SINTESIS PROTEIN MlKROBA DAN AKTIVITAS ENZIM SELULASE
RUMEN SAPI BALI
IN VITRO
DENGAN PENAMBAHAN
SABUN KALSIUM-MINY AK KEDELAI
SANTA LUSYA SIMANJUNTAK
DEPARTEMEN ILMU NUTRISI DAN TEKNOLOGI PAKAN
FAKULTASPETERNAKAN
INSTITUT PERT ANIAN BOGOR
BOGOR
2015
PERNY AT AAN MENGENAI SKRIPSI DAN SUMBER
INFORMASI SERTA PELIMP AHAN HAK CIPT A
Oengan ini saya menyatakan bahwa skripsi berjudul Sintesis Protein Mikroba dan Aktivitas Enzim Selulase Rumen Sapi Bali In Vitro dengan Penambahan Sabun Kalsium-Minyak Kedelai adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber inforrnasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telab disebutkan dalam teks dan dicantumkan dalam Oaftar Pustaka di bagian akhir skripsi ini.
Oengan ini saya melimpahkan hak cipta dari karya tulis saya kepada lnstitut Pertanian Bogor.
Bogor, April 2015
ABSTRAK
SANTA LUSYA SIMANJUNTAK. Sintesis Protein Mikroba dan Aktivitas Enzim Selulase Rumen Sapi Bali In Vitro dengan Penambahan Sabun Kalsium-Minyak Kedelai. Dibimbing oleh SRI SUHARTI dan DEWI APRI ASTUTI.
Minyak kedelai-sabun kalsium merupakan minyak kedelai yang diproteksi sabun sebagai sumber asam lemak tak jenuh ganda. Pemberian minyak kedelai-sabun kalsium dapat menghilangkan pengaruh negatif asam lemak tak jenuh pada mikroba rumen dan mengoptimaikan fermentasi rumen. Penelitian ini bertujuan untuk mengamati pengaruh penambahan minyak kedelai yang diproteksi sabun kalsium pada ransum terhadap konsentrasi amonia, sintesis protein mikroba dan aktivitas enzim selulase rumen sapi Bali secara in vitro. Penelitian ini
menggunakan Rancangan Acak Kelompok (RAK) dengan 3 perlakuan dan 4 kelompok sebagai ulangan berdasarkan waktu pengambilan cairan rumen. Perlakuan pada penelitian ini meliputi perlakuan ransum kontrol (K), perlakuan ransum kontrol ditambah minyak kedelai 5% (M), dan perlakuan ransum kontrol ditambah sabun kalsium-minyak kedelai 5% (S). Data diolah menggunakan analisis ragam (ANOV A). Hasil penelitian menunjukkan bahwa suplementasi sabun kalsium-minyak kedelai 5% tidak berpengaruh nyata terhadap nilai pH, konsentrasi amonia dan sintesis protein mikroba. Sebaliknya, penambahan sabun kalsium-minyak kedelai 5% dalam konsentrat mampu meningkatkan aktivitas enzim selulase.
Kata kunci: amonia, enzim selulase, minyak kedelai, sabun kalsium, sintesis protein mikroba rumen
ABSTRACT
SANTA LUSY A SlMANJUNT AK. Microbial Protein Synthesis and Activity of Cellulase Enzyme In Vitro with Addition of Soybean Oil Calcium Soap. Supervised by SRI SIHARTI and DEWI APRI ASTUTI.
Soybean oil-calcium soap is soap-protected soybean oil as a source of polyunsaturated fatty acids. Provision of soybean oil-calcium soap can removes negative effects of unsaturated fatty acids on rumen microbes and optimize rumen fermentation. The research was aimed to study the effect of soybean oil calcium soap on ammonia concentration, microbial protein synthesis and the activity of cellulase enzymes in vitro. The experiment was designed in completely randomized block design (RBD) with 3 treatments and 4 block based on time rumen sampling. The treatments were control diet (C), C
+ 5% soybean oil (M),
and C + calcium-soap of 5% soybean oil (S). The results showed that supplementation of calcium-soap soybean oil at level 5% did not significantly affect the pH value, NH3 concentration and microbial protein synthesis. Mean while it could increase the activity of cellulase enzyme.Keywords: ammonia, calcium soap, cellulose, rumen microbial protein synthesis, soybean oil
SINTESIS PROTEIN MIKROBA DAN AKTIVITAS ENZIM SELULASE
RUMEN SAPI BALI
IN VITRO
DENGAN PENAMBAHAN
SABUN KALSIUM-MINY AK KEDELAI
SANTA LUSYA SDdANJUNTAK
Skripsi
sebagai salah satu syarat untuk memperoleh gelar SaIjana Petemakan
pada
Departemen Ilmu Nutrisi dan Teknologi Pakan
DEPARTEMEN ILMU NUTRISI DAN TEKNOLOGI PAKAN
FAKULTASPETERNAKAN
INSTITUT PERT
AN IAN
BOGOR
Judul Skripsi: Sintesis Protein Mikroba dan Aktivitas Enzim Selulase Rumen Sapi
Nama NIM
Bali In Vitro dengan Penambahan Sabun Kalsium-Minyak Kedelai : Santa Lusya Simanjuntak
: 024100026
Oisetujui oleh
Dr Sri Suharti, SPt MSi Pembimbing I
Tanggal Lulus: (
05
MAY 2015
Prof Dr Ir Oewi Apri Astuti. MS Pembimbing II
Ketua Oepartemen
)
PRAKATA
Puji dan syukur penulis panjatkan kepada Tuhan Yang Maha Esa atas rahmat dan karunia-Nya sehingga penulis dapat menyelesaikan Skripsi ini. Skripsi yang berjudul " Sintesis Protein Mikroba dan Aktivitas Enzim Selulase Rumen Sapi Bali In Vitro dengan Penambahan Sabun Kalsium-Minyak Kedelai" merupakan salah satu syarat untuk mendapat gelar sarjana pada program mayor Ilmu Nutrisi dan Teknologi Pakan, Fakultas Petemakan, Institut Pertanian Bogor. Skripsi ini disusun berdasarkan hasil penelitian pada bulan Januari hingga Juli 2014.
Sapi bali adalah salah satu sapi lokal Indonesia yang sangat berpotensi sebagai sumber daging dalam memenuhi kebutuhan daging nasional. Tetapi kandungan asam lemak jenuh pada daging sapi Bali tinggi. Oleh karena itu diperlukan upaya yang dapat meningkatkan kandungan asam lemak tak jenuh pada daging sapi . Minyak kedelai merupakan salah satu minyak nabati yang kaya asam lemak tak jenuh. Pemberian minyak kedelai yang diproteksi dengan sabun kalsium berpotensi dalam meningkatkan kandungan asam lemak tidak jenuh, produk fermentasi dan sintesis protein mikroba. Penulisan Skripsi ini bertujuan untuk rnengamati penambahan minyak kedelai yang diproteksi sabun kalsium terhadap konsentrasi NH3, sintesis protein mikroba dan aktivitas enzim selulase
mikroba rumen sapi Bali secara in vitro.
Penulis menyadari bahwa skripsi ini masih jauh dari sempuma. Penulis berharap Skripsi ini bermanfaat bagi pembaca dan kekurangan yang ada pada tulisan ini dapat diperbaiki dalam tulisan selanjutnya
Bogor, April 2015
DAFTAR TABEL DAFT AR LAMP IRAN PENDAHULUAN METODE
Materi
Bahan dan Alat Ransum Perlakuan Lokasi dan Waktu Prosedur
Pembuatan Sabun Kalsium
DAFTARISI
F ermentasi in vitro
Pengukuran Konsentrasi NHJ
Perhitungan Sintesis Protein Mikroba Rumen Pengukuran Aktivitas Enzim Selulase
Rancangan Percobaan Perlakuan
Peubah Penelitian
HASIL DAN PEMBAHASAN
Pengaruh PerlakUan terhadap Produksi pH dan Amonia Sintesis Protein Mikroba (SPM)
Aktivitas Enzim Selulase SIMPULAN DAN SARAN
Simpulan Saran
DAFTAR PUSTAKA LAMPIRAN
RIWAYATHIDUP
UCAP AN TERIMA KASIH
DAFfAR TABEL
I Susunan ransum penelitian 4
2 Kandungan zat makanan ran sum komplit berdasarkan bahan kering 4 3 Rataan pH rumen dan konsentrasi NH3 pada perlakuan in vitro 8 4 Rataan sintesis protein mikroba rumen Hュセ@ 10mr') 10 5 Rataan aktivitas enzim selulase Hセュッャ@ mr jam-I) I I
DAFfAR LAMPIRAN
I Hasil analisis ragam konsentrasi arnonia 2 Hasil anal isis ragarn sintesis protein mikroba
PENDAHULUAN
Penyediaan dan peningkatan produktivitas ternak sapi lokal merupakan upaya dalam memenuhi permintaan daging dalam negeri serta menekan impor daging sapi. Sapi Bali merupakan salah satu sapi lokal asli Indonesia yang sangat potensial untuk dikembangkan. Namun, permasalahan dalam budidaya sapi Bali yaitu pertumbuhan anak sapi rang rendah dengan pertambahan bobot badan harian sekitar 0.58-0.68 kg harr (Parwati et al. 1999). Menurut Komariah (1997) bahwa kandungan asam lemak jenuh daging sapi Bali tinggi namun kandungan asam lemak tak jenuhnya rendah. Tingginya asam lemak jenuh dikarenakan semua lemak pakan terutama asam lemak tak jenuh yang melewati rumen men gal ami proses biohidrogenasi menjadi asam lemak jenuh. Selain itu, asam lemak takjenuh tinggi dalam pakan memberikan efek negatifterhadap fermentasi rumen.
Secara a1ami, daging sapi tinggi asam lemak tak jenuh karena sapi memiliki gen Stearoyl-CoA Desaturase (SCD). Oen SCD mengkode suatu enzim yang dapat mengkatalis proses konversi asam lemak jenuh ke asam lemak tidak jenuh. Selain itu, sapi yang diberi makan rumput dan ransum kaya asam lemak tak jenuh yang diproteksi akan menghasilkan daging sapi tinggi asam lemak tak jenuh terutama lemak omega-3 dan conjugated linoleic acid (Plank 2007). Lemak yang dikonsumsi ruminansia mengalami proses metabolisme di dalam rumen dan pasca rumen. Lemak pakan yang masuk ke dalam rumen mengalami dua proses yaitu proses hidrolisislliposis ikatan ester dan proses biohidrogenasi. Lipolisis lemak dilakukan oleh bakteri Anaerovibrio lipolylica dan Butyrivibrio fibrisolvens
(Harfoot dan Hazlewood 1997) dan terhidrolisis menjadi asam lemak bebas, gula, fosfat dan gliserol. Oula dan gliserol diubah menjadi VFA kemudian digunakan untuk sintesis sel mikroba rumen. Asam-asam lemak bebas tak jenuh mengalami isomerasi dari posisi cis menjadi trans dan proses biohidrogenasi menjadi asam lemak jenuh serta proses konjugasi asam lemak tak jenuh (Bauman dan Lock 2006) Proses biohidrogenasi merupakan proses untuk menghilangkan hidrogen yang terbentuk selama fennentasi rumen. Proses biohidrogenasi dilakukan dua grup bakteri yaitu bakteri grup A yang menghidrolisis asam lemak tak jenuh menjadi asam lemak trans 18:1 dan bakteri grup B yang menghidrolisis asam lemak trans 18:1 menjadi asam stearat (Lock et al. 2006; Bauman dan Lock 2006).
Proses pencernaan ruminansia sangat dipengaruhi oleh proses fennentasi di dalam rumen. Pemberian pakan harus memenuhi kebutuhan nutrien lernak, mensuplai nutrien bagi pertumbuhan mikroba rumen, dan menjaga kondisi optimum rumen. Salah satu nutrien yaitu lemak akan mengalami proses fermentasi di dalam rumen. Penggunaan lemak yang tinggi pada ruminansia dapat mengganggu proses fermentasi di dalam rumen. Pemberian minyak nabati tanpa diproteksi dapat menurunkan nilai pH rumen. Menurut Jenkins (1993) bahwa penambahan asam lemak tanpa diproteksi menurunkan pH rumen karena adanya penurunan aktivitas protozoa yang mengakibatkan kemampuan protozoa dalam menstabilkan pH juga menurun. Selain itu, penambahan lemak pada pakan menurunkan konsentrasi amonia. Penelitian Beauchemin et al. (2007) menyatakan bahwa penambahan 3.4% lemak dari biji bunga matahari, tallow, dan bunga matahari menurunkan konsentrasi amonia dibandingkan dengan kontrol (tanpa
2
penambahan minyak). Penurunan konsentrasi amonia akan berdampak pada pembentukan (sintesis) sel mikroba yang menurun karena amonia sebagai prekursor utama dan sumber nitrogen utama dalam sintesis protein mikroba. Pemberian lemak yang tidak diproteksi lebih dari 3%-4% akan mengurangi aktivitas mikroba rumen terutama bakteri pencema serat (termasuk aktivitas enzim) dan menekan pencemaan selulosa (Czerkawski et al. 1996) dan menurunkan penguraian serat oleh mikroba ( Harvantine dan Allen 2006). Asam lemak tak jenuh memiliki efek negatif pada mikroba rumen yaitu bersifat toksik bagi mikroba selulolitik. Kerja bakteri dipengaruhi oleh aktivitas enzim sebingga apabila aktivitas mikroba rumen terganggu juga mengganggu aktivitas enzim (Stewart 1977). Penelitian tentang pengaruh penambahan minyak/lemak terhadap aktivitas enzim belum banyak dilakukan khususnya pengaruh rninyak kedelai terhadap aktivitas enzim selulase belum ada.
Minyak kedelai merupakan salah satu minyak tanaman yang berpotensi untuk digunakan dalam memodifikasi komposisi asam lemak daging sapi. Minyak kedelai merupakan salah satu minyak yang banyak mengandung asam lemak poli tak jenuh (Polyunsaturated Fatty Acid, PUFA) cukup tinggi sekitar 84.6% yang terdiri dari asam oleat 23.3%, asam linoleat 53.7% dan linolenat 7.6% (O'Brien 2009). Sebelumnya sudah dikaji dalam pemilihan minyak yang akan digunakan yaitu minyak kedelai, minyak sawit, dan minyak biji bunga matahari, dilihat pengaruh yang diproteksi dengan sabun kalsium dengan yang tidak diproteksi. Hasil dari fermentasi yang terbaik adalah minyak kedelai (Bain et al. 2014).
Selain itu, ketersediaan minyak kedelai cukup banyak dan harganya cukup murah dibandingkan jenis minyak lainnya. Pemberian ransum yang kaya asam lemak tak jenuh merniliki korelasi positif dengan asam lemak tak jenuh pada daging. Penelitian Lubis dan Wina (1998) bahwa pemberian 10% kalsium lemak sawit dalam konsentrat meningkatkan asam lemak tak jenuh pada daging domba secara nyata yaitu asam linoleat (CI8:3), asam linolenat (CI8:2) dan asam erurat (C22:1), serta menurunkan kandungan total asam lemak jenuh dalam daging.
Penambahan minyak kaya PUF A yang diberikan dalam bentuk terproteksi untuk menghindari proses biohidrogenasi dalam rumen dan diharapkan tidak mengganggu aktivitas mikroba rumen. Teknologi sabun kalsium dapat melindungi asam lemak tak jenuh ganda (PUFA) sebesar 39.65% atau kehilangan 60.35% PUFA selama proses biohidrogenasi dalam rumen secara in vitro (Joseph 2007). Selain itu, penambahan 1emaklminyak tinggi dalam ransum temak memberikan dampak negatif yaitu membatasi pencemaan serat dan menurunkan aktivitas enzim. Menurut Suharti et al. (2015) bahwa suplementasi sabun kalsium dengan bahan minyak kanola dan flaxseed pada level 6% meningkatkan konsentrasi amonia dan produksi VF A total.
3 stabilitas struktur dinding sel dan sangat penting untuk sintesis potein mikroba rumen serta diperlukan oleh mikroba rumen untuk mencema selulosa.
Penelitian ini bertujuan untuk mengamati pengaruh penambahan minyak kedelai yang diproteksi sabun kalsium terhadap nilai pH, konsentrasi amorua, sintesis protein mikroba dan aktivitas enzim selulase secara in vitro.
METODE
Materi
Bahan dan A1at
Bahan yang digunakan untuk uji NH3 antara lain vaselin, larutan Na2C03
jenuh, larutan asam borat berindikator dan larutan H2S04 0.005 N. Bahan yang digunakan untuk uji sintesis protein mikroba adalah larutan reagen pembentukan kompleks, NaOH 2 N, NaOH 0.25 N dan reagen Folin-Ciocalteu. Bahan yang digunakan untuk uji aktivitas enzim selulase antara lain enzirn lisozim, larutan CC4, buffer phosphate 0.1 M (PH 6.8), larutan carboxymethyl cellulose (CMC) 1 %, larutan standar selulosa dan larutan ONS (Dinitrosalicylic Acid).
Peralatan yang digunakan adalah vakum, terrnos, kain penyaring, timbangan analitik, tabung reaksi, rak tabung reaksi, pipet volumetrik:, bulp, mikropipet, pH meter, tabung effendorf, tabung fermentor, waterbath, shaker waterbath, sentrifus, magnetic stirrer, cawan Conway, mikro sentrifuse, centrifoge beckman, vortex dan spektrofotometer.
Ransum Perlakuan
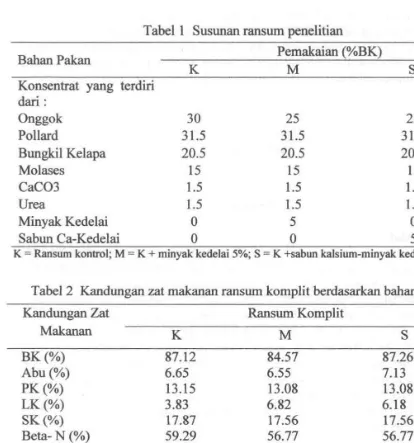
Pakan yang digunakan dalam penelitian adalah ransum disusun berdasarkan kebutuhan temak sapi potong 250 kg dengan protein kasar (PK) 13%, Total Digestible Nutrien (TON) 67% (Kearl 1982). Ransum terdiri atas hijauan dan konsentrat dengan rasio hijauan dan konsentrat adalah 40:60. Susunan ransum perlakuan yang digunakan dalam penelitian dapat dilihat pada Tabel I dan kandungan zat makanan ransum komplit berdasarkan bahan kering dapat dilihat pada Tabel 2.
4
Tabel I Susunan ransum penelitian
K
M
s
BahanPakan Pemakaian (%BK)
Konsentrat yang terdiri dari :
Onggok 30 25 25
Pollard 31.5 31.5 31.5
Bungkil Kelapa 20.5 20.5 20.5
Molases 15 15 15
Cacm 1.5 1.5 1.5
Urea 1.5 1.5 1.5
Minyak Kedelai 0 5 0
Sabun Ca-Kedelai 0 0 5
K - Ransurn kontrol; M - K + rninyak kedelai 5%; S - K +sabun kalsiurn-minyak kedelai 5 %
Tabel 2 Kandungan zat makanan ransum komplit berdasarkan bahan kering
Kandungan Zat Ransum Komplit
Makanan K M S
BK (%) 87.12 84.57 87.26
Abu (%) 6.65 6.55 7.13
PK (%) 13.15 13.08 13.08
LK (%) 3.83 6.82 6.18
SK(%) 17.87 17.56 17.56
Beta-N (%) 59.29 56.77 56.77
roN1 (%) 68.87 66.70 66.70
Ca (%) 0.70 0.70 0.70
P (%) 0.31 0.31 0.31
BK (Bahan Kering), PK (Protein Kasar), LK (Lemak Kasar), SK (Serat Kasar), Beta-N (Bahan Ekstrak tanpa Nitrogen), TON (TotaJ Digestibility Nutrient), Ca (Kalsium), P (Phosfor).
K セ@ Ransum kontrol; M セ@ K + rninyak kedelai 5%; S セ@ K +sabun kalsium-rninyak kedelai 5%
I : Hasil perhitungan TON berdasarkan rumus Sutardi (2001) TON セ@ 70.6 + 0.259 x PK + 1.01 x
LK - 0.76 x SK + 0.0991 x Beta-N. K セ@ Ransurn kontrol; M セ@ K + rninyak kedelai 5%; S セ@ K +sabun kalsium-minyak kedelai % %
Lokasi dan Waktu
[image:12.606.59.473.65.508.2]5
dengan stomach tube yang dipelihara di kandang A Fakultas Petemakan Institut Pertanian Bogor. Penelitian dilakukan dari Januari hingga Juli 2014.
Prosedur
Pembuatan Sabun Kalsium
Sabun kalsium dibuat menggunakan metode Kumar et al. (2006). Sebelum pembuatan sabun kalsium, dilakukan pengamatan terhadap bilangan penyabunan. Bilangan penyabunan adalah bilangan yang menyatakan jwnlah mg KOH yang dibutuhkan untuk menyabun I gram lemak/mioyak. Penentuan bilangan penyabunan minyak dilakukan dengan cara refluksi minyak yang ditambahkan larutan KOH beralkohol. Larutan KHO beralkohol dibuat dengan mencampurkan 1.4 gr KOH dan 50 ml alkohol 95%. Larutan KOH dibuat dengan mencampurkan KOH dan pelarut akuades sedangkan KOH beralkohol dibuat dengan mencampurkan KOH dan alkohol 95%. Sampel minyak sebanyak 5 ml dicampurkan dengan 50 ml larutan KOH beralkohol dimasukkan kedalam labu penangas berleher untuk direfluksi dan dititrasi sehingga didapat bilangan penyabunan dan jwnlah NaOH yang digunakan. Minyak dipanaskan dan ditambahkan larutan NaOH secara perlahan, strirrer selama 30 memt kemudian diteteskan larutan CaCh hingga membentuk endapan. Endapan tersebut ditempatkan pada aluminium foil hingga membeku dan kemudian dimasukkan ke oven 60°C selama 24 jam dan sabun kalsium siap untuk digunakan.
Fermentasi
in
vitroFerrnentasi in vitro dilakukan menggunakan metode Tilley dan Terry (1963). Cairan rumen diperoleh dari sapi Bali yang dipelihara di kandang A Fakultas Petemakan IPB. Cairan rumen dimasukkan ke dalam terrnos yang sebelurnnya diisi air panas 100°C, selanjutnya cairan rumen disaring dengan kain kasa. Tabung ferrnentor yang telah diisi 0.5 g sampel ransum perlakuan, kemudian ditambahkan \0 ml cairan rumen dan 40 ml larutan McDougall. Tabung dimasukkan ke dalam shaker water bath dengan suhu 39 °C dan dikocok dengan dialiri CO2 selama 30 detik. Tabung ferrnentor ditutup dengan karet berventilasi dan diferrnentasi selama 4 jam. Setelah 4 jam, tutup karet dibuka dan diukur pH rumen menggunakan pH meter. Cairan rumen ditetesi HgCh 2-3 tetes untuk membunuh milcroba dan disentrifuse dengan kecepatan 4000 rpm selama 10 mem!. Pengukuran Konsentrasi NHJ
6
dan dimiringkan untuk mencampur larutan Na2C03 jenuh dengan supernatan hingga merata. Cawan Conway dibiarkan dalam suhu ruang selama 24 jam. Setelah 24 jam, cawan Conway dibuka dan asam borat berindikator dititrasi dengan H2S04 0.005 N sampai terjadi perubahan warna dari biru menjadi merah jambu.
Konsentrasi NH3 dihitung dengan rumus: NH (roM) = Volume H2S04 x N H2S04 x 1000
3 9 sampel x BK sampel
Perhitungan Sintesis Protein Mikroha Rumen
Perhitungan sintesis protein mikroba menggunakan metode Makkar el al. (1982) kemudian dilanjutkan dengan metode Lowry's el al. (1951)
Tahapan sintesis protein sebagai berikut: I. Pembuatan reagen pembentukan kompleks
Reagen pembentukan kompleks dibuat dari tiga jenis larutan yaitu larutan A (2% b/v Na2C03 dalam NaOH 0.1 N), larutan B (0.5% b/v CuS04.5H20 dalam K-Na-Tartrat 1%). Larutan A sebanyak 50
mI
dicampurkan dengan larutan B sebanyak 1mI.
Reagen pembentukan kompleks hanya stabil selama satu hari.2. Larutan NaOH 2N
3. Pembuatan reagen folin-ciocalteu
Reagen folin-ciocalte dibuat dengan cara memasukkan 100 g sodium tungstate ke dalam labu erlenmeyer berukuran 500
mI,
kemudian tambahkan 25 g sodium molibdate, 700mI
akuades, 50 ml asam phosphate, dan 100 ml HC!. Campuran direfluks selama 10 jam kemudian tambahkan ISO mllithium sulfat, 50mI
akuades, dan beberapa tetes bromine. Campuran didihkan sekitar IS menit hingga bromine habis, dan dinginkan kembali lalu diencerkan dengan akuades I L serta disaring (filtrat berwarna kehijauan). Sebelum digunakan, filtrat harus diencerkan terlebih dahulu dengan perbandingan 1:5 (I bagian filtrat dengan 5 bagian akuades).Prosedur pengukuran sintesis protein mikroba dimulai dengan mempersiapkan alat destilasi. Cairan rumen sebanyak 20 ml didestilasi menggunakan magnetic stirrer dengan kecepatan 400 rpm selama 45 detik. Hal ini bertujuan untuk memisahkan bakteri dengan sampe!. Sampel disentrifuse kembali pada 408 gravitasi selama 5 menit, bertujuan untuk menurunkan populasi protozoa dan menghilangkan sisa partikel pakan, yang disebut dengan aliquot.
7
Supematan I ml ditambahkan larutan NaOH 2N sebanyak I ml, kemudian dihidrolisis pada suhu 100°C selama 10 men it pada penangas air. Sam pel didinginkan pada suhu ruangan lalu tambahkan 5.5 ml reagen pembentukan kompleks. Setelah itu, larutan dibiarkan pada suhu ruangan selama 10 menit dan ditambahkan 0.5 ml reagen Folin-Ciocalteu, homogenkan dengan vortex, lalu didiamkan selama 30-60 men it Gangan sampai lebih dari 60 men it). Absorbansi dibaca pada 670 nm.
Pengukuran Aktivitas Enzim Selulase
Pengukuran aktivitas enzim menggunakan metode Patra (2006). Pengukuran aktivitas enzim menggunakan cairan rumen perlakuan yang difermentasi selama 4 jam sebagai sumber enzim. Cairan rumen setelah inkubasi 4 jam dimasukkan kedalam beaker glass 100 ml, kemudian ditambahkan karbontetraklorida dan lisozim (konsentrasi 0.4 g 100 mrl buffer phosphate). Inkubasi pada suhu 40°C selama 3 jam, yang diikuti sonikasi pada suhu 4 °C. Setelah itu larutan disentrifuse pada kecepatan 14000 rpm selama 20 menil. Hasil sentrifuse berupa endapan dan supematan. Endapan dibuang dan supematan diambil sebagai sumber enzirn. Aktivitas enzim selulase dianalisis dengan inkubasi campuran larutan assay sebanyak 2 m!. Larutan assay terdiri dari larutan buffer fosfat 0.1 M (pH 6.8) sebanyak I ml, larutan carboxymethyl cellulose (CMC) 1% b/v (I g 100 mrl buffer fosfat) sebanyak 0.5 ml, dan larutan enzim ekstraseluler yang telah diencerkan sebanyak 0.5 m!. Campuran tersebut diinkubasi selama 60 menit pada suhu 39°C.
Gula-gula hasil reduksi yang dihasilkan diestimasi sebagai monosakarida dengan metode dinitrosalicylic acid (Miller 1959). Ambil sampel sebanyak J ml dan ditambahkan I ml air destiJasi (d. H20) dan 3 ml larutan DNS. Didihkan
selama 15 men it kemudian didinginkan selama 20 menit dalam air es. Setelah dingin, campuran tersebut diukur panjang gelombangnya (540 nm) dengan spektrofotometer. Prosedur untuk larutan blanko sarna dengan larutan sampel, namun larutan enzim (sarnpel) tidak dimasukkan. Aktivitas enzim didefinisikan sebagai jumlah enzim yang menghasilkan J mg monosakarida per jam per ml pada suhu 39
°c.
Akt· · S J I IVltas e u ase I1mo m Jam ( I I-I. .1) 8M Glukosa x AxFpxlOOO t
Keterangan: A Fp
Perlakuan
BM glukosa T
=
konsentrasi glukosa sarnpel = faktor pengencer (7.34) = 180= waktu dalam jam
Rancangan Percobaan
8
K : Kontrol (ransum dengan perbandingan hijauan:konsentrat
=
40:60) M : K +Minyak Kedelai 5%S : K+Sabun Kalsium-Minyak Kedelai 5% Persamaan matematik yang digunakan:
V lj = J1+ Ti + Ilj + E;j
Keterangan :
Yij : nilai pengamatan dari perlakuan ke-i pada kelompok ke-j
fl : rataan umum
tj : pengaruh aditif dari level sabun kalsium ke-i
I3j
:
pengaruh aditif dari kelompok ke-jEij :pengaruh galat percobaan pada sabun kalsium ke-i pada kelompok ke-j
Parameter NH3, pH dan SPM dianalisis menggunakan anal isis ragam
(analysis of variance) dan bita teljadi perbedaan dilanjutkan dengan uji jarak duncan (Steel dan Torrie 1993). Data aktivitas enzim selulase dianalisis secara deskriptif.
Peubab Penelitian
Peubah yang diamati dalam penelitian ini adalah konsentrasi amonia (NH3) dan pH rumen, sintesis protein mikroba, dan aktivitas enzim selulase rumen.
BASIL DAN PEMBAHASAN
Pengarub Perlakuan terbadap Produksi pH dan Amonia
Hasit anaJisis ragam menunjukkan bahwa suplementasi minyak kedelai 5% baik dalam bentuk minyak maupun terproteksi dengan sabun kalsium tidak memberikan pengaruh yang nyata terhadap nilai pH dan produksi amonia. Hasit pengukuran produksi pH dan amonia pada penelitian ini disajikan pada Tabel 3.
Tabel 3 Rataan pH dan konsentrasi N-NH3 rumen dengan penambahan minyak kedelai dalam konsentrat
Peubah
pH
N-NH3 (mM)
K
6.79 ± 0. 10 10.65± 1.71
Perlakuan M 6.79 ± 0.05
9.86±1.85
S 6.84 ± 0.07
9.67± 1.91
K - Ransum kontrol ; M - K + minyak kedelai 5%; S - K +sabun kalsium-minyak kedelai 5%
[image:16.601.84.464.522.606.2]9 mendegradasi pakan dan berlangsungnya proses fennentasi adalah 5.5-7.6. Jalc et at. (2007) melaporkan bahwa penambahan sebesar 3.5% asam lemak tak jenuh (oleat, linoleat, dan a-linolenat) pada pakan berbasis 80% lucerne and 20% barley belum memberikan perubahan terhadap nilai pH rumen yaitu berkisar 6.73-6.93 .
Nilai pH rumen berhubungan dengan fermentasi rumen. Jika pH rumen lebih rendah dari 6.2 maka proses pencemaan serat terganggu karena pH cairan rumen yang ideal untuk proses pencernaan seluJosa adalah 6.4-6.8 (Joseph 2007). pH rendah juga mengganggu mikroba khususnya protozoa. pH rendah dapat menurunkan kecemaan jaringan tumbuhan berserat dan menyebabkan energi dalam rumen dialihkan ke fungsi non-pertumbuhan, seperti menjaga kestabilan pH sehingga mengganggu sintesis sel mikroba (pathak 2008).
Penelitian ini menunjukkan bahwa pemberian minyak kedelai baik dalam bentuk terproteksi sabun kalsium maupun minyak tidak mempengaruhi nilai pH rumen karena jumlah pemberian minyak kedelai baik diproteksi sabun kalsium maupun dalam bentuk minyak masih dalam taraf yang rendah yaitu 5% dalam konsentrat. Selain itu, sabun kalsium tetap utuh dalam cairan rumen dengan pH netraJ dan terurai pada kondisi asam (PH 2-3) sehingga tidak mengganggu aktivitas mikroba rumen. Hal ini sesuai dengan penelitian Kowalski (1997) menyatakan bahwa penambahan sabun kalsium minyak kanola bersifat inert dalam rumen sehingga tidak mengubah pH rumen.
Rataan konsentrasi amonia yang dihasilkan dari semua perlakuan berkisar antara 7.53-12.40 mM. Konsentrasi amonia tersebut masih optimal untuk pertumbuhan mikroba rumen yaitu berkisar antara 6-21 mM (McDonald et at. 2002). Penambahan rninyak kedeJai dan sabun kalsium-minyak kedelai tidak mempengaruhi konsentrasi amonia Secara umum teIjadi kecenderungan penurunan amonia dengan penggunaan minyak kedelai atau sabun kalsium-rninyak kedelai. Penurunan amonia disebabkan pemanfaatan pakan oleh rnikroba rumen dalam kondisi normal untuk aktivitas pencemaan pakan dan perlakuan proteksi mengakibatkan protein ransum lolos dari degradasi rumen sehingga amonia menurun.
Konsentrasi amonia merupakan salah satu indikator untuk menilai degradasi protein dan sintesis protein bakteri. Penelitian Nurhanah (2014) bahwa pemberian rninyak kedelai terproteksi tidak nyata meningkatkan kecemaan protein kasar, tetapi penambahan rninyak kedelai terproteksi menghasilkan kecemaan protein kasar cenderung lebih tinggi dibandingkan ransum dengan minyak kedelai tidak diproteksi sabun kalsium. Kecemaan protein kasar yang tinggi merupakan indikator bahwa sebagian besar protein pakan didegradasi menjadi amonia sehingga meningkatnya kecernaan protein kasar diikuti konsentrasi amonia meningkat.
10
kedelai dan biji kedelai Iebih baik daripada perlakuan lemak yang diproteksi. Namun secara keseluruhan penambahan minyak kedelai pada ransum sapi Bali daIam bentuk sabun tidak memberikan pengaruh terhadap pertumbuhan mikroba sehlngga metabolisme protein dan fermentasi karbohidrat berIangsung secara optimal.
Sintesis Protein Mikroba (SPM)
Penambahan minyak kedelai daIam ransum baik daIam bentuk minyak maupun terproteksi sabun kalsium dengan level 5% tidak memberikan pengaruh terhadap sintesis protein mikroba dan efisiensi sintesis protein. Rataan sintesis protein mikroba rumen disajikan pada Tabel 4.
Tabel4 Rataan sintesis protein mikroba rumen (mg 10mr') Peubah
SPM Efisiensi SPM
K 24.52±12.8
11.69±6.1
PerIakuan M 27.08±12.33
I2.54±5.71
S 21.l3± 10.98
9.96±5.18
K - Ransum kontrol; M - K + minyak kedelai 5%; S - K +sabun kalsium-minyak kedelai % %
*
Efisiensi SPM=SPMxBOFR, BOFR (baban organik terfermentasi dalam rumen)=O.65xbahan organik tercerna (lAEA 1997)Hasil yang didapat menunjukkan bahwa perIakuan tidak memberikan pengaruh terhadap sintesis protein mikroba, artinya sintesis protein mikroba tidak terganggu dengan kehadiran sabun kalsium-minyak kedelai. Penambahan minyak kedelai dalam ransum cenderung meningkatkan sintesis protein mikroba. Hasil ini sesuai dengan penelitian Suhartati dan Suryapratama (2012) bahwa penambahan minyak kedelai 3% dalam ransum meningkatkan sintesis protein mikroba rumen. Menurut Pathak (2008) bahwa faktor-faktor yang mempengaruhl sintesis protein mikroba antara lain konsumsi bahan kering, suplai senyawa nitrogen, suplai energi terfermentasi, rasio hljauan:konsentrat pada ransum, sinkronisasi nitrogen dan energi, lingkungan rumen, laju makanan, vitamin dan mineraI. Amonia merupakan sumber nitrogen utama yang digunakan untuk pembentukan protein mikroba rumen. Sekitar 70%-80% protein didegradasi menjadi peptida dan asam amino serta diubah lanjut menjadi amonia. Menurut Fiorentini el af. (2013) bahwa sapi yang diberi makan minyak kedelai dan biji kedelai meningkatkan efisiensi sintesis protein mikroba dibandingkan minyak kedelai terproteksi.
II
pertumbuhan mikroba menurun. Energi yang dihasilkan tidak digunakan untuk sintesis protein mikroba, melainkan akwnulasi karbohidrat sel mikroba (Ointing 2005). Menurut Nolan dan Leng (1984) babwa amonia adalab sumber nitrogen utama untuk sintesis sel mikroba rumen. Sebanyak 80% bakteri rumen terutama bakteri selulolitik menggunakan ammonia sebagai satu-satunya sumber nitrogen untuk pertumbuhan. VF A merupakan produk akhir fermentasi karbohidrat dan sebagai sumber energi utama bagi lernak serta sebagai kerangka karbon untuk sintesis bakteri. Proses sintesis protein bakteri akan optimal apabila teIjadi keseimbangan produk amonia dan VF A. Kisaran VF A total yang mendukung sintesis protein mikroba yaitu 70-150 mM (McDonald e/ al. 2002). Konsentrasi amonia yang mendukung pertumbuhan mikroba rumen adalab 4-12 mM (Sutardi 1979).
Aktivitas Enzim Selulase
Hasil analisis deskriptif menunjukkan babwa suplementasi rninyak kedelai 5% tanpa diproteksi sabun kalsium (M) maupun diproteksi sabun kalsium (S) dalam ransum meningkatkan aktivitas enzim selulase secara in vitro. Hasil pengukuran aktivitas enzim selulase pada penelitian ini disajikan pada Tabel 5.
Tabel5 Peubab
Aktivitas Enzim Selulase
Rataan aktivitas enzim selulase (Ilmol mI-1 jam -I) Perlakuan
K M S
164±89.69 114.56±29.61 229.50±122.66
K Ransum kontrol; M - K + minyak kedelai 5%; S - K +sabun kalsium-minyak kedelai 5 %
Ransum yang diberi perlakuan sabun kalsium meningkatkan aktivitas enzim selulase sekitar 49%.Menurut Moharrery dan Das (2002) babwa aktivitas enzim selulase dari cell free rumen fluid (CFRF) 162.2-249.1 Ilmol mI-1 jam-I. Penambaban rninyak kedelai dalam bentuk sabun meningkatkan aktivitas enzim selulase. Aktivitas enzim selulase yang meningkat pada perlakuan sabun kalsium-rninyak kedelai 5% dalam konsentrat secara keseluruhan tidak diikuti peningkatan kecernaan pakan, tetapi kecernaan lemak pakan dan protein meningkat. Penurunan aktivitas enzim selulase yang diberi minyak kedelai dikarenakan pemberian rninyak mengganggu aktivitas mikroba selulolitik. Peningkatan aktivitas enzim selulase karena minyak kedelai yang diproteksi sabun kalsium tidak bersentuhan dengan partikel pakan sehingga akses permukaan membran sel mikroba rumen dengan pakan tidak terganggu akibatnya pencernaan serat terutama selulosa optimal.
Menurut Wina dan Susana (2013) babwa nilai kecernaan pakan in vitro
12
el af. (2000) bahwa aktivitas enzim dipengaruhi oleh banyak faktor yaitu suhu, pH
atau keasaman, konsentrasi atau jumlah enzim, dan inhibitor enzim. Sebaliknya, penurunan aktivitas enzim pada pemberian minyak kedelai tidak diproteksi sabun disebabkan minyak menutupi partikel pakan sehingga akses permukaan membran sel mikroba bersentuhan dengan pakan terhambat, akibatnya mengganggu fermentasi rumen serta mengurangi pencemaan serat (Tiven el al. 2011). Meningkatnya kecemaan ransum yang disuplementasi sabun kalsium sebagai indikasi meningkatnya aktivitas enzim seluJase yang dihasilkan oleh bakteri pendegradasi seluJosa.
Hasil penelitian ini didukung dengan hasil penelitian Aryani (2015) bahwa pemberian minyak kedelai tidak diproteksi maupun diproteksi sabun kalsium tidak memberikan pengaruh yang nyata terhadap popuJasi total bakteri tetapi secara deskriptifpemberian minyak kedelai-sabun kalsium meningkatkan popuJasi total bakteri 1.2 x \06 mJ·1 dibandingkan ransum minyak tidak terproteksi maupun kontrol yaitu 0.3 x 106mJ·l. Aktivitas seluJase dipengaruhi oleh ketersediaan mikroba rumen yaitu bakteri khususnya bakteri seluJolitik.
SIMPULAN DAN SARAN
Simpulan
Pemberian minyak kedelai dalam ransum yang diproteksi sabun maupun tidak diproteksi tidak mempengaruhi pH dan produksi amonia rumen, dan sintesis potein mikoba rumen. Namun sebaliknya, pemberian sabun kalisum-minyak kedelai 5% secara deskriptif cenderung meningkatkan aktivitas enzim seluJase rumen sapi Bali secara in vitro.
Saran
Perlu dilakukan penelitian lebih lanjut mengenai pengaruh pemberian minyak kedelai terproteksi terhadap aktivitas enzim lipase, populasi bakteri selulolitik dan lipolitik untuk melihat keterkaitan antara aktivitas enzim dan popuJasi bakteri spesifik.
DAFfAR PUSTAKA
13
Azizah N. 2011. Fortifikasi ekstrak lerak. dengan mineral mix (Ca, Mg, P, dan S) serta pengaruhnya terhadap karakteristik fermentasi dan sintesis protein bakteri in vitro. [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Bain A, Wiryawan KG, Astuti DA, Suharti S, Arman C. 2014. The effect of protected vegetable fermentation characteristics and nutrient digestibility of Bali cattle rumen fluid. Proceedings of the 16th AAAP Animal Science Congress Vol.
n
"Sustainable Livestock Production in the Perspective oj Food Security, Policy, Genetic Resources and Climate Changes". 10-14November, 2014. Yogyakarta, Indonesia. 308-311.
Bauman DE, Lock AL. 2006. Concepts in lipid digestion and metabolism in dairy cows. In: Eastridge ML, editor. Proceeding of Tri-State Dairy Nutrition Conference. Indiana, 25-26 April 2006. Port Wayne (Indiana): The Diho State University. p. 1-14.
Beauchemin KA, McGinn SM, Petit HV. 2007. Methane abatement strategies for cattle: lipid supplementation of diets. Can J Anim Sci. 87: 431-440.
Czerkawski JW, Blaxter KL, Wainman FW. 1996. The metabolism of oleic, linoleic, linolenic acids by sheep with reference to their effect on methane production. Br J Nutr. 20: 349-362.
Conway EJ, Byrne A. 1933. An absorption for the micro-determination of certain volatile substances. I. The micro-determination of ammonia. Department of Physiology and Biochemistry. University College, Dublin.
Fiorentini G, Messana JD, Dian PHM, Reis RA, Canesin RC, Pires AV, Berchielli IT. 2013. Digestibility, fermentation and rumen microbiota of crossbred heifers fed diets with different soybean oil availabilities in the rumen. Anim Feed Sci Technol. 181 :26-34. Abstract. http://dx.doi.orglI0.1016/j.anifeedsci.20 13.01.0 II.
Ointing SP. 2005. Sinkronisasi degradasi protein dan energi dalam rumen untuk memaksirnalkan produksi protein mikroba. Wartazoa. 15(1): 1-10.
Harfoot CG, Hazlewood GP. 1997. Lipid metabolism in the rumen. In: Hobson PN, Stewart CS. (ed.) The Rumen Microbial Ecosystem. London (UK): Chapman dan Hall. p. 82-42.
Harvatine KJ, Allen MS. 2006. Effects of fatty acid supplements on ruminal and total tract nutrient digestion in lactating dairy cows. J Dairy Sci. 89:
1092-1103.
Jalc D, Certik M, Kundrikova K, Namestkova P. 2007. Effect of unsaturated CI8 fatty acids (oleic, linoleic, and a-linolenic acid) on ruminal fermentation and production of fatty acid isomers in anartificial rumen. Vet Medic. 52(3): 87-94.
Jenkins TC. 1993. Lipid metabolism in the rumen..! Dairy Sci. 76: 3851-3863. Joseph G. 2007. Suplementasi sabun kalsium dalam pakan temak sebagai sumber
energi altematif untuk meningkatkan produksi daging yang berkualitas. [disertasi]. Bogor (ID): Institut Pertanian Bogor.
Kearl LC. 1982. Nutrient Requirements of Ruminants in Developing Countries.
Indonesia (ID): BPT Ciawi.
14
Kowalski ZM. 1997. Rumen fermentation, nutrient flow to the duodenum, anddigestibility in bulls fed calcium soaps of rapeseed fatty acids and soyabeanmeal coated with calcium soaps. Anim Feed Sci Technol. 69: 298 -303.
Kumar R, Sivaiah 1(, Ramana Reddy Y, Ekambram B, Reddy TJ, Reddy GVN. 2006. Effect of supplementation of clietary protected lipids on intake and nutrient utilization in Deccani lambs. Trop Anim Health Prod. 38: 151-158 Lock AL, Harvantine KJ, DrackJey JK, Bauman DE. 2006. Concepts in fat and
fatty acid digestion in ruminants. Proceedings Intermountain Nutrition Conference. New York (US): Cornell University. p. 85-100.
Lowry OH, Rosenbrough NJ, Farr AL, Randall RJ. 1951. Protein measurement with the folin phenol reagent. J Bioi Chern. 193: 265-275.
Lubis D, Wina E. 1998. Carcass production and meat quality of sheep fed high level of rumen bypass fat diets. Bull Anim Sci Suppl: 401-407.
Makkar HPS, Sharma, Dawra RK, Negi SS. 1982. Simple determination of microbial protein in rumen liquor. J Dairy Sci. 65: 2170-2173.
McDonald P, Edwart RA, Greenhall JFD, Morgan CA. 2002. Animal Nutrition.
6thed. New Jersey (US): Prentice Hall Publishing.
Miller GL. 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal Chern. 31(3):367-369.
Moharrery A, Das TK. 2002. Correlation between microbial enzyme activities in the rumen fluid of sheep under different treatments. Reprod Nutr Dev. 41: 513-529.
Nolan N , Leng RA. 1984. Nitrogen metabolism in th rumen. J Dairy Sci. 67(5):
1072-1089.
Nuraida L, Dewanti R, Hariyadi P, Buclijanto S. 2000. Eksplorasi karakterisasi dan produksi enzim lipase dengan aktivitas esterifikasi tinggi dari kapang incligenus. [Laporan Penelitian]. Bogor (ill): Institut Pertanian Bogor. Nurhanah S. 2014. Kecernaan nutrien ransum sapi bali dengan penambahan sabun
kalsium minyak kedelai secara in vitro. [skripsi]. Bogor (ID): Institut Pertanian Bogor.
O' Brien RD. 2009. Fats and Oils Formulating and Processing for Applications 3,d Edition. New York (US) : CRC Press.
Owens FN, Zinn R. 1988. Protein Metabolism of Ruminant Animal Digestive Physiology and Nutrition. New Jersey (US): Reston Boook Prentice Hall, Englewood Cliffs.
Parwati lA, Suyasa N, Guntoro S, Yasa MR. 1999. Pengaruh pemberian probiotik dan laser punktur dalam meningkatkan berat badan sapi Bali. Seminar Nasional Petemakan dan Veteriner 1999. p. 136-146.
Pathak AK. 2008. Various factors affecting microbial protein synthesis in the rumen. Vet World. 1(6): 186-189.
Patra AK. 2006. Effect of plant extracts on in vitro methanogenesis, enzyme activites and fermentation of feed in rumen liquor of buffallo. Anim Feed Sci Technol. 128: 276-291.
Planck N. 2007. Real Food: Hidup Bebas Penyakit dengan Maleanan Alami.
Yogyakarta (ID): B-First.
15 Stewart CS. 1977. Factors affecting the cellulolytic activity of rumen content.
Appl Environ Microbiol. 33(3): 497-502.
Suharti S, Nasution AR, Aliyah DN, Hidayah N. 2015. Potensi minyak kanola dan flaxseed terproteksi sabun kalsiwn untuk mengoptirnalkan fermentasi dan mikroba rumen sapi potong secara in vitro. Pros Sem Nas Masy Biodiv Indon. 1(1): 89-92.
Suryapratama W, Suhartati FM. 2012. Increasing rumen microbial protein synthesis with additional dietary substrate of Saccharomyces cerevisiae and soybean oil. Anim Product. 14(3): 155-159.
Sutardi, T. 1979. Ketahanan protein bahan makanan terhadap degradasi oleh mikroba rumen dan manfaatnya bagi peningkatan produktivitas ternak. [prosiding Seminar Penelitian dan Penunjang Peternakan). Bogor (ID): LPP IPB.
Sutardi T. 2001. Revitalisasi peternakan sapi perah melalui penggunaan ransum berbasis lirnbah perkebunan dan suplementasi mineral organik. Laporan Akhir RUT. Kantor Menteri Negara Riset dan Teknologi dan Lembaga Ilmu Pengetahuan Indonesia.
Tilley JMA, Terry RA. 1963. A. two stage technique for the in vitro digestion of forage corps. J British Grassland Soc. 18: 104-111.
Tiven NC, Yusiati LM, Rusman, Santoso U. 2011. Ketahanan asarn lemak tidak jenuh dalarn Crude Palm Oil terproteksi terhadap aktivitas mikrob rumen
domba in vitro. Med Pet. 34(1): 42-49.
16
Lampiran 1 Hasil analisis ragam konsentrasi amonia
SK Db JK KT F. Hit Sig.
Perlakuan 2 2.1549 1.0774 1.219 .359
Kelompok 3 24.6900 8.2300 9.313 .011
GaJat 6 5.3020 0.8837
Total 12 32.1469
Lampiran 2 Hasil anaJisis ragam sintesis protein mikroba
SK Db JK KT F. Hit Sig.
Perlakuan 2 71.267 35.6335 1.600 .277 Kelompok 3 1175.992 391.9972 17.600 .002
GaJat 6 133.639 22.2731
RIWAYAT HIDUP
Penulis dilabirkan pada tanggal 7 September 1992 di Pokanbaru, Sumatera Utara. Penulis adalab anak kedua dari empat bersaudara dari pasangan Bapak
Amran Simanjuntak dan Ibu Sondang Marilin Purba. Penulis pemab bersekolab di SDN 095132 Pokanbaru tabun 1998-2004, dilanjutkan di SMPN 3 Hutabayuraja tabun 2004-2007 dan SMAN 1 Pematangsiantar pada tabun 2007-2010. Penulis diterima sebagai mabasiswa di Institut Pertanian Bogor (IPB) melalui jalur Undangan Seleksi Masuk IPB (USMI) pada tabun 2010.
17
Selama menjalani pendidikan di IPB penulis aktif mengikuti beberapa organisasi kemabasiswaan yaitu UKM Persekutuan Mabasiswa Kristen (PMK) peri ode 2011-2012 sebagai pengurus Komisi Persekutuan PMK IPB, Himpunan Mabasiswa Nutrisi dan Makanan Temak (HIMASITER) periode 2011-2012 sebagai staf Biro Nutrition Comunity. Penulis juga lulus seleksi program kreatifitas mabasiswa bidang penelitian (PKMP) pada tabun 2012 sebagai Ketua dengan judul "Egg Richal: Pemanfaatan Tepung Jeroan Teripang (Holothuria scabra) sebagai Pakan Altematif Sumber Kalsiurn pada Puyuh (Cortunix cortunix japonica) dan menjadi penerima beasiswa Peningkatan Prestasi Akademik (PPA)
tabun 2011-2014.
UCAPAN TERIMA KASm
Puji dan syukur penulis panjatkan atas kasih dan karunia-Nya sehingga penulis dapat menyelesaikan skripsi ini. Penulis mengucapkan terima kasih kepada Dr. Sri Suharti, SPt MSi selaku dosen pembimbing skripsi pertama dan kepada Prof. Dr. Ir. Dewi Apri Astuti, MS selaku dosen pembimbing akademik sekaligus dosen pembimbing skipsi kedua atas bimbingan, motivasi, nasehat, saran dan dukungannya selama pelaksanaan penelitian hingga penyelesaian skripsi. Penulis mengucapkan terima kasih kepada Dr Ir M. Ridla, MAgr sebagai dosen pembabas seminar hasil pada tanggal 23 Oktober 2014 dan kepada Dr Indab Wijayanti STp MSi serta Dr Irma Isnatia Arief SPt MSi selaku dosen penguji siding pada tanggal 31 Maret 2015. Ucapan terimakasih juga penulis sampaikan kepada Penelitian Prioritas Nasional Masterplan Percepatan dan Perluasan Pembangunan Ekonomi Indonesia 2013-2014 (pERP!NAS MP3El 2013-2014) atas dana yang telab diberikan pada penelitian ini.
Ucapan terima kasih penulis sarnpaikan kepada Mama, Bapak, Bang Andre, Adek Tinos dan Rizki yang senantiasa menyemangati, mendoakan dan menguatkan mulai selama penelitian hingga penulisan skripsi ini selesai. Terima kasih kepada Ibu Dian Anggraeni dan Pak Bain atas bantuan dan dukungan selama penelitian. Terima kasih kepada satu tim penelitian yaitu Dinar dan Hanab atas keIjasama dan bantuannya selama penelitian Terima kasih juga penulis ucapkan kepada keluarga besar D.NET (!NTP 47), Kopral 47 serta Kosan Sinar Kencana atas dukungannya. , Z\G セLN ⦅@
,·;ti;",
-' .:) • . 0 C I... -, . 0'1>,
|セ M ' セセ i@
|M [セ@ 'LJ.
• • II
セN iBu B@... .i' .
..Ji