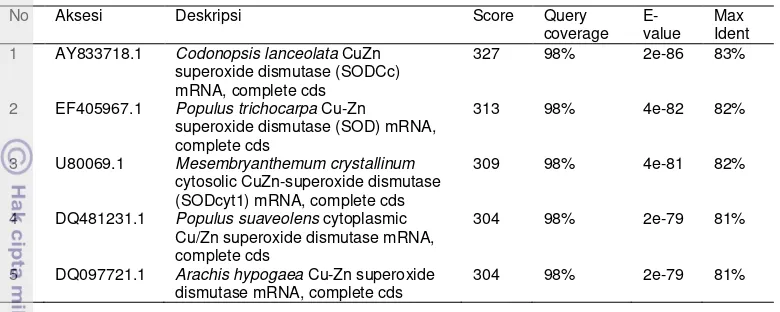
(CuZn-SOD) DARI Melastoma malabathricum L.
SALEHA HANNUM
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Dengan ini saya menyatakan bahwa disertasi berjudul “Isolasi, Pengklonan, dan Analisis Ekspresi Gen Penyandi Copper/Zinc Superoxide Dismutase (CuZn-SOD) dari Melastoma malabathricum L” adalah karya bersama saya dan pembimbing yang belum pernah diajukan dalam bentuk apapun
kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip
dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah
disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka.
Bogor, Januari 2012
SALEHA HANNUM. Isolation, Cloning, and Expression Analysis of Gene Coding Copper/Zinc Superoxide Dismutase (CuZn-SOD) from Melastoma malabathricum L. Supervised by SUHARSONO, UTUT WIDYASTUTI SUHARSONO, and ALEX HARTANA.
Melastoma malabathricum L. is an indigenous plant in tropical Southeast Asia with exceeding tolerance to aluminum (Al) stress in acid soils. Although increasing evidence suggests that Al-stress is accompanied by oxidative damages in plants, little has been described on the antioxidative components in this tolerant plant. Copper/zinc SOD (CuZn-SOD) is one of aluminum-induced genes. Analysis of the expression of genes induced by Al in M. malabathricum requires internal control genes. Actin is belong to housekeeping genes commonly used as an internal control. This study aimed to isolate and clone the cDNA fragment of MmACT encoding for actin of M. malabathricum, and isolate, clone and expression analysis of copper/zinc SOD (CuZn-SOD) from M. malabathricum L. Total RNA was isolated and used as template for cDNA synthesis by reverse transcription. Four of cDNAs clones were isolated, and were called MmACT1, MmACT2, MmACT3, and MmACT4. The size of MmACT1 and MmACT2 is 617 bp, whereas MmACT3 and MmACT4 is 735 bp. The sequence of MmACT cDNAs was registered in GenBank / EMBL / DDBJ database with accession numbers AB500686, AB500687, AB500688, and AB500689. Full length cDNA of MmCuZn-SOD had been successfully isolate by RACE. The size of MmCuZn-SOD is 824 bp that encoded a 152-amino acid polypeptide with a predicted pI value of 5.77, and showed high homology to other cytosolic CuZn-SODs from higher plants. Semi-quantitative RT-PCR showed that MmCuZn-SOD mRNA was highly expressed in the leaf tissues than stem and root. The Al treatment strongly induced the expression of MmCuZn-SOD both in the leaf and root tissues. These results suggested the fortification of the anti-oxidative defense system under Al stress in this acid soil-tolerant wild plant. Overexpression vectors was successfully constructed and each has been introduced into Agrobacterium tumefaciens LBA 4044 by triparental mating (TPM). N.benthamiana and Nicotiana tabacum transgenics had been obtained by mediated-A. tumefaciens. Analysis of segregation of T1 transgenic plants based on analysis of Khi-Quadrat (2) showed that the T1 transgenic plants accordance with 3:1 of Mendelian inheritance pattern. RNAi vector had been successfully constructed using GATEWAYTM cloning technology and it had been introduced into M. malabathricum L. mediated by Agrobacterium tumefaciens LBA 4404. Transgenic plants analyzes to Al stress showed that in the MS medium containing 1 mM Al (AlCl36H2O), the transgenic plants underwent growth suppression, whereas non-transgenic plants underwent growth normally. These results suggested that suppression of MmCuZn-SOD gene expression by RNAi to M. malabathricum L. caused the plant became sensitive to Al.
SALEHA HANNUM. Isolasi, Pengklonan, dan Analisis Ekspresi Gen Penyandi Copper/Zinc Superoxide Dismutase (CuZn-SOD) dari Melastoma malabathricum L. Dibimbing oleh SUHARSONO, UTUT WIDYASTUTI SUHARSONO, dan ALEX HARTANA.
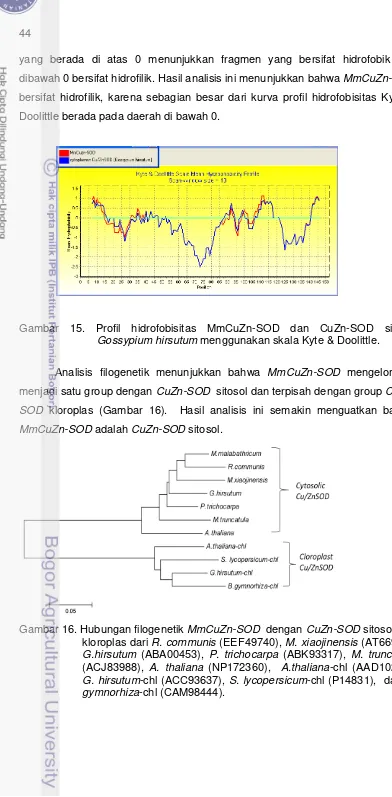
Melastoma malabathricum L. merupakan tanaman asli Asia Tenggara yang toleran terhadap cekaman aluminium (Al) pada tanah asam. Gen-gen yang ekspresinya diinduksi oleh Al diduga terlibat dalam sistem toleransi terhadap Al. Gen superoxside dismutase (SOD) merupakan salah satu gen yang ekspresinya diinduksi Al. SOD diduga memiliki peran dalam ketahanan M. malabathricum. Untuk mempelajari ekspresi gen secara kuantitatif pada M. malabathricum L, informasi housekeeping gene dari tumbuhan tersebut juga sangat dibutuhkan sebagai kontrol internal. Salah satu housekeeping gene yang populer dijadikan sebagai internal kontrol adalah gen aktin. Penelitian ini bertujuan mengisolasi dan mengklon fragmen gen penyandi aktin dari M. malabathricum L, selanjutnya melakukan isolasi, pengklonan, dan analisis ekspresi gen penyandi copper/zinc superoxide dismutase (CuZn-SOD) dari M. malabathricum L, dan mempelajari ekspresi gen MmCuZn-SOD melalui konstruksi ekspresi berlebih (overekspresi) di tanaman model Nicotiana benthamiana dan Nicotiana tabacum, dan pembungkaman gen MmCuZn-SOD melalui RNAi di M. malabathricum L. RNA total telah berhasil diisolasi dan dijadikan sebagai cetakan untuk sintesis cDNA total melalui transkripsi balik. Empat fragment cDNA yang menyandi aktin dari M. malabathricum telah berhasil diisolasi dan disisipkan ke dalam plasmid pGEM-T Easy. Keempat fragmen ini selanjutnya dinamakan fragmen MmACT1, MmACT2, MmACT3, dan MmACT4. Analisis urutan nukleotida menunjukkan bahwa fragmen MmACT1 dan MmACT2 berukuran 617 pb, dan fragmen MmACT3 dan MmACT4 berukuran 735 pb. Antar keempat fragmen cDNA MmACT ini memiliki kemiripan nukleotida sekitar 78%-99%, dan kemiripan asam amino sekitar 98% - 100%. Analisis hubungan filogenetik berdasarkan urutan asam amino menunjukkan bahwa pada ketidakmiripan 1% MmACT1, MmACT2, MmACT3 mengelompok dengan ACT5 Populus trichocarpha, sementara MmACT4 mengelompok dengan ACT9 P. trichocarpa dan ACT1 Gossypium hirsutum, dan kedua kelompok ini terpisah dengan aktin dari tumbuhan monokotil. Keempat fragmen MmACT ini telah didaftarkan di bank data GenBank/EMBL/DDBJ dengan nomor aksesi AB500686, AB500687, AB500688, and AB500689. Selanjutnya fragmen cDNA yang menyandi CuZn-SOD dari M. malabathricum juga telah berhasil diisolasi. Gen utuh MmCuZn-SOD yang berhasil diisolasi dengan metode RACE berukuran 824 pb, terdiri dari 459 pb open reading frame (ORF) yang menyandi 152 asam amino dengan prediksi nilai pI 5.77. Analisis filogenetik berdasarkan urutan asam amino menunjukkan bahwa MmCuZn-SOD memiliki kemiripan yang tinggi dengan CuZn-SOD sitosol tanaman tingkat tinggi. Berdasarkan analisis semi-kuantitatif RT-PCR menunjukkan bahwa MmCuZn-SOD terekspresi pada jaringan daun, batang dan akar. Perlakuan Al menginduksi ekspresi MmCuZn-SOD baik dalam jaringan daun maupun akar.
bahwa tanaman transgenik T1 yang diuji sesuai dengan pola pewarisan Mendel 3 : 1. Konstruksi pembungkaman gen MmCuZn-SOD juga telah berhasil dilakukan melalui RNAi. Vektor RNAi telah diintroduksikan ke tanaman M. malabathricum L. melalui Agrobacterium tumefaciens LBA4404 untuk mempelajari peranan gen MmCuZn-SOD dalam detoksifikasi cekaman Al. Uji toleransi tanaman transgenik terhadap cekaman Al menunjukkan bahwa pada media MS yang mengandung 1 mM Al (AlCl36H2O), tanaman transgenik mengalami hambatan pertumbuhan sampai mati, sementara non-transgenik tidak mengalami hambatan. Hal ini menunjukkan bahwa penghambatan ekspresi gen MmCuZn-SOD dengan RNAi pada tanaman M. malabathricum L. menyebabkan tanaman menjadi sensitive terhadap Al. Hal ini menjelaskan bahwa gen MmCuZN-SOD diduga berperan penting dalam detoksifikasi Al pada M. malabathricum L. Penelitian tentang isolasi, pengklonan, dan uji peranan gen merupakan penelitian yang masih jarang dilakukan. Saat ini isolasi fragmen gen penyandi aktin dari M. malabathricum L, isolasi, pengklonan, dan analisis ekspresi gen penyandi copper/zinc-superoxide dismutase (CuZn-SOD) dari M. malabathricum L, dan mempelajari ekspresi gen MmCuZn-SOD melalui konstruksi ekspresi berlebih (overexpression) di tanaman model Nicotiana benthamiana dan Nicotiana tabacum, dan pembungkaman gen MmCuZn-SOD melalui RNAi pada M. malabathricum L merupakan penelitian yang belum pernah dilakukan oleh peneliti lain di dunia. Oleh karena itu, keempat topik penelitian di atas adalah kebaruan (novelty) dalam penelitian ini.
©Hak Cipta milik IPB, Tahun 2012 Hak Cipta dilindungi Undang-Undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber.
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah.
b. Pengutipan tidak merugikan kepentingan yang wajar bagi IPB.
(CuZn-SOD) DARI Melastoma malabathricum L.
SALEHA HANNUM
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Biologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji Luar Komisi pada Ujian Tertutup, 19 Desember 2011: 1. Dr. Ir. Hajrial Aswidinnoor, M.Sc 2. Dr. Ir. Miftahudin, M.Si
Penguji Luar Komisi pada Ujian Terbuka 3 Januari 2012: 1. Dr. Ir. Satoto, M.S
Melastoma malabathricum L. Nama Mahasiswa : Saleha Hannum
Nomor Pokok : G361040041
Disetujui
Komisi Pembimbing
Prof.Dr. Ir. Suharsono, DEA Ketua
Dr. Ir. Utut W. Suharsono, M.Si Prof. Dr. Ir. Alex Hartana, M.Sc
Anggota Anggota
Diketahui
Ketua Program Studi Biologi
Dr. Ir. Dedy Duryadi Solihin, DEA
Dekan Sekolah Pascasarjana
Dr.Ir. Dahrul Syah, M.Sc. Agr.
Alhamdulillah, segala puji hanya untuk Allah SWT yang telah memberikan
kemudahan dan rahmat-Nya sehingga penulis dapat menyelesaikan penelitian
dan penulisan disertasi ini. Disertasi ini disusun berdasarkan hasil penelitian
yang dilakukan di Laboratorium Biotechnology Research Indonesia-The Netherland (BIORIN) Pusat Penelitian Sumberdaya Hayati dan Bioteknologi (PPSHB) IPB dan Laboratorium Plant Differentiation and Morphogenesis, Nara Institute Science and Technology (NAIST) Japan. Disertasi ini memuat hasil penelitian tentang isolasi, pengklonan, dan analisis ekspresi gen penyandi
copper/zinc superoxide dismutase dari tanaman Melastoma malabathricum L. Shalawat dan salam disampaikan kepada Rasulullah Muhammad SAW atas
keteladanannya.
Selama penelitian ini penulis telah banyak mendapat bantuan dan
bimbingan dari berbagai pihak. Pada kesempatan ini penulis menyampaikan
penghargaan yang tinggi dan ucapan terima kasih kepada Bapak Prof. Dr. Ir.
Suharsono, DEA selaku ketua komisi pembimbing, Ibu Dr. Ir. Utut Widyastuti
Suharsono, M.Si. dan Bapak Prof. Dr. Ir. Alex Hartana, M.Sc selaku anggota
komisi pembimbing atas segala bantuan, arahan, dan bimbingan yang telah
diberikan kepada penulis mulai dari tahap perencanaan, pelaksanaan, hingga
penulisan disertasi ini. Demikian juga penulis menyampaikan terimakasih dan
penghargaan yang tinggi kepada Prof. Dr. Akiho Yokota dan Dr. Kinya Akashi
yang telah memberikan arahan, bimbingan, dan fasilitas laboratorium di Nara Institute Sciences and Technology (NAIST) Japan. Kepada Tim Beasiswa Pendidikan Pascasarjana (BPPS), Rektor Universitas Sumatera Utara (USU)
Medan, Program Sandwich dan Hibah Pascasarjana dari Ditjen Dikti Depdiknas,
dan Hibah Kompetensi dengan judul:”Isolasi dan ekspresi gen dalam rangka perakitan tanaman yang toleran terhadap cekaman asam dan aluminium an.
Dr.Suharsono,DEA dengan nomor kontrak 039/HIKOM/DP2M/2008/ tanggal 13
agustus 2008, 219/SP2H/PP/DP2M/V/2009 tanggal 30 Mei 2009, dan 224/
SP2H/PP/DP2M/III/2010 tanggal 1 Maret 2010, terimakasih atas bantuannya
dalam menyediakan biaya pendidikan dan penelitian.
Terima kasih juga penulis sampaikan kepada Rektor, Dekan Fakultas
Pascasarjana (SPs) IPB, dan Ketua Program Studi Biologi SPs IPB, atas
kesempatan yang diberikan kepada penulis untuk mengikuti pendidikan di SPs
IPB Bogor. Kepada seluruh staf pengajar dan administrasi Sps IPB, penulis
menyampaikan banyak terimakasih atas ilmu dan kelancaran adminstrasi selama
penulis menjadi mahasiswa di SPs IPB. Penulis juga menyampaikan terimakasih
kepada staf PPSHB IPB atas bantuannya dalam kelancaran pelaksanaan
penelitian di laboratorium.
Penulis juga menyampaikan terimakasih kepada rekan-rekan mahasiswa
seperjuangan di Laboratorium BIORIN, yaitu Bu Srilis, Bu Yohana, Pak Ulung,
Bu Ratna, Pak Radit, Bu Ifa, Pak Asri, Muchdar, Anita, Nurul, Ophie, Delih,
Nikson, Ila, Iin, Lian, dan Mia, serta yang telah lulus, yaitu Pak Muzuni, Firdaus,
Agustina, Pak Hadi, Yasinta, Niken, Ulfa, Yassir, Jaya, Zahro, Lulu, Go To, Fajri,
Indah, dan Lita; rekan-rekan mahasiswa seperjuangan di Program Studi Biologi,
yaitu Bu Iin, Bu Dorly, Bu Nursahara, Bu Sri, dan Bu Ida atas dorongan dan
kerjasamanya. Terimakasih juga disampaikan kepada Pak Mulya, Mbak Pepi,
Pak Adi, Mbak Nia, Mbak Sarah, Pak Asep, Bu Dewi, Bu Ika, Bu Emi, Bu Eni,
dan Pak Iri atas bantuan dan kerjasamanya.
Ucapan terima kasih yang tulus ikhlas juga penulis sampaikan kepada
kedua orang tua penulis ayahanda dan ibunda (almarhum) yang senantiasa
mencurahkan cinta dan kasih sayangnya, dan selalu memanjatkan doa demi
kesuksesan penulis, serta dorongan moril sehingga penulis dapat menyelesaikan
pendidikan S3. Semoga Allah swt menyayangi mereka seperti menyayangi
penulis. Kepada kakak, abang, adek, dan keponakan penulis, terimakasih atas
segala perhatian, kasih sayang, pengorbanan, pengertian, dorongan moril, serta
doa yang diberikan kepada penulis selama ini.
Sebagai penutup, semoga karya ilmiah ini bermanfaat dan dapat
memberikan sumbangan bagi perkembangan ilmu pengetahuan di Indonesia.
Bogor, Januari 2012
Penulis dilahirkan di Pidoli Dolok pada tanggal 31 Agustus 1971, sebagai
anak ketiga dari enam orang bersaudara pasangan ayahanda H. Muhammad
Saleh Nasution, BA dan ibunda Hj. Masdewa Harahap (alm).
Penulis menyelesaikan pendidikan dasar di SDN 12 Kota Padang
Sidimpuan pada tahun 1984, pendidikan menengah dan lanjut di Madrasah
Tsanawiyah dan Madarasah Aliyah Pondok Pesantren K.H.Ahmad Dahlan
Sipirok, Tapanuli Selatan pada tahun 1987 dan 1990. Penulis melanjutkan
pendidikan di Universitas Andalas, Padang jurusan Biologi dan lulus 1996. Tahun
1998 melalui program DUE (Dikti) penulis melanjutkan pendidikan Pascasarjana
pada program studi Biologi di Institut Pertanian Bogor dan lulus 2001. Tahun
2000 penulis diangkat menjadi staf pengajar di Universitas Sumatera Utara,
Medan sampai sekarang. Tahun 2004 penulis mendapat kesempatan untuk
melanjutkan ke jenjang doktor pada program studi Biologi di Institut Pertanian
Bogor. Tahun 2008 penulis mendapat kesempatan melaksanakan penelitian di
Nara Institute Science and Technology (NAIST), Jepang selama 6 bulan melalui
Latar Belakang
Lahan asam merupakan salah satu lingkungan yang membatasi produksi
pertanian. Sekitar 50% lebih dari lahan pertanian di dunia adalah lahan asam
(Bot et al. 2000). Sementara Indonesia mempunyai sekitar 47,5 juta ha tanah Podsolik Merah Kuning (CSAR 1997) yang bersifat asam dengan kelarutan
aluminium (Al) yang tinggi.
Foy (1988) menjelaskan bahwa kemasaman tanah adalah faktor
cekaman terbesar yang mempengaruhi pertumbuhan tanaman dengan
keberadaan Al merupakan faktor pembatas pertumbuhan pada tanah asam.
Pada pH dibawah 5, Al menjadi terionisasi yang sangat beracun bagi tanaman
(Kinraide & Parker 1990; Kochian et al. 2004; Meriga et al. 2010 ). Aluminium telah bersifat racun bagi tanaman meskipun konsentrasinya masih sangat
rendah. Walaupun demikian Al yang membentuk ikatan dengan ligand adalah
tidak beracun bagi tanaman seperti aluminium silikat. Bentuk Al yang bersifat
toksik bagi tanaman adalah ion Al3+ yang dominan pada kondisi asam (Matsumoto 2000; Kochian et al. 2004). Kelarutan Al yang tinggi di dalam tanah
sangat merugikan tanaman karena dapat menghambat pertumbuhan akar
(Delhaize & Ryan 1995; Rout et al. 2001; Kochian et al. 2005).
Tumbuhan yang hidup di tanah asam umumnya adalah
tumbuh-tumbuhan yang dapat beradaptasi dengan lingkungan tersebut. Salah satu jenis
tumbuhan yang banyak dijumpai pada Tanah Podsolik Merah Kuning adalah
Melastoma (Tjitrosoedirdjo 1991). Melastoma merupakan anggota famili Melastomataceae, tersebar di daerah Tropik Asia dan seluruh Indonesia sebagai
gulma. Salah satu spesiesnya adalah Melastoma malabathricum L. yang banyak
dijumpai di lahan asam. Pertumbuhan akar M. malabathricum L. tidak mengalami gangguan pada pH 4.0 dan terganggu pada pH 3.0 (Muhaemin
2008). Tumbuhan ini dapat tumbuh dengan baik pada tanah asam yang
tumbuhan lain tidak tumbuh sehingga dapat dijadikan sebagai tumbuhan
M.affine D.Don. mampu mengakumulasi 8.81 g Al kg daun tua setelah 2 bulan perlakuan (Mutiasari 2008).
Respon toleransi tanaman terhadap Al sangat berkaitan dengan gen-gen
yang terlibat di dalamnya. Isolasi gen diperlukan untuk mengetahui regulasi
ekspresinya, sehingga dapat dimanfaatkan untuk perbaikan genetika tanaman
(Suharsono 2002). Gen-gen yang ekspresinya diinduksi oleh Al diduga terlibat
dalam sistem toleransi terhadap Al. Sedikitnya ada 30 gen yang ekspresinya
diinduksi Al (Darko et al. 2004), beberapa diantaranya adalah gen-gen yang mengkode enzim antioksidan, seperti glutathione-S-transferase (GST), ascorbate peroxidase (APX), catalase (Cat), dan superoxide dismutase (SOD) (Richards et al. 1998; Ezaki et al. 2000; Boscolo et al. 2003; Meriga et al. 2010).
Superoxide dismutase (SOD) termasuk kelompok metalloenzim yang
mampu menetralkan radikal bebas dengan mengkatalisis perubahan radikal
superoksida menjadi molekul O2 dan H2O2. Superoksida merupakan salah satu radikal bebas turunan (derivate) oksigen reaktif (ROS) yang umumnya terdapat
dalam sel tanaman sebagai hasil samping dari proses metabolisme normal.
Akumulasi radikal bebas dapat menyebabkan kerusakan makromolekul sel dan
bahkan kematian sel (Bowler et al. 1992; Scandalios 1993). Konsentrasi radikal
bebas di dalam sel tanaman dapat meningkat ketika tanaman merespon
cekaman biotik dan abiotik. Namun tanaman juga memiliki sistem pertahanan
yang dapat mencegah peningkatan radikal bebas ini dengan adanya enzim
antioksidan seperti SOD. Ada tiga tipe enzim SOD sesuai dengan logam yang
berperan sebagai kofaktor pada sisi aktif enzimnya, yaitu copper/zinc
(CuZn-SOD), besi (Fe-(CuZn-SOD), dan mangan (Mn-SOD). CuZn-SOD ditemukan di sitosol,
kloroplas, dan peroksisom; Mn-SOD di mitokhondria; dan Fe-SOD di kloroplas
(
Bannister et al. 1987; Bowler et al. 1992; Bueno et al. 1995; Kliebenstein et al.1998).
Aktivitas SOD telah dilaporkan meningkat dengan adanya cekaman
abiotik, seperti cahaya tinggi dan suhu rendah (Allen et al. 1997), sulfur dioksida
(Tseng et al. 2008), kekeringan (Fu & Huang 2001; Bian & Jian; 2009), dan aluminium (Cakmak & Horst 1991; Basu et al. 2001; Du et al. 2010). Brassica
napus yang tahan Al mengekspresikan gen SOD secara berlebih (Basu et al. 2001). Aktivitas enzim SOD juga meningkat dengan cekaman Al pada kedelai
(Meriga et al. 2010). Cekaman Al dapat menimbulkan cekaman oksidatif dengan
terbentuknya oksigen radikal (ROS) (Panda et al. 2003).
Gen penyandi SOD telah diisolasi dari jagung (Cannon et al. 1987), tomat
(Perl-Treves et al. 1988), sawi (Liu et al. 1998), dan Nicotiana plumbaginifolia (Alscher et al. 2002). Pada Arabidopsis thaliana, telah diisolasi tiga gen CuZnSOD, yaitu CSD1, CSD2, dan CSD3. CSD1 dan CSD2 terekspresi pada akar, daun, dan batang, dan masing-masing target proteinnya di sitosol dan
kloroplas. Sementara target potein CSD3 di peroksisom karena ujung
karboksilnya mengandung tripeptida Ala-Lys-Leu yang merupakan targeting signal peroksisom (Kliebenstein et al. 1998).
Pada M. malabathricum L., beberapa gen yang diduga terlibat dalam cekaman asam dan Al telah diisolasi seperti multidrug resistance associated protein (MRP) (Suharsono et al. 2008), metallothionein type 2 (Mt2) (Suharsono et al. 2009), H+-ATPase membran plasma (Muzuni et al. 2010), dan sitrat sintase (Mushofa 2011). Namun sampai saat ini belum ada informasi tentang gen
CuZn-SOD pada M. malabathricum L. yang diduga juga terlibat dalam toleransi terhadap cekaman asam dan Al. Untuk mempelajari ekspresi gen secara
kuantitatif pada M. malabathricum L, informasi housekeeping gene dari tumbuhan tersebut sangat dibutuhkan sebagai kontrol internal. Sampai saat ini
informasi tersebut belum ada. Menurut Maroufi et al. (2010) aktin termasuk salah satu kontrol internal yang paling stabil.
Beberapa metode dapat digunakan untuk mengisolasi gen antara lain
melalui penapisan terhadap pustaka genom dan pustaka cDNA, serta RT-PCR
(Reverse Transcription-Polymerase Chain Reaction). Selain itu ada juga yang
menggunakan metode RACE (Rapid Amplification of cDNA Ends) untuk
memperoleh gen utuh, yaitu sintesis cDNA dengan menggunakan mRNA
sebagai cetakan dan sekuen internal yang sudah diketahui urutan nukleotidanya
serta adapter pada ujung 3’ atau 5’ sebagai primer. Metode RACE telah digunakan untuk isolasi gen utuh (full length) mannose-binding lectin dari umbi Zephyranthes grandiflora (Kai et al. 2006), gen penyandi Gibberellin 20-Oxidase dari Helianthus annuus (Carzoli et al. 2008), dan gen penyandi H+-ATPase membran plasma dari M. malabathricum L. (Muzuni et al. 2010).
Peranan suatu gen dalam tanaman dapat dipelajari minimal dengan
pendekatan dua arah, yaitu meningkatkan ekspresi gen dengan mengkonstruksi
menurunkan ekspresi atau pembungkaman gen antara lain dengan
mengkonstruksi vektor RNAi (RNA interference). RNAi menyebabkan mRNA terdegradasi sehingga gen menjadi tidak berfungsi. Teknologi RNAi telah
digunakan untuk mempelajari peranan gen penyandi H+-ATPase pada M. malabathricum L. (Muzuni et al. 2011), membungkam gen OsGEN-L pada padi (Moritoh et al. 2005), dan menurunkan ekspresi gen ornithine decarboxylase pada tanaman Nicotiana tabacum L. (DeBoer et al. 2011). Pada penelitian ini,
telah dilakukan isolasi dan pengklonan fragmen gen penyandi aktin dari M. malabathricum L, selanjutnya dilakukan isolasi, pengklonan, dan analisis ekspresi gen penyandi copper/zinc-superoxide dismutase (CuZn-SOD) dari M. malabathricum L. Ekspresi gen dilakukan di tanaman model Nicotiana benthamiana dan Nicotiana tabacum, dan pembungkaman gen dilakukan di M. malabathricum L.
Tujuan Penelitian
Penelitian ini bertujuan :
1. Mengisolasi dan mengklon fragmen gen penyandi aktin dari Melastoma malabathricum L.
2. Mengisolasi, mengklon, dan menganalisis ekspresi gen penyandi copper/zinc
superoxide dismutase (CuZn-SOD) dari M.malabathricum L. (MmCuZn-SOD). 3. Mengkonstruksi vektor ekspresi gen MmCuZn-SOD untuk ekspresi berlebih
pada tanaman Nicotiana benthamiana dan Nicotiana tabacum.
4. Mempelajari peranan gen MmCuZn-SOD melalui konstruksi vektor RNAi untuk
pembungkaman gen pada tanaman M. malabathricum L.
Strategi Penelitian
Strategi yang digunakan untuk mencapai tujuan dengan membagi
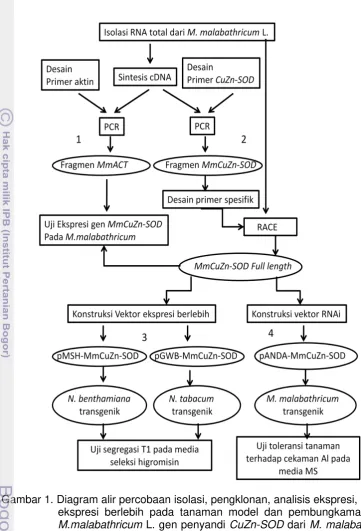
penelitian menjadi 4 aspek kajian (Gambar 1), yaitu:
1. Mengisolasi dan mengklon gen penyandi aktin dari M. malabathricum L.
2. Mengisolasi, mengklon, dan menganalisis ekspresi gen MmCuZn-SOD pada M. malabathricum L. yang diberi perlakuan cekaman abiotik.
3. Mengkonstruksi vektor ekspresi untuk ekspresi berlebih gen MmCuZnSOD pada tanaman model N. benthamiana dan N. tabacum.
BAB II
TINJAUAN PUSTAKA
Melastoma malabathricum L.
Melastoma adalah salah satu genus dari famili Melastomataceae yang termasuk dalam ordo Myrtales. Genus ini terdiri dari 22 spesies yang tersebar di
Asia Tenggara, India, Cina Selatan, Jepang dan Australia Utara (Meyer 2001).
Melastoma malabathricum L. merupakan salah satu spesies tumbuhan berkayu yang tumbuh di tanah asam dengan keasaman yang sangat tinggi dan miskin
unsur hara (seperti N dan P), dan tersebar di daerah tropis Asia, Australia, dan
Polynesia (Osaki et al. 1997). Di Indonesia tumbuhan ini juga banyak ditemukan
tumbuh di tanah asam, khususnya tanah Podsolik Merah Kuning dengan
kelarutan aluminium (Al) yang tinggi yang menjadi faktor pembatas bagi
pertumbuhan tanaman. Ketahanan tanaman M. malabathricum L. pada tanah asam berhubungan dengan kemampuannya mengakumulasi Al di daun tanpa
menyebabkan gejala keracunan dan bahkan Al memacu pertumbuhannya
(Watanabe et al. 1998), sehingga tanaman ini dikenal juga sebagai akumulator Al
(Watanabe & Osaki 2002; Watanabe et al. 2005).
Menurut Watanabe et al. (1997), M. malabathricum L. yang ditumbuhkan
pada media cair dengan cekaman Al sebesar 0.5 mM selama 6 minggu mampu
mengakumulasi Al lebih dari 10 g kg-1 pada daunnya, sementara pada daun muda tanaman ini mengakumulasi Al lebih dari 7 g kg-1. Analisis akumulasi Al pada M. affine D.Don (sinonim dari M. malabathricum L.) yang mendapat cekaman 3.2 mM Al pada pH 4 di dalam media cair menunjukkan bahwa M. affine D.Don mampu mengakumulasi 8.81 g Al kg-1 daun tua setelah 2 bulan perlakuan (Mutiasari 2008). Tanaman akumulator Al yang lain seperti tanaman
teh (Camellia sinensis (L.) Kuntze) dapat mengakumulasi Al hingga 30 g kg-1 pada daun tua dan 0.6 g kg-1 pada daun muda (Matsumoto et al. 1976). Daun Hydrangea macrophylla dapat mengakumulasi Al sebesar 3 g kg-1 (Ma et al. 1997).
Tanaman mencegah toksisitas Al melalui mekanisme ekslusi dan
mekanisme toleransi internal. Pada mekanisme ekslusi, Al didetoksifikasi
dengan mengeksudasi senyawa-senyawa organik yang dapat mengikat Al.
diserap tanaman. Bentuk-bentuk kimia Al pada tanaman toleran telah
diidentifikasi menggunakan spektroskopi Nuclear Magnetic Resonance (NMR). Menurut Watanabe et al. (1998) bentuk Al pada daun M. malabathricum L. adalah Al3+, Al-oksalat, Al(oksalat)2, dan Al-(oksalat)3. Sementara Watanabe & Osaki (2001) menjelaskan bahwa Al diangkut dari akar ke pucuk tanaman M. malabathricum dalam bentuk kompleks Al-sitrat. Jadi, asam organik dengan berat molekul kecil memainkan peranan penting dalam detoksifikasi internal pada
jaringan tanaman dan transport Al dari akar ke pucuk melalui pembentukan
kompleks asam organik. Hal yang sama juga ditemukan pada tanaman
akumulator Al lainnya seperti buckwheat (Ma & Hiradate 2000) dan teh (Morita et
al. 2008).
Respon toleransi tanaman terhadap Al sangat berkaitan dengan gen-gen
yang terlibat di dalamnya. Isolasi gen diperlukan untuk mengetahui regulasi
ekspresinya, sehingga dapat dimanfaatkan untuk perbaikan genetika tanaman
(Suharsono 2002). Gen-gen yang ekspresinya diinduksi oleh Al diduga terlibat
dalam sistem toleransi terhadap Al. Sedikitnya ada 30 gen yang ekspresinya
diinduksi Al yang telah dilaporkan (Darko et al. 2004). Beberapa diantaranya adalah gen-gen yang mengkode enzim antioksidan, seperti glutathione-S-transferase (GST), ascorbate peroxidase (APX), catalase (Cat), dan superoxide dismutase (SOD) (Richards et al. 1998; Ezaki et al. 2000; Boscolo et al. 2003; Meriga et al. 2010). Pada M. malabathricum L., gen yang diduga terlibat dalam toleransi tanaman terhadap cekaman asam dan Al telah diisolasi, yaitu multidrug
resistance associated protein (MRP) (Suharsono et al. 2008), metallothionein type 2 (Mt2) (Suharsono et al. 2009), H+-ATPase membran plasma (Muzuni et al. 2010), dan sitrat sintase (Mushofa 2011).
Toksisitas Aluminium
Secara normal, aluminium (Al) berada dalam bentuk oksida dan kompleks
aluminosilikat yang tidak larut dan tidak toksik. Pada pH netral, Al membentuk
kompleks dengan ion hidroksida yang tidak larut, sedangkan pada pH asam, Al
berada dalam bentuk Al3+ yang merupakan bentuk Al yang paling toksik (Kinraide & Parker 1990; Kinraide et al. 1994; Matsumoto 2000). Pada larutan dengan pH
yang lebih rendah dari 5.0, ion Al berada dalam bentuk oktahedral heksahidrat,
berkurang, Al(H2O)63+ mengalami deprotonasi menjadi Al(OH)2+ dan Al(OH)2+. Pada larutan netral menyebabkan Al(OH)3 mengendap dan larut kembali pada larutan basa dengan membentuk formasi tetrahedral, Al(OH)4- (Delhaize & Ryan, 1995; Marschner, 1995).
Keracunan Al merupakan salah satu kendala dalam produksi tanaman
pada tanah asam. Pada kondisi tersebut umumnya ketersediaan hara dan
kemampuan tanaman untuk menyerap hara sangat terbatas. Dari beberapa
percobaan diketahui bahwa penyerapan P, Ca, Mg, dan K oleh tanaman
berkurang secara nyata. Pada tanaman barley yang ditanam pada media yang
mengandung Al, kandungan Ca2+ dan K+ hanya setengahnya jika dibandingkan dengan kontrol (Matsumoto et al.1988). Al biasanya meningkatkan kandungan P
pada akar dan menurunkan kandungan P pada pucuk (Liang et al. 2001; Quartin
et al. 2001). Hal ini berhubungan dengan bentuk kompleks antara P dan Al pada akar yang menghambat transportasi P ke pucuk.
Aluminium dapat mengikat anion anorganik, seperti sulfat, fosfat, fluor,
dan silikat membentuk suatu kompleks yang mempunyai afinitas tinggi terhadap
oksigen atau air (Hodson & Evans 1995). Interaksi antara Al dengan anion
tersebut berpotensi untuk meningkatkan pH perakaran sekaligus dapat membuat
rancu pengaruh toksisitas Al dengan defisiensi unsur tertentu (seperti fosfat)
karena terbentuknya kompleks Al-fosfat (baik di larutan tanah maupun di dalam
sel) yang tidak tersedia bagi tanaman. Kemampuan tanaman untuk dapat
memanfaatkan kandungan P yang rendah secara efisien selalu dihubungkan
dengan sifat toleransi tanaman terhadap cekaman Al. Kation trivalen Al3+ menghambat transport Ca2+ secara efektif ke dalam akar, protoplasma dan membran vesikel. Hasil studi pada lipid bilayer menunjukkan bahwa Al dapat memblok Ca2+ dan saluran K+ (Ryan et al. 1997; Jones et al. 1998). Pada akar barley, perlakuan Al menurunkan kandungan Ca pada membran hingga 50% dan
menyebabkan penurunan aktivitas H+-ATPase dalam menghidrolisis ATP (Matsumoto & Yamaya 1988).
Pengaruh Aluminium pada Tanaman
Kelebihan konsentrasi Al dalam larutan tanah pada umumnya berakibat
buruk terhadap pertumbuhan tanaman, kecuali beberapa tanaman seperti teh
mineral tanah asam disebabkan dua hal yang sangat berkaitan, yaitu efek
langsung dengan menghambat penyerapan hara secara langsung, dan efek tidak
langsung dengan menghambat pertumbuhan sehingga secara tidak langsung
menghambat penyerapan hara (Marschner 1995).
Beberapa hasil penelitian menunjukkan bahwa target utama keracunan Al
adalah jaringan akar tanaman (Delhaize & Ryan 1995; Kochian et al. 2004). Penelitian pada gandum (Triticum aestivum) menggunakan kultivar yang sensitif
Al (Neepawa) dan toleran Al (PT741) menunjukkan bahwa setelah 3 hari
ditumbuhkan pada media yang mengandung berbagai konsentrasi Al, terlihat
penurunan pertambahan panjang akar pada kultivar sensitif sebanyak 57% pada
konsentrasi Al 25 µM, tetapi pada kultivar resisten Al belum berpengaruh (Basu
et al. 1994).
Aluminium banyak ditemukan pada inti dan dinding sel pada tanaman
yang sensitif. Pada dinding sel, penghambatan terjadi karena Al menggantikan
kedudukan Ca2+ pada lamella tengah. Aluminium berikatan dengan molekul pektin dinding sel atau komponen dinding sel yang bermuatan negatif pada
sel-sel epidermis dan korteks akar (Delhaize et al. 1993; Marienfeld et al. 2000; Schmohl & Horst 2000; Schmohl et al. 2000; Rout et al. 2001; Kochian et al. 2005). Gugus karboksil bebas pada molekul pektin yang terdemetilasi mengikat
ion Al toksik. Menurut Schmohl et al. (2000), perlakuan enzim pectin methylesterase (PME) pada suspensi sel Zea mays menurunkan resistensi terhadap Al, sehingga overekspresi PME pada tanaman kentang transgenik lebih
sensitif terhadap Al daripada non transgenik. Hal ini menunjukkan bahwa
matriks pektin pada apoplas sel-sel apikal akar berperanan penting dalam
memfasilitasi sinyal stress pada sitoskeleton sel-sel tersebut. Akumulasi Al yang
tinggi dalam apoplas akar merupakan karakteristik sensitifitas Al (Rincon &
Gonzales 1992; Schmohl & Horst 2000). Ikatan Al dengan gugus karboksil akan
menimbulkan ikatan yang kuat sehingga sel tidak dapat membesar (Marschner
1995). Pada inti sel, Al berasosiasi dengan DNA sehingga menghentikan proses
pembelahan sel pada meristem apikal (Matsumoto 1991; Rout et al. 2001). Aluminium dalam bentuk polimer memiliki muatan positif yang besar serta
memiliki banyak situs pengikatan. Polimer Al ini dapat mengikat fosfat yang ada
1991). Sedangkan menurut Silva et al. (2000) bahwa Al dapat terakumulasi dalam nukleus dengan konsentrasi yang rendah.
Pada membran sel, pengaruh Al lebih banyak disebabkan oleh adanya
perubahan atau kerusakan sifat permeabilitas. Pada membran sel akar barley, Al
ditemukan berasosiasi dengan gugus fosfolipid membran yang menyebabkan
kerusakan struktur membran atau perubahan dalam permeabilitas membran. Hal
ini menyebabkan penyerapan hara yang dikatalisis oleh pompa proton menjadi
terhambat (Matsumoto 1991; Rout et al. 2001). Ion Al yang bermuatan positif dapat berasosiasi dengan gugus fosfat dari ATP atau fosfolipid pada membran
yang akan mempengaruhi efektivitas transport proton (Kochian et al. 2004).
Toleransi Tanaman terhadap Cekaman Aluminium
Tanaman yang toleran terhadap Al tinggi mengembangkan mekanisme
toleransi melalui berbagai cara. Beberapa tanaman toleran Al mengeluarkan
asam-asam organik sebagai bahan pengkhelat Al pada daerah rizosfer.
Beberapa jenis tanaman diketahui mengeluarkan eksudat berupa asam sitrat,
seperti yang terjadi pada Phaseolus vulgaris (Miyasaka et al. 1991) dan kacang
kedelai (Yang et al. 2000). Pada tanaman gandum yang toleran Al mengeluarkan asam malat dari ujung akarnya (Delhaize et al. 1993). Tanaman
talas mengeluarkan eksudat asam oksalat (Ma & Miyasaka, 1998).
Taylor (1991) menyatakan bahwa resistensi Al dimediasi oleh protein
membran yang secara aktif mengeluarkan Al, sebagai enzim yang terlibat dalam
sintesis atau pengeluaran ligan kelator, atau enzim yang bertanggungjawab
terhadap sintesis komponen seluler yang mempunyai sifat mengubah resistensi
Al. Sopandie et al. (2003), juga mendapatkan tanaman kedelai yang toleran terhadap Al mengekspresikan suatu protein pada daerah meristem akar.
Beberapa tanaman dapat bertindak sebagai tanaman pengumpul
(akumulator) Al, karena dapat menyerap Al dan mengakumulasinya dalam
jaringan tanaman. Watanabe et al. (1998) melaporkan bahwa tanaman M. malabathricum L. mampu mengakumulasi Al dalam jaringan mesofil daun
maupun dalam jaringan penyusun akar, terutama pada jaringan epidermis dan
endodermis. Tanaman teh (Camellia sinensis L.) dapat mengakumulasi Al pada
mesofil daunnya dapat mengandung 19.000 mg Al kg-1 bobot kering daun (Gonzalez-Santana et al. 2011).
Gen-Gen yang Berhubungan dengan Toleransi Aluminium
Taylor (1991) mengungkapkan bahwa respon toleransi tanaman terhadap
Al sangat berkaitan dengan gen yang terlibat di dalamnya. Beberapa
gen-gen telah diketahui baik secara langsung maupun tidak langsung mengen-gendalikan
toleransi terhadap Al. Aniol (1995) menunjukkan bahwa pada lengan panjang
kromosom 2D tanaman gandum terdapat faktor genetik yang mencegah
akumulasi Al pada meristem apikal akar. Delhaize et al. (1993) menyatakan bahwa toleransi Al pada gandum dikendalikan oleh gen dominan Alt yang mengendalikan ekspresi malat ketika tanaman tersebut mengalami cekaman Al.
Hal yang sama juga dilaporkan Sasaki et al. (2004), bahwa gen ALMT1 yang terlibat dalam eksudasi malat dapat meningkatkan toleransi terhadap Al pada sel
tembakau. Eksudasi berbagai asam organik seperti malat, sitrat, dan oksalat
terjadi pada tanaman yang diberi cekaman Al (Delhaize et al. 1993; Kidd et al. 2001; Kochian et al. 2005). Pembungkaman gen penyandi H+-ATPase membran plasma melalui RNAi pada tanaman M. malabathricum L. transgenik menunjukkan kepekaan yang lebih tinggi terhadap cekaman 3.2 mM Al dan pH 4
dibandingkan tanaman non transgenik. Hal ini menunjukkan bahwa gen ini
berperan dalam toleransi M.malabathricum L. terhadap cekaman Al (Muzuni 2011).
Delhaize et al. (2004) melaporkan bahwa ekspresi berlebih Al-inducible malate transporter (ALMT) meningkatkan toleransi Al pada tanaman Hordeum vulgare. Overekspresi SbMATE yang menyandi putative citrate transporter dapat meningkatkan toleransi terhadap cekaman Al pada Arabidopsis dan gandum (Magalhaes et al. 2007; Liu et al. 2009). Gen ALS3 yang menyandi ABC transporter- like protein juga diperlukan dalam toleransi Al pada Arabidopsis dan
dapat berperan untuk mendistribusikan akumulasi Al dari jaringan yang sensitif
untuk melindungi pertumbuhan akar dari keracunan Al (Larsen et al. 2005).
Cekaman Al dapat menyebabkan pembentukan cekaman oksidatif.
Salah satu mekanisme keracunan Al adalah terjadinya peroksidasi lipid yang
oksidatif, sehingga meningkatkan aktivitas beberapa enzim cekaman oksidatif
(Ricards et al. 1998; Cakmak & Horst 1991; Foyer & Noctor 2005).
Richards et al. (1998) telah berhasil mengisolasi gen-gen cekaman oksidatif dari Arabidopsis thaliana yang ekspresinya terinduksi oleh cekaman Al,
yaitu gen-gen penyandi metallothionein-like protein, glutathione-s-transferase (GST), peroksidase, dan superoxide dismutase (SOD). Hal yang sama juga dilakukan oleh Ezaki et al. (1995) yang berhasil mengisolasi gen-gen tembakau
yang menyandikan GST, Peroxidase (PER), dan GDP Dissociation Proteinase Inhibitors (GDI) yang diinduksi oleh cekaman Al. Pada tanaman kedelai, Anwar et al. (2000) berhasil mengklon fragmen cDNA dari gen-gen kedelai yang toleran terhadap cekaman Al, antara lain gmali1 (Glycine max aluminum induced), gmali14, gmali49, dan gmali50, masing-masing menyandikan H+-ATPase membran plasma, protein histon H3, NADH-dehidrogenase dan Auxin-induced
protein. Semua gen tersebut di atas terekspresi untuk mempertahankan diri dari
cekaman lingkungan. Pada Melastoma affine (sinonim M. malabathricum), Suharsono et al. (2009) telah berhasil mengisolasi gen metallothionein type 2 (MaMt2) yang ekspresinya diinduksi oleh cekaman Al (Trisnaningrum 2009).
Superoxide Dismutase (SOD)
Superoxide dismutase (SOD) termasuk kelompok metalloenzim yang mampu menetralkan radikal bebas dengan mengkatalisis dismutase radikal
superoksida menjadi molekul O2 dan H2O2 (Bowler et al. 1992; Fridovich 1995; Tseng et al. 2008). Pada kondisi normal, radikal superoksida yang merupakan
derivat reactive oxygen species (ROS) terdapat di dalam sel tanaman sebagai produk sampingan dari proses metabolisme. Akumulasi ROS dapat
menyebabkan kerusakan berbagai fungsi seluler seperti kerusakan DNA,
protein, dan peroksidasi lipid, sehingga enzim ini sangat penting sebagai
antioksidan untuk pertahanan pada hampir semua sel yang terpapar oksigen
(Bowler et al. 1992; Fridovich 1995; Dong et al. 2009). Akumulasi ROS dapat disebabkan oleh berbagai cekaman lingkungan (Allen et al. 1997: Foyer & Noctor
2005).
SOD merupakan enzim antioksidan yang berada pada garis depan
sebagai pertahanan terhadap radikal superoksida (Fridovich 1995). Ada tiga tipe
enzimnya, yaitu copper-zinc (CuZn-SOD), besi (Fe-SOD), dan mangan
(Mn-SOD). CuZn-SOD ditemukan di sitosol, kloroplas, dan peroksisom, Mn-SOD di
mitokhondria dan Fe-SOD di kloroplas (Bannister et al. 1987; Bowler et al. 1992; Bueno et al. 1995; Kliebenstein et al. 1998; Dong et al. 2009). Pemberian logam
Mn, Cu, Zn, atau Fe ke dalam media kultur dapat meningkatkan ekspresi dan
aktivitas SOD. Shi et al. (2005) melaporkan bahwa aktivitas SOD, terutama aktivitas Mn-SOD meningkat seiring dengan peningkatan Mn pada tanaman
mentimun, sedangkan menurut Fernando and Miguel (2000) aktivitas SOD tidak
dipengaruhi oleh konsentrasi Mn pada padi. Pada Arabidopsis thaliana yang ditumbuhkan pada media dengan konsentrasi Cu2+ yang tinggi mampu meningkatkan aktivitas total SOD (Maria et al. 2007).
Aktivitas SOD telah dilaporkan meningkat dengan adanya cekaman
abiotik, seperti cahaya tinggi (Allen et al. 1997) sulfur dioksida (Tseng et al. 2007;
Tseng et al. 2008), ozon (Pitcher and Zilinskas 1996), kekeringan (Mittler & Zilinskas 1994; Fu & Huang 2001; Bian & Jian; 2009), suhu rendah
(Hernandez-Nistal et al. 2002; Lee & Lee 2000; Gao et al. 2009), garam ( Sreenivasulu et al.
2000), dan aluminium (Basu et al. 2001; Darko et al. 2004; Du et al. 2010; Meriga
et al. 2010). Tang et al. (2006) dan Lim et al (2007) melaporkan ekspresi gen CuZnSOD yang meningkat terhadap berbagai cekaman lingkungan pada tanaman kentang transgenik.
Basu et al. (2001) melaporkan bahwa Brassica napus yang tahan Al mengekspresikan gen SOD secara berlebih. Aktivitas enzim SOD juga meningkat dengan cekaman Al pada kedelai ( Cakmak & Horst 1991; Du et al. 2010), gandum (Darko et al. 2004), dan padi (Meriga et al. 2010). Menurut Du et
al. (2010) perlakuan Al meningkatkan aktivitas SOD pada akar dan kalus dari 2 genotipe kedelai (Al-tolerant PI 416937 (PI) dan Al-sensitive Young). Aktivitas
SOD pada kedua genotipe tersebut berbeda dalam merespon cekaman Al dan
bergantung pada konsentrasi Al dan lama perlakuan. Aktivitas SOD pada akar
yang tahan (PI) lebih tinggi dibandingkan akar yang rentan (Young) pada lama
perlakuan cekaman Al 36 atau 48 jam. Hal ini menunjukkan bahwa peningkatan
aktivitas enzim antioksidan merupakan salah satu mekanisme toleransi Al. Pada
akar barley, SOD juga terlibat dalam mekanisme detoksifikasi Al pada dosis
Peixoto et al. (1999) melaporkan bahwa aktivitas SOD pada akar gandum
yang resisten Al lebih tinggi dibandingkan gandum yang tahan Al pada kondisi
tanpa cekaman Al, namun dengan perlakuan cekaman Al, aktivitas SOD
meningkat lebih tinggi pada kultivar gandum yang tahan dibandingkan yang
resisten. Tingginya peningkatan aktivitas SOD pada gandum yang tahan dengan
perlakuan Al kemungkinan adalah hasil dari peningkatan konsentrasi H2O2. Salah satu mekanisme toksisitas Al adalah menyebabkan peroksidasi lipid
(Gutteridge et al. 1985; Cakmak & Horst 1991).
Gen penyandi SOD telah diisolasi dari jagung (Cannon et al. 1987), tomat
(Perl-Treves et al. 1988), sawi (Liu et al. 1998), dan Nicotiana plumbaginifolia (Alscher et al. 2002). Pada Arabidopsis thaliana, telah diisolasi tiga gen CuZnSOD, yaitu CSD1, CSD2, dan CSD3. CSD1 dan CSD2 terekspresi pada akar, daun, dan batang, dan masing-masing target proteinnya di sitosol dan
kloroplas. Sementara target potein CSD3 di peroksisom karena ujung
karboksilnya mengandung tripeptida Ala-Lys-Leu yang merupakan targeting signal peroksisom (Kliebenstein et al. 1998).
Pengklonan DNA dengan Rekombinasi Situs Spesifik
Teknologi pengklonan DNA sangat penting dalam bidang biologi,
terutama genetika dan biologi molekular. Pengklonan DNA dibutuhkan untuk
analisis fungsional gen dan ekspresi gen. Saat ini telah berkembang teknik
pengklonan DNA dengan menggunakan prinsip rekombinasi situs spesifik yang
lebih efisien dibandingkan dengan teknik pengklonan yang menggunakan enzim
restriksi (Hartley et al. 2000).
Pengklonan sistem Gateway merupakan salah satu metode pengklonan dengan rekombinasi situs spesifik secara in vitro yang ditemukan dan
dikomersialisasikan oleh Invitrogen sejak akhir 1990-an. Metode Gateway cloning adalah metode biologi molekuler untuk mentransfer fragmen DNA antar plasmid dengan efisien menggunakan situs rekombinasi, yaitu situs " gateway att", dan dua enzim campuran, yang disebut LR Clonase dan BP Clonase. Gateway cloning secara efektif telah menggantikan enzim restriksi dan ligase. Sistem ini memerlukan awal penyisipan fragmen DNA ke plasmid dengan dua
sekuens rekombinasi pengapit yang disebut "att L1" dan "att L2", untuk
Nakagawa et al. 2007). Metode ini sangat efisien, karena keberhasilannya lebih
dari 90% (Patton 2000; Freuler et al. 2008).
Secara umum, teknologi Gateway melibatkan proses dua-langkah. Gen target pertama diklon ke entry vector melalui suatu reaksi yang disebut reaksi BP
dengan enzim BP clonase, dan menghasilkan entry clone. Ketika membuat entry
clone, maka perlu untuk mengubah urutan ujung gen target agar kompatibel dengan situs rekombinasi Gateway (situs pengenalan rekombinase), namun tidak melibatkan enzim restriksi selama proses pengklonan secara keseluruhan.
Selanjutnya, gen target yang ada di dalam entry vector (entry clone) disubklon ke
destination vector (plasmid biner) melalui reaksi LR menggunakan enzim LR clonase. Jadi, hanya dengan sekali membuat entry clone, maka entry clone dapat digunakan ke berbagai plasmid biner sesuai dengan tujuan hanya dengan
reaksi LR, dan ini merupakan salah satu keuntungan dari teknologi Gateway (Invitrogen Co. 2003 ; Katzen 2007; Xu & Li 2008).
Salah satu entry vector yang tersedia saat ini adalah pENTR™/D-TOPO® kloning kit (Invitrogen), dengan menambahkan 4 nukleotida (CACC) pada primer
5'-PCR (forward) untuk amplifikasi gen, produk PCR yang berujung rata sudah
terarah untuk diklon ke vektor TOPO untuk menghasilkan clone entry (Gambar 2). Dengan demikian, teknologi TOPO dengan mudah menghasilkan klon entry (Xu and Li 2008). DNA sasaran yang yang telah tersisipi di klon entry akan lebih
mudah masuk ke vektor biner dengan hanya menggunakan reaksi LR (Hartley et
al. 2000; Patton 2000).
Tekhnologi gateway ini telah diaplikasikan untuk konstruksi vektor dalam
analisis ekpresi berlebih (overexpression) (Karimi et al 2002; Curtis & Grossniklaus 2003; Earley et al. 2006), antisense (Karimi et al. 2002), RNAi (Muzuni 2011), analisis promoter (Curtis & Grossniklaus 2003; Earley et al. 2006; Karimi et al. 2007), analisis ekspresi gen induksi (inducible gene) (Joube`s
et al. 2004; Brand et al. 2006; de Schutter et al. 2007), dan analisis ekspresi beberapa gen (multisite) (Karimi et al. 2005).
RNA interference (RNAi)
RNA interference (RNAi) merupakan potongan kecil RNA yang dapat menginduksi penghancuran mRNA tertentu sebelum dapat menyandi protein di
dalam sitoplasma. Prinsip dasarnya adalah masuknya double-stranded RNA (dsRNA) ke dalam sitoplasma akan membungkam ekspresi suatu gen di tingkat
post-transkripsi (Fire et al. 1998; Kalantidis et al. 2008). Pada awalnya, proses gangguan (interference) menggunakan RNA tidak berhasil, karena para peneliti
mengunakan dsRNA dengan panjang lebih dari 30 nukleotida. Hal ini
menyebabkan supresi dari gen yang tidak seharusnya terbungkam (non-specific
suppression gene). Pada perkembangannya, penggunaan dsRNA dengan nukleotida yang lebih pendek, 21-23 nukleotida, berhasil membungkam ekspresi
gen yang dikehendaki pada sel mamalia, yang dikenal dengan small interfering RNA (siRNA) (Fire et al. 1998; Waterhouse et al. 2001; Hannon 2002; Pickford & Cogoni 2003)
Penghambatan ekspresi gen dengan memasukkan dsRNA ini,
sebenarnya ditemukan secara tidak sengaja oleh Napoli et al (1990) ketika bermaksud meningkatkan ekspresi warna bunga Petunia. Namun hasil yang
mereka peroleh dengan memasukkan dsRNA yang komplementer dengan gen
yang berperan dalam biosintesia warna bunga tidak seperti yang mereka
harapkan. Ekspresi warna bunga yang diharapkan adalah menjadi ungu tua
sebagaimana umumnya warna bunga Petunia, namun mereka mendapatkan
sebaliknya, yaitu bunga Petunia yang berwarna ungu keputih-putihan
Penemuan ini merangsang berbagai kelompok peneliti mengikutinya dengan
tujuan untuk mempelajari efek tertekannya (suppression) ekspresi gen akibat
Mekanisme dasar RNAi dalam membungkam gen berhasil dijelaskan Fire
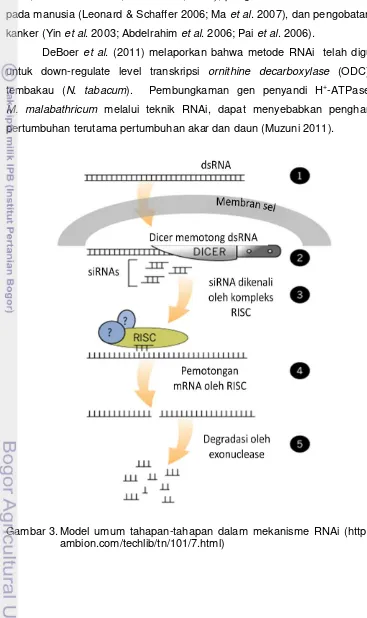
et al. (1998) dan Montgomery & Fire (1998) dengan menggunakan Caenorhabditis elegans. Mekanisme ini terdiri dari beberapa proses (Gambar 3): 1. Rantai dsRNA masuk kedalam sitoplasma sel (baik dalam bentuk alami
ataupun sintetis) dan akan langsung dikenali oleh enzim dicer (RNAse tipe
III). Enzim ini akan memotong rantai dsRNA menjadi rantai yang pendek-pendek (21 pb, termasuk 2 nukleotida dengan ujung 3’ di kedua ujungnya). 2. Dicer-dicer tersebut bersama co-factor lainnya akan sangat aktif
memotong-motong dsRNA sehingga akan terdapat banyak
potongan-potongan kecil dsRNA, yang disebut dengan small interfering RNA (siRNA) yang masih memiliki rantai ganda.
3. Selanjutnya siRNA akan dikenali dan ditangkap oleh kompleks
protein yang mengandung ribonuklease (ribonuclease-containing
multi-protein complex) atau diistilahkan dengan RNA-Induced Silencing Complex (RISC). RISC mengandung enzim Argonaut, pada tahap ini siRNA akan terdenaturasi menjadi utas tunggal yang akan mengaktifkan
RISC.
4. RISC yang aktif akan segera mencari mRNA hasil transkripsi yang keluar
dari inti sel, dan siRNA di dalam RISC akan dengan tepat mengenali target
dan mengikat pasangan basa komplemennya di mRNA.
5. Setiap RISC mengandung aktifitas enzim endonuclease (Argonaut
subunit) yang bertugas memotong target mRNA menjadi bagian-bagian
kecil, sehingga tidak dapat ditranslasi menjadi protein. Pada hewan,
mRNA yang terpotong ini akan teridentifikasi oleh sel sebagai mRNA yang
rusak (aberrant mRNA) dan langsung terdegradasi secara alami dengan
mekanisme endogenous. Sementara pada tumbuhan, potongan-potongan
mRNA ini selain terdegradasi secara alami oleh metabolisme sel, dapat
juga menjadi cetakan yang akan teridentifikasi oleh enzim RNA-dependent
RNA polymerase (RdRp) untuk melakukan polimerisasi dan menjadikan mRNA yang awalnya berutas tunggal menjadi dsRNA. Selanjutnya
dsRNA ini akan kembali teridentifikasi oleh dicer dan seterusnya
berulang-ulang.
Teknologi RNAi ini telah digunakan untuk mempelajari fungsi suatu gen
gen fungsional untuk mengidentifikasi target terapi pada hewan (Soutchek et al.
2004; Shen et al. 2005; Raoul et al; 2005), pengobatan infeksi virus berbahaya pada manusia (Leonard & Schaffer 2006; Ma et al. 2007), dan pengobatan terapi
kanker (Yin et al. 2003; Abdelrahim et al. 2006; Pai et al. 2006).
DeBoer et al. (2011) melaporkan bahwa metode RNAi telah digunakan
untuk down-regulate level transkripsi ornithine decarboxylase (ODC) pada tembakau (N. tabacum). Pembungkaman gen penyandi H+-ATPase pada M. malabathricum melalui teknik RNAi, dapat menyebabkan penghambatan pertumbuhan terutama pertumbuhan akar dan daun (Muzuni 2011).
Teknik Transformasi
Metode transformasi pada tanaman dapat dilakukan menggunakan
partikel bombardemen dan menggunakan bakteri Agrobacterium tumefaciens (Dai et al. 2001). Teknik transformasi dengan partikel bombardemen adalah mentransfer gen yang dioperasikan secara fisik menggunakan partikel yang
dibungkus DNA langsung ke sel atau jaringan tanaman (Klein et al. 1992;Kikkert
et al. 2004). Teknik ini secara ekonomi lebih mahal dibandingkan dengan menggunakan bakteri A. tumefaciens. Teknik transformasi menggunakan A. tumefaciens mampu mentransfer gen kedalam eksplan tanaman dan mempunyai regenerasi tinggi (Hiei & Komari 2008).
Kemampuan bakteri A. tumefaciens mentransformasi sel tanaman berhubungan dengan adanya T-DNA yang dapat berintegrasi ke dalam genom
tanaman. T-DNA adalah suatu bagian pada tumor inducing (Ti) plasmid yang
terdapat di dalam sel A. tumefaciens. Ti-plasmid berukuran sekitar 200-800 kb dan T-region (T-DNA)nya sendiri berukuran sekitar 10% nya (10-30 kb). T-region
ini dibatasi oleh dua sekuen pembatas (border) yaitu right border (RB) dan left border (LB) yang mengapit T-region. Bagian lain dari Ti-plasmid yang tidak kalah pentingnya adalah vir-region yang mengandung sejumlah gen-gen virulen (virA,
virB, virC, virD, virE, virF,virG dan virH) yang berfungsi didalam proses transfer T-DNA ke dalam sel tanaman (Zambryski et al. 1983: Gelvin 2000; Karami et al.
2009).
Interaksi antara A. tumefaciens dengan sel tanaman didahului dengan penginderaan (sensing) A. tumefaciens terhadap sel yang luka. Mekanisme ini terjadi secara kimiawi dimana sel tanaman yang luka menghasilkan suatu
metabolit yang berperan sebagai isyarat bagi A. tumefaciens. Metabolit tersebut dapat berupa senyawa gula, asam, asam amino atau senyawa fenol (Tinland
1996; Karami et al. 2009). Adanya isyarat tersebut maka A. tumefaciens akan bergerak aktif menuju ke sel sasaran. Gerakan yang bersifat kemotaksis ini
dipandu oleh senyawa yang disekresikan oleh sel tanaman rentan yang luka.
dalam sintesis berbagai suatu senyawa glukan, yaitu chvA, chvB, dan exoC
(Sheng & Citovsky 1996; Tinland 1996; Gelvin 2000).
Tahap selanjutnya adalah induksi faktor virulensi (vir) yang akan
mengatur proses pemotongan dan transfer T-DNA kel sel tanaman. Beberapa
metabolit yang disekresi oleh tanaman, akan menginduksi faktor virulensi.
Metabolit tersebut adalah asetosiringon, hidroksi asetosiringone, koniferil alkohol
dan etil firulat (Gelvin 2000; Kumar et al. 2006; Sarangi et al. 2011). Proses transfer T-DNA diwali dengan dideteksinya senyawa fenol dari sel tanaman yang
luka oleh A. tumefaciens. Hal ini menyebabkan terjadinya proses aktivasi ekspresi gen virulensi. Protein dari virA ini akan menginduksi fosforilasi produk dari virG yang selanjutnya mengaktifkan ekspresi berbagai vir lainnya. Protein yang dihasilkan oleh gen vir akan memotong T-DNA pada kedua sekuen berulang yang mengapit T-DNA yaitu batas kiri (LB) dan batas kanan (RB).
Transfer bersifat polar, yaitu bergerak dari batas kanan ke batas kiri. Jika batas
kanan dibuang maka transfer T-DNA tidak akan terjadi, tetapi jika batas kiri yang
dibuang maka transfer T-DNA tetap terjadi. Selama induksi, T-DNA dipotong
tepat pada utas bawah di dalam batas berulang oleh protein virD. Akibat
kejadian ini dihasilkan utas T yang berutas tunggal, kemudian utas tunggal ini
dipindahkan melalui membrane bakteri ke dalam sitoplasma sehingga
terintegrasi ke dalam genom tanaman inang (Sheng & Citovsky 1996; Gelvin
2000; Karami et al. 2009).
Hal penting dalam proses transformasi melalui A. tumefaciens ini adalah
transfer T-DNA ke inti tanaman target yang diinduksi oleh ekspresi gen-gen vir serta ekspresi gen-gen yang tertransformasi. Selain itu, integrasi T-DNA yang
membawa transgen ke dalam genom resipien, akan mengalami sedikit
pengaturan kembali secara intra dan intermolekuler, untuk memulihkan sistim
transkripsi dan translasi genom tanaman resipiennya. Transformasi melalui
A. tumefaciens lebih menjamin kestabilan genom tanaman resipien (Slamet-Loedin 1994; Sheng & Citovsky 1996; Gelvin 2003).
Transfer dengan sistim A. tumefaciens ini biasanya menggunakan vektor
biner (binary vector). Pada sistem ini digunakan dua plasmid, yaitu plasmid Ti
yang mengandung bagian virulen, dan plasmid kedua yang mengandung T-DNA
menemukan sisi pemotongan yang unik dengan enzim restriksi pada plasmid Ti
yang berukuran sangat besar (Slamet-Loedin 1994).
Agrobacterium adalah bakteri yang hidup bebas dalam tanah dan dapat menimbulkan penyakit pada tanaman yang terinfeksi. Pada budidaya pertanian
penyakit ini tergolong penting dan sebagian besar terjadi pada tanaman dikotil.
Menurut Miller & Bassler (2001) terdapat dua species Agrobacterium yang bersifat patogen yaitu A. tumefaciens sebagai penyebab penyakit tumor (crown gall) dan A. rhizogenes sebagai penyebab penyakit akar rambut (hairy root) pada berbagai tanaman dikotil yang peka.
Beberapa hal yang perlu diperhatikan dalam transfer gen yaitu konstruksi
gen dengan jenis promoter yang tepat dan dapat terekspresi pada jaringan target
yang diinginkan, kemampuan jaringan target untuk menerima gen asing, dan
kemampuan beregenerasi dari jaringan target. Keberhasilan transformasi
genetik tanaman ditandai dengan terintegrasinya gen yang diintroduksikan ke
dalam genom tanaman dan terekspresi serta tetap terpelihara dalam seluruh
proses pembelahan sel sampai regenerasi tanaman. Untuk pembuktian
terintegrasinya gen asing, umumnya digunakan gen penanda, misalnya gen gus yang menyandikan β-glucuronidase, yang ekspresinya ditunjukkan dengan timbulnya warna biru setelah uji histokimia (Jefferson et al. 1987). Untuk mengetahui integrasi gen kedalam tanaman juga dapat dilakukan dengan
menggunakan marka seleksi resistensi terhadap antibiotik (Hiei & Komari 2008).
Aktin
Aktin adalah protein yang sangat penting bagi sel eukariotik. Protein ini
berperan penting dalam membentuk jaringan yang memberikan dukungan
mekanik sel, menentukan bentuk sel, pergerakan sel, dan juga pembelahan sel
(Vantard & Blanchoin 2002; Blessing et al. 2004). Aktin juga penting dalam morphogenesis sel pada tumbuhan, sebagai komponen dinding sel, terlibat
dalam pertumbuhan rambut akar, sel trikom, tabung pollen, perpanjangan sel
dan apikal meristem (Gilliland et al. 2003). Kehilangan ekspresi gen ACT11 pada
jaringan vegetatif dapat menyebabkan perubahan morfologi organ tanaman
Arabidopsis (Kandasamy et al. 2002), dan pembungkaman gen aktin GhACT1 pada tanaman kapas menyebabkan terhambatnya pemanjangan serat kapas (Li
dengan protein lain seperti actin depolarizing factor (Chen et al. 2002) dan Rho family GTPase (Fu et al. 2002).
Aktin disandi oleh multigene family pada tanaman (Li et al. 2005; Feng et
al.2006). Pada Arabidopsis thaliana, famili gen aktinnya terdiri dari 10 gen yang berbeda, delapan diantaranya adalah gen fungsional dan dua gen adalah
pseudogen (McDowell et al. 1996). Li et al. (2005) telah mengisolasi 15 gen aktin (GhACT) dari kapas.
Pada Arabidopsis ada dua kelompok gen aktin yaitu kelompok vegetatif,
yang diekspresikan dominan pada daun, batang, akar, petal, dan sepal, dan
kelompok generatif yang diekspresikan secara kuat pada pollen, ovule, dan
jaringan embrionik (McDowell et al. 1996; Kandasamy et al. 1999). Gen-gen aktin Arabidopsis tersebut tersebar di kromosom 1, 2, 3, dan 5 (McKinney et al. 1996). Adapun pada kedelai dikenal dengan mu-aktin, kappa-aktin, dan
lambda-aktin (McLean et al. 1990).
Gen aktin termasuk housekeeping gene (Tu et al. 2007), yaitu gen yang memiliki tingkat eksperesi yang stabil di berbagai jaringan pada semua tahapan
perkembangan. Sifat gen yang seperti ini menjadikan gen aktin digunakan
sebagai kontrol internal pada analisis ekspresi, khususnya analisis ekspresi gen
dengan metode qRT-PCR (quantitative real time polymerase chain reaction)
yang merupakan metode analisis ekspresi yang berkembang saat ini (Bas et al.
2004). Gen aktin adalah salah satu gen yang paling sering digunakan sebagai
internal kontrol pada studi qRT-PCR (Bezier et al. 2002; Thomas et al. 2003; Chen et al. 2010).
Gen aktin telah digunakan sebagai kontrol ekspresi gen pada kentang
(Nicot et al. 2005), kedelai (Jan et al. 2008), gandum (Paolacci et al. 2009),dan padi (Zhang et al. 2009). Menurut Maroufi et al. (2010), aktin termasuk salah satu kontrol internal yang paling stabil pada uji ekspresi gen di daun dan akar
Cichorium intybus. Hal yang sama direkomendasikan untuk uji ekspresi gen pada padi (Ambavaram & Preira 2011) dan uji ekspresi gen perkembangan
bantalan buah pada kurma China (Sun et al. 2009). Bahkan Olbrich et al. (2008)
hanya merekomendasikan aktin sebagai kontrol internal untuk qRT-PCR pada
tanaman Fagus sylvatica L., setelah menguji kestabilan beberapa housekeeping
DARI Melastoma malabathricum L.
Abstrak
Melastoma malabathricum L. adalah tumbuhan akumulator aluminium (Al) yang dapat tumbuh baik pada tanah asam dengan kelarutan Al yang tinggi, sehingga dapat digunakan sebagai tanaman model untuk toleransi terhadap cekaman aluminium dan asam. Ekspresi gen-gen yang diinduksi oleh aluminium pada tanaman M. malabathricum memerlukan gen kontrol internal. Aktin adalah termasuk gen housekeeping yang biasa digunakan sebagai kontrol internal. Penelitian ini bertujuan mengisolasi dan mengklon fragmen cDNA MmACT yang menyandi aktin dari M. malabathricum. RNA total telah berhasil diisolasi dan dijadikan sebagai cetakan untuk sintesis cDNA total melalui transkripsi balik. Empat fragment cDNA yang menyandi aktin dari M. malabathricum
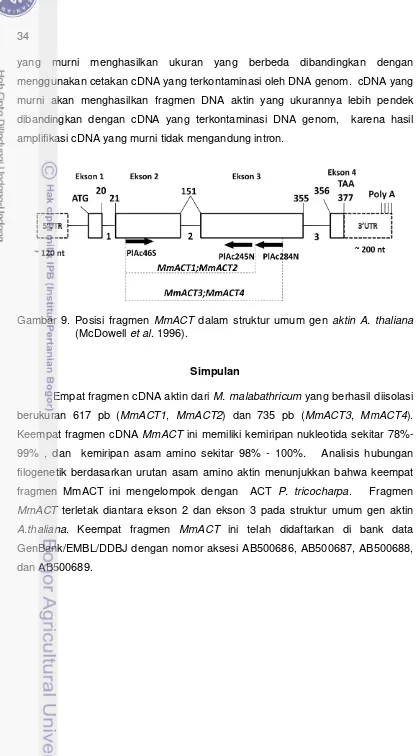
telah berhasil diisolasi dan disisipkan ke dalam plasmid pGEM-T Easy. Keempat fragmen ini selanjutnya dinamakan fragmen MmACT1, MmACT2, MmACT3, dan
MmACT4. Analisis urutan nukleotida menunjukkan bahwa fragmen MmACT1 dan
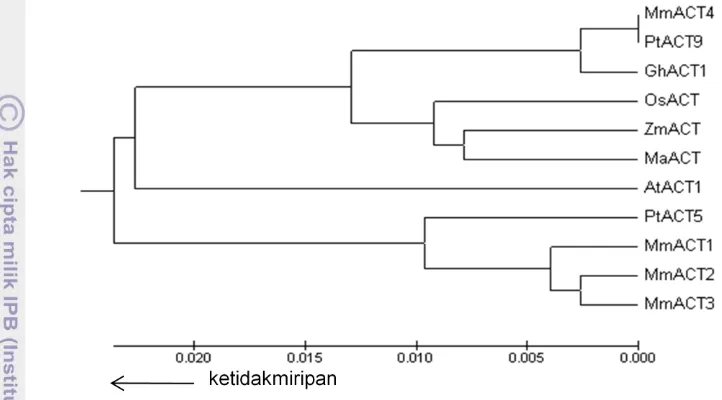
MmACT2 berukuran 617 pb, dan fragmen MmACT3 dan MmACT4 berukuran 735 pb. Antar keempat fragmen cDNA MmACT ini memiliki kemiripan nukleotida sekitar 78%-99%, dan kemiripan asam amino sekitar 98% - 100%. Analisis hubungan filogenetik berdasarkan urutan asam amino menunjukkan bahwa pada ketidakmiripan 1% MmACT1,
MmACT2, MmACT3 mengelompok dengan ACT5 Populus trichocarpha, sementara
MmACT4 mengelompok dengan ACT9 P. trichocarpa dan ACT1 Gossypium hirsutum, dan kedua kelompok ini terpisah dengan aktin dari tumbuhan monokotil. Keempat fragmen MmACT ini telah didaftarkan di bank data GenBank/EMBL/DDBJ dengan nomor aksesi AB500686, AB500687, AB500688, and AB500689.
Abstract
Melastoma malabathricum is accumulator plant of aluminum (Al) which can grow well in acidic soil with high Al solubility, thereby it can be used as a model plant for tolerance to aluminum and acid stresses. Analysis of the expression of genes induced by Al in M. malabathricum requires internal control genes. Actin is belong to housekeeping genes commonly used as an internal control. This study aimed to isolate and clone the cDNA fragment of MmACT encoding for actin of M. malabathricum. Total RNA was isolated and used as template for cDNA synthesis by reverse transcription. Four cDNA fragments encoding for actin from M. malabathricum have been isolated and inserted into plasmid pGEM-T Easy. Four of cDNAs clones were isolated, and were called MmACT1,
MmACT2, MmACT3, and MmACT4. The size of MmACT1 and MmACT2 is 617 bp, whereas MmACT3 and MmACT4 is 735 bp. Comparing among these four actin cDNA showed that they are about 78%-99% similarities based on nucleotide sequence and about 98%-100% similarities based on amino acid sequence. Analysis of phylogenetic relationship based on amino acid sequence showed that at 1% dissimilarity the
MmACT1, MmACT2, MmACT3 and the ACT5Populus trichocarpha are clustered in one group, while the MmACT4 is grouped with ACT9 P. trichocarpa and ACT1 Gossypium hirsutum, and these two groups are separated with actin group of monocotyl plants. The sequence of MmACT cDNAs was registered in GenBank / EMBL / DDBJ database with accession numbers AB500686, AB500687, AB500688, and AB500689.