BIOSYSTEMATICS OF
PANDANACEAE
IN JAVA
SRI ENDARTI RAHAYU
THE GRADUATE SCHOOL
BOGOR AGRICULTURAL UNIVERSITY
BOGOR
STATEMENT OF RESEARCH ORIGINALITY AND
INFORMATION SOURCE
This is to certify that my dissertation entitled Biosystematics of Pandanaceaein Java is my own work and never been submitted to any institution before. All the incorporated data and information are valid and stated clearly in the text and listed in the references.
Bogor, Januari 2011
ABSTRACT
SRI ENDARTI RAHAYU. Biosystematics of Pandanaceae in Java. Supervised by ALEX HARTANA, TATIK CHIKMAWATI, KUSWATA KARTAWINATA, and MIEN A. RIFAI.
Pandan family (Pandanaceae) is represented in Java by two genera: Freycinetia Gaud. and Pandanus Parkins. Since the studies by Backer and Bakhuizen van den Brink in 1968 and Stone in 1972, there were no further exploration on the pandan flora of the island have been made, thus the pandan flora remains largely unknown. Taxonomical problem as far as the Java pandans concerned are centered not only on species status ofP. odoratissimus L.f and P. tecroriusvar.littoraliswhich are regarded as synonym, but also the species status which are given as synonym ofP. furcatus Roxb. by Backer and Bakhuizen van den Brink. Backer and Bakhuizen van den Brink included of P. bantamensis Koord., P. oviger Martelli, P. pseudolais Warb. and P. scabrifolius Martelli as synonym to P. furcatus Roxb. This Backer and Bakhuizen van den Brink’s classification is in contrast with Stone who stated that these species were regarded as four different species. For the reason of this taxonomical problem, therefore an effort has been made to redescribe these species in detail, using morphological, anatomical and molecular data such as sequence data of atpB-rbcL IGS, since a satisfactory classification depends upon the interpretation of many characters as possible. In this study we only found three of four species mentioned above, viz. P. bantamensis Koord., P. pseudolais Warb., and P. scabrifolius Martelli. The result showed that P. bantamensis Koord., P. pseudolais Warb, and P. scabrifolius Martelli are treated as three different species, and P. odoratissimus L.f and P. tectorius var.littoralis are treated as two different species. The ISSR marker showed that six species of Freycinetia and thirteen species of Pandanus from Java have high genetic diversity, although Freycinetia has a bit lower than Pandanus. This research showed seven species of Freycinetia could be recognized, viz. F. angustifolia Bl., F. funicularis Merr., .F. imbricata Bl., F. insignisBl.,F. javanicaBl.,F. scandensGaud., andF. sumatranaHemsl.; sixteen species of Pandanus, viz. P. amaryllifolius Roxb., P. bantamensis Koord., P. bidur Jungh., P. dubius Spreng., P. faviger Backer, P. kurzii Merr., P. labyrinthicus Kurz, P. multifurcatus Fagerl., P. nitidus Kurz, P. odoratissimus L.f., P. polycephalus Lam., P. pseudolais Warb., P. scabrifolius Martelli, P. spinistigmaticusFagerl., P. tectorius Parkins. with two varieties, viz. P. tectorius var. littoralis, P. tectorius var. samak, one cultivar, i.e Pandanus tectorius cv. Sanderi; P. utilis Bory; one variety P. leram Jones var. andamanensium (Kurz) Stone, and one cultivar,P. spuriusMiq. cv. Putat.Pandanaceaein Java are most diverse in lowland rain forest and hill forest.
ABSTRAK
SRI ENDARTI RAHAYU. Biosystematics of Pandanaceae in Java. Di bawah bimbingan ALEX HARTANA, TATIK CHIKMAWATI, KUSWATA KARTAWINATA dan MIEN A. RIFAI.
Pandanaceae di Jawa diwakili oleh dua marga, yaitu Freycinetia Gaud. danPandanusParkins. Sejak penelitian oleh Backer dan Bakhuizen van den Brink pada tahun 1968 dan Stone pada tahun 1972, belum pernah dilakukan eksplorasi lebih jauh tentang flora pandan di Jawa sehingga sampai saat ini banyak hal yang belum diketahui tentang flora pandan tersebut. Masalah taksonomi tentang Pandanaceae yang ada di Jawa, tidak hanya tentang status spesies dari P. odoratissimus L.f dan P. tectorius var. littoralis yang dianggap sinonim, tetapi juga status spesies yang merupakan sinonim dari P. furcatus Roxb, yaitu P. bantamensisKoord.,P. oviger Martelli,P. pseudolais Warb., dan P. scabrifolius Martelli. Backer dan Bakhuizen van den Brink memasukkan P. bantamensis Koord., P. oviger Martelli, P. pseudolais Warb., dan P. scabrifolius Martelli ke dalam satu spesies, yaituP. furcatusRoxb., sedangkan menurut Stone ke empat spesies tersebut adalah empat spesies yang berbeda. Berdasarkan latar belakang permasalahan tersebut perlu dilakukan penelitian untuk dapat mendeskripsikan kembali spesies-spesies tersebut dengan menggunakan data morfologi, anatomi dan molekular, seperti sekuen antar genatpB-rbcL, karena suatu klasifikasi yang paling memuaskan tergantung pada interpretasi dari sebanyak mungkin karakter. Dalam penelitian ini hanya ditemukan tiga dari empat spesies tersebut di atas, yaitu P. bantamensisKoord., P. pseudolais Warb., dan P. scabrifolius Martelli. Hasil penelitian menunjukkan bahwa P. bantamensis Koord., P. pseudolais Warb., dan P. scabrifolius Martelli diperlakukan sebagai tiga spesies yang berbeda, dan P. odoratissimus L.f dan P. tecrorius var. littoralis diperlakukan sebagai dua spesies yang berbeda. Penelitian ISSR menunjukkan bahwa ke enam spesies Freycinetia dan 13 spesies Pandanus di Jawa memiliki keanekaragaman genetik yang tinggi, walaupun keanekaragaman genetik Freycinetia sedikit lebih rendah bila dibandingkan denganPandanus.Hasil penelitian menunjukkan bahwa di Jawa dapat dikenali tujuh spesies Freycinetia dan 16 spesies Pandanus. Ketujuh spesiesFreycinetia tersebut mencakupF. angustifoliaBl.,F. funicularis Merr., F. imbricata Bl., F. insignis Bl., F. javanicaBl., F. scandensGaud., dan F. sumatrana Hemsl. Keenambelas spesies Pandanus tersebut terdiri dari P. amaryllifolius Roxb.,P. bantamensis Koord.,P. bidur Jungh.,P. dubius Spreng, P. favigerBacker,P. kurziiMerr.,P. labyrinthicusKurz, P. multifurcatusFagerl., P. nitidus Kurz, P. odoratissimus L.f., P. polycephalus Lam., P. pseudolais Warb., P. scabrifoliusMartelli, P. spinistigmaticus Fagerl.,P. tectorius Parkins, dengan dua varietas, yakniP. tectoriusvar.littoralis, P. tectoriusvar.samak, satu kultivar, yaitu Pandanus tectoriuscv. Sanderi, P. utilisBory; satu varietas, P. leramJones var.andamanensium( Kurz) Stone, dan satu kultivar,P. spuriusMiq. cv. Putat. Di Jawa, keanekaragaman Pandanaceae yang paling tinggi terdapat di hutan pegunungan bawah dan bukit pegunungan.
SUMMARY
SRI ENDARTI RAHAYU. Biosystematics ofPandanaceaein Java. Supervised by ALEX HARTANA, TATIK CHIKMAWATI, KUSWATA KARTAWINATA, and MIEN A. RIFAI.
Pandan family (Pandanaceae) is represented in Java by two genera, Freycinetia Gaud. and Pandanus Parkins. Since the studies by Backer and Bakhuizen van den Brink in 1968 and Stone in 1972, there were no further exploration on the pandan flora of the island have been made, thus the pandan flora remains largely unknown. Taxonomical problem as far as the Java pandans concerned, are centered not only on the species status ofPandanus odoratissimus L.f andP. tecrorius var.littoralis which are considered as synonym, but also the species status which are given as synonym ofPandanus furcatusRoxb. by Backer and Bakhuizen van den Brink. Under the latter they included of P. bantamensis Koord.,P. oviger Martelli,P. pseudolaisWarb., and P. scabrifoliusMartelli into one species ofP. furcatusRoxb. This classification is in contrast with Stone who stated that these species were regarded as four different species. Therefore an effort has been made to redescribe these species in detail, using morphological, anatomical and molecular data such as sequence data ofatpB-rbcL IGS to provide better understanding of morphological, anatomical and molecular characters in supporting taxa delimitation and its distribution in Java.
This study was based mainly on available specimens at the Herbarium Bogoriemse (BO), National Herbarium Netherlands, Leiden (L) and Herbarium of the Royal Botanical Gardens Kew (K) and new collection specimens obtained from field work in different location in Java. In addition, living plants grown in botanical garden were also studied. Five species that was planted in Bogor Botanical Garden, viz. Pandanus kurzii, P. labyrinthicus, P. multifurcatus, P. polycephalusandP. spinistigmaticuswere also examined.
Characters of leaf shape, leaf apex, the morphology of leaf auricles and type of pistillate inflorecence were found useful in delimitation and identification of Javanese Freycinetia, while character of habit, the surface of stem, present or absent of prop root, the surface of prop root, the leaf shape, leaf apex, the armature of leaf margins and midrib, the colour of leaf margin and midrib teeth, the distinctness or indistinctness of tertiary cross vein, present or absent of apical ventral pleats, the texture of leaves in dry state, phalange shapes, the position of infructescence, position of seed chamber and stigma shape are proved useful for distinguishing species of JavanesePandanus.
the epidermal tissue including the stomata proves to be a great value in identification ofPandanusspecies in Java.
Morphological, anatomical and comparative sequence data of atpB-rbcL IGS were able to solve the taxonomical problem of Pandanus furcatus complex and P. tectorius complex and as a result P. bantamensis Koord, P. pseudolais Warb. and P. scabrifolius Martelli treated as three different species, and P. odoratissimusL.f andP. tectoriusvar.littoralis treated as two different species.
ISSR (Inter Simple Sequence Repeat) study showed that six species of Freycinetia and thirteen species of Pandanus from Java have high genetic diversity, although Freycinetia has a bit lower of genetic diversity than Pandanus, while Principal component analysis (PCA) for Javanese thirteen Pandanusand six JavaneseFreycinetiashowed a bit different in clustering pattern and species relationships compared to the cluster analysis.
This study showed that in Java there are seven species of Freycinetia, viz. F. angustifolia Bl., F. funicularis Merr., F. imbricata Bl., F. insignis Bl., F. javanica Bl.,F. scandensGaud., andF. sumatranaHemsl.; and sixteen species of Pandanus, viz.P. amaryllifoliusRoxb., P. bantamensisKoord., P. bidur Jungh., P. dubius Spreng., P. faviger Backer, P. kurzii Merr., P. labyrinthicusKurz, P. multifurcatus Fagerl., P. nitidus Kurz, P. odoratissimus L.f., P. polycephalus Lam., P. pseudolais Warb., P. scabrifolius Martelli., P. spinistigmaticus Fagerl., P. tectoriusParkins, with two varieties, viz.P.tectoriusvar.littoralis,P. tectorius var. samak, one cultivar, i.e Pandanus tectorius cv. Sanderi, P. utilis Bory; one variety P. leram Jones var. andamanensium (Kurz) Stone, and one cultivar, P. spuriusMiq. cv. Putat.
@ Hak cipta milik IPB, tahun 2011
Hak cipta dilindungi Undang-undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik ataupun tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB.
BIOSYSTEMATICS OF
PANDANACEAE
IN JAVA
SRI ENDARTI RAHAYU
Dissertation
Submitted as a part of the requirement for the fulfillment of Doctor degree in Biology
THE GRADUATE SCHOOL
BOGOR AGRICULTURAL UNIVERSITY
BOGOR
Examiners in comprehensive examination: Dr. Rugayah MSc
Researcher In Herbarium Bogoriense, LIPI Cibinong
Dr. Rita Megia, DEA
Lecturer in Biology Department, Faculty of Mathematics and natural Sciences, Bogor Agricultural University, Bogor
Examiners in final examination:
Dr. Sri Soedarmiyati Tirtosoedirdjo MSc
Lecturer in Biology Department, Faculty of Mathematics and natural Sciences, Bogor Agricultural University, Bogor
Dr. Ruliyana Susanti MSi
ACKNOWLEDGMENT
This work could not have been accomplished without the kind assistance
of many persons and institutions. I would like to thank to my advisors Prof. Dr.
Ir. Alex Hartana MSc (chairman of the advisory committee), Dr. Ir. Tatik
Chikmawati MSi, Dr. Kuswata Kartawinata and Prof. Dr. Mien A. Rifai
(members of the advisory committee) for their advices, guidance, encouragement
and valuable discussion throughout this study. Prof. dr. Peter C van Welzen of
the National Herbarium Netherlands – Leiden University are gratefully acknowledged for his kind assistance in offering permits and providing facilitation
for my study at Leiden, Dr. Jef Veldkamp for helping to translate German
literatures, Dr. Barbara Gravendeel and MCM Eurlings for allowing me to work
in the laboratory, introducing me with PCR and guiding me how to determine
DNA sequence with sequencer in Van der Klauw Laboratory, Leiden University,
Dr. William J. Baker, Dr. Timothy M.A Ulteridge arranged my visit to Kew
Herbarium and allowing me working withPandanaceaecollection, Dr. Rogier de
Kok from Kew Herbarium for helping me translate German literature, Dr. Tim
Fulcher to let me work with sequencer in the Jodrell Laboratory of Royal Botanic
Gardens Kew. I sincerely thank to Dr. Ir. Dedy Duryadi Solihin, DEA for his
advice, Dr. Eko Baroto Walujo for giving me opportunity to studyPandanaceae
specimens and conduct anatomical study at the Laboratory of Anatomy,
Morphology and Cytology, Herbarium Bogoriense (BO), Cibinong. I Also would
like to thank the Directors and Curators of National Herbarium of Netherland (L)
and Herbarium of Kew Botanical Garden (K) for giving me an opportuninty to
study Pandanaceae specimen kept at L and K, Dr. Irawati for giving me
opportunity to study living collection of Pandanaceae in Bogor Botanical
Garden.
I am grateful to Dr. Rugayah MSc, Dr. Rita Megia, DEA, Dr. Sri
Sudarmiyati Tjitrosoedirdjo, MSc and Dr. Ruliyana Susanti MSi are
acknowledged for their willingness being examiners in my final examination.
Many people have helped me through good advice, suggestions and supports. In
particular. I would like to thank to Dr. Sunaryo, Drs. Erlin Rachman, Eka
technical supports during my anatomical study, Dr. Elizabeth A. Widjaja, for her
kindness in giving the specimens of Freycinetia and giving references of
Pandanaceae, Dr. Arry P. Keim, for his valuable discussion and critics on
Pandanaceae., Dra, Tutie Djarwaningsih, MSi, for her help for publication in
Floribunda, Dr. Fitmawati, Dr. Soaloan Sinaga, Sulassih SP for their help and
technical supports during ISSR study and Dr. Nunik Sri Ariyanti for her help in
making keys.
Rector and the Dean of Fakultas Biologi Universitas Nasional, Jakarta,
Koordinator Kopertis Wilayah III, Rector and Dean of Sekolah Pascasarjana
Institut Pertanian Bogor (IPB) for the opportunity given to me to undertake this
study. Financial support came from BPPS Dikti Depdiknas, Universitas Nasional,
Fundamental research grant from DP2M-Dikti Depdiknas, International
Publication grant from Dikti Depdiknas, Sandwich program from Dikti
Depdiknas, and from Yayasan Damandiri.
I sincerely thank to all staff at Herbarium Bogoriense for sharing their
experties in different fields, all my colleagues in Fakultas Biologi Universitas
Nasional, and all post graduate student of Botany – Taxonomny IPB, Dr. Nurhaidah Iriany Sinaga MSi, Dr. Nursahara Pasaribu MSc., Dr. Pudji Widodo,
Dr. Iris Rengganis and Dra. Ratna Siahaan MSi who always offered good
suggestion and words of hope during the hard times of my study.
Finally, my deepest appreciation is also sent to my parents, my brothers
and sisters, my brothers and sisters in law, especially my husband Drs. Darmanto
and my daughters Dea Anggraini and Della Anggiani for their deep understanding
and great patient and for giving me their moral support.
Bogor, January 2011
CURRICULUM VITAE
Sri Endarti Rahayu was born on 25 August, 1958 in Bandung, West Java.
She spent her childhood with her parents in Bandung until she graduated from the
Padjadjaran University in 1984.
Since 1988 she has been working at Biology Faculty at National
University, Jakarta. In September 1991, she had an opportunity to continue study
at IPB sponsored by TMPD Dikti Depdiknas., and admitted to the degree of MSi
in October 1995.
In September 2004 she had an opportunity to continue study at IPB for her
PhD sponsored by BPPS Dikti Depdiknas. In 2008, she had the opportunity to
follow sandwich program, sponsored by Directorate General of Higher Education
(DGHE) of Indonesia to study Biosystematics of Pandanaceae in Java for three
months at Nationaal Herbarium of The Netherlands (NHN), Leiden University
and one month at Kew Herbarium of Royal Botanic Garden, Kew, UK.
A part of dissertation has been published under two different title articles :
(1) Genetic Diversity of Pandanus and Freycinetia from Java based on ISSR
markers, Floribunda 3(4) : 95 – 103. 2007; (2) A taxonomic study of the Pandanus furcatus andPandanus tectorius complexes with special emphasis on
plants from Java submitted to the 8th International Flora Malesiana Symposium in
TABLES OF CONTENTS
Page
LIST OF TABLES ... ix
LIST OF FIGURES ... x
GENERAL INTRODUCTION Taxonomical Aspects ofPandanaceaein Java and Its Systematic Problems ... 1
Objectives ... 3
MORPHOLOGY OF JAVANESE SPECIES OFPANDANACEAE Introduction ... 5
Materials and Methods ... 6
Results and Discussion ... 6
Morphology of JavaneseFreycinetia ... 6
Morphology of JavanesePandanus ... 12
LEAF ANATOMICAL FEATURES INPANDANUS (PANDANACEAE) FROM JAVA Introduction ... 23
Materials and Methods ... 24
Results and Discussion ... 25
A TAXONOMIC STUDY OF PANDANUS FURCATUSCOMPLEX AND PANDANUS TECTORIUSCOMPLEX WITH SPECIAL EMPHASIS ON PLANTS FROM JAVA Introduction ... 39
Materials and Methods ... 40
Results and Discussion ... 41
GENETIC DIVERSITY OF JAVANESEPANDANUSAND FREYCINETIA BASED ON ISSR MARKER Introduction ... 51
Materials and Methods ... 53
Results and Discussion ... 56
THE DISTRIBUTION AND ECOLOGY OFPANDANACEAEIN JAVA Introduction ... 67
Materials and Methods ... 67
Page
SPECIFIC PART :
A. BIOLOGICAL FLORA OF JAVA :Pandanus tectoriusParkinson 75
B TAXONOMIC TREATMENT ... 87
GENERAL DISCUSSION ... 137
CONCLUSION ... 143
LIST OF TABLES
Page
4.1 Morphological characters of species recognized within
Pandanus furcatuscomplex ... 41
4.2 Summary of contrasting characters of species recognized within Pandanus furcatuscomplex. Leaf base colour, scale bar = 5 cm; Peduncle, scale bar = 13 cm; Cephalia shape, scale bar = 10 cm;
Style shape, scale bar = 1 cm ... 42
4.3 Morphological characters of species recognized within
Pandanus tectoriuscomplex ... 43
4.4 Summary of contrasting characters of species recognized within Pandanus tectoriuscomplex; Prickles on leaf apex, scale bar = 2 cm; Cephalium shape, scale bar = 10 cm ; Phalange apex,
scale bar = 1 cm ... 44
4.5 Polymorphic sites inatpB-rbcL IGS sequence of
Pandanus furcatuscomplex ... 45
4. 6 Polymorphic sites inatpB-rbcL IGS sequence of
Pandanus tectoriuscomplex ... 46
5.1 Thirteen species ofPandanusof Java used in this study ... 54
5.2 Six species ofFreycinetiaof Java used in this study ... 55
5.3 ISSR primers sequence and amplified results on thirteen species of
Pandanusof Java ... 58
5.4 ISSR primers sequence and amplified results on six species of
Freycinetiaof Java ... 58
5.5 Nei and Li’s similaritymatrix of thirteenPandanusspecies of Java
by using ISSR primers ... 59
5.6 Nei and Li’s similarity matrix of sixFreycinetia species of Java
by using ISSR primers ... 60
LIST OF FIGURES
Page
2.1 Habit ofFreycinetiaspp.: A.F. javanica, scale bar = 15 cm;
B. F. sumatrana, scale bar = 2 m. ... 7
2.2 Auricles type ofFreycinetiaspp.: A. Membranous (F. javanica) B. Coriaceous (F. insignis).Auricles was shown with arrow. Scale bar = 2 cm ... 9
2.3 Inflorescence type ofFreycinetiaspp.: A. Umbel (F. javanica), B. Racemiform (F. angustifolia), C. Lateral (F. funicularis).
Scale bar = 1 cm ... 11
2.4 Berries type ofFreycinetiaspp.: A. Obconic (F. angustifolia)
B. Lageniform (F. insignis), scale bar = 25 mm ... 12
2.5 Habit ofPandanusspp.: A. Shrub(P. amaryllifolius), scale bar = 10 cm; B. Tree (P. odoratissimus.), scale bar = 1 m ... 13
2.6 Prop root type ofPandanusspp.: A. Slender (P. spuriuscv. Putat): B. Stout (P. odoratissimus); C. No prop root (P. kurzii);
D. Muricate in longitudinal line (P. utilis). Scale bar for A = 75 cm, Scale bar for B, C and D = 5 cm ... 14
2.7 Prickles type ofPandanus leaves: A. White prickles (P. tectorius
var. littoralis); B. Red prickles (P. utilis), scale bar = 1 cm ... 15
2.8 Inflorescence type ofPandanusspp.: A. Staminate inflorescence erect (P. labyrinthicus); B. Long staminate inflorencence (P. utilis); C. Pistillate inflorescence in spike (P. odoratissimus); D. Raceme of spikes (P. polycephalus). Scale bar for A and B = 10 cm, scale bar for C and D = 2 cm ... 18
2.9 Stamen types ofPandanusspp.: A. Sessile(P. tectoriusvar. littoralis) B. Pedicellate (P. utilis), scale bar = 2 cm ... 18
2.10 Stigma type ofPandanusspp.: A. Sessile (P. utilis), B. On style
(P. scabrifolius), scale bar = 1 cm ... 19
Page
3.1 Light micrographs of leaves: (A)- transverse leaf section ofP. multifurcatusFagerl.; (B)- elongated abaxial epidermal cells of P. bantamensisKoord.; (C)- undulate adaxial anticlinal cell walls ofP. kurziiMerr ; (D)- adaxial epidermis cells with cubical crystals ofP. bantamensisKoord.;(E)- adaxial stomata with lobed papillae ofP. utilisBory; (F)- costal zone in the abaxial epidermis of P. amaryllifoliusRoxb. Scale bar for A & F = 100 µm.; Scale bar
for B, C and D = 50 µm; scale bar for E = 20 µm ... 26
3.2 Light micrographs of leaves: (A)- abaxial epidermal cells of P. dubiusSpreng.; (B)- abaxial papillae in lateral subsidiary cells of P. tectoriusvar.littoralis; (C)- abaxial papillae on epidermal cells of P. spuriusMiq. cv Putat; (D)- hypodermis ofP. labyrinthicusKurz; (E)- crystal cells in outermost hypodermis ofP. pseudolaisWarb.; (F)- circular palisade cells ofP. amaryllifoliusRoxb. Scale bar for
A, D, E. & F = 50 µm.; scale bar for B & C = 20 µm ... 29
3.3 Light micrographs of leaves: (A)- sclerenchyma in spongy tissue of P. tectoriusParkinson cv.Sanderi, (B)- raphide sacs in internal palisade ofP. amaryllifoliusRoxb.; (C)- unspecialized abaxial stomata ofP. tectoriusParkinson cv. Sanderi ; (D)- abaxial papillose lateral subsidiary cells ofP. odoratissimusL.f ; (E)- abaxial papillose terminal and lateral subsidiary cells ofP. scabrifoliusMartelli; (F)- abaxial papillose neighbouring and subsidiary cells ofP. kurzii Merr.; (G)- abaxial, overarching, lobed or dendritic papillae ofP. utilis Bory. Scale bar for A & B = 100 µm, scale bar for C, D, E, F &
G = 20 µm ... 31
5.1 ISSR profile ofPandanusspp and Freycinetiaspp of Java using primer ISSR4;M : Marker, S1:P. bantamensis, S2:P. tectoriusvar.
littoralis, S5:P. odoratissimus, S6:F .javanica, S7:F. angustifolia, S14:P. nitidus,S15:F. angustifolia, S20:F. sumatrana,
S22:F. imbricata, S24:P. kurzii, ... 57
5.2 ISSR profile of Pandanusspp and Freycinetiaspp of Java using primer ISSR7.; M: Marker, S3:P. bidur,
S4:P. spinistigmaticus,S8:P. pseudolais, S13:P. multifurcatus S14: P. nitidus, S21:P. amaryllifolius, S23:P. polycephalus,
S24:P.kurzii, S25:P. scabrifoliusand S26: P.dubius... 57
5.3 UPGMA dendrogram based on ISSR analysis of thirteen
Pandanusspecies of Java ... 60
5.4 UPGMA dendrogram based on ISSR analysis of sixFreycinetia
Page
5.5. Principal component analysis for thirteen JavanesePandanus
based on ISSR data ... 62
5.6. Principal component analysis for six JavaneseFreycinetiabased
on ISSR data ... 63
6.1 Diversity ofPandanaceaein different habitat in Java, BF (Beach Formation); LRF (Lowland Rain Forest); HF (Hill Forest); LMF
GENERAL INTRODUCTION
Taxonomical Aspects ofPandanaceaein Java and Its Systematic Problems
Pandanaceae was first recognized by Robert Brown in 1810. The
Pandanaceae, the sole representative of the Pandanales, is arborescent or
scandent monocotyledons confined to the Old World tropics and subtropics (Cox
1990). Pandanaceae is a large family, consist of 4 genera: Freycinetia Gaud.,
Pandanus Parkins., Sararanga Hemsl., and Martellidendron Callm. & Chassot
(Callmanderet al. 2003).Pandanaceaecommonly known as screw pine or screw
palm has a unique growing form. The large serrated leaves up the trunk are
formed in a circular motion, giving it the screw like look. The family is important
in several regions, wherein it has contributed to the fundamental structure and
physiognomy of the vegetation (Stone 1983b).
Pandan family (Pandanaceae) is represented in Java by two genera:
Freycinetia Gaud. andPandanus Parkins. (Backer and Bakhuizen van den Brink
1968; Stone 1972). The genus Freycinetia was described in 1824 by the French
botanist Gaudichaudii (Stone 1970), and may be called climbing pandans. This
genus has been known to occur in Java for a long time, since most of the oldest
taxa nomenclaturally speaking are typified by specimens from Java (Stone
1968), and according to Stone (1970) the real home ofFreycinetiaamong others
is Java. The name Pandanus originates from the Malay word pandan, it was
latinized and first published by George Eberhard Rumpf (Rumphius) whose
Herbarium Amboinensis of 1743 contains the description of eleven species of
which eight are illustrated (Stone 1965). Java harbours a large number of species
of Pandanus,not less than fifteen species of Pandanus are mentioned in the key
given by Backer and Bakhuizen van den Brink (1968). It becomes evident that
Pandanusin Java is rich in species, considering the widely variable morphology
The last attempt at a comprehensive treatment of Pandanaceae of Java
was Backer and Bakhuizen van den Brink (1968) in their “Flora of Java” in which
they recognized seven species of Freycinetia and fifteen species of Pandanus.
Since a short visit to Hortus Bogorienses by Stone (1972), there were no further
exploration on the pandan flora of the island have been made, thus the pandan
flora remains largely unknown.
Since a large number of morphological characters are now known for
Freycinetia and Pandanus species, it appears useful to consider their use in
identifyingPandanaceaeof Java, because morphological data are regarded as the
most appropriate and the most rapid mean for identification and for constructing
map of diversity of plant (Davis & Heywood 1963). Morphological characters
have the great advantage over other characters that we can see the plant variability
easier. Unfortunately complete material is not always available for the various
numbers of a group being studied, and sometimes we need to identify incomplete
material or small fragment of material. In this case we use anatomical characters.
It is now realized that anatomical characters is as valuable as morphological ones
(Stace 1989). Some of these anatomical characters are so diagnostic that they are
now commonly used in identification or identification of fragment of plants
(incomplete material). Valuable taxonomic evidence has been obtained from the
study of leaf epidermis and stomata ofPandanus, and the appearance of particular
anatomical characters seems sufficiently constant. In this study we are able to
propose anatomical key to species forPandanusin Java. Taxonomical problem as
far as the Java pandans concerned, are centered not only on the species status ofP.
odoratissimusandP. tecroriusvar.littoralis, which are considered as a synonym,
but also the species status which are given as synonyms of Pandanus furcatus
Roxb. Backer and Bakhuizen van den Brink (1968) included P. bantamensis
Koord.,P. oviger Martelli,P. pseudolaisWarb., and P. scabrifoliusMartelli into
one species ofP. furcatusRob. This classification is in contrast with Stone (1972)
who stated that these species were regarded as four different species. In this study
P. pseudolaisWarb, P. scabrifoliusMartelli. Therefore an effort has been made
to redescribe these species in detail, using morphological, anatomical and
molecular data, i.e sequence data of atpB-rbcL IGS since a satisfactory
classification depends upon the interpretation of as many characters as possible
(Davis & Heywood 1963). Molecular data can be useful in solving different kinds
of taxonomic problem, and choloroplast marker such as atpB-rbcL IGS can be
particularly valuable at lower taxonomical level (Soltis & Soltis 1998).
The pandans in general tend to be horticulturally interesting and often
curious or unusually elegant, and tend to cause considerable public interest. The
species that are already completely well known in cultivation are few, viz. P.
dubius, P. spurius cv. Putat, P. tectorius cv. Sanderi and P. utilis. In view of its
importance and considering the large number of wild species germsplasm
available for Freycinetia and Pandanus in Java, the genetic analysis through
molecular marker is a prerequisite to have a deep insight of the genome
organization in the wild species. It is imperative, threfore to establish strategies
for the preservation of Freycinetia and Pandanus germplasm. In this study,
genetic diversity have been assessed using ISSR marker. This analysis is a
preliminary step to ensure the conservation and the development of genetic
resources, and to know the most important location to conserve Pandanaceae in
Java, we studied the distribution and ecology ofPandanaceaein Java.
Every chapter was written in different style, following the format of the
journal we are going to publish these articles, e,g. the article of Biological Flora of
Java:Pandanus tectoriusParkinson is in preparation for publication in Journal of
Ecology.
The objectives of the study
Our objective were to unravel species which is having taxonomical
problem. A combined morphological, anatomical and molecular marker was
undertaken to evaluate the suitability of the species concept; to determine genetic
MORPHOLOGY OF JAVANESE SPECIES OF
PANDANACEAE
Introduction
The last attempt at a comprehensive treatment of Pandanaceae of Java
was Backer and Bakhuizen van den Brink (1968) in their “Flora of Java” in which they recognized seven species of Freycinetia and fifteen species of Pandanus.
Since a short visit to Hortus Bogorienses by Stone (1972), there were no further
exploration on the pandan flora of the island have been made, thus the pandan
flora remains largely unknown.
Since a large number of characters are now known for Freycinetia and
Pandanusspecies, it appears useful to consider their use in identifying plants from
Java. The species are invariably classified by the feature of the female for several
reasons (Kam 1971). First, the male flowers are very short-lived, generally they
bloom and decay within 2 or 3 days. Furthermore, there seem to be a higher
percentage of females in any given population. Thirdly, the fruit offers a greater
number of useful diagnostic features. The male inflorescences are generally
strongly scented, and sometimes offer powering fragrance. The male flowers
themselves are simply masses of stamens gathered into units representing
branching systems of various ranks. Fieldwork carried out for this study has
provided stronger foundation for understanding morphological variation within
the species.
Characters of leaf shape, leaf apex, the morphology of leaf auricles and
type of pistillate inflorescence were found useful in delimitation and identification
of Javanese Freycinetia, while characters of habit, the surface of stem, present or
absent of prop root, the surface of prop root, the leaf shape, leaf apex, the
armature of leaf margins and midrib, the colour of leaf margin and midrib teeth,
the distinctness or indistinctness of tertiary cross vein, present or absent of apical
ventral pleats, phalange shapes, the position of infructescence, position of seed
chamber and stigma shape are proved useful for distinguishing among species of
This study was undertaken to have a better understanding on the
morphology of the family in order to make a better species delimitation,
particularly for species found in Java.
Materials and Methods
Morphological study on Javanese species of Pandanaceae has been conducted.
This study was based mainly on available specimens at the BO, K and L and new
collection specimens obtained from field work in different location in Java. In
addition, living plants grown in botanical garden were also studied. Five species
that was planted in Bogor Botanical Garden, viz. Pandanus kurzii, P.
labyrinthicus, P. multifurcatus,P. polycephalusandP. spinistigmaticus were also
examined. The process of undertaking in this study followed the methods
described by Rifai (1976) and Vogel (1987).
Basic morphological characters such as habit, stem, leaves, inflorescence,
staminate flower, pistillate flower, fruit and their details were used to describe and
recognize taxa; all morphological data was used for producing the description of
each taxon, while the key to species was constructed from the diagnostic
characters only.
The morphological species concept was applied as a framework to define
taxa, in which distinction is based on perceived discontinuities in morphological
variation (Davis & Heywood 1963).
Results and Discussion
Morphology of JavaneseFreycinetia
Habit
All species of Freycinetia found in Java are climber with different size.
Three species of Freycinetia (F. imbricata, F. javanica and F. scandens) are
smallish to medium climber, while F. insignis and F. sumatrana are medium
climbers, and the smallish climber is found inF. angustifolia.
The stems are found hanging on trees and branch richly, such as found in
F. angustifoliaandF. javanica(Fig. 2.1 A); while inF. imbricatathe main stem
makes the trunk of the host tree invisible, such as found in F. insignis and F.
sumatrana(Fig.2.1 B).
Figure 2.1 Habit ofFreycinetiaspp.: A.F. javanica, scale bar = 15 cm B. F. sumatrana, scale bar = 2 m.
Stem
Stems of JavaneseFreycinetiavary in size, nodes and colour. In term of
size, F. angustifolia, F. imbricata, F. javanica and F. scandens are included in
slender species, whileF. funicularis, F. insignisandF. sumatranaare included in
robust species. Variation is observed in stem shape. This study showed that F.
angustifolia, F. imbricata,F. insignis, F.scandens, andF. sumatranahad terete
stem, whereasF. javanicahad subterete to terete stem.
Variation is also observed in internode shape. This study showed that F.
angustifolia, F. imbricata, F. insignis, F. javanica, and F. sumatranahad terete
internode, whereas F. scandens had subterete to terete internode. The surface of
the stem can be sulcate or sulcate to canaliculate. The colour also varies from
yellowish, greenish brown to reddish brown.
Leaves
The leaves ofFreycinetiaare usually green to dark green coloured on the
upper surface, but paler green on the lower surface. In Java F. angustifolia
possesses the smallest and most slender leaves, whereas F. sumatrana has the
most robust leaves. The leaves arrangement in species of Freycinetia found in
Java is alternate and imbricate. Four species of Freycinetia (F. angustifolia, F. A
A
funicularis,F. javanicaandF. scandens) have alternate leaves, and the other three
species (F. imbricata,F. insignisand F. sumatrana) had imbricate leaves.
Leaves are simple blade and usually linear-lanceolate in outline, but
variation do exist in some species.F. scandenshave variation in leaf shape, from
elliptic, oblanceolate to lanceolate. Leaf apex can be abruptly attenuate below the
apex to gradually attenuate toward the subulate apex, whereas in F. sumatrana it
is long tapering to a slender subulate tip. The margin usually was armed in basal
part, apical part and upper midrib of its leaves with serrate prickles. The basal part
ofF. insignisandF. scandens are denticulate, and basal part of F. sumatrana is
dentate. The colour of the prickles may be wholly straw colour or with brown
tipped as in F. angustifolia. The laminar part can be chartaceous, subcoriaceous
to coriaceous.
Auricles
The auricle is an organ of flange-like extensions found on the leaf-sheath,
and can be easily seen on the young leaf. Auricles are usually membranous (Fig.
2.2 A), transparent, fragile or coriaceous (Fig.2.2 B). Auricles can be regarded as
a good identification character of Freycinetia in the field, because in herbarium
specimen they are hardly seen in fine condition. The shape, size, texture, nature of
margin, nerves and colour of auricle vary between species. Freycinetia
angustifolia possess the smallest auricle, while F. sumatrana possess the biggest
auricle. Colour of auricles also varies from pale green in F. insignisto brownish
green–brownis yellow in F. javanica.
InFreycinetia, they presented in varying from tapered or rounded to the
apex or adnate to the apex. The margin is entire or denticulate to spinulose at apex
(F. insignis), or armed almost to the base (F. imbricata). The lamina is usually
membranous inF. javanica(Fig. 2.2 A) or coriaceous inF. insignis(Fig. 2.2 B),
some of them fragmenting transversally in F. sumatrana or fragmenting laterally
in F. scandens, with 4 widely spaced nerved in F. insignis or 1-2 septate in F.
Figure 2.2 Auricles type ofFreycinetiaspp.: A. Membranous (F. javanica) B. Coriaceous (F. insignis).Auricles was shown with arrow. Scale bar = 2 cm
Bracts
Bracts are persistence or caduceus, located on the peduncle. Bracts vary in
shapes and sizes. Bracts are usually ovate, cymbiform to lanceolate, with entire
margin to slightly armed with prickles. The sizes of bracts are in concurrent with
their habits. Slender species like F. angustifolia, F. imbricata and F. javanica
possess minute bracts, while robust species such as F. insignisandF.sumatrana
have robust bracts as well. Their apex have various shapes, varying according to
the species from acute to acuminate in F. imbricata, acute to cuspidate in F.
javanica) mucronate to aristate in F. angustifolia, and aristate with aculeate
prickles inF. sumatrana.
Peduncle and pedicel
The peduncle usually is straight and short. The peduncle may be slender or
robust, densely or sparsely pubescent or rarely glabrous, only in F. insignis the
peduncle had bract scar. Usually the pedicell is longer than peduncle. The pedicel
shape usually is half-terete, slender (F. scandens), usually as thick as peduncle,
or only somewhat stouter than the peduncle with 0.2-1.5 cm diameter (F.
insignis). The surface of pedicels can be glabrous or covered with indumentum.
F. angutifolia, F. javanica and F. scandens had glabrous pedicel, while F.
imbricataandF. insignishad puberolous to hirsute pedicell.
Inflorescentia
The inflorescence in most ofFreycinetiafound in Java are usually located
on terminal part of the stems (i.e terminal inflorescence). Lateral inflorescence
are less often seen. This study showed that there is one species that possess such
feature, viz.F. funicularis. In lateral inflorescence, not only bract that is present,
but also prophyll, that is located in the upper part of bracts.
Freycinetia is mostly dioecious, which means that male and female
flowers are formed on different plant. Staminate inflorescence is invariably
terminal on a normal leafy shoot, usually a raceme of spikes, each of the three or
four spikes. Staminate inflorescence are more rarely seen than that of pistillate
inflorescence, due to the anthesis which takes only one to three days (Stone
1983b). Similar to other genera of Pandanaceae, Freycinetia identification is
therefore mainly based on the structure of female inflorescence and infruitescence.
In Freycinetia, the spikes (both male and female) tend to be closely
adjacent and often ternate, so that the open ripe inflorescence seems to be an
umbel like in F. javanica (Fig. 2.3 A) or pseudoumbel. In F. angustifolia, the
inflorescence is racemiform (Fig. 2.3 B), whereas in F. funicularis, the
inflorescence is lateral (Fig. 2.3 C).
Male flowers (Stamen)
The stamens are simple, without branched filaments, and each terminate is
in a small rather short anther or sessile stamens, and staminate pedicel is absent.
Female flowers (Pistil)
The pistil consists of multiovulate carpel with carpels separated to base or
carpel united. Each carpel tipped by a stigma, and the stigma is sessile. The
stigmas vary in number and shape, from 2 to 5 in number, and from ovate (F.
javanica), depressed ovate-suborbicular (F. angustifolia, F. imbricata, and F.
insignis), pentagonal-suborbicular (F. scandens) or protude (F. funicularis) in
Fig.2.3 Inflorescence type ofFreycinetiaspp.: A. Umbel (F. javanica), B. Racemiform (F. angustifolia), C. Lateral (F. funicularis). Scale bar = 1 cm
Cephalia
Cephalium (plural: cephalia) is the complex fruit in Pandanaceae. Some
of the important morphological character inFreycinetiaare found in the cephalia.
In the Javanese species of Freycinetia cephalia vary in shapes, size and colours.
Four species (F. funicularis F. insignis, F. javanica, and F. sumatrana) have
cylindric cephalia. F. angustifolia have cylindric to oblanceolate cephalia, F.
scandens have cylindric to oblong cephalia, and F. imbricata have narrowly
elliptic to broadly oblong cephalia. The number of cephalia per inflorescence is
usually three, two or four.
A B
Berries
Berry is the simple fleshy fruit ofFreycinetia. InFreycinetia a cephalium
consisted of numerous berries. A berry contain many fused ovules, thus a
multiovulate fruit, while inPandanusthe simple fruit is always uniovulate. In this
circumstances a cephalium can be regarded as a complex fruit. The shape of
berries vary from obconic like in F. angustifolia (Fig. 2.4 A) obovate (F.
funicularis), oblong (F. javanica) and subpyramidal such as in F. sumatrana.
Three species, viz. F. imbricata, F. insignis(Fig. 2.4 B) andF. scandens are the
species observed with various shapes of berry from pentagonal to lageniform,
pentagonal, lageniform to oblong, and pentagonal, lageniform, ovate to oblong
respectively. The apical part of a berry is usually harder and stiffer, while the
basal part is usually fleshy.
Fig.2.4 Berries type ofFreycinetiaspp.: A. Obconic (F. angustifolia) B. Lageniform (F. insignis), scale bar = 25 mm
Morphology of JavanesePandanus
Habit
UnlikeFreycinetiathat have climbing habit,Pandanusspecies habit found
in Java vary from shrub to tree. Shrub species are P. amaryllifolius(Fig. 2.5 A),
P. kurzii, P. nitidusandP. polycephalus. Tree species include P. bantamensis,P.
bidur, P. dubius, P. faviger. P. labyrinthicus, P. leram var. andamanensium, P.
multifurcatus, P. odoratissimus) (Figure 2.4 B),P. pseudolais, P. scabrifolius, P.
spinistigmaticus,P. spuriuscv Putat,P. tectoriusandP. utilis.
Figure 2.5 Habit ofPandanusspp.: A. Shrub(P. amaryllifolius), scale bar = 10 cm ; B. Tree (P. odoratissimus.), scale bar = 1 m
Stems
Stems of Javanese Pandanus vary in size, colour and branch. Slender
stem is found in P. amaryllifolius or short stem in P. kurzii. Erect stem and
unbranched are found in P. bantamensis,P. dubius andP. pseudolais, while P.
labyrinthicus, P. multifurcatus, P. nitidus, P. odoratissimus and P.
spinistigmaticus had spreading branches, whereas P. faviger, P. tectorius var.
littoralis and P. utilis had dichotomous branching. Surface of stem can be
smooth likeP. kurzii, sulcate inP. amaryllifolius, abundant rootlet to thorny in
P. bidur, P. dubius, P. labyrinthicus, P. multifurcatus, P. nitidus, P.
odoratissimus, P. spinistigmaticus, and P. tectorius var. littoralis or ringed by
leaf scars in P. polycephalusandP. utilis. Colour of stem also varies from grey
to green or brownish.
Prop root
A prop root is a root formed from the stem, usually close to the ground
which helps hold the stem erect and anchor the plant. There are variation in prop
root sizes, number and surfaces. Prop root can be small and few in P.
amaryllifolius, small and abundant in P. labyrinthicus, P. odoratissimus and P.
tectoriusvar.samak, slender in P. spurius cv Putat (Fig. 2.6 A), some arise from
lateral branches inP.nitidusandP. spuriuscv Putat, long and stout as inP. bidur,
P. dubius, P. faviger, P. leram var. andamanensium, P. multifurcatus, P.
odoratissimus(Fig. 2.6 B) andP. spinistigmaticus or absent in P. kurzii(Fig. 2.6
C). Surface of the prop root can be smooth in P. scabrifolius, muricate in
longitudinal line such as in P. utilis (Fig. 2.6 D), P. tectorius var. littoralis, P.
pseudolais, andP. nitidus or armed with prickles or spine inP. spinistigmaticus,
P. polycephalusand P. multifurcatus.
Figure 2.6 Prop root type ofPandanusspp.: A. Slender (P. spuriuscv. Putat): B. Stout (P. odoratissimus) ; C. No prop root (P. kurzii) :
D. Muricate in longitudinal line (P. utilis). Scale bar for A = 75 cm, Scale bar for B, C and D = 5 cm.
Marginal Spine
Leaves usually set with prickles along its length or part of it. The prickles
decrease in size nearing the leaf apex (P. utilis, P. bantamensis, P. pseudolais, P.
tectoriusvar. littoralis, andP. nitidus), always antrorse (ascending) but those near
leaf base sometimes retrorse, or absent. Their margin are usually prickly,
A B
occasionally entire (P. spuriuscv. Putat), or prickly very near and at apex only (P.
amaryllifolius), or occasionally smaller at leaf base, larger at midsection and
decreasing in size nearing the leaf apex (P. bidur, P. dubius, P. odoratissimus,
andP. scabrifolius). Twin lateral pleats are often well distinguished, smooth (P.
bantamensis, P. nitidus, P. pseudolais, and P. scabrifolius), or in some species
prickly serrate or absent (P. odoratissimus, P. spinistigmaticus, P. spurius cv.
Putat, and P. tectorius var. littoralis). Some species have white spine in P.
tectorius var. littoralis (Fig. 2.7 A), green prickles with brown tipped (P.
bantamensis, P. pseudolais, andP. scabrifolius) or yellowish green (P.dubius) or
red prickles inP. utilis(Fig. 2.7 B).
Figure 2.7 Prickles type ofPandanus leaves: A. White prickles (P. tectorius var. littoralis) ; B. Red prickles (P. utilis), scale bar = 1 cm.
Leaves
The leaves of Pandanus are usually dark green coloured, glossy and
glaucous on upper (adaxial) surface, but paler green on the lower surface, whereas
in P. tectoriuscv. Sanderi have longitudinal of white to pale yellow band from
centre to leaf margin. Compared with the other members ofPandanaceae, the
size of leaf in Pandanus is noticeably longer and bigger. In Java P.
amaryllifolius has the smallest and most slender leaves, whereas P. pseudolais
possess the longest leaves (299-574.5 cm long). The leave arrangement in all
species of Pandanus found in Java is spiral. The leaves are simple and usually
ensiform to linear in shapes. However variation do exist as in some species, the
leaves are observed in ligulate such as found inP.labyrinthicus, P. multifurcatus,
P. odoratissimus and P. spurius cv. Putat. The apical part can be abruptly
terminating in a point to gradually long tapering to subulate apex. Caudate apices
are observed only in one species, i.eP. dubius. The margin can be entire as inP.
spuriuscv. Putat or noticeably armed with spines throughout the length, except in
P. amaryllifoliuspossess prickles only in the apical part of its leaves. The laminar
part can be chartaceous, thin coriaceous, coriaceous to thick coriaceous.
Longitudinal lines are more prominent abaxially, with tertiary cross vein on both
surfaces. In P. amaryllifolius, P. bantamensis, P. bidur, P. dubius, P. kurzii, P.
pseudolais, and P. scabrifolius tertiary cross vein form a network of meshes,
oblong or rhombic meshes, whereas in P. leram var. andamanensium tertiary
cross vein makes tesselate appearance. The basal part of leaves of P. kurzii, P.
polycephalus and P. scabrifolius are whitish, reddish in P. utilis, or reddish
brown inP. bantamensis, and P. pseudolais.
Bracts
Bract are covering inflorescence and usually formed by the three levels of
trichiously arranged bracts. The interior bract covers the inflorescence, while the
other two namely exterior and middle bracts protect the interior bract and the
inflorescence inside.
Bracts vary in shapes and sizes. Bracts are usually lanceolate in shape,
with margins armed with prickles to spines. The sizes of bracts are in concurrent
with their habits. Robust species such as P. bantamensis, P. pseudolais and P.
scabrifolius have robust bracts as well, while slender species like P. nitidus
possess slender and minute bracts. The exterior bracts are leaf-like in forms,
except inP. kurziithe exterior bracts are cymbiform in shape. The inner bracts are
shorter and lighter in colour.
The colour of bracts also varies. The colour of exterior and interior bracts
can be different. The differences can be seen within the same bract. The colour of
the apical part, middle and basal parts may not be uniform such as in P.
odoratissimus, and in few species the colour of bracts are uniform such as inP.
polycephalus (white) and P. utilis (green). The apex of bracts are observed
abruptly acute inP. utilis, gradually tapering to a subulate tip inP. labyrinthicus,
Inflorescentia
Pandanus are dioecious, which means that male and female flowers are
produced on separate plants. The inflorescence in all species ofPandanusin Java
is usually found in apical part of the stem (i.e terminal inflorescence). Staminate
inflorescences are fragrant and usually pendent, except in P. labyrinthicus
staminate inflorescence is erect (Fig. 2.8 A). Staminate inflorescence is also varies
in sizes. In P. odoratissimusflowering part of spike is 21.3-33 cm long and bear
9-18 lateral racemes, whereas inP. utilisflowering part of spike is 41-43 cm long
and bear 20 lateral racemes (Fig. 2.8 B). Staminate inflorescence are more rarely
seen than that pistillate inflorescences, thereforePandanusidentification is mainly
based on the structure of female inflorescences and infructescence.
Pistillate inflorescences may be represented by a single cephalium (head)
as in P. odoratissimus (Fig. 2.8 C) or by s spike of several cephalia. In P.
favigerthe pistillate are observed spike of 3 cephalia, whereas inP. polycephalus
pistillate inflorescence is raceme of spike (Fig. 2.8 D).
Peduncle
The peduncle is stout and glabrous, generally straight, whereas in P.
bantamensis,P. tectoriusvar. littoralis, andP. utilisthey are slightly curved at the
end; in P. pseudolais it is curved at the end. Their shapes are usually 3-sided,
whereas in P. polycephalus it is obtusely trigonous. Their size is larger at apex
and decrease nearing the base. Their colour is varied according to the species from
whitish green (P. bantamensis) to green (P. pseudolais, andP. utilis).
Male Flower (Staminate)
The stamens are to be borne in small or large clusters on short or long axes
(P. labyrinthicus, P. odoratissimus, P. spurius cv. Putat, P. tectorius var.
littoralis, andP. utilis). Most of the male flowers are sessile (Fig. 2.9 A), whereas
Figure 2.8 Inflorescence type ofPandanusspp.: A. Staminate inflorescence erect (P. labyrinthicus); B. Long staminate inflorencence (P. utilis); C. Pistillate inflorescence in spike (P. odoratissimus); D. Raceme of spikes (P. polycephalus). Scale bar for A and B = 10 cm, scale bar for C and D = 2 cm.
Figure 2.9 Stamen types ofPandanusspp.: A. Sessile(P. tectoriusvar. littoralis) B. Pedicellate (P. utilis), scale bar = 2 cm
C D
A B
Female Flower (Pistillate)
Female flowers consist of a single uniovulate carpel with a single stigma.
The stigma may be sessile as in P. bidur, P. dubius. P. odoratissimus, P.
polycephalus, P. tectorius var. littoralis, and P. utilis (Fig. 2.10 A) or borne on
style in P. bantamensis, P. kurzii, P. nitidus, P. pseudolais, and P. scabrifolius
(Fig. 2.10 B). The stigmas have various shapes, varying according to the species
from linear (P. kurzii) or rounded (P. polycephalus), cordate, elliptic, lanceolate,
lip-like (P. bidur) or separating in the forkes (P. bantamensiis, and P.
scabrifolius).
Figure 2.10 Stigma type ofPandanusspp.: A. Sessile (P. utilis), B. On style (P. scabrifolius), scale bar = 1 cm
Cephalia
Some of the important morphological character for species identification
in Pandanus are in their cephalia (singular, cephalium). In Javanese species of
Pandanuscepalia vary in shapes, sizes and colours. Five species, i.eP. kurzii, P.
odoratissimus, P. scabrifolius, P. tectorius var. littoralis and P. utilis have
subglobose cephalia, while the other species such as inP. bantamensis,P. nitidus,
P. polycephalus and P. pseudolais possess ellipsoid to oblong cephalia. The
number of cephalia per infruitescence is usually one, other numbers such as three,
five or six are also present but less seen.
In Pandanus, a cephalium can be consisted of numerous drupes or
phalanges. Drupes is composed of free, a single carpel while phalanges
(polydrupes) are carpels that are permanently fused into compound structure.
Drupes or phalanges are of great importance in Pandanus classification. The
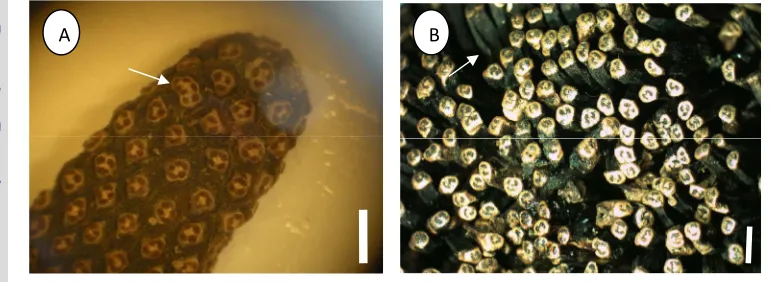
result of this study showed that seven species. i.e P. bantamensis (Fig. 2. 11 A
dan B), P. faviger, P. kurzii. P. nitidus, P. polycephalus, P. pseudolais and P.
scabrifolius have cephalium consisted of numerous drupes, while the other
species such as in P. bidur, P. dubius, P. leram var. andamanensium, P.
odoratissimus, P. tectorius var.littoralis andP. utilis (Fig. 2.11 C and D) have
cephalia consisted of numerous phalanges. Drupe varies in shape from conical,
cuneate to oblanceolate, while phalange shapes vary from clavate, cuneate,
obovate to oblong.
Drupes usually have one stigma while phalanges (polydrupes) have more
than one stigma. Some species have sessile stigma, while in other species stigma
is on style. The surfaces of cephalium can be smooth, such as in P. utilis or
covered by flat scales like inP. kurzii (Fig. 2.11 E), while in the surface of the
apical phalanges of Pandanus usually have no cracks on centre apical sinuses,
except in Pandanus tectorius var. littoralis, there are cracks on the centre of
apical sinuses on its the surface apical of the phalange (Fig. 2.11F ). The apical
part of drupe of phalange is usually harder, and the basal part is usually fibrous
Figure 2.11 Cephalia type ofPandanusspp.: A. Cephalia consist of numerous drupes (P. bantamensis); B. Drupe (P. bantamensis); C. Cephalia consist of numerous phalanges (P. utilis); D. Phalange (P. utilis); E. Cephalia covered by flat scales (P. kurzii); F. Apical phalange with cracks on the centre of apical sinuses (P.tectoriusvar. littoralis). Scale bar for A and C = 20 cm.; scale for B, D, E and F = 1 cm.
A B
C D
Conclusion
In this study, based on gross morphological characters, seven species of
Freycinetia can be recognized, viz. F. angustifolia Bl., F. funicularis Merr., F.
imbricataBl., F. insignis Bl., F. javanica Bl., F. scandensGaud., F. sumatrana
Hemsl; and sixteen species of Pandanus, viz. P. amaryllifolius Roxb., P.
bantamensis Koord., P. bidur Jungh., P. dubius Spreng., P. faviger Backer, P.
kurziiMerr., P. labyrinthicus Kurz, P. multifurcatus Fagerl., P. nitidus Kurz,P.
odoratissimus L.f., P. polycephalus Lam., P. pseudolais Warb., P. scabrifolius
Martelli.,P. spinistigmaticusFagerl., P. tectoriusParkins, with two varieties, viz.
P. tectorius var. littoralis, P. tectorius var. samak, one cultivar, i.e Pandanus
tectorius cv. Sanderi, P. utilis Bory; one variety P. leram Jones var.
andamanensium (Kurz) Stone, and one cultivar, P. spurius Miq. cv. Putat are
LEAF ANATOMICAL FEATURES IN
PANDANUS
(PANDANACEAE) FROM JAVA
Introduction
Pandanus Parkinson is characterized by a number of very obvious
features, forming a unique combination of characters. These distinctive characters
are generally an erect stem (sometimes sprawling), basally supported by many
long stilt and prop roots, branching more or less dichotomously, usually rather
prickly because of short, blunt or sharp specialized adventitious roots; leaves in 3
regular, close-set spirals, on the rounded or slightly 3-angled trunks, leaves entire,
long, narrow, deeply channelled along the midrib and pleated once on each side,
the tip, margin, midrib (below) and the pleats (above, sometimes) set with stout or
fine prickles (Stone 1965).
Pandanus, with three other genera,FreycinetiaGaud., SararangaHemsl.,
and Martellidendron Callm. & Chassot form the Pandanaceae. Pandanus
contains more than 600 species which are distributed throughout the tropics of the
Old World and this large genus is internally very diverse (Kam 1971). The last
revisions of thePandanaceaeof Java were made by Backer & Bakhuizen van den
Brink (1968) and Stone (1972). They recognized 16 species and produced some
classifications, which invariably used morphological characters of the pistillate
flowers and fruits as diagnostic features. It is, therefore virtually impossible to
identify sterile or staminate plants (Kam 1971).
Edeoga and Ikem (2001) showed that leaf epidermal features are as useful
in systematic botany as for instance DNA sequences or chemical compounds. The
taxonomic value of leaf anatomical features was considered in some detail by
Stace (1965). Examining the shape of epidermal, guard and subsidiary cells of
stomata may prove useful for the identification of selected plant species (Jakubska
2007). Stone (1976) constantly reaffirmed that the variation in the epidermal
tissue (including stomata) is of great value in systematic botany. Tomlinson
(1965) founded a classification of stomatal types based on progressive
complexity, and this system was used by Kam (1971) to demonstrate that there is
Pandanus.The aims of the present study are to describe the epidermal variation in
15 Javanese species ofPandanusand to evaluate the usefulness of the characters
for identification and classification purposes.
Materials and Methods
The survey was largely based on fresh material collected from the field,
from plants cultivated at the Bogor Botanical Garden and from herbarium material
supplied by the Herbarium Bogoriense, Bogor. A full list of names and sources is
provided in the appendix 1.
Leaves from living and dried specimens were used. Dried leaves were
boiled in water for a few minutes to soften the leaf until they became unfolded
and were ready for epidermal scrapping. Fresh leaves were fixed in 70% FAA.
Leaf samples were prepared according to the modified method of Johansen
(1940). The fresh or dried leaves were placed in a tube with 10% nitric acid and
kept in boiling water for about 10-15 minutes. Nitric acid softens the leaf tissue
and facilitates peeling of the epidermis.
Both epidermal layers were stripped off gently by brushing away
unwanted tissue with the help of a pointed needle and forceps, after which the
epidermis was stained with safranin. An excess of safranin was washed off and
the epidermis was temporarely mounted in an aquaous glycerol solution.
For sectioning, a rotary microtome was used to make 15-20 µm thick
sections. The ribbons were placed on clean slides smeared with a thin film of
Haupt’s albumen and allowed to dry, after which a drop of water was added prior to mounting. The slides were placed on a hot plate at 40oC for a few minutes to
let the ribbons expanding and they were stored overnight. In the next day the
slides were immersed for 2-5 minutes in a solution of xylene and absolute alcohol
(1:1 ratio v/v). The slides were then transferred to another solution of xylene and
alcohol in the ratio 1:3 (v/v) for few minutes, after which they were washed in a
series of 95%, 90%, 70% and 50% alcohol. The slides were stained with a few
drops of fast green and counterstained with safranin for two minutes, then
dehydrated in a series of 50%, 70%, 80%, 90% xylene/alcohol solution and