Keanekaragaman Polen Dari Beberapa Spesies Stingless Bee Pada Perkebunan Kelapa Sawit Dan Karet
Teks penuh
Gambar

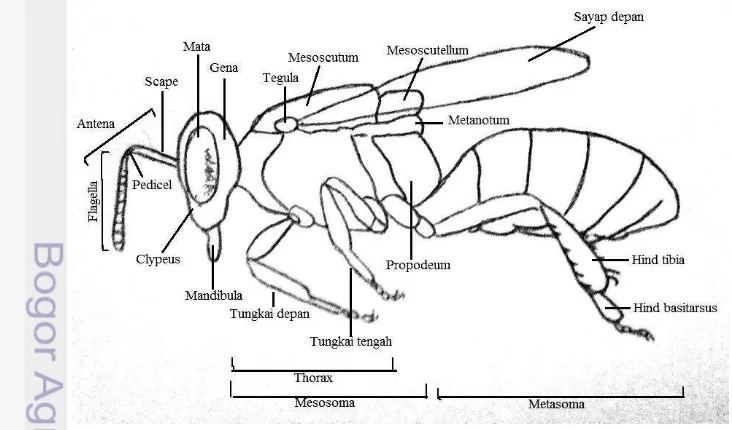

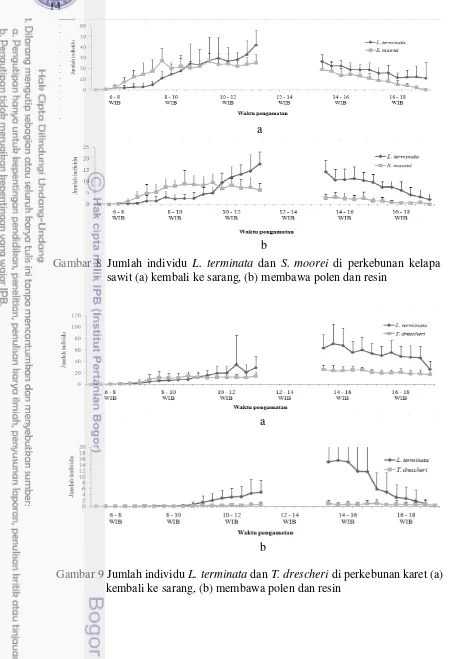

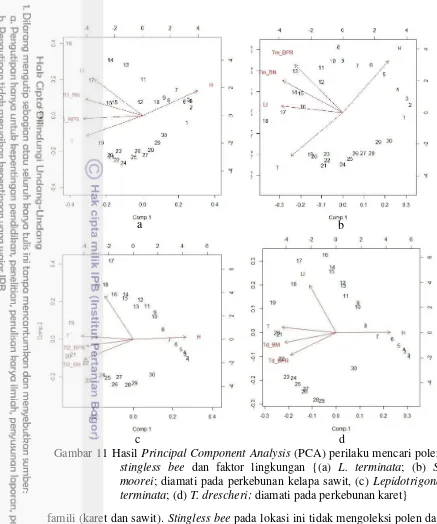

Dokumen terkait
Lima isolat bakteri nitritasi asal lahan perkebunan kelapa sawit dan karet dengan aktivitas penurunan konsentrasi amonium tertinggi Lima isolat bakteri nitratasi asal lahan
Pada perkebunan FELDA Gunung Besout 6 hanya didapatkan 6 famili dengan 14 spesies dari Hymenoptera parasitoid (Hanysyam et al. Hal ini membuktikan bahwa ekosistem pada
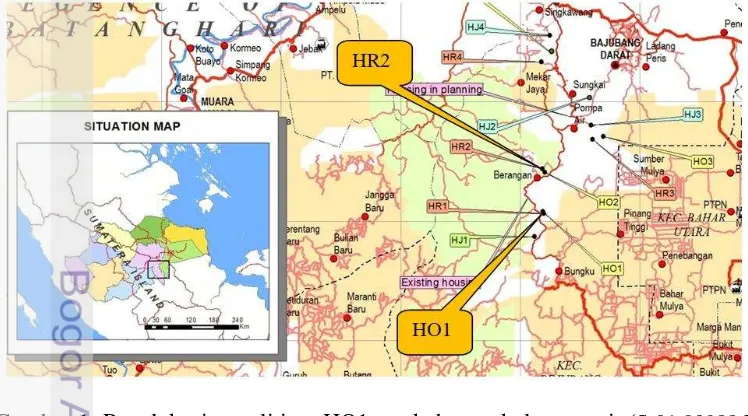
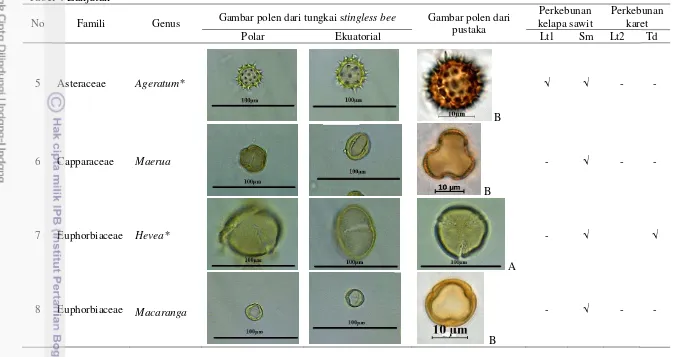
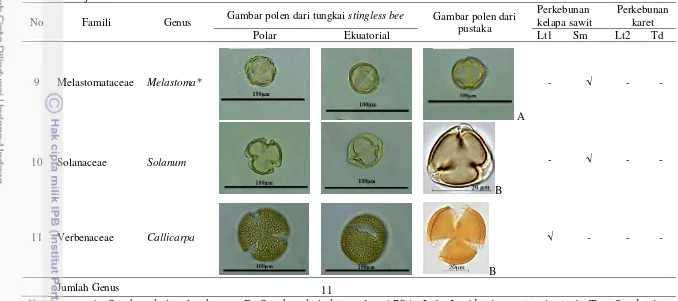
Penelitian ini meliputi beberapa tahap, yaitu pelepasan dan penghitungan jumlah polen kelapa sawit yang menempel pada tubuh kumbang betina, penyusunan peta
Bagi pemilik perkebunan tumbuhan bawah pada lahan kelapa sawit menimbulkan dampak negatif terhadap perkembangan kelapa sawit baik pada tanah gambut maupun tanah
Kemungkinan terdapat keanekaragaman tinggi pada perkebunan kelapa sawit yang berbatasan langsung atau dekat dengan hutan, kemudian semakin jauh dari hutan maka
KESIMPULAN Adapun kesimpulan dari penelitian ini yaitu komposisi gulma pada lahan perkebunan kelapa sawit terdapat 13 famili, 20 spesies, dan 7590 individu gulma, frekuensi gulma
Terbatasnya lahan sebagai sumber hijauan, maka pola pengembangan usaha kambing dapat dilakukan melalui pendekatan integrasi dengan sistem usaha perkebunan (kelapa sawit/karet)
Hasil penelitian ini dalam alih komoditi tanam dari karet ke sawit adalah 1 Alih komoditi terjadi karena perkebunan karet yang sudah mulai tua serta hasil dari karet sudah mulai
