,3 g ditambah dengan nitrogen cair digerus dengan mortar dan pistil sampai berbentuk bubuk. Bubuk daun tersebut kemudian dimasukkan ke dalam tabung mikro 2 ml dan ditambah 1 ml bufer ekstrak, yang mengandung 50 mg/ml polyvinilpyrrolidone (PVP) dan 60 µl 10% sodium dedocyl sulfate (SDS), serta ditambah 10 µl mercapto etanol. Tabung tersebut divorteks sampai rata lalu dimasukkan ke dalam penangas air dengan suhu 65 oC selama 30 menit. Setelah itu, tabung didinginkan sampai dengan suhu ruang dan ditambahkan 750 µl chloroform : isoamil alcohol (CI 24:1). Kemudian tabung yang berisi ekstraktan divorteks dan disentrifugasi pada 11.000 rpm selama 10 menit.
Supernatan yang terbentuk pada proses tersebut dipisahkan dan diletakkan di tabung baru. Supernatan tersebut kemudian ditambahkan 1 ml chloroform
kemudian divorteks dan disentrifugasi pada 11.000 rpm pada suhu 4 oC selama 10 menit. Supernatan yang dihasilkan kemudian ditambah 1 ml isopropanol dingin kemudian dihomogenkan, dengan cara membalikkan tabung beberapa kali, lalu diinkubasi pada suhu -20 oC selama 30 menit dan disentrifugasi pada 11.000 rpm pada suhu 4 oC selama 10 menit. Pelet diresuspensi dengan 200 µl TE pH 8.0 (10 mM Tris Cl pH 8.0 dan 1 mM EDTA pH 8.0), kemudian dihomogenkan dengan cara membalikkan tabung beberapa kali kemudian diinkubasi pada suhu 37 oC selama 1 jam. Pelet yang dihasilkan ditambah 200 µl
membalikkan tabung beberapa kali lalu diinkubasi pada suhu -20 oC selama 1 malam (over night) dan disentrifugasi pada 14.000 rpm pada suhu 4 oC selama 10 menit. Cairan tersebut dibuang kemudian pelet dicuci dengan 500 µl ethanol
dingin 75%, dikeringkan dan diresuspensi dengan 100 µl bufer TE kemudian disimpan pada suhu -20 oC sampai digunakan pada metode selanjutnya.
Amplifikasi Gen Protein Selubung RTBV
Amplifikasi dilakukan dengan metode PCR menggunakan primer spesifik RTBV yaitu RTBV-2L (5’-GGTCTTGGATGGATGGTAGA-3’) dan RTBV-2R (5’-GCTGAGGTGCTACATAGGTT-3’). Sepasang primer tersebut didesain untuk mengamplifikasi pada bagian gen protein selubung sampai sebagian gen protease aspartat RTBV, yang menghasilkan produk 1497 bp (Venkitesh et al.
1994). Sebanyak 0,4 pmol masing-masing primer, 2 unit Taq DNA polymerase (Invitrogen, TECH-LINE USA), 1x bufer PCR, 1,5 mM MgCl2, 0,2 mM dNTP,
dan 2 ml DNA template dalam volume akhir 25 µl digunakan dalam amplifikasi. Amplifikasi ini dilakukan pada DNA thermal cycler (Gene Amp PCR System 9700, PE Applied Biosystems, USA). Amplifikasi tersebut didahului dengan denaturasi awal selama 5 menit pada 94 °C. Satu siklus amplifikasi meliputi denaturasi 1 menit pada 94 °C, penempelan primer (annealing) selama 1 menit pada 55 °C, sintesis selama 2 menit pada 72 °C dengan pengulangan sebanyak 34 kali, kemudian untuk tahapan sintesis ditambah 10 menit pada 72 °C.
Pemotongan DNA Protein Selubung RTBV dengan Enzim Retriksi
tersebut diinkubasikan pada suhu 37 °C selama 3 hari untuk enzim PstI, NsiI dan
EcoRV sedangkan enzim BclI pada suhu 50 °C selama 3 hari.
Situs pemotongan DNA protein selubung RTBV dengan enzim restriksi :
Elektroforesis dan Visualisasi
Penyiapan Gel Agarose. Gel agarose dengan konsentrasi 1,5% disiapkan dengan mencampurkan bubuk agarose dengan bufer Tris-acetat EDTA (TAE) 1x (0,045 M Tris-acetat, 0,01 M EDTA) dan dimasukkan ke dalam tabung erlenmeyer 100 ml. Campuran dipanaskan dalam microwave sampai agarose larut sempurna kemudian didinginkan sampai kira-kira 60 oC lalu ditambahkan
ethidium bromida. Sebelumnya, aparatus pencetak gel dibersihkan, dikeringkan, kemudian diletakkan pada permukaan yang datar. ”Sisir” gel diletakkan di bagian atas aparatus pencetak gel (± 0,5-1,0 mm dari atas). Selanjutnya larutan agarose
dituang ke dalam aparatus pencetak gel. Setelah gel agarose mengeras (30-45 menit), dengan hati-hati sisir gel dilepas dan gel diletakkan dalam bak
elektroforesis (Bio-Rad Power PAC 300, USA). Selanjutnya ditambahkan bufer elektroforesis TAE 1 x hingga gel agarose terendam.
Elektroforesis DNA Hasil PCR dan DNA Hasil Restriksi. Sebanyak 7 µl DNA hasil PCR sebagai kontrol tanpa pemotongan dan 15 µl untuk DNA hasil restriksi dimasukkan ke dalam sumuran gel dengan pipet mikro kemudian salah satu sumuran gel dimasukkan 10 µl marker 1 kb DNA ladder (Fermentas, USA). Elektroforesis dilakukan dengan tegangan 50 volt selama 60 menit. Hasil elektroforesis divisualisasikan dengan transluminator ultraviolet (Sambrook et al.
1989). Pita DNA yang terbentuk pada hasil elektroforesis tersebut diamati dan dipotret dengan menggunakan kamera digital.
5’C T G C A G3’ 3’G A C G T C5’ Pst I
5’A T G C A T3’ 3’G A C G T C5’ Nsi I
5’G A T A T C3’ 3’C T A T A G5’ EcoRV
Gejala Infeksi, Masa Inkubasi, dan Kejadian Penyakit pada Tanaman Padi yang Diinokulasi Virus Tungro
Tanaman sumber inokulum virus tungro yang diambil dari lapangan adalah tanaman padi yang menunjukkan gejala penyakit tungro berupa perubahan warna daun dari hijau menjadi kuning sampai kuning-oranye yang didasarkan pada deskripsi gejala dari Puslitbangtan (2007). Hasil penularan pada padi varietas rentan IR64 menunjukkan bahwa terdapat perbedaan respon tanaman terhadap beberapa isolat virus tungro.
Tabel 3 Masa inkubasi dan kejadian penyakit pada tanaman padi IR64 yang diinokulasi virus tungro
Asal isolat Masa Inkubasi (hari)
Kejadian Penyakit ** (%)
Bogor
GH* 1 15-17 60
GH* 2 15-17 60
GH* 3 15-17 60
GH* 4 15-17 60
Sidrap 10-14 100
Mamuju 10-14 100
* Galur Harapan
** Persentase jumlah tanaman yang terinfeksi berdasarkan jumlah tanaman yang menunjukkan gejala dibandingkan dengan jumlah tanaman yang diinokulasi
Masa inkubasi virus tungro pada isolat Sidrap dan Mamuju pada varietas rentan IR64 lebih cepat yaitu 10-14 hari (Tabel 3) dibandingkan dengan isolat Bogor (GH 1-4) dengan waktu 15-17 hari (Tabel 3). Kejadian penyakit pada isolat Sulawesi (Sidrap dan Mamuju) 100%, sedangkan pada isolat Bogor 60%. Hal tersebut menunjukkan bahwa isolat Sidrap dan Mamuju mampu menyebabkan penyakit lebih cepat dan banyak dibandingkan isolat Bogor.
virus adalah tanaman yang mampu menghambat replikasi dan penyebaran virus di dalam tanaman atau perkembangan gejala, sehingga konsentrasi virus di dalam tanaman menjadi rendah, sedangkan tanaman rentan adalah tanaman yang tidak mampu menghambat replikasi dan penyebaran virus di dalam tanaman yang dicirikan dengan konsentrasi virus yang tinggi dan masa inkubasi atau munculnya gejala yang cepat (Goodman et al. 1986). Hal ini terjadi pada isolat Sidrap dan Mamuju yang berasal dari varietas padi TN1 dan Ciherang yang telah diketahui rentan terhadap virus tungro (Cabautan et al. 1995 dan Syam 2007).
Amplifikasi Gen Protein Selubung RTBV
Gen protein selubung RTBV berhasil diamplifikasi dari DNA total tanaman padi IR64 yang diinokulasi dengan 6 isolat virus tungro. Amplifikasi gen tersebut dilakukan dengan menggunakan primer spesifik RTBV-2R dan RTBV-2L (Venkitesh et al 1994). Hasil amplifikasi menunjukkan ukuran fragmen DNA yang sama dari semua isolat yaitu 1500 bp (Gambar 3). Hal tersebut sesuai dengan laporan Venkitesh et al. (1994) yang menyatakan bahwa virus tungro dapat diamplifikasi pada bagian gen protein selubung sampai sebagian gen
aspartic protease (PR) RTBV yang menghasilkan produk 1496 bp.
Gambar 3 Hasil amplifikasi gen protein selubung RTBV dengan PCR menggunakan primer RTBV-2R dan RTBV-2L. M= Marker 1 kb DNA ladder (Fermentas, USA), 1= Kontrol negatif (Tanaman Sehat), 2= Kontrol positif Isolat No 35 (Ladja 2008), 3= Isolat GH 1, 4= Isolat GH 2, 5= Isolat GH 3, 6= Isolat GH 4, 7= Isolat Sidrap (Sulawesi Selatan), 8= Isolat Mamuju (Sulawesi Barat) bp= base pairs
4 5 6 7 8 3
2 1 M
±1000 bp
Karakteristik Gen Protein Selubung RTBV dengan PCR-RFLP
A B C D
Gambar 4 Hasil PCR-RFLP dari DNA hasil amplifikasi gen protein selubung RTBV dengan enzim EcoRV (a), NsiI (b), PstI (c) dan BclI (d), M= Marker 1 kb DNA ladder (Fermentas, USA), 1= tidak dipotong, 2= Isolat GH 1, 3= Isolat GH 2, 4= Isolat GH 3, 5= Isolat GH 4, 6= Isolat Sidrap (Sulawesi Selatan), 7= Isolat Mamuju (Sulawesi Barat), bp= base pairs
4 5 6 7 3
2 1 M
4 5 6 7 3
2 1 M
4 5 6 7 3
2 1 M
4 5 6 7
3 2 1 M
± 1000 bp ± 1500 bp
4 5 6 7 3
2 1 M
± 250 bp ± 1000 bp
± 1500 bp
± 1270 bp
± 230 bp
4 5 6 7
3 2 1 M
± 500 bp ± 1000 bp ± 1500 bp
± 474bp ± 1023 bp
4 5 6 7
3 2 1 M
± 701 bp ± 1220 bp ± 750 bp
± 523 bp ± 273 bp ± 250 bp
± 500 bp ± 1000 bp ± 1500 bp
4 5 6 7
Tabel 4 Hasil pemotongan DNA gen protein selubung RTBV hasil PCR dengan empat enzim EcoRV, NsiI, PstI dan BclI
Isolat RTBV Hasil Pemotongan
EcoRV NsiI PstI BclI Bogor
GH 1 - - - -
GH 2 - - - -
GH 3 - - - -
GH 4 - - - -
Sidrap + + ++ -
Mamuju + + ++ -
bp: base pair, (-) Tidak terpotong.
EcoRV (+) Kira-kira berukuran 1270 bp dan 230 bp
NsiI (+) Kira-kira berukuran 1023 bp dan 474 bp
PstI (++) Kira-kira berukuran 1220 bp, 701 bp, 523 bp, dan 273bp
Ukuran-ukuran fragmen DNA tersebut (Gambar 4 dan Tabel 4) dianalogikan dengan ukuran fragmen DNA gen protein selubung RTBV dari empat isolat Philiphina yang disajikan pada Tabel 5. Ukuran-ukuran fragmen DNA tersebut diperoleh dari analisis menggunakan Premier Biosoft Internasional 2004 (USA) dengan primer yang sama dalam penelitian ini. Sama halnya dengan hasil penelitian Suprihanto (2005) dan Agustina (2007), pada penelitian ini ditemukan gen protein selubung RTBV yang terpotong oleh EcoRV menjadi dua fragmen yang kira-kira berukuran ±1270 bp dan ±230 bp yang tidak ditemukan pada gen protein selubung protein RTBV isolat-isolat dari Philiphina.
Tabel 5 Ukuran fragmen yang terbentuk pada pemotongan gen protein selubung empat strain RTBV dari Philipina menggunakan enzim EcoRV, PstI,
BclI* Isolat RTBV/Sumber
Ukuran Fragmen (bp)
Pst1 Nsi1 EcoRV Bcl1 Phi-1/X57924 1220, 277 1023, 474 1409, 88 959, 538 G1/ AF113830 701, 524, 272 1023, 474 772, 651, 74 958, 539 G2/AF113831 1220, 277 1023, 474 1409, 88 959, 538 Ic/AF113832 701, 523, 273 1023, 474 771, 523, 203 957, 540
Pemotongan fragmen DNA gen protein selubung RTBV hasil PCR dengan enzim EcoRV, NsiI, PstI, dan BclI pada galur harapan isolat Bogor ternyata tidak membedakan semua isolat (Isolat GH 1-4). Hal tersebut terjadi karena semua galur harapan isolat Bogor tidak terpotong oleh keempat enzim yang digunakan. Pemotongan dengan enzim EcoRV, NsiI, PstI, dan BclI memperlihatkan perbedaan antara isolat Bogor dengan isolat Bogor yang digunakan Agustina (2007) dan isolat Jabar-2 (Bogor) yang digunakan Suprihanto (2005). Hal ini terjadi karena isolat Bogor yang digunakan Agustina (2007) dan isolat Jabar-2 (Bogor) yang digunakan Suprihanto (2005) diambil dari daerah yang berbeda dengan isolat Bogor dalam penelitian ini. Hal ini menunjukkan dalam satu daerah yang sama dalam lokasi yang berbeda memungkinkan adanya variasi virulensi (Suprihanto 2005). Menurut Arboleda et al. (1999) dalam satu lokasi bisa ditemukan lebih dari satu isolat RTBV yang berbeda.
Fragmen DNA gen protein selubung RTBV isolat Sidrap dan Mamuju, sama halnya dengan isolat gen protein selubung RTBV Bogor, tidak terpotong oleh enzim BclI. Namun fragmen DNA isolat RTBV Sidrap dan Mamuju terpotong oleh ketiga enzim lainnya, yaitu enzim EcoRV, NsiI, dan PstI dengan pola dan ukuran yang sama. Isolat RTBV Sidrap dan Mamuju kemungkinan mempunyai urutan nukleotida pada fragmen amplikon gen protein selubung RTBV yang mirip, sehingga dapat terpotong oleh ketiga enzim tersebut. Gen protein selubung RTBV isolat Sidrap dan Mamuju memiliki kekerabatan yang dekat dengan gen protein selubung RTBV isolat Sulawesi Selatan (Lanrang) (Suprihanto 2005), karena dapat terpotong oleh enzim EcoRV, NsiI, dan PstI. Gen protein selubung RTBV Sidrap dan Mamuju meskipun terpotong oleh ketiga enzim tersebut, yaitu EcoRV, NsiI, dan PstI, tetapi pada pemotongan dengan
Tabel 6 Hasil pemotongan DNA gen protein selubung isolat RTBV hasil PCR dengan empat enzim EcoRV, NsiI, PstI, dan BclI dari penelitian sebelumnya
Isolat RTBV Hasil Pemotongan Sumber
EcoRV NsiI PstI BclI
1 + + - + Agustina (2007)
3 ++ - + + Agustina (2007)
5 - + - + Agustina (2007)
7 + + - + Agustina (2007)
9 ++ + - - Agustina (2007)
Jabar-2 (Bogor) + + - TAD Suprihanto (2005) Sulawesi Selatan (Lanrang) + + + TAD Suprihanto (2005)
bp= base pair, (-) Tidak terpotong, (TAD) Tidak Ada Data.
EcoRV (+) Kira-kira berukuran 1270 bp dan 227 bp
EcoRV (++) Kira-kira berukuran 771 bp, 523 bp, dan 203 bp
NsiI (+) Kira-kira berukuran 1023 bp dan 474 bp
PstI (+) Kira-kira berukuran 1220 bp dan 277 bp
BclI (+) Kira-kira berukuran 959 bp dan 538 b
tertentu dan adanya perubahan lingkungan juga dapat menyebabkan perbedaan virus tungro.
Kesimpulan
Virus tungro isolat Bogor dengan teknik PCR-RFLP tidak menunjukkan adanya keragaman gen protein selubung RTBV karena tidak terpotong oleh enzim
EcoRV, NsiI, PstI, maupun BclI. Virus tungro isolat Sidrap dan Mamuju mempunyai pola fragmen DNA gen protein selubung RTBV yang mirip karena semua isolat tidak terpotong oleh enzim BclI dan terpotong oleh enzim EcoRV,
NsiI, dan PstI. Ada perbedaan yang nyata antara RTBV isolat Bogor, Sidrap dan Mamuju.
Saran
Affandi, Adjid A, Wardoyo, Mulyono BXF, Soenarso, Syahruddin S. 1977.
Pedoman Bercocok Tanam Padi Palawija Sayur-Sayuran. Jakarta; Badan Pegendali BIMAS.
Agrios GN. 1997. Plant Pathology. 4th ED. New York: academic press, inc. Agustina ER. 2007. Keragaman gen protein rice tungro bacilliform badnavirus
pada suatu luasan hamparan padi di Desa Ciherang, Kecamatan Darmaga, Kabupaten Bogor [Skripsi]. Bogor: Fakultas Pertanian, Institut Pertanian Bogor.
Arboleda M, Cruz SF, Azzam O. 1999. Preliminary analysis of genetic variation of RTBV in two provinces of the Philippines. Di dalam: Chancellor TCB, Azzam O, Heong KL editor. Rice Tungro Management. Los Banos: IRRI. hlm 11-16.
Arfianis. 2006. Diferensiasi empat isolat rice tungro bacilliform badnavirus
dengan PCR-RFLP [Skripsi]. Bogor: Fakultas Pertanian, Institut Pertanian Bogor.
Azzam O, Cabunagan RC, Chancellor TCB. 2000. Maintenance of virus infected rice plants. Di dalam: Method for Evaluating Resistance to Rice Tungro Disease. Discussion Paper. IRRI. hlm 14-16.
Azzam O, Chancellor TCB. 2002a. The biology, epidemiology, and management of rice tungro viruses in the Philippines. Plant Disease 86: 77-81.
Azzam O, Chancellor TCB. 2002b. The biology, epidemiology, and management of rice tungro disease in Asia. Plant Disease 86: 88-100.
Bengkulu Ekspress. 2010. Diserang tungro, panen turun 10 %. www.bengkuluekspress.com. [20 Mei 2010].
Burhanuddin. 2004. Penyebaran penyakit tungro di propinsi Papua. Prosiding Seminar Nasional Teknologi Pertanian. BPTP Papua. Puslitbang Sosial Ekonomi Pertanian. Badan litbang Pertanian. Deptan. Jayapura, 5-6 Oktober 2004. hlm 707-710.
Cabautan PQ, Hibino H. 1984. Detection of spherical and bacilliform virus particles in tungro-infected plants by leafhopper transmission. IRRN 9: 18−19.
Cabautan PQ, Cabunagan RC, Koganezawa H. 1995. Biological variants of rice tungro viruses in the Philippines. Phytopathology 85: 77-81.
ISOLAT SIDRAP, MAMUJU DAN BOGOR
LARA HIKMAHAYATI
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
LARA HIKMAHAYATI. Keragaman Gen Protein Selubung Rice Tungro Bacilliform Badnavirus Isolat Sidrap, Mamuju dan Bogor. Dibimbing oleh ENDANG NURHAYATI.
Tungro merupakan penyakit padi yang disebabkan oleh infeksi kompleks dua virus, rice tungro bacilliform badnavirus (RTBV) dan rice tungro spherical waikavirus (RTSV). Beberapa isolat virus tungro dilaporkan memiliki keragaman genetika pada gen protein selubung RTBV. Penelitian ini bertujuan untuk mempelajari keragaman gen protein selubung RTBV isolat Sidrap, Mamuju dan Bogor, dengan metode PCR-RFLP. Isolat virus tungro diperoleh dari dua daerah endemik tungro di Indonesia yaitu varietas TN 1 dari Sidrap (Sulawesi Selatan), varietas Ciherang dari Mamuju (Sulawesi Barat), serta Galur Harapan (GH) dari Bogor (Jawa Barat). Isolat RTBV Sulawesi diperoleh dari koleksi Loka Penelitian Penyakit Tungro, Lanrang, Sidrap, Sulawesi Selatan. Isolat RTBV Bogor diperoleh dari pertanaman padi milik petani di Ciputih, Kecamatan Darmaga, Kabupaten Bogor, Jawa Barat. Semua isolat dipelihara pada tanaman padi varietas rentan IR64 di rumah kaca dengan ditularkan menggunakan vektor Nephotettix virescens Distant (Hemiptera: Cicadellidae). Tanaman padi yang menunjukkan gejala penyakit tungro dipanen pada 10-15 hari setelah inokulasi. DNA total tanaman padi terinfeksi virus tungro diekstraksi dan gen protein selubung RTBV diamplifikasi menggunakan sepasang primer spesifik RTBV-2L dan RTBV-2R. Hasil amplifikasi dipotong dengan 4 enzim restriksi yaitu EcoRV, NsiI, PstIdan
BclI. Pemotongan amplikon hasil PCR menggunakan enzim restriksi pada isolat Bogor tidak menunjukkan adanya keragaman gen protein selubung RTBV karena tidak terpotong oleh keempat enzim tersebut. Sedangkan isolat Sidrap dan Mamuju mempunyai pola fragmen DNA gen protein selubung RTBV yang mirip karena semua isolat tidak terpotong oleh enzim BclI dan terpotong oleh enzim
EcoRV, Nsi1, dan PstI.
Latar Belakang
Penyakit tungro adalah salah satu penyakit penting pada tanaman padi di
Asia terutama Asia Selatan dan Asia Tenggara. Penyakit ini disebabkan oleh
infeksi dua virus,
rice tungro bacilliform badnavirus
(RTBV) dan
rice tungro
spherical waikavirus
(RTSV) (Hibino
et al
. 1978). Kedua virus itu ditularkan
secara semipersisten oleh vektor yang mempunyai karakter mobilitas yang tinggi
yaitu wereng hijau
Nephotettix virescens
Distant (Hibino dan Cabunagan 1986).
Luas serangan penyakit tungro cukup tinggi bila dibandingkan penyakit
lainnya yang terdapat pada pertanaman padi. Menurut Soetarto
et al.
(2001),
rata-rata luas serangan penyakit tungro setiap tahun mencapai 12.000 ha. Bahkan pada
saat terjadi ledakan, luas serangan di satu propinsi saja dapat menyamai atau
berkali lipat rata-rata luas serangan tahunan nasional. Di Sulawesi Selatan pada
MT 1972/1973 epidemi penyakit tungro mencapai 43.151 ha yang tersebar di
Kabupaten Pinrang, Sidrap, Luwu, dan Polmas (Halteren dan Sama 1973). Pada
akhir tahun 1995 di Surakarta, Jawa tengah ledakan penyakit tungro menyebabkan
sekitar 12.340 ha sawah puso (gagal panen), dan nilai kehilangan hasil akibat
penyakit tersebut diperkirakan setara dengan Rp 25 milyar. Keberadaan penyakit
tungro tersebut ditemukan pula di beberapa daerah di Jawa Barat seperti
Purwakarta, Cianjur, Sukabumi, dan Bogor (Puslitbangtan 1995). Pada tahun 2004
luas serangan penyakit tungro di Sulawesi Tengah mencapai 217 ha terjadi di
Donggala, Parigi, Banggai dan Tolitoli. Sebelumnya, pada musim tanam 2002
serangan virus tungro terjadi di Kabupaten Donggala, Tolitoli dan Parigi Moutong
(Negara
et al
. 2004).
Isolat virus tungro dari beberapa daerah endemis tungro di Indonesia
menunjukkan adanya variasi virulensi. Perbedaan virulensi tersebut dapat dilihat
dari variasi gen protein selubung
yang dapat diketahui melalui pengujian secara
molekuler menggunakan metode PCR-RFLP (Suprihanto 2005). Arfianis (2006)
juga melaporkan adanya keragaman pada gen protein selubung RTBV dari empat
Penggunaan varietas tahan dan galur harapan (GH) tahan tungro merupakan salah satu bentuk dari pengendalian penyakit ini. Varietas tahan penyakit tungro diklasifikasikan sebagai varietas yang tahan terhadap wereng hijau sebagai penular (vektor) patogen dan tahan terhadap virus yang merupakan patogen penyebab penyakit tungro (Widiarta dan Said 2007). Galur harapan merupakan galur-galur padi yang telah lolos dalam pengujian multilokasi yang kemudian dapat dijadikan sebagai varietas baru (Pakki et al. 2007).
Penelitian mengenai variasi virulensi virus tungro yang dapat diketahui melalui keragaman pada gen protein selubung RTBV, sangat bermanfaat sebagai sumber informasi bagi para peneliti dalam upaya pengendalian virus tungro. Selama ini penelitian mengenai keragaman pada gen protein selubung RTBV sebagian besar dilakukan di wilayah endemik tungro di Jawa Barat, sehingga informasi mengenai variasi virulensi spesifik di wilayah endemik tungro lainnya masih kurang. Selain itu, pengujian terhadap keragaman gen protein selubung RTBV pada galur harapan juga belum ada. Oleh karena itu, penelitian mengenai variasi virulensi virus tungro pada wilayah endemik lain selain Jawa Barat serta pada galur harapan perlu dilakukan.
Tujuan Penelitian
Penelitian ini dilakukan untuk mempelajari keragaman gen protein selubung RTBV Isolat Sidrap, Mamuju, dan Bogor dengan metode PCR-RFLP.
Manfaat Penelitian
Tanaman Padi
Padi merupakan tanaman pangan berupa rumput berumpun yang juga merupakan makanan pokok utama bagi lebih dari sepertiga penduduk dunia. Dari semua padi yang diproduksi dan dikonsumsi lebih dari 90% terpusat di Asia. Sebagai sumber pemberi tenaga, beras merupakan bahan makanan utama untuk ratusan juta umat manusia, terutama bagi umat manusia yang menduduki belahan timur dari benua ini. Oleh karenanya tidaklah mengherankan bahwa tanaman padi yang terluas terdapat di negara-negara Asia yang seluruh penduduknya sebagian besar memperoleh tenaganya dari beras sebagai sumbernya (Siregar 1980).
Tanaman padi diduga berasal dari negara India dan Cina kemudian menyebar ke negara-negara lain. Bukti sejarah memperlihatkan bahwa penanaman padi di Zhejiang, Cina sudah dimulai pada 3.000 tahun SM. Fosil butir padi dan gabah ditemukan di Hastinapur, Uttar Pradesh, India sekitar 100-800 SM. Tanaman padi di Indonesia berasal dari perantau-perantau Malaysia yang membawa tanaman padi sekitar tahun 1.500 SM (Siregar 1980).
Botani dan Morfologi Padi
Tanaman padi (Oryza sativa L.) termasuk golongan tumbuhan Gramineae yaitu tumbuhan yang ditandai dengan batang yang tersusun dari beberapa ruas. Tanaman ini juga termasuk golongan tanaman setahun/semusim (Affandi 1997). Bentuk batangnya bulat dan berongga, daunnya memanjang seperti pita yang berdiri pada ruas-ruas batang dan mempunyai sebuah malai yang terdapat pada ujung batang (Siregar 1980).
lihatkan percabangan dimana cabang yang terpendek menjadi lidah daun (ligula), dan bagian yang terpanjang dan terbesar menjadi kelopak. Di dekat lidah daun dan daun kelopak terdapat dua embel sebelah kiri dan kanan yang disebut auricle. Daun kelopak yang membalut ruas yang paling atas dari batang umumnya disebut daun bendera (flag-leaf). Tepat dimana daun pelepah teratas muncul ruas yang disebut bulir padi. Bulir terdiri dari ruas-ruas pendek. Tiap-tiap ruas sebelah kiri dan kanannya timbul cabang-cabang bulir, dan pada ujung tiap-tiap cabang bulir terdapat bunga padi. Bunga padi terdiri dari tangkai bunga, perhiasan bunga dan daun mahkota yang terdiri dari dua belahan yang tidak sama besarnya (Siregar 1980).
Taksonomi Padi
Padi (Oryza sativa L.) merupakan tumbuhan berbiji tunggal (monokotil), dengan urutan taksonomi (Siregar 1980):
Kingdom : Plantae Subkingdom : Tracheobionta Superdivisi : Spermatophyta Divisi : Magnoliophyta
Class : Liliopsida (Monocotyledons) Subclass : Commelinidae
Family : Poaceae Genus : Oryza Spesies : Oryza sativa L.
Varietas Padi
tahan terhadap tumbangnya pertanaman, mutu beras, dan rasa nasi yang lebih enak (Siregar 1980).
Keunggulan dari varietas tersebut tidak bersifat kekal atau abadi. Predikat unggul yang diberikan pada suatu varietas yang tertentu hanya berlaku sebelum ditemukannya varietas baru yang dapat menandingi varietas terdahuluya dalam sifat-sifatnya (Siregar 1980).
Selain varietas unggul tersebut, padi memiliki beberapa varietas yang resisten, moderat (intermediet), dan rentan terhadap hama dan penyakit. Varietas resisten dan moderat terhadap penyakit tungro diklasifikasikan tahan terhadap wereng hijau sebagai penular (vektor) patogen dan tahan terhadap virus yang merupakan penyebab penyakit tungro.
[image:48.595.94.519.521.738.2]Varietas tahan wereng hijau dikelompokkan berdasarkan sumber gen tetua tahannya menjadi golongan T1, T2, T3, dan T4 (Tabel 1). Anjuran penggunaan varietas tahan wereng hijau adalah (1) di Jawa Barat dapat ditanam varietas tahan golongan T1, T2, dan T4, (2) Jawa Tengah dapat menanam semua golongan varietas tahan, (3) Yogyakarta dianjurkan menanam varietas tahan dari golongan T2 dan T4, (4) Jawa Timur dan Bali hanya dianjurkan menanam varietas tahan golongan T4, dan (5) NTB dianjurkan menanam varietas tahan virus (Widiarta dan Said 2007).
Tabel 1 Varietas tahan wereng hijau untuk mengendalikan penyakit tungro
Golongan Varietas Gen Tahan
To IR5, Pelita, Atomita, Cisadane, Cikapundung, dan Lusi -
T1 IR20, IR30, IR26, IR46, Citarum, dan Serayu Glh1
T2 IR32, IR38, IR36, IR47, Semeru, Asahan, Ciliwung,
Krueng Aceh dan Bengawan Solo Glh 6
T3 IR50, IR48, IR54, IR52 dan IR64 Glh 5
T4 IR66, IR70, IR72, IR68, Barumun, dan Klara. Glh 4
Varietas tahan virus tungro yang telah dilepas antara lain Tukad Petanu, Tukad Unda, Tukad Balian, Kalimas, dan Bondoyudo yang sesuai di setiap daerah (Tabel 2). Varietas Tukad Petanu dapat dianjurkan untuk ditanam di seluruh daerah endemis, sedangkan Tukad Unda dianjurkan ditanam di NTB dan di Sulawesi Selatan. Petani di derah Bali dan Sulawesi Selatan dianjurkan menanam varietas Tukad Balian dan Bondoyudo. Varietas Kalimas dan Bondoyudo diketahui tahan terhadap penyakit tungro di Jawa Timur (Widiarta dan Said 2007).
Tabel 2 Pewilayahan kesesuaian varietas tahan virus tungro
Varietas
Kesesuaian Daerah
Jabar Jateng Jatim Bali Mataram Sulsel
Tukad Petanu + + + + + +
Tukad Unda - - - - + +
Tukad Balian - - + + - +
Bondoyudo - - + + - +
Kalimas - - + - - -
Sumber: Widiarta dan Said 2007 (+): Sesuai (tungro < 50%) (-): Tidak (tungro > 50%)
Dee-Geo-Woo-Gen dan Tsai-Yuan-Chung yang dirakit pada tahun 1949 oleh pemulia tanaman di Taichung District Agricultural Improvement Station, Taiwan. TN1 adalah varietas padi yang rentan terhadap virus tungro dan wereng hijau (Cabautan et al. 1995).
Penyakit Tungro
Arti Penting Penyakit Tungro
Tungro yang berarti ‘pertumbuhan terhambat’, untuk pertama kali ditemukan di Philiphina pada tahun 1963 dan merupakan penyakit yang sangat merugikan. Penyakit ini disebabkan oleh virus yang ditularkan wereng daun, terutama Nephotettix virescens Distant (Semangun 1991).
Di Indonesia, penyakit tungro mula-mula hanya terbatas penyebarannya di daerah tertentu seperti Sulawesi Selatan, Kalimantan Selatan, Nusa Tenggara Barat (NTB), dan Sulawesi Utara. Penyakit ini kemudian menyebar ke Jawa Timur, Jawa Tengah, dan Yogyakarta. Selanjutnya pada awal tahun 1970-an ledakan penyakit tungro dilaporkan terjadi di beberapa daerah sentra produksi padi di Indonesia. Ledakan penyakit tungro terjadi di Bali pada tahun 1980, yang meliputi Kabupaten Badung, Tabanan, dan Gianyar (Rachim 2000).
Daerah endemis penyakit tungro di Indonesia terpusat di daerah sentra produksi padi seperti Sulawesi Selatan, Jawa, Bali dan Nusa Tenggara Barat (Hasanuddin et al. 1995), serta propinsi Sulawesi Tenggara, Papua, dan Sulawesi Tengah terutama di daerah Parigi Moutong (Burhanuddin 2004). Menurut data Balai Informasi Pertanian Palu, luas serangan penyakit tungro di Sulawesi Tengah pada tahun 2004 mencapai 217 ha terjadi di Donggala, Parigi, Banggai dan Tolitoli. Sebelumnya, pada musim tanam 2002 serangan virus tungro terjadi di Kabupaten Donggala, Tolitoli dan Parigi Moutong (Negara et al. 2004).
dalam Agustina 2007). Di Indonesia, kerugian yang dirasakan oleh petani akibat serangan virus tungro pernah terjadi di Sulawesi Selatan tahun 1972, di Bali tahun 1980, dan di Surakarta tahun 1995, dengan kerugian ditaksir milyaran rupiah (Talanca et al. 2007). Pada musim tanam 2005/2006 lalu, virus tungro telah menyerang tanaman padi di NTB dan Manokwari dengan tingkat serangan sedang sampai berat. Bahkan pada musim tanam tahun ini pun produksi padi menurun hingga lebih dari 10% di Bengkulu (Bengkulu Express 2010). Diperkirakan kehilangan hasil akibat serangan virus tungro di seluruh Indonesia rata-rata 12.000 ha/tahun atau kerugiannya senilai Rp 48 miliar/tahun (asumsi harga gabah Rp 1.000/kg). Karenanya penyakit ini perlu diantisipasi, terutama di daerah endemis seperti Sulawesi Selatan, Sulawesi Tengah, NTB, Papua, Jawa Tengah, Jawa Timur, dan Bali (Puslitbangtan 2007).
Gejala Penyakit Tungro
Penyakit tungro disebabkan oleh dua jenis virus yaitu rice tungro bacilliform badnavirus (RTBV) dan rice tungro spherical waikavirus (RTSV). Tanaman padi yang terinfeksi kedua virus tersebut menjadi kerdil dan kelihatan belang serta perubahan warna kuning sampai oranye pada daun. Gejala tungro yang khas dapat disebabkan oleh RTBV dan gejala dapat diperkuat dengan kehadiran RTSV (Agrios 1997).
Gambar 1 Gejala serangan virus tungro pada daun tanaman padi. Warna daun yang menguning (orange), dimulai dari ujung daun dan berkembang kebagian lamina daun bawah ([email protected] ).
Tanaman yang terinfeksi virus tungro tumbuh kerdil, jumlah anakan sedikit, helaian daun dan pelepah daun memendek. Pada bagian bawah helaian daun muda terjepit oleh pelepah daun sehingga daunnya terpuntir atau menggulung sedikit. Malai pendek, gabah tidak terisi sempurna atau kebanyakan hampa dan terdapat bercak-bercak coklat yang menutupi malai. Infeksi virus tungro pada tanaman tua (umur di atas 50 hari setelah tanam) kurang berpengaruh terhadap produksi dan tanaman tidak menampakkan gejala serangan sampai panen (Ling 1972).
Penurunan jumlah anakan sangat tinggi bila infeksi terjadi pada stadium pertumbuhan sangat awal. Jumlah anakan tanaman padi dipengaruhi umur dan mungkin akan meningkat bila infeksi virus tungro setelah tanaman berumur lebih dari satu bulan. Namun jumlah anakan akan tetap sedikit jika selama infeksi terjadi, stadium petumbuhan terhambat (Ling 1972).
Penularan Penyakit Tungro
Virus tungro ditularkan secara semipersisten oleh wereng daun
Serangga penular virus tungro terutama adalah wereng hijau (N. virescens Distant, N. nigropictus (Stal), N. malayanus dan N. parvus). Serangga lain yang dapat juga sebagai penular virus tungro, namun kurang efisien adalah wereng loreng Recilia dorsalis (Motsch). Rentang efisiensi penularan virus oleh populasi
N. virescens antara 35 - 83%, dibandingkan dengan N. nigropictus yang rentang efisiensinya antara 0 - 27%. Spesies wereng hijau lainnya seperti N. malayanus
dan N. parvus memiliki kemampuan menularkan virus berturut-turut 40% dan 7% lebih rendah dari N. virescens (Deptan 1986).
Nephotettix sp. dikenal sebagai wereng hijau. Serangga ini menyerang bagian daun tanaman padi. Serangga dewasa tersebut berukuran 4-6 mm. Telurnya berbentuk bulat panjang atau lonjong berwarna terang (kuning pucat), berukuran 1,3 x 0,30 mm. Telur ini diletakkan berderet sebanyak 5-25 butir. Wereng daun betina mampu bertelur 200-300 butir yang diletakkan di dalam jaringan pelepah daun. Telur tersebut menetas setelah 4-8 hari dan membentuk serangga muda (nimfa). Nimfa ini mengalami 5 kali ganti kulit selama 16-18 hari, kemudian menjadi dewasa setelah 2-3 hari. Terdapat dua jenis Nephotettix sp yang dominan yaitu N. virescens dan N. nigropictus. Spesies N. virescens
berwarna hijau kekuningan dengan ujung kepala meruncing. N. virescens jantan mempunyai ukuran 4 mm dan N. virescens betina 6 mm, sedangkan nimfa
N. virescens berwarna hijau kekuningan hingga hijau tua. Spesies N. nigropictus
berwarna hijau tua dengan ujung kepalanya agak tumpul dan dilengkapi dengan
garis pita hitam yang jelas di atas bagian kepalanya. Serangga jantan
N. nigropictus berukuran 3,6 mm, sedangkan nimfa N. nigropictus berwarna kuning coklat hingga gelap (Deptan 1986).
Cabautan dan Hibino (1984) melaporkan bahwa wereng hijau dapat memindahkan RTSV dari tanaman padi yang hanya terinfeksi RTSV, tetapi tidak mampu memindahkan RTBV dari tanaman yang hanya terinfeksi RTBV. RTBV hanya dapat dipindahkan oleh wereng hijau dari tanaman yang telah terinfeksi RTSV. Dengan demikian RTBV merupakan virus dependent sedangkan RTSV berfungsi sebagai helper. Kedua partikel virus tersebut bersifat noncirculative, yaitu dalam tubuh vektor virus tidak dapat ditularkan dari imago ke telur maupun stadia perkembangan imago (Ling 1966). Disamping itu virus tungro juga tidak dapat ditularkan melalui biji, tanah, air dan secara mekanis (misal pergesekan antara bagian tanaman yang sakit dengan yang sehat). Nimfa wereng hijau juga dapat menularkan virus tungro, tetapi menjadi tidak infektif setelah ganti kulit (Widiarta 2005).
Fluktuasi kepadatan populasi vektor virus tungro sangat mempengaruhi keberadaan tanaman terinfeksi virus tungro bila sumber inokulum virus ini sudah ada di lapang. Persentase tanaman terinfeksi virus tungro yang tinggi pada musim hujan (Desember hingga April) bertepatan dengan kepadatan populasi wereng hijau yang tinggi pada periode yang sama. Sebaliknya pada musim kemarau (Mei sampai November), persentase tanaman terinfeksi virus tungro yang rendah bersamaan dengan kepadatan populasi wereng hijau yang lebih rendah daripada musim hujan (Widiarta 2005).
Virus Tungro
RTSV termasuk kedalam famili Sequiviridae genus Waikavirus. RTSV mempunyai genom poliadenil ssRNA, unipartit, terbungkus partikel isometrik dengan diameter 30 nm (Hibino et al. 1978). Genom RNA RTSV kira-kira 11 kb (kilo base) dan protein selubungnya terbentuk dari dua jenis molekul protein (Agrios 1997).
RTBV termasuk famili Caulimoviridae dan genus Badnavirus. Bentuk partikel RTBV adalah bacilliform dengan diameter 30-35 nm dan panjang kira-kira 100-300 nm yang bervariasi antar isolat (Hibino et al. 1978). Asam nukleat RTBV adalah DNA utas ganda dan bulat lebih kurang 8 kb. Asam nukleat tersebut mengandung dua daerah yang tidak bersambung yang merupakan hasil dari proses replikasi oleh reverse transcriptase dan empat open reading frames
Keragaman Gen Protein Selubung RTBV
Pendiagnosisan keragaman virus tungro dengan melihat gejala dan analisis RFLP telah dilakukan pada isolat dari Philippina (Azzam dan Chancellor 2002a) dan Indonesia (Suprihanto 2005). Dari diagnosis tersebut diperoleh pengetahuan bahwa virus tungro memiliki perbedaan pada setiap lokasi. Populasi virus tungro secara geografi dilaporkan hanya stabil pada periode waktu tertentu. Ini menunjukkan bahwa virus tungro memberikan respon yang berbeda terhadap perubahan lingkungan dan inang. Studi lingkungan menunjukkan bahwa pada satu lokasi virus tungro yang memiliki keragaman secara genetik dan biologi dapat hidup berdampingan (Azzam dan Chancellor 2002b).
Cabautan et al. (1995) melaporkan bahwa ada keragaman pada empat strain RTBV (G1, G2, Ic dan L) dari Philippina berdasarkan gejala yang berbeda pada varietas padi FK135 dan TN1. Uji RFLP terhadap genom empat strain RTBV di atas menunjukkan pola RFLP yang beragam. Suprihanto (2005) juga melakukan uji penularan virus tungro pada tanaman diferensial FK 135 dan TN1, dan uji PCR-RFLP terhadap delapan isolat RTBV yang diambil dari daerah endemis tungro di Indonesia. Berdasarkan gejala yang diamati dan pola PCR-RFLP diketahui bahwa delapan isolat RTBV berturut-turut menyebabkan gejala yang berbeda terutama pada warna daun dan keragaman pada gen protein selubungnya. Demikian juga Arfianis (2006) melakukan uji diferensiasi dengan PCR-RFLP empat isolat RTBV yang diambil dari daerah endemis tungro di Jawa Barat. Hasil yang diperoleh pun menunjukkan adanya keragaman pada gen protein selubung RTBV. Pengujian terhadap variasi genetik protein selubung RTBV pada tingkat luasan hamparan padi disuatu wilayah pada varietas juga dilaporkan oleh Agustina (2007). Hasil yang diperoleh yaitu terdapatnya keragaman pada gen protein selubung RTBV.
Polymerase Chain Reaction
Restriction Fragment Length Polymorphism ( PCR-RFLP)
B. Mullis, seorang peneliti di perusahaan CETUS Corporation. Metode PCR sangat sensitif karena dapat digunakan untuk melipatgandakan suatu molekul DNA. Metode ini juga sering digunakan untuk memisahkan gen-gen kanopi tunggal dari sekelompok sekuen genom. PCR mensyaratkan bagian tertentu sekuen DNA yang dilipatgandakan harus diketahui terlebih dahulu sebelum proses pelipatgandaan tersebut dilakukan. Sekuen yang diketahui tersebut penting untuk menyediakan primer, yaitu suatu sekuen oligonukleotida pendek yang berfungsi mengawali sintesis rantai DNA dalam reaksi berantai polimerase (Yuwono 2006 ). Proses PCR pada dasarnya terdiri atas tiga tahap reaksi dengan kondisi suhu yang berbeda secara berulang dalam beberapa siklus tertentu yaitu denaturasi, annealing (penempelan primer) dan ekstensi primer (sintesis DNA). Dengan reaksi amplifikasi DNA secara simultan, maka jumlah sasaran akhir telah dilipatgandakan secara eksponensial (Mc Pherson et al. 1992 dalam Mutaqin 2000). Proses sintesis inilah yang membuat sensitifitas teknik PCR semakin tinggi, karena dari jumlah molekul DNA yang sedikit dapat dikopi menjadi berlipat ganda (Takahashi et al. 1993).