ENKAPSULASI BAKTERI KITINOLITIK PADA BENIH CABAI UNTUK MENGHAMBAT SERANGAN Sclerotium rolfsii
TESIS
Oleh
NETTI IRAWATI 117030036/ BIO
PROGRAM PASCASARJANA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM UNIVERSITAS SUMATERA UTARA
ENKAPSULASI BAKTERI KITINOLITIK PADA BENIH CABAI UNTUK MENGHAMBAT SERANGAN Sclerotium rolfsii
TESIS
Diajukan sebagai salah satu syarat untuk memperoleh gelar Magister Sains dalam Progrm Studi Magister Ilmu Biologi pada Program Pascasarjana
Fakultas MIPA Universitas Sumatera Utara
Oleh
NETTI IRAWATI/BIO 117030036/ BIO
PROGRAM PASCASARJANA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM UNIVERSITAS SUMATERA UTARA
PENGESAHAN TESIS
Judul Tesis : ENKAPSULASI BAKTERI
KITINOLITIK PADA BENIH CABAI UNTUK MENGHAMBAT SERANGAN Sclerotium rolfsii
Nama Mahasiswa : NETTI IRAWATI
Nomor Induk Mahasiswa : 117030036 Program Studi : Magister Biologi
Fakultas : Matematika dan Ilmu Pengetahuan Alam Universitas Sumatera Utara
Menyetujui Komisi Pembimbing
Pembimbing I
Prof. Dr. Dwi Suryanto, M.Sc. NIP. 1964 0409 199403 1 003
Pembimbing II
Prof. Dr.Erman Munir, M.Sc. NIP. 1965 1101 199103 1 002
Ketua Program Studi
Prof. Dr. Syafruddin Ilyas, M.Biomed. NIP. 1966 0209 199203 1 003
Dekan
PERNYATAAN ORISINALITAS
ENKAPSULASI BAKTERI KITINOLITIK PADA BENIH CABAI UNTUK MENGHAMBAT SERANGAN Sclerotium rolfsii
TESIS
Dengan ini saya nyatakan bahwa saya mengakui semua karya tesis ini adalah hasil kerja saya sendiri kecuali kutipan dan ringkasan yang tiap satunya telah dijelaskan sumbernya dengan benar.
Medan, Agustus 2013
PERNYATAAN PERSETUJUAN PUBLIKASI KARYA ILMIAH UNTUK KEPENTINGAN AKADEMIS
Sebagai sivitas akademika Universitas Sumatera Utara, saya yang bertanda tangan di bawah ini:
Nama : Netti Irawati
NIM : 117030036
Program Studi : Biologi Jenis Karya Ilmiah : Tesis
demi pengembangan ilmu pengetahuan, menyetujui untuk memberikan kepada Universitas Sumatera Utara Hak Bebas Royalti Non-Eksklusif (Non-Exclusive Royalty Free Right) atas tesis saya yang berjudul:
ENKAPSULASI BAKTERI KITINOLITIK PADA BENIH CABAI UNTUK MENGHAMBAT SERANGAN Sclerotium rolfsii
beserta perangkat yang ada (jika diperlukan). Dengan hak bebas Royalti noneksklusif ini, Universitas Sumatera Utara berhak menyimpan, mengalih media, memformat, mengelola dalam bentuk data base, merawat dan mempublikasikan tesis saya tanpa meminta izin dari saya selama tetap mencantumkan nama saya sebagai penulis dan sebagai pemegang dan atau sebagai pemilik hak cipta.
Demikian surat pernyataan ini dibuat dengan sebenarnya.
Medan, 28 Agustus 2013
Telah diuji pada
Tanggal : 28 Agustus 2013
PANITIA PENGUJI TESIS
Ketua : Prof. Dr. Dwi Suryanto, M.Sc Anggota : 1. Prof. Dr. Erman Munir, M.Sc
RIWAYAT HIDUP
DATA PRIBADI
Nama lengkap berikut gelar : Netti Irawati, S.Si
Tempat dan Tanggal lahir : Karang Tengah, 27 Maret 1985 Alamat Rumah : Jalan Raya Menteng Gg. Arhalim Telepon/ Faks/ HP : 08566312504
e-mail : netti_sept@yahoo.com
Instansi Tempat Bekerja : IFDS Siti Hajar
Alamat Kantor : Jln. Djamin Ginting km 11 Medan Telepon/ Faks/HP : 061-77812577
DATA PENDIDIKAN
KATA PENGANTAR
Puji dan syukur penulis panjatkan ke hadirat Allah SWT yang telah melimpahkan rahmat dan karunia Nya, serta shalawat beriring salam penulis sampaikan kepada Nabi Muhammad SAW, sehingga penulis dapat menyelesaikan penulisan tesis yang berjudul ”Enkapsulasi Bakteri Kitinolitik pada Benih Cabai untuk Menghambat Serangan Sclerotium rolfsii”, sebagai salah satu syarat untuk meraih gelar Magister Sains di Program pascasarjana Biologi Fakultas Matematika dan Ilmu Pengetahuan Alam Universitas Sumatera Utara Medan.
Dalam kesempatan ini penulis mengucapkan terima kasih yang sebesar-besarnya kepada: Bapak Prof. Dr. Dwi Suryanto, M.Sc sebagai Dosen Pembimbing 1 dan Bapak Prof. Dr. Erman Munir, M.Sc selaku Dosen Pembimbing 2, Ibu Dr. It Jamilah M.Sc dan Ibu Dr. Suci Rahayu, M.Si selaku Dosen Penguji yang telah memberikan banyak saran dan arahan dalam penulisan tesis ini. Bapak Ketua Departemen Pascasarjana Biologi, Bapak Prof. Dr. Syafruddin Ilyas, M. Biomed dan seluruh staff pengajar dan pegawai di Pascasarjana Biologi.
Terima kasih kepada DITLITABMAS DIKTI yang telah mendanai penelitian ini melalui Hibah Unggulan Perguruan Tinggi Universitas Sumatera Utara. Terima kasih kepada Sekretaris Jenderal Pendidikan Tinggi (SEKJEN DIKTI) yang memberikan Beasiswa Unggulan. Terima kasih disampaikan kepada Laboratorium Mikrobiologi dan Laboratorium Sentral Departemen Biologi Fakultas Matematika dan Ilmu Pengetahuan Alam beserta Compost Center Universitas Sumatera Utara yang telah memberikan fasilitas selama penelitian ini berlangsung.
ENKAPSULASI BAKTERI KITINOLITIK PADA BENIH CABAI UNTUK MENGHAMBAT SERANGAN Sclerotium rolfsii
ABSTRAK
Penelitian mengenai enkapsulasi bakteri kitinolitik dengan menggunakan beberapa matriks seperti alginat, tapioka, carboksimetilselulosa dan gum arabik untuk menghambat serangan Sclerotium rolfsii pada benih cabai merah telah dilakukan. Enam isolat bakteri kitinolitik diujikan secara in vitro untuk mengetahui kemampuannya dalam menghambat pertumbuhan S. rolfsii jamur penyebab rebah kecambah pada cabai merah. Uji viabilitas sel bakteri pada benih yang terlapis dengan enkapsulasi bakteri kitinolitik dilakukan dengan menghitung sel hidup melalui cawan sebar. Uji kemampuan bakteri yang terenkapsulasi dilakukan dengan menanam benih terenkapsulasi pada media tanam yang sudah diinokulasi dengan konidia S. rolfsii. Bacillus sp. BK17 dan Enterobacter sp. PB17 menunjukkan kemampuan yang lebih baik dalam menghambat pertumbuhan S. rolfsii sebesar 11,40 mm dan sebesar 2,18 mm. Tingkat serangan S. rofsii pada benih cabai merah mencapai 64,71% dan mengalami penurunan sebesar 47,06% - 50% ketika benih direndam terlebih dahulu dalam larutan Enterobacter sp. PB17 dan Bacillus sp. BK17 sebelum melakukan persemaian. Enkapsulasi Bacillus sp. BK17 dengan menggunakan gum arabik menunjukkan viabilitas sel yang lebih baik dibandingkan dengan yang lainnya. Penanaman benih yang sudah dienkapsulasi dengan Bacillus sp. BK17 dan Enterobacter sp. PB17 pada rumah kasa menunjukkan adanya penurunan rebah pada kecambah cabai merah sebesar 100% dan 80%.
ENCAPSULATION OF CHITINOLYTIC BACTERIA IN CHILLI SEED TO INHIBIT ATTACK OF Sclerotium rolfsii
ABSTRACT
A study of the encapsulation of chitinolytic bacteria in alginate, tapioca, carboxymethylcellulose and gum arabic in coated chilli seeds to inhibit Sclerotium rolfsii growth has been done. Six isolates of chitinolytic bacteria were examined in vitro to know their ability to inhibit the growth of S. rolfsii, a causal agent of damping-off on red chillies. Examination of bacterial viability on coated seeds was done by using spread plate method. Examination ability of encapsulated bacteria was done by planting the seeds in the growing media inoculated with conidia of S. rolfsii. Bacillus sp. BK17 and Enterobacter PB17 showed the higher inhibition rate to inhibit the growth of S. rolfsii with inhibition zone of 11.40 mm and 2, 18 mm, respectively. Chitinolytic bacteria treatment could reduce damping-off by 47,61-50% compared to that of (+) control which causing damping-damping-off by 63,71%. Encapsulated Bacillus Sp. BK17 with gum arabic showed higher cell viability compared to that of others. Coated seeds with encapsulated Bacillus sp. BK17 and Enterobacter sp. PB17 showed to decrease damping-off in vitro by 100% and 80 %.
DAFTAR ISI
Halaman
LEMBAR PENGESAHAN i
PERNYATAAN ORISINALITAS ii
PERNYATAAN PERSETUJUAN PUBLIKASI KARYA ILMIAH
UNTUK KEPENTINGAN AKADEMIS iii
RIWAYAT HIDUP v
KATA PENGANTAR vi
ABSTRAK viii
ABSTRACT ix
DAFTAR ISI x
DAFTAR TABEL xii
DAFTAR GAMBAR xiii
DAFTAR LAMPIRAN xv
BAB I PENDAHULUAN 1
1.1. Latar Belakang 1
1.2. Perumusan Masalah 3
1.3. Tujuan Penelitian 3
1.4. Hipotesis 4
1.5. Manfaat Penelitian 4
BAB 2 TINJAUAN PUSTAKA 5
2.1. 2.2. 2.3. 2.4. 2.5.
Karakteristik Tanaman Cabai
Penyakit Rebah Kecambah (Damping-off) pada Tanaman Cabai
Faktor yang Mempengaruhi Pertumbuhan Jamur Potensi Bakteri sebagai Agen Biokontrol
Enkapsulasi a. Alginat
b. Karboksimetil Selulosa (CMC) c. Tapioka
d. Gum Arabik
5 6 8 9 11 12 13 14 16
BAB 3 BAHAN DAN METODE 18
3.1. 3.2.
Waktu dan Tempat Alat dan Bahan
3.3. 3.4. 3.5. 3.6. 3.7. 3.8. 3.9. 3.10. Isolat
Uji Antagonis Bakteri Kitinolitik In Vitro Pengamatan Struktur Hifa Jamur Abnormal Asai Patogenitas S. rolfsii terhadap Benih Cabai Preparasi Suspensi Isolat Bakteri
Enkapsulasi Benih Cabai Merah
Asai Viabilitas Bakteri Kitinolitik dalam Enkapsulasi Asai Patogenitas S. rolfsii terhadap Benih Cabai Terenkapsulasi di Rumah Kasa
19 19 19 20 20 21 22 23
BAB 4 HASIL DAN PEMBAHASAN 24
4.1. 4.2. 4.3. 4.4. 4.5.
Kemampuan Antagonis Bakteri Kitinolitik secara In Vitro
Abnormalitas Hifa Jamur S. rolfsii setelah Uji Antagonis dengan Bakteri Kitinolitik
Patogenitas S. rolfsii pada Benih Cabai
Viabilitas Bakteri Kitinolitik dalam Enkapsulasi Patogenitas S. rolfsii terhadap Benih Cabai Terenkapsulasi 24 26 28 30 32
BAB 5 KESIMPULAN DAN SARAN 36
5.1. 5.2. Kesimpulan Saran 36 36
DAFTAR PUSTAKA 37
DAFTAR TABEL
Nomor Tabel
Judul Halaman
1 Kombinasi perlakuan enkapsulasi benih dengan 4 matriks Pembawa
22
2 Penghambatan pertumbuhan jamur S. rolfsii oleh bakteri kitinolitik
24
DAFTAR GAMBAR
Nomor Gambar
Judul Halaman
1 2 3 4 5 6 7 8 9 10 11 12
Gejala serangan Sclerotium rolfsii adanya miselium pada batang tua
Struktur natrium alginat
Struktur CMC
Struktur penyusun pati
Struktur penyusun gum arabik
Penghambatan pertumbuhan miselium jamur S. rolfsii pada media MGMK + yeast
Perubahan morfologi hifa jamur S. rolfsii setelah uji antagonis dengan bakteri antagonis pada media MGMK yeast (a). hifa normal, (b). hifa menggulung pada bagian ujung hifa, (c). hifa lisis pada bagian dinding hifa, (d).hifa bengkok, (e). isi hifa lisis, (f). hifa putus
Persentase rebah kecambah pada benih cabai (BK17 = benih cabai + Bacillus sp. BK17, PB17 = Benih cabai + Enterobacter sp. PB17)
Benih cabai yang normal (a) dan yang terserang S. rolfsii (b) setelah 30 hari 1. Hifa S. rolfsii pada pangkal batang, 2. Batang yang menguning dengan daun yang sudah habis berguguran.
Viabilitas sel bakteri pada benih terenkapsulasi selama penyimpanan
DAFTAR LAMPIRAN
Nomor Lampiran
Judul Halaman
1 2 3 4
Alur kerja asai antagonisme in vitro Alur kerja preparasi suspesi isolat bakteri Komposisi medium
Foto penelitian
46 47 48 48
ENKAPSULASI BAKTERI KITINOLITIK PADA BENIH CABAI UNTUK MENGHAMBAT SERANGAN Sclerotium rolfsii
ABSTRAK
Penelitian mengenai enkapsulasi bakteri kitinolitik dengan menggunakan beberapa matriks seperti alginat, tapioka, carboksimetilselulosa dan gum arabik untuk menghambat serangan Sclerotium rolfsii pada benih cabai merah telah dilakukan. Enam isolat bakteri kitinolitik diujikan secara in vitro untuk mengetahui kemampuannya dalam menghambat pertumbuhan S. rolfsii jamur penyebab rebah kecambah pada cabai merah. Uji viabilitas sel bakteri pada benih yang terlapis dengan enkapsulasi bakteri kitinolitik dilakukan dengan menghitung sel hidup melalui cawan sebar. Uji kemampuan bakteri yang terenkapsulasi dilakukan dengan menanam benih terenkapsulasi pada media tanam yang sudah diinokulasi dengan konidia S. rolfsii. Bacillus sp. BK17 dan Enterobacter sp. PB17 menunjukkan kemampuan yang lebih baik dalam menghambat pertumbuhan S. rolfsii sebesar 11,40 mm dan sebesar 2,18 mm. Tingkat serangan S. rofsii pada benih cabai merah mencapai 64,71% dan mengalami penurunan sebesar 47,06% - 50% ketika benih direndam terlebih dahulu dalam larutan Enterobacter sp. PB17 dan Bacillus sp. BK17 sebelum melakukan persemaian. Enkapsulasi Bacillus sp. BK17 dengan menggunakan gum arabik menunjukkan viabilitas sel yang lebih baik dibandingkan dengan yang lainnya. Penanaman benih yang sudah dienkapsulasi dengan Bacillus sp. BK17 dan Enterobacter sp. PB17 pada rumah kasa menunjukkan adanya penurunan rebah pada kecambah cabai merah sebesar 100% dan 80%.
ENCAPSULATION OF CHITINOLYTIC BACTERIA IN CHILLI SEED TO INHIBIT ATTACK OF Sclerotium rolfsii
ABSTRACT
A study of the encapsulation of chitinolytic bacteria in alginate, tapioca, carboxymethylcellulose and gum arabic in coated chilli seeds to inhibit Sclerotium rolfsii growth has been done. Six isolates of chitinolytic bacteria were examined in vitro to know their ability to inhibit the growth of S. rolfsii, a causal agent of damping-off on red chillies. Examination of bacterial viability on coated seeds was done by using spread plate method. Examination ability of encapsulated bacteria was done by planting the seeds in the growing media inoculated with conidia of S. rolfsii. Bacillus sp. BK17 and Enterobacter PB17 showed the higher inhibition rate to inhibit the growth of S. rolfsii with inhibition zone of 11.40 mm and 2, 18 mm, respectively. Chitinolytic bacteria treatment could reduce damping-off by 47,61-50% compared to that of (+) control which causing damping-damping-off by 63,71%. Encapsulated Bacillus Sp. BK17 with gum arabic showed higher cell viability compared to that of others. Coated seeds with encapsulated Bacillus sp. BK17 and Enterobacter sp. PB17 showed to decrease damping-off in vitro by 100% and 80 %.
BAB 1
PENDAHULUAN
1.1. Latar Belakang
Cabai merah (Capsicum annuum L.) merupakan tanaman sayuran yang sangat penting di Indonesia. Hal ini ditunjukkan dengan areal pertanaman cabai merah yang terluas diantara tanaman sayuran. Pada tahun 2004, luas panen cabai mencapai 194.588 ha dan produksinya mencapai 1.100.514 ton. Produktivitas cabai merah di Indonesia sekitar 5.66 ton/ha. Produktivitas ini jauh lebih rendah dibandingkan potensinya yaitu 12-15 ton/ha (Departemen Pertanian, 2004). Penggunaan benih bermutu rendah dan infeksi penyakit merupakan penyebab utama rendahnya produktivitas cabai tersebut. Penurunan hasil panen di lapangan akibat penyakit rebah kecambah pada musim hujan cukup tinggi yaitu mencapai 80%, sedangkan pada musim kemarau sekitar 20-30% (Widodo, 2007). Di Kabupaten Garut misalnya penurunan hasil panen akibat rebah kecambah mencapai 60-70% (Oktaviane, 2013).
lainnya seperti sambiloto (Hartati et al., 2008), famili Solanaceae (Kim dan Weon, 2003), tanaman bayam (Cumming, 2009), tomat (Yusniawaty, 2009) dan kedelai (Malinda et al., 2012).
Tanaman cabai adalah tanaman yang tidak ditanam langsung di lahan tetapi harus lebih dahulu disemai, setelah kecambah berumur 7-12 hari bibit dipindah ke lapangan. Menurut Setiadi (2004) persemaian benih memiliki peranan yang penting dalam penentuan hasil panen. Sehingga kualitas persemaian cabai perlu diperhatikan agar hasil panen sesuai dengan yang diharapkan. Hingga saat ini, varietas cabai komersial berdaya hasil tinggi dan tahan terhadap penyakit rebah kecambah masih belum ada. Umumnya spesies cabai yang terserang rebah kecambah akan mati sebelum muncul ke permukaan tanah.
Pada umumnya pengendalian penyakit rebah kecambah masih bergantung kepada fungisida, namun karena fungisida memberikan dampak yang negatif bagi lingkungan, para peneliti berupaya mencari alternatif lain yang bisa digunakan untuk pengendalikan rebah kecambah tanpa mengganggu keseimbangan lingkungan diantaranya: penggunaan ekstrak tanaman seperti rimpang jahe, umbi bawang putih, daun alamanda, daun neem, dan biji kalijira (Islam dan Faruq, 2012) dan menggunakan agen biokontrol seperti Trichoderma harzianum dan Trichoderma virens (Jinantana dan Sariah, 1998; Istikorini, 2006; Mukarlina et
al., 2010), dan Pseudomonas putida (Boer et al., 2003). Penggunaan mikroorganisme sebagai agen pengendali penyakit rebah kecambah bisa secara langsung (Seikh et al., 2006; Suprapta, 2012) maupun dengan cara enkapsulasi (Bashan, 1986).
(Kim et al., 1996), Lactobacillus plantarum terenkapsulasi susu skim dan gum arab (Rizqiati et al., 2009) dan Methylobacterium spp. terenkapsulasi beberapa komposisi bahan pelapis (alginat, gum arabik) (Eka, 2009). Enkapsulasi dengan menambahkan subtansi prebiotik dalam produk merupakan salah satu faktor yang dapat digunakan untuk meningkatkan viabilitas organisme pada produk (Kneifel et al., 1993). Dalam penelitian ini perlu dilakukan penelitian mengenai potensi enkapsulasi bahan pelapis alginat, CMC, tapioka dan gum arab terhadap benih cabai merah dengan memanfaatkan isolat bakteri kitinolitik yang potensial sebagai pengendali hayati jamur patogen.
1.2. Perumusan Masalah
Penggunaan fungisida yang tidak bijaksana dapat menimbulkan masalah pencemaran lingkungan, gangguan keseimbangan ekologis dan residu yang ditinggalkannya dapat bersifat racun dan karsinogenik. Sehingga perlu segera diupayakan pengurangan penggunaan fungisida kimiawi dan mengalihkannya pada jenis fungisida yang aman bagi lingkungan, yakni dengan cara pengendalian hayati dengan menggunakan mikroorganisme. Namun kualitas penggunaan mikroorganisme perlu diperhatikan seperti membuat formulasi baru dalam peningkatan efektifitas mikroorganisme sebagai pengendali hayati. Salah satu alternatif formulasi yang digunakan adalah dengan enkapsulasi benih. Benih yang terenkapsulasi diharapkan memiliki kemampuan yang tinggi dalam pengendalian hayati untuk itu perlu dilakukan pengujian bahan enkapsulasi yang baik untuk melapisi benih cabai merah dan bakteri kitinolitik.
1.3. Tujuan Penelitian
Penelitian ini bertujuan untuk mengetahui:
1. Kemampuan bakteri kitinolitik dalam menghambat pertumbuhan jamur S. rolfsii pada benih cabai.
3. Viabilitas bakteri kitinolitik dan benih terenkapsulasi bakteri kitinolitik dengan beberapa pembawa.
4. Kemampuan bakteri kitinolitik terenkapsulasi dalam menghambat pertumbuhan jamur S. rolfsii.
1.4. Hipotesis
1. Bakteri kitinolitik mampu menghambat pertumbuhan jamur S. rolfsii pada benih cabai.
2. Viabilitas bakteri kitinolitik dan benih terenkapsulasi bakteri kitinolitik pada bahan alginat, CMC, tapioka dan gum arabik berbeda.
3. Bakteri kitinolitik dalam bentuk enkapsulasi pada benih cabai merah mampu menghambat pertumbuhan jamur S. rolfsii.
1.5. Manfaat Penelitian
BAB 2
TINJAUAN PUSTAKA
2.1 Karakteristik Tanaman Cabai
Cabai merupakan tanaman perdu dari famili terung-terungan (Solanaceae) yang diduga memiliki 90 genus dan sekitar 2.000 spesies yang terdiri dari tumbuhan herba, semak, dan tumbuhan kerdil lainnya. Tanaman cabai sendiri diperkirakan ada sekitar 20 spesies yang sebagian besar tumbuh di tempat asalnya Amerika. Di antaranya yang sudah dimanfaatkan baru beberapa spesies saja, yaitu cabai besar (Capsicum annuum), cabai kecil (Capsicum frustescens), Capsicum baccatum, Capsicum pubescens, dan Capsicum chinense (Setiadi, 2004). Beberapa sifat tanaman cabai yang dapat digunakan untuk membedakan antar varietas di antaranya adalah percabangan tanaman, pembungaan tanaman, ukuran ruas, dan tipe buahnya (Prajnanta, 1999). Bunga pada tanaman cabai terdapat pada ruas batang dan jumlahnya bervariasi antara 1-8 bunga tiap ruas tergantung pada spesiesnya. Cabai besar (C. Annuum) mempunyai satu bunga tiap ruas, sedangkan cabai rawit (C. frutescens) mempunyai 1-3 bunga tiap ruas. Ukuran ruas tanaman cabai bervariasi dari pendek sampai panjang. Makin banyak ruas makin banyak jumlah bunganya, dan diharapkan semakin banyak pula produksi buahnya. Buah cabai bervariasi antara lain dalam bentuk, ukuran, warna, tebal kulit, jumlah rongga, permukaan kulit dan tingkat kepedasannya.
dan Sulawesi, sedangkan cabai merah keriting buahnya bergelombang atau keriting, ramping, kulit buah tipis, berumur lebih lama, lebih tahan simpan, dan rasanya pedas. Tipe ini banyak diusahakan di Jawa Barat dan Sumatera. Cabai paprika buahnya berbentuk segi empat panjang dan biasa dipanen saat matang hijau (Semangun, 2007). Umur cabai sangat bervariasi tergantung jenis cabai. Tanaman cabai besar dan keriting yang ditanam di dataran rendah sudah dapat dipanen pertama kali umur 70 –75 hari setelah tanam, sedangkan waktu panen di dataran tinggi lebih lambat yaitu sekitar 4 – 5 bulan setelah tanam. Panen dapat terus-menerus dilakukan sampai tanaman berumur 6 – 7 bulan. Pemanenan dapat dilakukan dalam 3 – 4 hari sekali atau paling lama satu minggu sekali (Setiadi, 2004).
Tanaman cabai akan tumbuh baik pada lahan dataran rendah yang tanahnya gembur dan kaya bahan organik, tekstur ringan sampai sedang, pH tanah berkisar antara 5.5 – 6.8, drainase baik dan cukup tersedia unsur hara bagi pertumbuhannya. Kisaran suhu optimum bagi pertumbuhannya adalah 18 – 30 oC Secara geografis tanaman cabai dapat tumbuh pada ketinggian 0 – 1200 m di atas permukaan laut. Pada dataran tinggi yang berkabut dan kelembabannya tinggi, tanaman cabai mudah terserang penyakit. Cabai akan tumbuh baik pada daerah yang rata-rata curah hujan tahunannya antara 600–1250 mm dengan bulan kering 3–8,5 bulan dan pada tingkat penyinaran matahari lebih dari 45 % .
2.2. Penyakit Rebah Kecambah (Damping-off) pada Tanaman Cabai
Rebah kecambah (damping–off) sering terjadi di persemaian cabai maupun terung. Biji dapat membusuk di dalam tanah, atau semai-semai dapat mati sebelum muncul ke permukaan tanah. Batang semai muda yang masih lunak jika terserang rebah kecambah pangkalnya menjadi basah dan mengkerut sehingga menyebabkan semai rebah dan mati (Semangun, 2007).
dilaporkan bahwa penyakit rebah kecambah juga bisa disebabkan oleh jamur patogen tanaman diantaranya dari genus Phytium, Rhizoctonia dan Sclerotium (Singh, 1998; Agrios, 1997). S. rolfsii merupakan salah satu jamur patogen yang dapat menyebabkan rebah kecambah pada tanaman cabai (Semangun, 2007; Lamidi, 1986; Dange, 2006; Yusniawaty, 2009). Jamur ini merupakan jamur tular tanah yang dapat bertahan lama dalam bentuk sklerotia di dalam tanah, pupuk kandang, dan sisa-sisa tanaman sakit. Di samping itu, jamur tersebut dapat menyebar melalui air irigasi dan benih pada lahan yang ditanami secara terus menerus dengan tanaman inang yang sama, sehingga mengakibatkan turunnya produksi tanaman yang akan dipanen (Timper et al., 2001).
Jamur S. rolfsii menyebabkan gejala rebah kecambah pada persemaian dan busuk batang pada tanaman inang yang lebih tua. Apabila tanaman muda yang terserang maka akan cepat mati, tetapi bila tanaman yang lebih tua terserang, pucuk tanaman menjadi kuning, layu dan kemudian mati. Akibatnya bagian dari batang dan akar pada batas tanah menjadi busuk dan sebagian ditutupi oleh bercak putih yaitu miselium yang berwarna putih (Gambar 1) sampai terbentuk sklerotia coklat (Agrios, 1997).
Gambar 1. Gejala serangan S. rolfsii adanya miselium pada batang tua (Thu et al., 2013)
tanaman tersebut dicabut akan terlihat benang-benang halus pada bagian yang sakit. Benang-benang ini berwarna putih dan padanya terdapat butiran yang mula-mula berwarna putih kemudian coklat dan akhirnya coklat tua. Butir ini disebut sklerotium yang sangat mudah terlepas dari benang-benangnya. Miselium putih yang terjalin mengelilingi jaringan tanaman yang terserang sering terlihat seperti jalinan benang halus pada bagian tanaman yang sakit berwarna coklat tua dan dikelilingi oleh benda-benda kecil yang bentuknya menyerupai biji lada, yang dihasilkan pada bagian permukan tanaman yang terinfeksi dan dekat permukaan tanah. Tanaman muda dapat dikelilingi sklerotia dan mati.
Sclerotium rolfsii termasuk dalam subdivisi Deuteromycotina, Kelas
Deuteromycetes, subkelas Hyphomycetidae dan ordo Agonomycetales (Alexopoules dan Mims, 1979). S. rolfsii adalah sejenis jamur yang mempunyai miselium yang terdiri dari benang-benang, berwarna putih, tersusun seperti bulu atau kipas. S. rolfsii tidak membentuk spora. Untuk pemencaran dan untuk mempertahankan diri jamur membentuk sejumlah sklerotium yang semula berwarna putih, kelak menjadi coklat. Butir-butir ini mudah sekali lepas dan terangkut oleh air. Sklerotium mempunyai kulit yang kuat sehingga tahan terhadap suhu tinggi dan kekeringan. Di dalam tanah sklerotium dapat bertahan sampai 6 - 7 tahun. Ukuran diameter sklerotia 0,05-2 mm dan bentuk perkecambahan sklerotia dispersif seperti kapas berwarna putih. Ukuran terkecil dari diameter koloni sclerotium adalah 0,61 cm dan ukuran terbesarnya 1,71 cm sedangkan untuk pengukuran kecepatan pertumbuhan miselium yang terlambat adalah 3,1 mm/hari dan yang tercepat adalah 8,54 mm/hari yang dapat dilihat pada hari kedua dan hari ketujuh penelitian (Magenda, 2011).
2.3. Faktor yang Mempengaruhi Pertumbuhan Jamur
adalah pada kelembaban relatif (Rh) 12% - 15%. Pertumbuhan optimum S. rolfsii pada suhu 27-30o C dan sklerotia tidak dapat bertahan pada suhu dibawah 0oC sehingga suhu dingin bisa membatasi penyebaran jamur ini (Singleton et al., 1992; Singh, 1998). Sklerotia yang dihasilkan memiliki kemampuan yang baik dalam beradaptasi dengan lingkungan sehingga penyebaran sklerotia di sekitar inang dapat menjangkau wilayah yang cukup luas. Tingkat pertumbuhan sklerotia pada tanah dipengaruhi oleh bentuk perkecambahan hifa, kerusakan jaringan tanaman, dan kelarutan nutrisi (Punja dan Gregon, 1981; Punja et al., 1985).
Pada media biakan, jamur dapat tumbuh pada pH 1.4 – 8.8 (Agrios, 1997), dengan suhu optimum pertumbuhan 30 - 35oC sedangkan suhu minimumnya adalah 8oC dan suhu maksiumnya 40oC. Hifa vegetatif tidak dapat bertahan pada suhu dibawah 0oC. Pada kondisi tersebut hifa akan mati dalam jangka waktu 24 jam. Sedangkan sklerotianya dapat tahan selama 48 jam pada suhu -10oC (Watkins, 1961). Miseliumnya mampu tumbuh dan menyerang biji-biji dalam tanah dengan kandungan air yang jauh lebih rendah dari pada kandungan air yang dibutuhkan untuk pertumbuhan biji (Epps et al., 1951). Keadaan suhu, kelembaban dan aerasi yang baik di atas permukaan tanah atau beberapa sentimeter dalam tanah dapat menguntungkan perkembangan miselia jamur dan akan meningkatkan patogenitas jamur. Jamur S. rolfsii memiliki dua fase pertumbuhan yang secara ekologi berbeda. Pertama adalah fase perkembangan miseliun yang berupa miselium putih dan tebal dan sering disebut sebagai jamur putih. Fase tersebut juga dikenal sebagai fase pertumbuhan atau fase patogenik dari jamur. Fase yang kedua adalah fase produksi sklerotia jamur yang memungkinkan jamur dapat bertahan pada keadaan yang tidak cocok (Boyle, 1961).
2.4. Potensi Bakteri sebagai Biokontrol
penyakit dengan menggunakan mikroorganisme sebagai agen biokontrol (Martin dan Hancock, 1987; Wijayanti, 2003; Harni, 2007). Mikroorganisme yang digunakan sebagai agen biokontrol adalah mikroorganisme yang mampu menggunakan dinding sel jamur patogen sebagai sumber nutrisinya. Dinding sel jamur terdiri dari glukan, selulosa, kitin, manosa, glukosa, asam amino dan lemak (Landecker, 1996). Salah satu pengembangan teknologi yang cocok untuk mengendalikan jamur patogen tanaman adalah pemanfaatan mikroba kitinolitik yang memiliki aktivitas kitinase. Mikroba kitinolitik mampu menghidrolisis senyawa kitin yang merupakan salah satu penyusun dinding sel kapang patogen. Terdegradasinya senyawa tersebut menyebabkan jamur patogen menjadi lemah atau mati. Dengan demikian mikroba kitinolitik berpotensi digunakan sebagai biofungisida untuk mengendalikan kapang patogen yang memiliki kitin sebagai struktur dinding selnya (Nildayanti, 2011). Bakteri kitinolitik merupakan mikroba yang memiliki kemampuan mendegradasi kitin karena menghasilkan enzim kitinase. Berbagai kelompok bakteri dilaporkan memiliki aktivitas kitinolitik diantaranya yaitu Pseudomonas fluorescens, Bacillus dan Streptomyces (Djatmiko et al., 2007; Papuangan, 2009). Bacillus apiarius (Muharni dan Hary, 2011) Serratia marcescens (Wijayanti, 2003).
Pengendali hayati patogen tanaman yaitu bakteri yang disebarkan melalui tanah sudah dipelajari sebagai suatu alternatif untuk mengendalikan jamur patogen tanaman. Beberapa bakteri yang dimanfaatkan sebagai biokontrol yaitu P. putida terhadap Fusarium oxysporum (Boer et al., 2003), P. fluorescens, Bacillus
Pemanfaatan bakteri kitinolitik sebagai pengendali jamur patogen tanaman diterapkan dengan cara enkapsulasi pada benih tanaman yang akan dijadikan target penelitian (Bashan, 1986; Schisler et al., 2004; Dawar et al., 2008; Hameeda et al.,2010; Suarez et al., 2011). Metode enkapsulasi ini diharapkan dapat meningkatkan ketahanan benih tanaman terhadap jamur patogen yang menyerang tanaman, sehingga pemanfaatan mikroorganisme sebagai pengendali hayati dapat lebih efektif dan berkesinambungan.
2.5. Enkapsulasi
Enkapsulasi adalah suatu proses pembungkusan suatu bahan atau campuran beberapa bahan dengan bahan lain. Bahan yang dibungkus atau bahan yang ditangkap biasanya berupa cairan, walaupun ada juga yang berbentuk partikel padat atau gas yang disebut sebagai bahan inti atau bahan aktif atau bahan internal, sedangkan bahan yang berfungsi sebagai pembungkus disebut sebagai dinding atau bahan pembawa atau membran. Proses enkapsulasi banyak digunakan untuk mempertahankan flavor, asam, lipida, enzim, mikroorganisme, pemanis buatan, vitamin, mineral, air, bahan pengembang, warna dan garam (Risch, 1995).
spray chilling, ekstruksi sentrifugal, pemisahan suspensi rotasional, dan kompleksasi inklusi.
Beberapa bahan yang bisa digunakan untuk enkapsulasi yaitu: a. Alginat
Alginat adalah zat koloidal hidrofilik yang diekstrasi dari ganggang laut Macrocystis pyrifera dan Ascophyllum nodosum yang bersifat biokompatibel dan
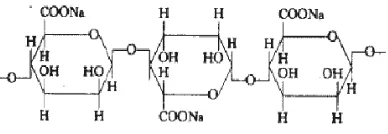
[image:30.612.221.415.372.439.2]biodegradabel terdiri dari β-D manuronat dan α–L guluronat yang dihubungkan dengan ikatan (1-4) dengan berbagai perbandingan (Colwell et al., 1985). Alginat yang tersedia secara komersial adalah dalam bentuk garamnya yaitu natrium alginat (Gambar 2). Keunikan natrium alginat yaitu perubahannya menjadi hidrogel dengan 95% molekul air di dalamnya, yang merupakan syarat penting untuk penggunaan dalam menjebak senyawa. Ketika natrium alginat bertemu dengan kation divalent seperti Ca+2 menghasilkan pembentukan gel dimana residu G dari alginat yang mengikat ion Ca+2 (Wang et al., 2006).
Gambar. 2. Struktur natrium alginat
et al., 2008), cabai dengan isolat Bacillus sp. (Suryanto et al., 2012), padi dengan
isolat Bacillus megaterium (Chumthong et al., 2008).
b. Karboksimetil Selulosa (CMC)
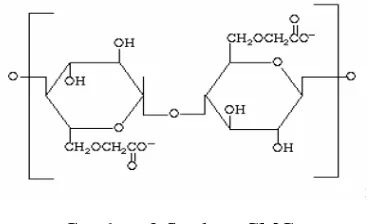
[image:31.612.230.415.361.473.2]Karboksimetil selulosa atau Carboxymethyl Cellulose (CMC) banyak digunakan pada berbagai industri seperti: detergen, cat, keramik, tekstil, kertas dan makanan. Fungsi CMC disini adalah sebagai pengental, penstabil emulsi atau suspensi dan bahan pengikat (Wijayani et al., 2005). CMC adalah polisakarida anionik linear yang larut dalam air dan merupakan gom alami yang dimodifikasi secara kimia. Bubuk CMC yang telah dimurnikan berwarna putih sampai krem, mengalir bebas, tidak berasa, dan tidak berbau (Yundhana, 2008). Fungsi dasar CMC adalah untuk mengikat air, menstabilkan komponen lain, dan mencegah pengerutan. Struktur CMC mempunyai kerangka D-glukopiranosa yang berikatan β- (1,4) dari polimer selulosa (Gambar 3).
Gambar. 3 Struktur CMC
Perbedaan cara membuat CMC mempengaruhi derajat substitusi, tetapi secara umum derajat substitusi berkisar dari 0.4 sampai 1.4 per unit monomer CMC diproduksi dengan cara mencampurkan selulosa dari pulp kayu atau kapas dengan larutan NaOH. Selulosa-alkali ini kemudian direaksikan dengan Namonokloroasetat atau asam monokloroasetat menghasilkan Na- CMC dan NaCl (Awalludin, 2004). Berbeda dengan turunan selulosa lainnya. CMC mengandung garam karboksil yang membuatnya lebih mudah larut dalam air.
dan cenderung mengendap. Stabilitas maksimum CMC terjadi pada pH 7 sampai 9. CMC dengan tingkat kemurnian tinggi yang dikenal sebagai gom selulosa, telah digunakan secara luas dalam bidang industri makanan dan farmasi. Dalam industri makanan, CMC digunakan sebagai pengental, pencegah pengerutan, dan pencegah pembentukan kristal es. Sifat CMC yang tidak larut dalam asam lambung, tetapi larut dalam cairan basa di usus, menyebabkan CMC digunakan untuk pembuatan tablet atau serbuk obat dengan cara salut enteric (Awalludin 2004).
Natrium CMC adalah garam dari asam karboksilat. Pada pH 3.0 atau lebih rendah, CMC akan kembali menjadi bentuk asam bebas tidak larut. Sifat yang paling berguna dari CMC adalah daya pengentalannya. Viskositas larutan hampir tidak terpengaruh pada pH 5−7; pada pH<3, viskositas mungkin meningkat dan pengendapan bentuk asam bebas dari CMC dapat terjadi; pada pH>10 terjadi sedikit penurunan viskositas. Viskositas larutan CMC menurun dengan meningkatnya suhu (Yundhana, 2008). Beberapa penelitian yang telah memanfaatkan CMC sebagai bahan enkapsulasi diantaranya pada benih padi dengan isolat P. fluorescens (Nandakumar et al., 2001), benih jagung dengan isolat B. substilis (Muis dan Arcadio, 2006).
c. Tapioka
(1:6) unit glukosa ini jumlahnya sangat sedikit dalam suatu molekul pati, berkisar antara 4−5%. Namun, jumlah molekul dengan rantai cabang, yaitu amilopektin, sangat banyak dengan DP berkisar antara 105 dan 3x106 unit glukosa (Jacobs dan Delcour, 1998).
[image:33.612.176.466.394.477.2]Granula pati tapioka berbentuk semi bulat sampai bulat dengan salah satu dari bagian ujungnya mengerucut dengan ukuran 5-35 mm. Suhu gelatinisasi berkisar antara 52-64 0C, kristalinitas 38%, kekuatan pembengkakan sebesar 42 dan kelarutan 31%. Kekuatan pembengkakan dan kelarutan tapioka lebih kecil dari pati kentang, tetapi lebih besar dari pati jagung (Rickard et al., 1991). Suhu gelatinisasi pati tapioka berada di antara pati jagung waxy dan pati jagung reguler. Viskositasnya lebih rendah dibandingkan dengan pati jagung waxy. Apabila dilakukan pendinginan, larutan pati mengalami retrogradasi untuk menghasilkan suatu gel yang halus. Larutan pati tapioka lebih jernih dibandingkan pati-pati native yang lainnya (National Starch, 2005).
Gambar. 4. Struktur penyusun pati
Penggunaan tapioka sebagai bahan enkapsulasi telah dilakukan oleh beberapa peneliti diantaranya Azospirillum brasilense sebagai pupuk organik (Wijayanti, 2010). Benih cabai terhadap F. oxysporum (Suryanto et al., 2012). Enkapsulasi terhadap P. fluorescens dan B. putida (Charpentier et al., 1998).
d. Gum Arabik
Gum arabik telah digunakan selama setidaknya 4.000 tahun (FAO, 1999). Substansi gum Arabik merupakan suatu metabolit sekunder yang dihasilkan olah tanaman yang berasal dari spesies Acacia senegal dan tanaman lainnya yang masih dekat kekerabatannya. Pohon Acacia senegal banyak ditemukan di Sudan, Senegal, Nigeria dan daerah semiarid. Eksudat kering tidak berbau, tidak berwarna dan hambar, dan sering teerjadi sebagai respons terhadap infeksi pada bagian pohon yang luka. Larut dalam air walaupun dalam air dingin dan air panas dengan konsentrasi mencapai 50 %. Viskositas larutan bervariasi tergantung tipe gum arabik, pH dan ikatan ionik. pH memiliki peranan penting dalam memodifiksi viskositas gum arabik. pH < 5 dan pH > 9,5 menyebabkan pengurangan viskositas ion arabates. Namun pH yang mendekati pH netral dapat meningkatkan viskositas gum arabik (Mahmoud, 2000). Viskositas maksimum dicapai antara pH 6 dan 7. Gum arabik digunakan sebagai koloid protektif dan emulsifier.
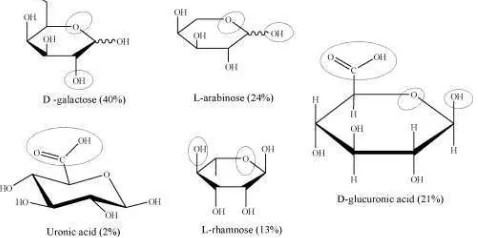
Gum arabik adalah campuran kompleks dari oligosakarida arabinogalactan, polisakarida, glikoprotein. Strukur kimianya bersifat netral atau sedikit asam pada rantai cabang. Komposisi kimia yang terikat pada rantai utama gum arabik berbeda-beda, hal ini dipengaruhi oleh beberapa faktor yaitu musim, iklim, umur pohon, curah hujan, waktu eksudasi, dan faktor lainnya. Rantai utama gum arabik terdiri dari β-(1→3) berikatan dengan unit D-galactopyranosyl. Rantai sampingnya terdiri dua sampai lima β-(1→3) berikatan dengan unit D-galactopyranosyl, berikatan dengan rantai utama pada ikatan 1,6. Baik rantai utama dan rantai samping dari gum arabik mengandung unit dari α-L-arabinofuranosyl-, α-L-rhamnopyranosyl, β-D-glucuronopyranosyl, dan 4-O-methyl β-D-glucuronopyranosyl (Gambar 5) (Verbeken et al., 2003; Ali et al., 2009). Tergantung pada sumber tanamannya, glycan pada gum arabik pada seyal acacia mengandung proporsi L-arabinosa yang lebih besar terhadap D-galaktosa (Motlagh et al., 2006; Chaplin, 2007). Penggunaan gum arabik sebagai bahan enkapsulasi mikroba telah banyak dilakukan diantaranya pada benih jagung dengan isolat B. substillis (Muis dan Arcadio, 2006), benih kacang hijau terhadap B. thuringiensis (Sheikh et al., 2006).
BAB 3
BAHAN DAN METODE
3.1. Waktu dan Tempat
Penelitian ini dilakukan mulai bulan Januari sampai dengan Juli 2013 bertempat di Laboratorium Mikrobiologi dan Laboratorium Sentral, Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam, Laboratorium Analisis Bahan Pangan Fakultas Pertanian, dan Compost Center Universitas Sumatera Utara, Medan.
3.2. Alat dan Bahan
Alat-alat yang dipergunakan adalah tabung reaksi, cawa Petri, jarum ose, bunsen, gelas beaker, Erlenmeyer, pipet mikro, gelas ukur, spatula, pipet volum, propipet, kertas saring, corong, hot plate, vorteks, pinset, stirer, jangka sorong, autoclave, shaker water bath, oven, inkubator, timbangan analitik, sentrifugasi, spektrofotometer, polibaq, gunting, aluminium foil. Bahan-bahan utama yang diperlukan pada penelitian aquades, potato dextrosa agar (PDA), dextrosa, yeast ekstrak (0,3%), alginat, karboksimetil selulosa (CMC), tapioka, gum arabik, NaCl 0,85 %, NaNO3, larutan Mc Farland, HCL 10 N, nutrien agar (NA), kloroks 1 %,
3.3. Isolat
Isolat bakteri kitinolitik yang digunakan diperoleh dari Laboratorium Mikrobiologi Departemen Biologi FMIPA USU Bacillus sp. BK13, Enterobacter sp. BK15, Bacillus sp. BK17, PB08, PB15 dan Enterobacter sp. PB17. Isolat jamur yang digunakan adalah S. rolfsii yang merupakan koleksi Laboratorium Mikrobiologi FMIPA USU.
3.4. Uji Antagonis Bakteri Kitinolitik In Vitro
Kemampuan bakteri kitinolitik dalam menghambat pertumbuhan S. rolfsii diuji dengan asai antagonisme in vitro. Biakan S. rolfsii diinokulasikan pada bagian tengah media PDA yeast. Kultur diinkubasi selama 24 jam. Setelah 24 jam sebanyak 30 µl (≈ 108 sel/ml) sel bakteri diinokulasikan ke dalam media tersebut dengan menggunakan cakram kosong (Oxoid) pada kedua sisi isolat jamur dengan jarak 3 cm. Biakan diinkubasi pada suhu ruang. Zona hambat terhadap miselia S. rolfsii diamati mulai hari kedua sampai ketujuh. Besarnya zona hambat dihitung dengan mengukur selisih jari-jari pertumbuhan jamur normal dengan jari-jari pertumbuhan jamur yang terhambat oleh bakteri.
3.5. Pengamatan Struktur Hifa Jamur Abnormal
3.6. Asai Patogenitas S. rolfsii Terhadap Benih Cabai
Biakan S. rolfsii yang telah diremajakan di cawan petri selama 7 hari diinokulasikan pada 100 ml media GYB di dalam labu erlenmeyer 250 ml dan diinkubasi pada suhu ruang selama 10 hari. Sebanyak 100 ml suspensi biakan S. rolfsii dicampur dengan 500 g campuran pasir, tanah dan kompos steril (nisbah
2:1:1) (Mulyati, 2009) di dalam nampan plastik berukuran 30 cm x 22 cm x 10 cm. Pada tiap nampan ditanam 30 benih cabai terenkapsulasi lalu ditutup dengan plastik lalu diamati selama 30 hari. Benih yang ditanam pada media yang tidak diinokulasi S. rolfsii digunakan sebagai kontrol negatif (K (+)) sedangkan benih yang ditanam pada media yang diinokulasi S. rolfsii digunakan sebagai kontrol positif (K (-)). Peubah yang diamati adalah tanaman yang terserang rebah kecambah selama masa persemaian 30 hari. Persentase rebah kecambah dihitung dari jumlah kecambah yang rebah dibagi jumlah seluruh kecambah yang tumbuh. Peubah yang diamati adalah tanaman yang terserang rebah kecambah, tinggi tanaman, dan jumlah daun selama persemaian 30 hari. Pengurangan persentasi rebah kecambah dihitung dari rumus :
Pengurangan rebah kecambah =
Reisolasi ulang dilakukan terhadap S. rolfsii dengan memotong jaringan pada pangkal batang kecambah yang menunjukkan gejala rebah kecambah. Jaringan tersebut didesinfeksi dengan larutan 2% NaClO, dicuci dengan air steril sebanyak tiga kali dan ditanam pada media PDA. Isolat yang diperoleh diuji kembali patogenitasnya.
3.7. Preparasi Suspensi Isolat Bakteri
Isolat yang akan diujikan diinokulasikan pada campuran media dekstrosa sodium nitrat (NaNO3) 0,03 %, yeast ekstrak 0,2 g dan koloidal kitin sebanyak 2%
selanjutnya diguncang dengan kecepatan 200 rpm selama 4 hari pada suhu ruang. Kultur Enterobacter sp. PB17 kemudian disentrifugasi pada 3000 rpm selama 10 ∑(Kontrol(-)-∑(Kontrol(+) - ∑kecambah rebah
menit, pelet yang diperoleh dipindahkan pada penyangga fosfat (100 mM, pH 7) selanjutnya disuspensikan kembali dengan akuades (1:2 v/v) diukur absorbansinya dengan spektrofotometer pada panjang gelombang 600 nm. Kultur Bacillus sp. BK17 disentrifugasi pada 3000 rpm selama 10 menit, pelet yang diperoleh dipindahkan pada penyangga fosfat (100 mM, pH 7) selanjutnya disuspensikan kembali dengan akuades (1:2 v/v) diinkubasi pada suhu 65oC untuk membunuh sel vegetatifnya selama 15 menit. Kerapatan spora diukur absorbansinya dengan spektrofotometer pada panjang gelombang 660 nm (Pungnoo et al., 2005).
3.8. Enkapsulasi Benih Cabai Merah
Metode enkapsulasi yang digunakan merupakan modifikasi dari Chumthong (2008). Proses enkapsulasi diawali dengan preparasi suspensi isolat bakteri yang diuji, selanjutnya 5 g benih cabai merah dicampur dengan Enterobacter sp. PB17 dengan kerapatan sel OD 600 = 0,5 dan endospora Bacillus sp. BK17 dengan kerapatan spora OD 660 = 0,5 masing-masing sebanyak 10 ml. Selanjutnya dicampur dengan 85 g laktosa monohidrat, 5 g PVP, 50 g tepung talk dan 10 g natrium alginat dan dikeringanginkan. Enkapsulasi tapioka, 5 g benih cabai merah direndam ke dalam 10 ml suspensi Enterobacter sp. PB17 dan endospora Bacillus sp. BK17, kemudian dicampur dengan 3 ml gliserol, 25 g tapioka dan 50 g tepung talk. Semua bahan dicampur kemudian ditambahkan akuades steril sampai volumenya menjadi 100 ml lalu dikeringanginkan.
Enkapsulasi menggunakan CMC digunakan metode modifikasi dari Vidhyasekaran dan Muthamilan (1999), 5 g benih cabai merah dicampur dengan suspensi Enterobacter sp. PB17 dan endospora Bacillus sp. BK17 masing-masing sebanyak 10 ml. Selanjutnya dicampur dengan 100 g tepung talk, 1,5 g CaCO3
dan 5 g CMC lalu dikeringanginkan, sedangkan untuk enkapsulasi dengan menggunakan gum arabik, 5 g benih cabai merah dicampur dengan suspensi Enterobacter sp. PB17 dan endospora Bacillus sp. BK17 masing-masing sebanyak
3.9. Asai Viabilitas Bakteri Kitinolitik Dalam Enkapsulasi
Jumlah koloni kultur bakteri kitinolitik ditentukan dengan menggunakan standard plate count. Satu gram cabai merah terenkapsulasi diambil, kemudian digerus lalu dicampurkan dengan 10 ml akuades steril, dipipet sebanyak 0,1 ml lalu ditumbuhkan pada media MGMK (Suryanto, 2001). Jumlah koloni bakteri kitinolitik yang tumbuh dihitung pada media MGMK yang ditandai dengan zona bening di sekitar koloni. Perhitungan jumlah koloni dilakukan pada minggu ke- 1,2,3 dan 4.
Kombinasi perlakuan enkapsulasi sel bakteri Bacillus sp. BK17 dan Enterobacter sp. PB17 terhadap benih cabai merah dengan menggunakan matriks
[image:40.612.134.507.375.711.2]pembawa alginat, tapioka, gum arabik dan CMC dapat dilihat pada Tabel 1 di bawah ini.
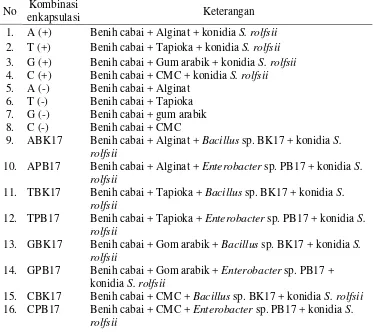
Tabel 1. Kombinasi perlakuan enkapsulasi benih dengan 4 matriks pembawa
No Kombinasi
enkapsulasi Keterangan
1. A (+) Benih cabai + Alginat + konidia S. rolfsii 2. T (+) Benih cabai + Tapioka + konidia S. rolfsii 3. G (+) Benih cabai + Gum arabik + konidia S. rolfsii 4. 5. 6. 7. 8. C (+) A (-) T (-) G (-) C (-)
Benih cabai + CMC + konidia S. rolfsii Benih cabai + Alginat
Benih cabai + Tapioka Benih cabai + gum arabik Benih cabai + CMC
9. ABK17 Benih cabai + Alginat + Bacillus sp. BK17 + konidia S. rolfsii
10. APB17 Benih cabai + Alginat + Enterobacter sp. PB17 + konidia S. rolfsii
11. TBK17 Benih cabai + Tapioka + Bacillus sp. BK17 + konidia S. rolfsii
12. TPB17 Benih cabai + Tapioka + Enterobacter sp. PB17 + konidia S. rolfsii
13. GBK17 Benih cabai + Gom arabik + Bacillus sp. BK17 + konidia S. rolfsii
14. GPB17 Benih cabai + Gom arabik + Enterobacter sp. PB17 + konidia S. rolfsii
15. 16.
CBK17 CPB17
3.10. Asai Patogenitas S. rolfsii terhadap Benih Cabai Terenkapsulasi di Rumah Kasa
Benih cabai yang telah dienkapsulasi ditanam pada polibeg yang telah diisi dengan campuran kompos, tanah dan pasir (nisbah 2:1:1) (Mulyati, 2009) dengan 3 perlakuan. Perlakuan pertama benih yang sudah terenkapulasi oleh bakteri Enterobacter sp. PB17 dan Bacillus sp. BK17 ditanam pada media tanam diberi
BAB 4
HASIL DAN PEMBAHASAN
4.1. Kemampuan Antagonis Bakteri Kitinolitik secara In Vitro
Hasil uji antagonisme 6 isolat bakteri kitinolitik terhadap jamur S. rolfsii menunjukkan bahwa keenam isolat kitinolitik mampu menghambat pertumbuhan S. rolfsii dengan kemampuan yang berbeda-beda. Zona hambat umumnya mulai
[image:42.612.129.507.406.526.2]teramati pada hari kedua dan jarak zona hambat terus bertambah, dan setelah hari ketujuh umumnya tidak terjadi lagi pertambahan jarak zona hambatan seperti terlihat pada Tabel 2 di bawah ini.
Tabel.2. Penghambatan pertumbuhan jamur S. rolfsii oleh bakteri kitinolitik No. Isolat Rata-rata zona hambatan (mm) hari ke
2 3 4 5 6 7
1. Bacillus sp. BK13 2,10 4,69 7,76 2,01 0,00 0,00 2. Enterobacter sp. BK15 2,23 5,65 5,15 1,69 0,00 0,00 3. Bacillus sp. BK17 1,79 3,81 8,33 13,50 13,90 11,40 4. PB08 0,55 1,74 0,00 0,00 0,00 0,00 5. PB15 1,53 0,75 0,05 0,00 0,00 0,00 6. Enterobacter sp. PB 17 1,19 3,88 6,13 6,68 2,18 0,00
Pada pengamatan hari ketujuh dari keenam isolat tersebut, isolat yang menunjukkan efektivitas paling tinggi dalam menghambat pertumbuhan jamur S. rolfsii adalah Bacillus sp. BK17 dengan jari-jari zona hambat sebesar 11,40 mm
dan diikuti oleh Enterobacter sp. PB17 dengan jari-jari zona hambat sebesar 2,18 cm pada hari keenam. Isolat yang paling lemah dalam menghambat jamur S. rolfsii adalah BK08 yang hanya mampu menghambat sampai hari ketiga sebesar
daya hambat sampai penurunan daya hambat. Perbedaan daya hambat masing-masing isolat ini bisa terjadi karena dipengaruhi oleh adanya perbedaan kemampuan bakteri kitinolitik dalam menghasilkan metabolit / protein antifungi seperti enzim kitinase. Media antagonistik yang mengandung kitin menginduksi bakteri dalam menghasilkan enzim kitinase untuk menguraikan kitin sebagai sumber karbonnya. Produksi enzim dipengaruhi jenis karbon yang diformulasikan ke dalam media uji dengan pengaturan pH dan jumlahnya (El-katatny et al., 2000).
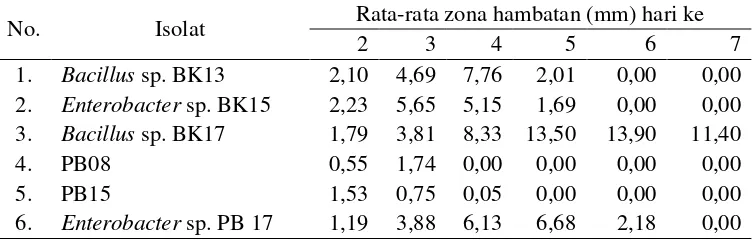
Menurut Ferniah et al., (2003) kitin merupakan induser bagi enzim kitinase, dengan memecah kitin menjadi ketooligosakarida sampai dengan N-asetil D-glukosamin yang akan mengalami deN-asetiliasi dan deaminasi dan menghasilkankan molekul-molekul glukosa. Ketika koloni bakteri yang ada pada media sudah semakin tumbuh dan mendekati koloni jamur yang ada kitin pada dinding sel jamur digunakan sebagai sumber karbon untuk pertumbuhan bakteri, sehingga dinding sel jamur mengalami kerusakan yang mengakibatkan pertumbuhan jamur menjadi terhambat. Kandungan kitin yang terdapat pada dinding sel jamur juga mempengaruhi besarnya zona hambat isolat pada masing-masing jamur. Semakin besar kandungan kitin pada dinding sel maka semakin besar zona hambat yang terbentuk. Bentuk zona hambatan tersebut berupa cerukan penipisan elevasi seperti terlihat pada Gambar 6.
[image:43.612.146.495.506.676.2]Mekanisme penghambatan yang terjadi pada uji antagonisme dapat diamati dengan terbentuknya zona bening sebagai zona penghambatan pertumbuhan jamur S. rolfsii oleh bakteri kitinolitik. Zona bening ini terbentuk karena terjadi pemutusan ikatan β-1,4 homopolimer N-asetilglukosamin pada kitin oleh kitinase menjadi monomer N-asetilglukosamin. Perbedaan zona bening yang dihasilkan disebabkan adanya perbedaan aktivitas kitinase pada isolat Enterobacter sp. PB17 dan Bacillus sp. BK17.
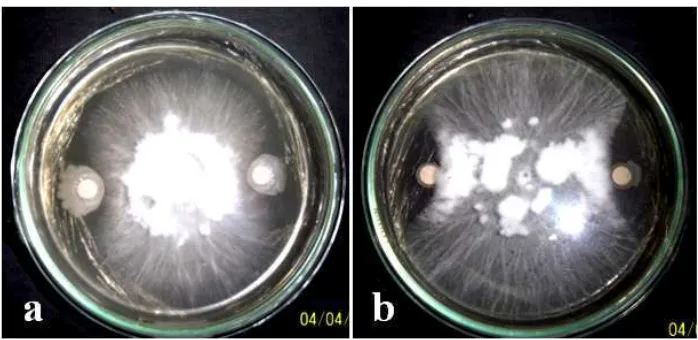
4.2. Abnormalitas Hifa Jamur S. rolfsii setelah Uji Antagonis dengan Bakteri Kitinolitik
Pengamatan hifa abnormal dilakukan setelah dilakukan uji antagonis antara bakteri kitinolitik Bacillus sp. BK17 dan Enterobacter sp. PB17 dengan jamur S. rolfsii dengan pengamatan selama 7 hari. Setelah 7 hari pengamatan dan dilanjutkan dengan pengamatan secara mikroskopis ditemukan adanya perbedaan struktur hifa antara hifa jamur yang normal dengan hifa yang mengalami abnormalitas (Gambar. 7). Struktur hifa yang abnormal terlihat menggulung pada bagian ujungnya, lisis pada dinding selnya, lisis pada isi selnya, bengkok dan putus.
Gambar 7. Perubahan morfologi hifa jamur S. rolfsii setelah uji antagonis dengan bakteri antagonis pada media MGMK + yeast (a). hifa normal, (b). hifa menggulung pada bagian ujung hifa, (c). hifa lisis pada bagian dinding hifa, (d).hifa bengkok, (e). isi hifa lisis, (f). hifa putus
Rhizoctonia dan Phytopthora (Gohel et al., 2003), G. boninense, P. citrinum dan
F. oxysporum (Suryanto et al., 2011).
4.3. Patogenitas S. rolfsii pada Benih Cabai
Dari hasil uji patogenitas S. rolfsii terhadap benih cabai merah diperoleh bahwa S. rolfsii penyebab penyakit rebah kecambah, dengan persentase rebah kecambahnya
mencapai 64,71% dari benih yang tumbuh pada kontrol positif (K (+)) (Gambar 8). Menurut Agrios (1997), jamur S. rolfsii menyebabkan rebah kecambah pada persemaian dan busuk batang pada tanaman inang yang lebih tua. Apabila tanaman muda yang terserang tanaman cepat mati, tetapi bila tanaman yang lebih tua terserang, pucuk tanaman menjadi kuning layu dan kemudian mati. Akibatnya bagian dari batang dan akar pada batas tanah menjadi busuk dan sebagian ditutupi oleh bercak putih yaitu miselium yang berwarna putih sampai terbentuk sklerotia coklat.
Gambar 8. Persentase rebah kecambah pada benih cabai (BK17 = benih cabai + Bacillus sp. BK17, PB17 = Benih cabai + Enterobacter sp. PB17).
[image:46.612.187.464.393.578.2]sp. BK17 dan Enterobacter sp. PB17. Serangan jamur S. rolfsii bisa dihambat karena dinding sel tersusun oleh kitin, sehingga enzim kitinase yang dihasilkan oleh Enterobacter sp. PB17 dan Bacillus sp. BK17 dapat menghidrolisis kitin yang ada pada dinding sel jamur S. rolfsii. Menurut El-Katatny et al., (2000), pengendalian hayati jamur dengan menggunakan mikroorganisme kitinolitik didasarkan pada kemampuan mikroorganisme tersebut dalam menghasilkan kitinase dan β-1,3 glukanase yang dapat melisiskan dinding sel jamur. Pelisisan dinding sel jamur ini bisa terjadi karena salah satu penyusun diinding sel jamur adalah kitin, sehingga enzim kitinase dapat menghidrolisis kitin yang ada pada dinding sel jamur (Sing et al., 1999; Quecine et al., 2008). Kandungan kitin pada jamur bervariasi dari 4 – 9 % berat kering sel, tergantung spesies atau strain jamurnya (Rajarathanam et al., 1998).
Gambar 9. Benih cabai yang normal (a) dan yang terserang S. rolfsii (b) setelah 30 hari, 1. Hifa S. rolfsii pada pangkal batang, 2. Batang yang menguning dengan daun yang sudah gugur.
Menurut Maryudani dan Sudarmadi (1976), gejala serangan S. rolfsii mula-mula diawali dengan menguning dan melayunya daun-daun, kemudian diikuti dengan membusuknya batang. Kulit dan kayu pada pangkal batang serta akar tanaman yang terserang rusak. Pada tingkat penyerangan belum lanjut, bila tanaman tersebut dicabut akan terlihat benang-benang halus pada bagian yang sakit.
4.4. Viabilitas Bakteri Kitinolitik Dalam Enkapsulasi
Gambar 10. Viabilitas sel bakteri pada benih terenkapsulasi selama penyimpanan
Dari gambar di atas setelah dilakukan perhitungan selama 4 minggu viabilitas sel tertinggi ditemukan pada benih yang dienkapsulasi dengan gum arab dan Bacillus sp. BK17 (GBK17) sebesar 198 x 1010 cfu/gr, kemudian diikuti oleh benih yang dienkapsulasi dengan CMC Bacillus sp. BK17 (CBK17) sebesar 182 x 1010 cfu/gr, sedangkan viabilitas sel terendah ditemukan pada benih yang dienkapsulasi dengan alginat dan Enterobacter sp. PB17 (APB17) sebesar 8 x 1010 cfu/gr. Hal ini menunjukkan bahwa formulasi terbaik yang bisa digunakan sebagai bahan enkapsulasi bakteri pada benih adalah gum arabik. Gum arabik adalah campuran kompleks dari oligosakarida arabinogalactan, polisakarida, glikoprotein. Rantai utama gum arabik terdiri dari β-(1→3) berikatan dengan unit D-galactopyranosyl. Rantai sampingnya terdiri dua sampai lima β-(1→3)-berikatan dengan unit D-galactopyranosyl, β-(1→3)-berikatan dengan rantai utama pada ikatan 1,6. Baik rantai utama dan rantai samping dari gum arabik mengandung unit dari α-L-arabinofuranosyl-, α-L -rhamnopyranosyl, β-D-glucuronopyranosyl, dan 4-O-methyl β-D-glucuronopyranosyl (Verbeken et al., 2003; Ali et al., 2009).
PB17 dan Bacillus sp. BK17 bertahan dalam kondisi lingkungan yang baru yang kering. Jika dibandingkan jumlah sel antara bakteri Enterobacter sp. PB17 dan Bacillus sp. BK17 pada 1 gram benih terenkapsulasi, bakteri Bacillus sp. BK17 lebih banyak dibandingkan dengan Enterobacter sp. PB17. Hal ini menunjukkan bahwa Bacillus sp. BK17 memiliki viabilitas yang lebih tinggi dibandingkan dengan Enterobacter sp. PB17. Hal ini dikarenakan Bacillus sp. BK17 mampu menghasilkan spora untuk bertahan hidup di kondisi yang ekstrim.
Bacillus merupakan salah satu bakteri yang dapat membentuk endospora
pada kondisi lingkungan yang tidak menguntungkan bagi dirinya. Spora yang terbentuk merupakan struktur bertahan dari Bacillus. Spora ini dapat bertahan dalam waktu yang lama hingga mencapai puluhan tahun. Namun kemampuan bertahan spora bacillus dipengaruhi oleh jenis media atau bahan yang digunakan untuk penyimpanan (Sulistiani, 2009).
4.5. Patogenitas S. rolfsii pada Benih Cabai Terenkasulasi
Pengamatan jumlah benih yang tumbuh dilakukan selama 6 minggu. Dari pengamatan diketahui bahwa jumlah benih yang tumbuh (Gambar 11) lebih banyak pada benih yang dienkapsulasi oleh tapioka daripada yang dienkapsulasi oleh bahan enkapsulasi lainnya yaitu sebesar 80% pada TBK17, diikuti oleh gum arabik dan CMC sebesar 60%, selanjutnya diikuti oleh alginat sebesar 0-20%. Hal ini menunjukkan bahwa kapsul yang terbuat dari tapioka lebih mudah ditembus oleh benih ketika berkecambah. Disamping itu jumlah benih yang paling banyak tumbuh adalah benih yang sudah dicampur dengan isolat Bacillus sp. BK17. Hal ini menandakan bahwa Bacillus memiliki peranan dalam pertumbuhan benih cabai.
mempunyai kemampuan untuk memproduksi hormon asam indolasetat, asam giberelat, sitokonin, dan etilen di dalam tanaman (Sulistiani, 2009; Susanto, 2008).
Gambar 11. Persentase benih cabai yang tumbuh dan yang rebah
Benih cabai yang telah terenkapsulasi bakteri kitinolitik ditumbuhkan pada medium tanah yang telah diberikan suspensi S. rolfsii. Pengamatan dilakukan terhadap persentase rebah kecambah, tinggi kecambah dan jumlah daun dari minggu ke-0 sampai minggu ke-4. Kecambah yang ditumbuhkan pada tanah yang mengandung S. rolfsii sangat rentan terserang jamur ini. Hal ini terlihat pada perlakuan C(+) dan G(+) masing-masing 40% dan 20% dan benih yang dienkapsulasi dengan bakteri Enterobacter sp. PB17 pada TPB17 dan GPB17 masing - masing sebesar 20% sedangkan K (-) tidak ada benih yang rebah (Gambar 11).
pangkalnya menjadi basah dan mengerut sehingga semai roboh dan mati (Semangun, 2007). S. rolfsii merupakan salah satu jamur patogen yang dapat menyebabkan rebah kecambah pada tanaman cabai (Semangun, 2007; Lamidi, 1986; Dange, 2006; Yusniawaty, 2009).
[image:52.612.147.498.420.609.2]Benih yang dienkapsulasi dengan Bacillus sp. BK17 memiliki kemampuan yang lebih tinggi dibandingkan dengan Enterobacter sp. PB17 dalam menghambat serangan S. rolfii. Hal ini menunjukkan bahwa kemampuan enzim kitinase pada Bacillus sp. BK17 lebih baik dalam mendegradasi kitin yang ada pada hifa S. rolfsii dibandingkan dengan enzim kitinase pada Enterobacter sp. PB17. Kitinase merupakan enzim yang mampu menghambat perkembangan jamur patogen dengan menghidrolisis polimer kitin sebagai salah satu komponen dinding sel hifa cendawan (Gohel et al., 2003), dan secara langsung menghidrolisis dinding miselia jamur dengan pelepasan elisitor endogen oleh aktivitas kitinase yang kemudian memicu reaksi ketahanan sistemik pada inang. Oleh sebab itu, kitinase dikenal sebagai salah satu protein anti cendawan (Gohel et al., 2003, Wang et al., 2006).
Gambar 12. Jumlah rata-rata daun dan tinggi tanaman setelah 6 minggu
dan ABK17 dan diikuti oleh C(+) sebanyak 10 helai (Gambar 12), sedangkan jumlah daun yang paling sedikit ditemukan pada APB17 sebanyak 2 helai. Kecambah yang tertinggi juga ditemukan pada benih yang dienkapsulasi oleh tapioka TBK17 dan T (-) setinggi 13 cm dan diikuti oleh ABK17 dan C (+) sebesar 10, 50 cm, sedangkan tanaman yang terendah ditemukan pada benih APB17 sebesar 2 cm. Adanya variasi jumlah daun dan tinggi kecambah pada masing-masing perlakuan disebabkan adanya perbedaan usia perkecambahan pada masing-masing perlakuan. Benih yang memiliki jumlah daun terbanyak adalah benih yang tumbuh lebih awal dibandingkan dengan benih yang lain. Perbedaan jenis matrik pembawa pada masing-masing perlakuan mempengaruhi kemampuan benih dalam berkecambah. Semakin sederhana komponennya maka semakin mudah bagi benih untuk tumbuh dan berkecambah.
BAB 5
KESIMPULAN DAN SARAN
5.1 Kesimpulan
Adapun kesimpulan dari penelitian ini adalah:
1. Bacillus sp. BK17 merupakan bakteri kitinolitik yang terbaik dalam menghambat pertumbuhan jamur S. rolfsii pada benih cabai merah terekapsulasi maupun yang tidak terenkapsulasi.
2. Gum arabik merupakan bahan yang baik digunakan untuk enkapsulasi benih dengan kemampuan viabilitas bakteri yang tertinggi dibandingkan dengan bahan enkapsulasi lainnya.
3. Semua bakteri kitinolitik yang terenkapsulasi alginat, CMC, tapioka dan gum arabik yang diuji mengalami penurunan viabilitas sel pada minggu kedua namun tingkat penurunannya semakin kecil dan cenderung stabil pada minggu keempat.
4. Benih yang sudah dienkapsulasi dengan Bacillus sp. BK17 dan Enterobacter sp. PB17 memiliki viabilitas yang baik dibandingkan benih yang lainnya.
5.2 Saran
DAFTAR PUSTAKA
Agrios, G. N. 1997. Plant Pathology. 2nd Edition. Academic Press, New York. hlm 416.
Alexopoulos, C. J. dan Charles W. M. 1979. Introductory Mycology. 4th ed. John Wille & Sons Inc., New York. hlm. 632.
Ali, B. H., Amal Z. dan Gerald B. 2009. Biological Effects of Gum Arabic: A review of some recent research. Food and Chem Toxicol. 47: 1–8.
Awalludin A. 2004. Karboksimetilasi Selulosa Bakteri. Skripsi. Fakultas Matematika dan Ilmu Pengetahuan Alam. Institut Pertanian Bogor. Backman, P. A, Brannnen P. M dan Mahaffe W. F. 1994. Plant Respon and
Disease Control Followin Seed Inoculation with Bacillus sp. Di dalam: Ryder MH, Stephen PM, Bowen GD, editor. Improving Plant Production with Rhizosphere Bacteria. Australia: Pruc Third Int Work PGPR South Australia.
Bashan, Y. 1986. Alginate Beads as Synthetic Inoculant Carriers for Slow Release of Bacteria That Affect Plant Growth. Appl Environ Microbiol. 51(5): 1089-1098.
Boer, M. D., Peter B., Frondo K., Joost J. B., Keurentjes, Lentse V. D. S., vanLoon L. D dan Bakker A. H. M. 2003. Control of Fusarium Wilt of Radish by Combining Pseudomonas putida Strains that have Different Disease Suppressive Mechanisms. Section of Phytopathology. Netherlands Utrecth University. hlm. 626-632.
Boyle, L.W. 1961. The Ecology of Sclerotium rolfsii with Emphasis on The Role of Saprophytic Media. Phytopathol. 51:118-119.
Charpentier, A. C, Gadille P., Digat B dan Benoit J. P. 1998.Microencapsulation of Rhizobacteria by Spray-Drying: Formulation And Survival Studies. J. Microencapsul. 15:639-59.
Chaplin, M. F. 2007. Water Structure and Science. http://www.lsbu.ac.uk/water/. Diakses 21 Januari 2013.
Sheath Blight: Formulation, Bacterial Viability and Efficacy Testing. J Microb Biotechnol. 24: 2499-2507. DOI: 10.1007/s11274-008-9774-7.
Colwell, R. R., Pariser E. R dan Sinskey A. J. 1985. Biotechnology of Marine Polysaccharides. Massachusetts Institute of Technology. Sea Grant Collenge Programe. hlm. 474.
Cummings, J. A. 2009. Greenhouse Evaluation of Seed and Drench Treatments for Organic Management of Soilborne Pathogens of Spinach. Plant Disease. doi:10.1094/PDIS-93-12-1281.93:1281-1292.
Dange, V. 2006. Studies on Root Rot of Chilli Caused by Sclerotium rolfsii sacc. Tesis. Department of Plant Pathology College of Agriculture, Dharwad University of Agricultural Sciences, Dharwad.
Dawar, S., Marium T dan Zakia M.T. 2008. Aplication of Bacillus species in Control of Meloidogyne Javanica (Treub) Chitwood on Cowpea and Mash Bean. J Bot. 40: 439-444.
Departeman Pertanian. 2004. Pusat Data dan Informasi Pertanian. Tim Penyusun. http://www.deptan.go.id. Diakses 13 Nopember 2012.
Djatmiko, H. A., Triwidodo A., Bambang, H dan Bambang H. S. 2007. Potensi Tiga Genus Bakteri dari Tiga Rizosfer Tanaman sebagai Agensia Pengendali Hayati Penyakit Lincat. J Ilmu-ilmu Pertanian Indonesia. 9(1).
Dziedzic, S. Z dan Kearsley M.W. 1995. The technology of Starch Production. Di dalam Kearsley, M.W. dan S.Z. Dziedzic. Handbook of Starch Hydrolysis Products and Their Derivatives. Blackie Academic & Prof., London. hlm. 24.
Eka, S. P. 2009. Pengaruh Komposisi Bahan Pelapis dan Methylobacterium spp. Terhadap Daya Simpan Benih dan Vigor Bibit Kacang Panjang (Vigna sinensis L.). Tesis. Fakultas Pertanian. Institut Pertanian Bogor. Bogor.
El-Katatny, M. H., Somitsch W., Robra1 K.-H., El-Katatny M. S dan Gübitz1. G.
M. 2000. Production of Chitinase and β-1,3-glucanase by Trichoderma
harzianum for Control of the Phytopathogenic Fungus Sclerotium rolfsii. Food technol. Biotechnol. 38:173–180.
Epps, W. M., Patterson J. C dan Freeman I. E. 1951. Physiology and Parasitism of Sclerotium rolfsii. Phytopathol. 41: 245-256.
FAO .1999. Gum Arabik. (Food and Nutrition Paper 52, addendum 7). FAO, Rome.
Kentang (Solanum tuberosum). Fakultas Matematika Dan Ilmu Pengetahuan Alam Universitas Diponegoro. Semarang.
Frazier, W. C dan Westhoff D. C. 1998. Food microbiology. Mc Graw-Hill Book Co. New York.
Gohel, V, Singh A., Vimal M., Ashwini D dan Chhatpar H. S. 2003. Bioprospecting and Antifungal Potential of Chitinolytic Microorganism. African J Biotechnol. 5: 54-72.
Hameeda, B., Harini G., Rupela O. P, Kumar R. J. V. D. K dan Gopal R. 2010. Biological Control of Chickpea Collar Rot by Co-inoculation of Antagonistic Bacteria and Compatible Rhizobia. Indian J Microbiol .50:419–424.
Harni, R., Abdul M., Supramana dan Ika M. 2007. Potensi Bakteri Endofit Pengendali Nematoda Peluka Akar (Pratylenchus brachyurus) pada Nilam. Hayati J Bioscie.14: 7-12
Hartati, S.Y., Taufik E., Supriadi dan Karyani N. 2008. Karakteristik Fisiologis Isolat Sclerotium sp. Asal Tanaman Sambiloto. J Littri.14: 25 – 29. Hustiany, R. 2006. Modifikasi Asilasi dan Suksinilasi Pati Tapioka sebagai bahan
enkapsulasi Komponen flavor. Disertasi. Institut Pertanian Bogor. Bogor. Islam, M. T dan Faruq A. N. 2012. Effect of Some Medicinal Plant Extracts on Damping-off Disease of Winter Vegetable. J World App Scie. 17: 1498-1503.
Istikorini, Y. 2006. Efektifitas Cendawan Endofit Untuk Mengendalikan Penyakit Antraknosa, Meningkatkan Pertumbuhan dan Hasil cabai (Capsicum annum L.). Skripsi. Fakultas Pertanian. Institut Pertanian Bogor.
Jacobs, H. dan Delcour J. A. 1998. Hydrothermal modifications of granular starch, with retention of the granular structure: a review. J Agric. Food Chem. 46:2895-2905.
Jinantana, J dan Sariah M. 1998. Potential for Biological Control of Sclerotium Foot Rot of Chilli by Trichoderma spp. Pertanika. J Trap Agric Sci. 21: 1 -10.
Kim K.I., Baek Y.J dan Yoon Y.H. 1996. Effects of Rehydration Media And Immobilisation In Calcium-Alginate On The Survival of Lactobacillus casei and Bifidobacterium bifidum. J Dairy Sci. 18:193–198.
King, A. H. 1995. Encapsulation of Food Ingredients: Revew of Available Technology, Focusing on Hydrocolloids. Abstrak. AS Symposium Series, Volume 590. American Chemical Society.
Kloepper, J. W dan Schroth M. N. 1981. Development of a Powder Formulation of Rhizobacteria for Inoculation of Potato Seed Pieces. Phytopathol. 71: 590-592.
Kneifel, W., Jaros D dan Erhard F. 1993. Microflora and Acidification Properties ofyoghurt and Yoghurt Related Products Fermented With Commercially Available Starter Cultures. Int J Food Microbiol. 18:179–189.
Laitila, A., Alakomi H-L, Raaska L, Mattila-Sandholm T dan Haikara A. 2002. Activities of Two Lactobacillus Plantarum Strains against Fusarium Moulds In Vitro and In Malting of Barley. J of App Microbiol. 93: 566– 576.
Lamidi, 1986. Faktor-Faktor yang Mempengaruhi Perkecambahan dan Kepekaan Sklerotia Sclerotium rolfsii Sacco terhadap Serangan Trichoderma sp.