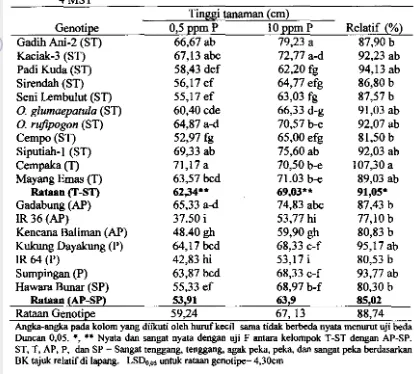
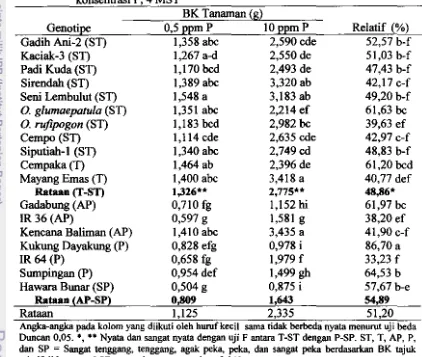
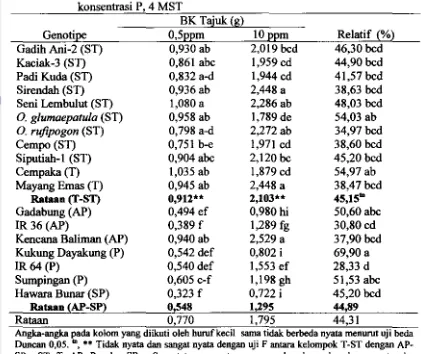
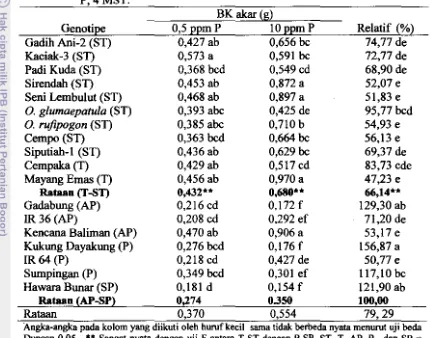
DI TANAH SAWAH
ABD. AZIZ SYARIF
SEKOLAII PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN
Saya menyatakan dengan sebenar-benamya bahwa semua pemyataan dalam disertasi saya yang berjudul Adaptasi daD Ketenggangan Genotipe Padi terhadap De .... siensi Fodor di Tanab Sawah merupakan gagasan dan basil penelitian disertasi saya sendiri, dengan bimbingan Komisi Pembimbing; kecuali
yang dengan jelas dinyatakan rujukannya. Disertasi ini belum pemah diajukan
untuk memperoleh gelar pada program sejenis di perguruan tinggi yang lain. Semua data dan infonnasi yang digunakan te1ah dinyatakan secara jelas dan diperiksa kebenarannya.
Bogor, Agustus 2005
Defisiensi Fosfor di Tanah Sawah. Dibimbing oleh DIDY SOPANDIE, M.A.
CHOZIN, TRlKOESOEMANINGTY AS, KOMARUDDIN IDRlS, dan
SUWARNO.
Defisiensi hara fesfor adalah salah satu kendala produksi padi sawah di
Indonesia yang meliputi luas sekitar 1,27 juta hektar. Penggunaan genotipe atan
varietas yang tenggang terhadap defisiensi hara ini merupakan salah satu
pendekatan yang dapa! digunakan dalam penanggulangan kendala tersebut.
Perakitan varietas tenggang dan berdayahasil tinggi memerlukan identifikasi tetua yang memiliki sifat tenggang. Pengetahuan tentang mekanisme ketenggangan atau karakter yang berhubungan dengan ketenggangan akan sangat membantu
dalam penapisan genotipe untuk mendapatkan tetua tenggang.
Suatu penelitian yang terdiri dari satu percobaan Japang (pada sawah defisien P) dan empat percobaan rumah kaca menggunakan medium larutan hara telah dilaksanakao untuk mengevaluasi keragaman ketenggangan genotipe padi
terhadap P rendah dan untuk mengidentifikasi genotipe-genotipe yang tenggang dan karakter yang berkaitan dengan ketenggangan, dan mempelajari mekanisme adaptasi ketenggangan padi terhadap defisiensi P. HasH penelitian menunjukkan
terdapat keragaman ketenggangan terhadap P rendah di antara plasmanutfah (120 genotipe) yang diuji. Sejumlah 14 genotipe sangat tenggang berdasarkan bobot kering tajuk relatif dan 28 genotipe berdasarkan jumlah anakan relatif berhasil diidentiftkasi. Genotipe tenggang terhadap P rendah di lapang menunjukkan kemampuan penyerapan dan penggunaan internal hara P pada kondisi P rendah yang Iebih tinggi dibanding genotipe peka.
Terlihat variasi mekanisme peningkatan serapan P pada kondisi P rendah di
antara genoti pe tenggang. Mekanisme tersebut pembentukan akar yang tinggi,
peningkatan eksudasi asam organik, dan affinitas karier akar terhadap P yang tinggi. Namun ada genotipe (Gadih Ani-2) yang memiliki ketiga mekaoisme tersebut. Ditemukan beberapa mekanisme adaptasi tanaman terhadap P rendah di larutan hara yakni peningkatan nisbah akar-tajuk, peningkatan panjang akar
terpanjang, nisbah efisiensi dan penggunaan P tanaman dan tajuk, serta nisbah
kadar dan kandungan P akar-tajuk. Namun, mekanisme ini tidak konsisten dengan
perbedaan ketenggangan genotipe terhadap P rendah di lapang. Terdapat
ABSTRACT
ABD. AZIZ SYARIF. Adaptation and Tolerance of Rice Genotypes to Phosphorus Deficiency in Paddy soil. Supervised by DIDY SOPANDIE, M. A. CHOZIN, TRIKOESOEMANINGTY AS, KOMARUDDIN IDRIS, and SUWARNO.
Phosphorus deficiency is one of the constraints in rice production in Indonesia, affecting about 1.27 million hectares of lowland (paddy) rice field. The use of tolerant genotype to low phosphorus (deficient) soil has been believed to be a sound approach to ease the problem. However, tailoring tolerant and high yielding varieties requires identification of tolerant genotypes used as parent material. Infonnation on the mechanisms as well as the characters related to the tolerance will be advantageous for accurate parental selection.
A research consisting of five experiments has been perfonned to evaluate the variability of low P tolerance among rice gennplasm and to identify to tolerant genotypes as well as the mechanisms and plant characters underlying the tolerance. The results showed that there was phenotypic and genotypic variability of low P tolerance among gennplasm evaluated. Fourteen genotypes have been identified as highly tolerant based on relative shoot dryweight and 28 genotypes based on relative tillering ability. Tolerant genotypes showed higher phosphorus uptake and internal use efficiency than those of sensitive genotypes, indicating both external and internal efficiency.
DITANAHSAWAH
ABD. AZIZ SY ARIF
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Departemen Agronomi dan Hortikultura
SEKOLAHPASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Disertasi: Adaptasi dan Ketenggangan Genotipe Padi terhadap Defisiensi Fosfor di Tanah Sawah
Nama
NIM
: Abd. Aziz Syarif : P03600005
Disetujui, Komisi Pembimbing
Prof. Dr. Jr. Didy Sopandie. M.Agr.
Prof. Dr. Ir. .A. Chozin M.A
Dr. Ir. Komaruddin Idris. M.S.
Anggota
Ketua Program Studi Agronomi
Aセ@
U
Dr. Jr. Satnyas I1yas, M.S.Tanggal Ujian: 26 Juli 2005
Ketua
.----Dr. Ir. T rikoesoemaningtyas, M.Sc.
Anggota
Dr. Jr. Suwamo. M.S.
Anggota
セIi。「@ PascasaJjana
セセi。ヲゥゥ、。@ Manuwoto, M.Sc.
U
Syukur Alhamdulillah penulis panjatkan ke hadirat Allah SWT yang telah
melimpahkan karuniaNya sehingga penulis dapat menyelesaikan penelitian dan penulisan karya ilmiah yang menjadi tugas Studi Doktor pada Program Studi Agronomi Institut Pertanian Bogor.
Ucapan terima kasih sebesar-besarnya penulis sampaikan kepada Prof. Dr. Jr. Didy Sopandie, M.Agr., Prof. Dr. Ir. M.A. Chozin, M.Agr., Dr. Ir. Trikoesoemaningtyas, M.Sc., Dr. Ir. Komaruddin Idris, M.S., dan Dr. Ir. Suwarno, M.S. yang telah memberi saran, baik pada persiapan dan pelaksanaan penelitian, maupun penulisan hasilnya. Terima kasih yang sarna juga disampaikan pada Kelompok Surnberdaya Genetik pada Balai Besar Penelitian Bioteknologi dan Sumberdaya Genetik Pertanian, dan Instalasi Penelitian BALITP A di Muara yang telah menyediakan plasmanutfah padi yang digunakan
pada peneiitian ini. Kepada Pusat Penelitian dan Pengembangan Tanah dan
Agroklimat yang memberi fasilitas rumah kaca dan laboratorium juga disampaikan terima kasih. Kepada Badan Penelitian dan Pengembangan Pertanian yang telah rnemberi kesempatan dan dukungan dana bagi penulis dalam melaksanakan tugas belajar di IPB penulis sampaikan penghargaan. Dukungan keluarga dan rekan-rekan seperjuangan juga sangat membantu, untuk itu penulis ucapkan terima kasih setulus-tulusnya.
Akhimya, penulis berharap karya i1miah ini dapat melengkapi infonnasi ilmiah yang sudah ada dan dapat digunakan untuk kemajuan ilmu dan teknologi, khususnya di bidang pertanian.
Bogor, Agustus 2005
RIWAYATHIDUP
Penulis dilahirkan di Kanagarian Sumpur, Kabupaten Tanah Datar
(Swnatera Barat) pada tanggal 4 Maret 1959 sebagai anak ketujuh dari Ibunda Khadijab Hamid (almarhwnab) dan Bapak Syarif Ali (almarhwn). Pendidikan
dasar diperoleh di Kanagarian Swnpur, sedangkan SMP dan SMA di Padang.
Pendidikan sarjana di tempuh pada Fakultas Pertanian Universitas Andalas. lulus
pada tabun 1983. Pada tabun 1984 penulis diterima di Jurusan Agronomi
Fakultas PascasaJjana IPB sebagai petugas belajar dari Badan Litbang Pertanian
dengan beasiswa NARP 11, dan lulus pada tabun 1987. Selanjutnya penulis
mendapat kesempatan untuk mengikuti studi program doktor pada Program Studi Agronomi IPB pada tabun 2000 dengan beasiswa P AA TP dari Badan Litbang
Pertanian.
Penulis beketja di BPTP (Balai Pengkajian Teknologi Pertanian) Swnatera
Barat yang berkedudukan di Sukarami, Kabupaten Solok, pada kelompok peneliti
DAFTAR TABEL ... Ix
DAFTAR GAMBAR ... .... xii
DAFT AR LAMPIRAN ... XIII PENDAHULUAN Latar Belakang ... I Tujuan Penelitian ... 3
Hipotesis ... 3
Manfaat Penelitian ... 3
Kerangka Penelitian ... 3
TINJAUAN PUSTAKA ... 5
EV ALUASI KETENGGANGAN GENOTIPE PAD! TERHADAP P RENDAH D! TANAH SAWAH Abstrak ... ... ... ... 18
Abstract... .... ... ... ... ... ... 18
Pendahuluan ... 19
Bahan dan Metode ... 19
Hasil dan Pembahasan ... 21
Simpulan ... ... 32
Daftar Pustaks ... 33
TANGGAP GENOTIPE TANAMAN PADI TERHADAP DOSIS P D! LARUT AN HARA Abstrak ... '" ... ... .... ... ... ... ... ... 36
Abstract ... ... ... ... ... ... 36
Pendahu1uan ... 37
Bahan dan Metode ... 37
Hasil dan Pembahasan ... ... 39
Simpulan ... 61
Daftar Pustaks ... 62
PERTUMBUHAN DAN SERAPAN P GENOTIPE TANAMAN PADI PADA DUA BENTUK SUMBER P Abstrak ... 64
Abstract... ... ... ... ... ... 64
Pendahu1uan ... .... ... .... ... ... 65
Bahan dan Metode ... 65
Hasil dan Pembahasan ... 66
Simpulan ... 87
Daftar Pustaks ... 87
PENGARUH STATUS P TANAMAN PADA BEBERAPA GENOTIPE PAD! TERHADAP EKSUDASI ASAM ORGANIK Abstrak ... 89
Abstract ... _... 89
Halaman
Pendahuluan... ... 90
Bahan dan Metode... ... ... ... ... .... 90
Hasil dan Pembahasan... ... ... 91
Simpulan... ... ... 97
Daftar Pustaka... ... ... .... 97
PENGUJIAN K1NETIKA PENYERAPAN P PADA BEBERAPA GENOTIPE TANAMAN PADI Abstrak... 99
Abstract... ... .... ... ... 99
Pendahuluan... 100
Bahan dan Metode... 100
Hasil dan Pembahasan... lOl Simpulan... ... ... ... 103
Daftar Pustaka... ... ... 103
PEMBAHASAN UMUM... .... ... 105
SIMPULAN DAN SARAN... llO
DAFTAR PUSTAKA... III
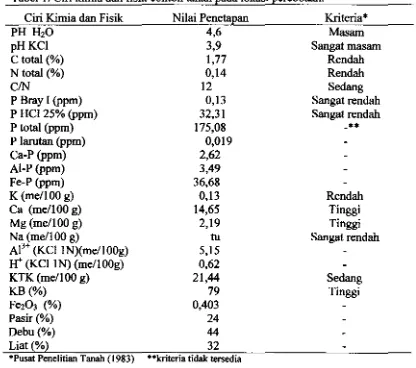
HalamaD I Ciri kimia dan fisik contoh tanah pada lokasi percobaan... 22
2 Kuadrat tengab pengaruh dosis P terhadap pertumbuhan 120
genotipe padi di tanah sawab... 24
3 Ragam dan koefisien ragam fenotipik dan genotipik ketenggangan
120 genotipe padi terhadap P rendab di tanah sawah... 25
4 Bobot kering tajuk genotipe padi sangat tenggang pada dua dosis
pemupukan P di tanah sawab... ... 26
5 Jumlab anakan genotipe padi sangat tenggang berdasarkan jumlab
anakan relatif. ... 27
6 Bobot kering tajuk, serta kadar, serapan, nisbah efisiensi, dan efisiensi
penggnnaan P genotipe peka dan tenggang di tanah sawah... 32
7 Bobot kering tajuk, jumlab anakan, dan tinggi tanaman padi yang
digunakan pada percohaan pengaruh dosis P di larutan hara... 38
8 Kuadrat tengah pengaruh dosis P terhadap pertumbuhan 18 genotipe
padi di larutan hara... 40
9 Tinggi tanaman 18 genotipe padi di larutan hara pada dua dosis P... ... 41
10 Jumlab anakan 18 genotipe padi eli larutan hara pada dua dosis P... 42
II Bobot kering tanarnan 18 genotipe padi di larutan hara pada dua dosis
P ... __ .... 43
12 Bobot kering tajuk 18 genotipe padi eli larutan ham pada dua dosis P.... 45
13 Sobot kering akar 18 genotipe padi di larutan hara pada dua dosis P... 46
14 Nisbab hobot kering akar-tajuk 18 genotipe padi di larutan ham pada
dua dosis P ... ... ... 48
15 Panjang akar terpanjang 18 genotipe padi eli larutan hara pada dua
dosis P... 59
16 Kuadrat tengah pengaruh dosis P terhadap serapan dan efisiensi
penggnnaan P di larutan hara... 50
17 Kadar P tanaman 18 genotipe padi di larutan hara pada dua dosis P... 51
Halaman 18 Serapan P per tanaman 18 genotipe padi di larutan hara pada dua dosis
P... 53
19 Penurunan pH medium 18 genotipe padi di larutan bata pada dua dosis
P...
5520 Efisiensi serapan P akar 18 genotipe padi di larutan hara pada dua dosis
P...
5721 Nisbah efisiensi P 18 genotipe padi di larutan hara pada dua dosis P... 59
22 Efisiensi penggunaan P 18 genotipe padi di larutan bata pada dua dosis
P... 60
23 Bobot kering tanaman 10 genotipe padi pada dua sumber P di larutan
hara...
6724 Serapan P 10 genotipe padi pada dua sumber P di larutan bata... ... 68
25 Penurunan pH medium 10 genotipe padi pada dua sumber P di larutan
hara... 69
26 Penunman nisbah kadar amonium-nitrat medium 1 0 genotipe padi pada dua sumber P di larutan bata... ... ... 70
27 Bobot kering akar 10 genotipe padi pada dua sumber P di larutan
bara...
7128 Bobol kering tajuk 10 genotipe padi pada dua sumber P di larutan
bara...
7229 Nisbah akar tajuk 10 genotipe padi pada dua sumber P di larutan
hara...
7330 Kadar P tajuk 10 genotipe padi pada dua sumber P di larutan bata.. ... 74
31 Kadar P akar 10 genotipe padi pada dua sumber P di larutan bata.. ... 75
32 Nisbah kadar P akar-tajuk 10 genotipe padi pada dua sumber P di
larutan
bara...
7633 Serapan P akar 10 genotipe padi pada dua snmber P di larutan bata... 77
34 Serapan P tajuk 10 genotipe padi pada dua sumber P di 1arutan bata. .... 78
35 Nisbah P akar-tajuk 10 genotipe padi pada dua sumber P di larutan
bata... 79
hara... 81
37 Efisiensi serapan akar 10 genotipe padi pada dua sumber P di larutan
hara... 82
38 Nisbah efisiensi P akar 10 genotipe padi pada dua sumber P di larutan
hara... 83
39 Efisiensi penggunaan P akar 10 genotipe padi pada dua sumber P di
larotan hara... 84
40 Nisbah efisiensi P tajuk 10 genotipe padi pada dua sumber P di larutan
hara... 85
41 Efisiensi penggunan P tajuk 10 genotipe padi pada dua sumber P di
larutao
hara... ...
8642 Eksudasi asam orgaoik per taoamao empat genotipe padi... 93
43 Eksudasi asam orgaoik per g tajuk taoamao empat genotipe padi... 94
44 Kadar P tajuk dan akar empat genotipe padi dengan dan tanpa
pelaparan P... 95
45 Bobot kering akar empat genotipe padi dengan dan taopa pelaparan P... 96
46 Parameter kinetika serapan P empat genotipe padi... 102
DAFTAR GAMBAR
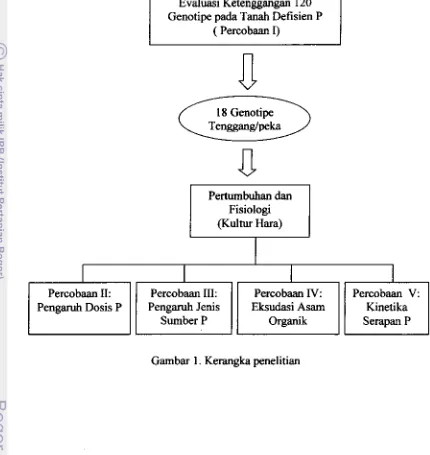
Halaman Kerangka Penelitian ... _... 4
2 Kurva erapan P contoh tanah lokasi percobaan... 23 3 Kurva semi log erapan P contoh tanah lokasi percobaan... 23
4 Pengaruh pemberian P terhadap pertumbuhan tanaman padi di tanah
sawab... ... 24
5 Sebaran 120 genotipe padi menurut tingkat ketenggangan terhadap
defisiensi P pada tanah sawab. ... ... 25
6 Korelasi bobot kering tajuk relatif denganjumlab anakan relatif 120
genotipe padi... ... ... 28
7 Korelasi bobot kering relatif 120 genotipe denganjumlab anakan dan
tinggi tanaman pada kondisi tanpa pupuk P... 29
8 Korelasi tingkat ketenggangan genotipe padi terhaciap P rendah dengan
bobot kering tajuk pada kondisi tanpa dan dengan pupuk P... 30
9 Respon genotipe tanaman padi dengan empat tingkat ketenggangan
terhadap pemupukan P ... 31
10 Korelasi jumlah anakan relatif di larutan hara dengan nilai relatif di
lapang... 42
11 Korelasi tingkat ketenggangan genotipe padi di lapangan dengan
serapan P tanaman dilarutan hara pada 0,5 ppm P... 54
12 Korelasi bobot kering akar dengan dengan serapan P tanaman padi
pada 0,5 ppm P di larutan hara... .... ... 54
13 Korelasi bobot kering akar per tanaman dengan penurunan pH medium 56
14 Korelasi efisiensi serapan akar relatif tanaman padi dengan di larutan
hara
dengan ketenggangan terhadap P rendab di lapang.. ... 5815 Korelasi efisiensi penggunaan P di larutan
hara
pada IO ppm P dengan ketenggangan di lapang... ... 6116 Korelasi hobot kering akar relatif dengan kandungan P akar relatif
tanaman padi di larutan
hara...
7717 Korelasi nisbab BK akar-tajuk relatif dengan nisbab P akar-tajuk relatif 80
kering akar ... .
19 Laju serapan P empat genotipe tanarnan padi ... 10 I
DAFTAR LAMPIRAN
Halaman
1 Genotipe padi pada percobaan evaluasi ketenggangan di lapang.. ... 119
2 Komposisi dan konsentrasi larutan hara yoshida... ... 122 3 Pengaruh pemberian P terhadap tinggi tanaman 120 genotipe padi di
tanah sawah... ... 123
4 Pengaruh pemberian P terhadap jumlah anakan 120 genotipe padi di
tanah sawah... 126
5 Pengaruh pemberian P terhadap bobot kering tajuk 120 genotipe padi
di tanah sawah... 129
Latar Belakang
Usaha pemenuhan kebutuhan beras merupakan program yang berkelanjutan
untuk memenuhi kebutuban yang terus meningkat disehabkan oleh pertumbuban
jumlah penduduk (Hasanuddin ef al. 2000). Dari berbagai agroekosistem pertanaman padi, padi sawah irigasi merupakan andalan utama untuk pencukupan
produksi. Hal ioi adalah karena luas dan produktivitasnya yang lebih tinggi
dibanding pada agroekosistem lainnya.
Defisiensi ham fosfor (P) adalah salah satu kendala dalam sistem produksi
tanaman padi. Defisiensi ini terdapat secara luas pada hampir semua ekosistem
pertanaman padi (Dobermann dan Fairhust 2000). Tipe-tipe tanab dengan kecenderungan terdapat defisiensi P adalah; tanab bertekstur kasar dengan kandungan bahan organik rendah, tanah dengan pelapukan Ian jut Ultisols,
Oxisols, tanab sawah terdegradasi, tanab berkapur, tanab salin, tanab sodik, tanah
vulkanis dengan sorpsi P tinggi (Andisols), serta tanah gambut dan sulfat masam dengan kandungan Al dan Fe aktif tinggi. Defisiensi dapat juga teIjadi karena pemupukan yang tidak seimbang atau penanarnan varietas yang peka. Vertisols dan Inceptisols juga berpotensi menimbulkan defisiensi P pada tanaman padi (De Datta 1981).
Di Indonesia telah dipetakan status P sekitar 7,5 juta ha sawah, dari jumlah
uu terdapat 17% (1,27 juta ha) berstatus P rendah, 43% (3,24 juta ha) berstatus
sedang, dan 40% (2,99 juta hal berstatus tinggi (Sotyan dan Adimibardja 2001).
Akibat kenaikan harga pupuk menyusul penghapusan suhsidi harganya, diperkirakan luas sawah berstatus P rendah akan semakin bertambah akibat penurunan penggunaan pupuk.
Pemberian pupuk fosfat adalah salah satu usaha untuk mengatasi defisiensi
P pada tanaman padi. Namun, beberapa basil penelitian menunjukkan bahwa efisiensi pemupukan fosfat pada tanaman padi sangat rendah, hanya 15-20% dari P yang diberikan dapat diserap tanaman (Baharsyah 1990). Menurut De Datta (1981) efisiensi pemupukan P pada tanaman padi umurnnya kurang dari 10%.
Rendahnya efisiensi pemupukan P ini adalah karena terjadinya transformasi
2
kalsium, magnesium, dan mangan menjadi bentuk yang sukar lamt dalam air (Tisdale et al. 1985).
Penggunaan genotipe yang tenggang dengan kemampuan yang tinggi dalam menyerap dan menggunakan P secara efisien adalah salah satu pendekatan yang dianjurkan untuk menjamin keberlanjutan (sustainability) sistem produksi (Duncan dan Baligar 1990; Subbarao et al. 1997). Genotipe yang efisien akan mengurangi kebutuban pemberian pupuk P, memungkiukan penggunaan pupuk yang kurang larnt namun lebih murah seperti fosfat alam, dan memperpanjang efek residu pemupukan (Polle dan Konzak 1990). Sifat tenggang terbadap defisiensi P dapat dimanfaatkan untuk meningkatkan daya hasil tanaman pada tanab defisien P. Wissua dan Ae (200 I) berbasil meningkatkan daya basil padi gogo Nipponbare yang kurang tenggang P sebesar 250% dengan memanfaatkan sifat tenggang defisiensi P pada varielas lokal (Ianras) Kasalath.
Identifikasi plasmanutfah merupakan tahap awal dari program untuk merakit varietas yang tenggang terhadap kondisi kahat hara. Identifikasi ini biasanya juga meliputi infonnasi mekanisme morfologis, fisiologis,
dan
biokimia yang potensial terlibat dalam sifat efisien tersebut ( Gourley et af. 1993; Crowley dan Rengel 2000). Informasi tentang mekanisme ini dapat digunakan dalam mengembangkan teknik penapisan genotipe (plasmanutfah) yang akurat (Rengel 2000). Penelitian-penelitian tentang variasi genotipik dan mekanisme adaptasi tanaman terhadapstres hara P telah banyak dilaporkan (Blum 1994; Rao et al 1999; Caradus 1990).
Namun. penelitian-penelitian tersebut umumnya dilakukan pada tanaman selain padi seperti terigu, barley, jagung, kede1ai. dan tanaman pakan ternak. Penelitian mekanisme adaptasi tanaman padi terhadap defisiensi P, khususnya pada plasmanutfah padi Indonesia masih sangat terbatas. Di samping itu, kebanyakan penelitian tersebut bersifat parsial, hanya mene1iti satu atau dua aspek adaptasi seperti efisiensi serapan P, eksudasi asam organik, kinetika serapan P dan lain-lain. Publikasi yang tersedia juga lebih banyak melaporkan basil penelitian lapang atau rumah kaca yang menggunakan larutan hara. Laporan penelitian yang
Tujuan Penelitian
Penelitian ini bertujuan untuk (1) mengidentifikasi variasi ketenggangan plasmanutfah padi (terutama varielas lokal) pada P rendah di tanah sawah, (2)
mengidentifikasi mekanisme adaptasi dan ketenggangan genotipe padi terhadap
defisiensi P di tanah sawah, (3) mengidentifikasi karakter-karakter tanaman yang berkaitan dengan ketenggangan terhadap P rendah di tanah sawah.
Hipotesis
Hipotesis yang dikemukakan pada penelitian ini adalah (1) terdapat variasi
ketenggangan terhadap P rendah di tanah sawah di antara plasma nutfah padi, (2) perbedaan ketenggangan terhadap P rendah di tanah sawah di antara plasmanutfah padi dapat diterangkan dengan perbedaan fisiologis seperti efisiensi penggunaan P
internal, perubahan pH medium, eksudasi asam organik, dan afinitas akar terhadap
ion P, serta pertumbuhan (morfologis) tananum seperti pertumbuban akar.
Manfaat Penelitian
Hasil peneiitian ini diharapkan memberi manfaat (1) sebagai masukan
dalam program perbaikan varietas padi untuk ketenggangan terhadap P rendah
pada sawah melalui identifikasi plasmanutfah yang tengang, (2) memperkaya infonnasi ilmiah tentang mekanisme adaptasi dan ketenggangan genotipe padi
terhadap P rendah di tanah sawah.
Kerangka Penelitiao
4
dilaksanakan di Laboratorium Kimia dan Uji Taoah Balai Penelitiao Taoah
Bogor, sedangkan analisis asam organik eksudat akar dilakukan di Laboratorium Balai Besar Penelitian Pasca Panen Bogor.
I
Percobaan II:
Pengaruh Dosis P
Evaluasi Ketenggaogao 120 Genotipe pada Tanah Defisien P
( Percobaao I)
I
D
18 Genotipe Tenggang/peka
Pertumbnbao dan
Fisiologi
(Kultur Hara)
I
Percobaan III: Percobaao IV:
Pengaruh Jenis Eksudasi Asam
SumberP Organik
Gambar 1. Kerangka penelitian
I
Percobaan V: Kinetika
[image:20.593.89.519.150.605.2]Fungsi dan Gejala Defisiensi Fodor pada Tanaman
Fungsi Fosfor
Fosfor tennasuk hara makro dibutuhkan tanaman dalam jumlah yang
banyak seperti halnya N, K, Co, Mg, dan S. Dalam tanaman P dijumpai dengan
kadar 0,1 - 0,4%, lebih rendah dari kadar N dan K (Tisdale et 0/. 1985). Secara
garis besar, fungsi P pada tanaman dapat digolongkan dalam tiga bagian
(Marschner 1995). Fnngsi pertama adalab sebagai penynsnn makromolekul. Dua
contoh utama atau terpenting dari makromolekul yang melibatkan P adalah asam nukleat (DNA, RNA) dan fosfolipid biomembran. Asam nukleat adalab senyawa
yang berperan dalam pewarisan sifat dan perkembangan tanaman. Pada
biomembran P membentuk ikatan atau jembatan antara digliserida dan molekul
lainnya seperti asam amino, amina, atau alkohol, membentuk fosfatidilkolin (lesitin) yang menjaga intergritas membran. Fnngsi kedua dari P adalab sebagai
unsur pembentuk senyawa penyimpan dan perpindahan energi. Dua senyawa
kaya energi yang paling umum adalab ATP dan ADP. Energi dalam ATP/ADP terletak pada ikatan pirofosfat yang pemecahannya akan me1epaskan energi, yang
dikenal dengan proses fosforilasi. A TP merupakan sumber energi untuk hampir
semua proses biologi yang membutuhkan energi. Unsur P juga diperlukan dalam
proses fotosintesis yakni pada fotofosporilasi dan pembentukan ribulosa
1.5-bifosfat. Fungsi ketiga P adalah sebagai regulator reaksi biokimia melalui fosforilasi yang dapat mengaktivasi atau inaktivasi protein yang dianggap sebagai faktor kunci dalam transduksi sinyal.
Secara agronomis unsur P diketahui berperan dalam percepatan pematangan biji, kekuatan batang sereal, serta mutu buah, hijauan, dan biji-bijian.
Ketenggangan tanaman terhadap penyakit juga meningkat pada tanaman yang
mendapat cukup P (Tisdale et 0/. 1985). Benih yang dihasilkan dari tanaman
yang mendapat cukup P akan memiliki daya kecambah dan vigor yang tinggi
6
Pada tanaman padi P dipedukan dalam perkembangan akar, mempercepat
pemblUlgaan dan pematangan (terutama pada suhu rendah), serta mendorong
pembentukan anakan dan biji (De Datta 1981).
Gejala Defisiensi Fosfor
Gejala khas defisiensi sering sukar terlihat. tidak seperti gejala defisiensi unsur lainnya seperti K dan Mg. Kekerdilan dan pengurangan jumlab anakan
pada tanaman monokotil atau cabang pada dikotil, daun pendek dan tegak, serta
penundaan pembungaan adalab gejola yang umum pada kebanyakan tanaman
(Rao dan Terry 1989). Penurunan luas dan jumlab daun juga merupakan gejala defisiensi P akihat tertekannya perkembangan sel epidennis daun (Lynch et al. 1991). Tanarnan yang defisien P juga sering memperlihatkan daun sempit bewama bijau gelap (Rao dan Terry 1989). Hal ini adalab karena pertambaban luas daun lebili tertekan dibandingkan pembentukan kloroplas dan klorofil.
Pada tanaman padi defisiensi P mengakibatkan pertumbuban tanarnan yang
kerdil dengan pengurangan jumlab anakan, daun sempit, pendek, kaku dan
bewama bijau kotor. Jumlab daun, malai, dan biji per molai juga berkurang.
Daun muda kelibatan sebat, tetapi daun tua berwarna kecoklatan dan mati. Pada
tanaman yang dapat membentuk antosianin, dapat muncul gejala daun berwama merah atau ungu. Defisiensi P pada tingkat sedang sukar diamati di lapang (Dobennann dan Fairhust 2000).
Penyerapan Fosfor
Tanaman menyerap P dari larutao tanah terutama dalam bentuk ortofosfat primer dan sekunder (H,PO, dan HPol') dan sedikit dalam bentuk senyawa
organik (Tisdale et al 1985). Orthofosfat sekunder lebili dominan pada pH di
alas 7,22, namun tanaman menyerap P ini lebili lambat dibandiug orthophosfat primer. Bagian tanarnan yaug aktif menyerap P adalab jaringan mnda dekat ujung
akar. Konsentrasi P yang relatif tinggi menumpuk di ujung akar diikuti oleh
akumulasi yang rendab pada bagian pemanjangan, kemudian oleh akumulasi
Penyempan P oleh tanaman dari tanah adalah penyempan aktif karena melawan gradien konsentrasi (Clarkson dan Grignon 1991). Kadar P larutan
tanah di luar sel akar umumnya hanya 111M atau kumng, sedangkan kadar dalarn sitoplasma adalah 103 sarnpai 10' lebih tinggi. Kedua larutan dengan perbedaan
konsentrasi yang besar ini dipisahkan oleh membran plasma dengan ketebalan hanya sekitar 8 nm. Untuk membawa 1 mol P ke dalam akar sel dibutuhkan
energi minimal 25-40 kJ, setara dengan energi bebas yang dilepas dari hidrolisis I mol A TP. Energi dari hidrolisis A TP digunakan lUltuk memompa proton keluar membran plasma, menciptakan graciien pH antara sitoplasma dan apoplas (dinding
sel). Ion fosfat (anion) akan masuk ke dalam sitopiasma bersama proton
(symport) ataupun OHlanion antiport yang difasilitasi oleh protein khusus
(transporter). Beberapa gen yang menyandi transporter P dan terutama terekspresi di akar telah berhasil diisolasi (Smith 2000). Ekspresi gen tersebut dipengaruhi oleh status P tanaman, kekumngan P meningkatkan tmnskripsi tmnsporter di akar.
Berbeda dengan nitrogen, fosfor yang disemp tanaman tidak mengalarni reduksi, tetapi tetap dalarn bentuk oksidatif tertinggi (Marschner 1995). Setelah diserap. fosfat dapat tetap sebagai P inorganik atau teresteriftkasi (melalui gugus hidroksil) dengan mntai karbon (C-O-P) sebagai ester P sederhana (gnla P) atau
terikat dengan P lainnya dengan ikatan pirofosfat kaya energi (ATP, ADP) atau
diester (C-P-C).
Pergerakan ion fosfat menuju akar tanaman terdiri dari dua cara yakni aliran massa dan difusi (Tisdale et al. 1985). Alimn massa adalah pergemkan ion
mengikuti pergerakan air menuju akar yang teIjadi sebagai akibat transpirasi.
Berdasarkan perhitungan besaran transpirasi dan konsentrasi ion P dalam tanah, sumbangan aliran massa dalam penyediaan P untuk tanaman dianggap kurang
bemrti. Dengan asumsi kadar ion P tanah (tanpa pemupukan) 0,05 ppm, alimn massa hanya menyumbang 1 % kebutuhan P tanarnan. Pada tanah yang dipupuk
sumbangan alimn massa dapat lebib tinggi karena pemupukan meningkatkan
konsentrasi ion P. Kadar P larut pada zona reaksi pupuk-tanarnan dapat mencapai 2 sarnpai 14 ppm. Narnun, keadaan ini hanya berlangsung sementam karena
8
Pergerakan secara difusi merupakan mekanisme pergerakan P menuju akar yang paling penting. kecuali pada tanah dengan kadar P sangat tinggi. Difusi P sangat dipengaruhi oleh beberapa faktor tanah yakni kadar air tanah, kapasitas penyangga P tanah, temperatur, dan bentuk lintasan difusi.
Bentuk dan Keseimbangan Fodor dalam Tanah
Secara garis besar fosfor (P) tanah digolongkan ke dalam P organik dan P anorganik, tergantlmg pada bentuk senyawanya (Tisdale el al. 1985). Fraksi organik ditemukan dalam humus dan senyawa organik yang berkaitan atau tidak
berkaitan dengan humus. Tanaman dapat menyerap P organik larot tertentu
seperti asam nukleat dan fitin. Namun, peranan P organik lebih sebagai cadangan P yang ketersediaannya bagi tanaman tergantung pada proses mineralisasi.
P anorganik terdapat dalam dua bentuk yakni P larutan dan P fase padatan yang dikatagorikan dalam tiga bentuk (Wijaya -Adi dan Sudjadi 1987). Bentuk
pertama adalah P lahil yakni bentuk yang cepat mengadakan keseimbangan dengan P larutan. Bentuk kedua adalah P metastabil, yang sedang-Iambat
mengadakan keseimbangan dengan P larutan. Bentuk ketiga yang rnerupakan P stabil adalah bentuk yang tidak membentuk keseimbangan dengan P larutan.
Reaksi keseimbangan diantara ketiga bentuk tersebut secara sederhana dapat digambarkan sebagai berikut:
I 2 3
P-Iarutan セ@ • P-Iabil 4 • P-metastabil 4 • P-stabil
Reaksi I berjalan cepat, reaksi 2 lamba!, dan reaksi 3 amat lambat. Reaksi I
adalah proses erapan, karena itu P lahil disebut P tererap, sedangkan reaksi 2 dan
3 adalah proses fiksasi yang berlangsung lambat. P metastabil dan stabil disebut P
non labil.
Ketersediaan Fosfor pada Tanab Tropis
rendah (mengandWlg <10% mineral dapat lapuk dalam fraksi pasir dan debu), dan
23% (1,1 milyar ha) memiliki kapasitas ftksasi tinggi. Fosfor merupakan ham
pembatas utama pada kebanyakan tanab-tanab ini (Uexkull dan Mutert 1995).
Tanab-tanab dengan keterbatasan P di daerab tropis umwnnya (43%)
tergolong pada ordo Oxisols dan Ultisols (Sanchez dan Uehara 1980).
Tanab-tanah ordo lainnya, terutama subgrup rodik dan oksik dari Alfisols dan Inceptisols juga memiliki keterbatasan P karena komposisi minera1nya yang tennasuk
pengerap P tioggi. Oxisols dan Ultisols memilki kapasitas fiksasi P sedang sampai tinggi karena adanya permukaan yang luas Wltuk sorbsi P akihat tingginya
kandungan amorfus dan mikrokristalin aluminium dan besi oksihidroksida.
Dari berbagai jenis tanah yang terdapat di Indonesia, Oxisols
dan
Ultisolsmerupakan jenis tanah yang dominan, yakni sekitar 36,4% dari luas tanah
(Muljadi dan SoepraptohaIjo 1975). Pemanfaatan tanab-tanab ini Wltuk budidaya
padi sawah menghadapi kendala kekurangan P, sehingga memerlukan pemupukan
P yang cukup tinggi (Roechan dan Sudarman 1982).
Ketenediaan Fosfor pada Tanah Sawah
Pengenangan dapat mengakibatkan peningkatan ketersediaan P tanab
(Sanchez 1976). Peningkatan ini disebabkan oleh beberapa faktor; (a) reduksi ferifosfat yang sukar larut menjadi ferofosfat yang lebih mudah larut, (b) lepasnya
P larut pereduksi (reductant soluble/occluded), (c) hidrolisis senyaw. AI-P atau
Fe-P, (d) mineralisasi P organik akibat kenaikan pH, dan (e) peningkatan difusi
fosfat. Desorbsi P dari permukaan liat dan oksida aluminium dan besi, serta
pelepasan fosfat dari aluminium dan besi fosfat oleh anion organik juga
merupakan faktor peningkatan ketersediaan P akibat penggenangan (Patrick dan
Mabapatra 1968).
Peningkatan ketersediaan P umumnya teIjadi pada awal penggenangan kemudian menurun setelah dua sampai empat minggu akibat presipitasi oleh Fe2+ dan adsorpsi oleh partikelliat dan hidroksida AI (Dobennaon dan Fairhust 2000).
Peningkatan juga tidak terjadi pada semua jenis tanab. Pada tanab Ultisols tidak terlihat perbedaan P terekstrak Bray-2 antara yang tergenang dan tidak tergenang
10
padi sawah tidal< membutuhkan pupuk P. Defisiensi P terdapat secara luas pada
sawah denganjenis tanah Vertisols, Ultisols, dan Oxisols.
Adaptasi Tanaman terhadap Fosfor Rendah
Seleksi alarn untuk adaptasi terhadap masalah tanah adalah suatu proses
yang umum diamati. Hal ini terlihat dari adaDya berbagai ekotipe edaftk dalam
populasi vegetasi alami (Epstein dan Jefferies 1964). Tumbuhan yang adaptif
terhadap kondisi tanah kahat hara memiliki sifat yang lebih baik dalam menyerap bam, membuat ham lebih tersedia, atau mentranslokasikan hara pada limbung
(sink) yang sesuai (Blum 1988). Karena ito, dalarn konteks praktis dan bukti
fisiologis, ketenggangan terhadap defisiensi ham sebagian besar adalah
merupakan mekanisme penghindaran (avoidance), dimana tanaman yang
tenggang dapat menghindari terjadinya konsentrasi ham yang rendah pada jaringan. Sebalikoya, ketenggangan adalah kerusakan yang relatif kecil pada
proses fisiologis walaupun jaringan tumbuhan memiliki kandungan ham tertentu di bawah kadar kritis. Situasi ini jarang ditemui, dan dinyatakan sebagai efisiensi metabolis atau tenggang.
Seeara wnum adaptasi tanaman ini dicapai melalui mekanisme peningkatan
penyerapao dan peningkatan efisiensi penggunaan (Rao el al. 1999). Peningkatan
penyerapan dicapai melalui perbedaan morfologi (pertumbuhan dan distribusi, diameter, rambut akar)
dan
fisiologi akar (system penyerapan dan mobilisasi P pada rizosfir), sedangkan efisiensi pengunaan dicapai melalui partisi (mobilisasi) P dalam tanaman dan efisiensi penggunaan pada level selular.Peningkatan Penyerapan Fosfor
Peningkatan serapan hara per tanamao dapat dicapai dengao (a) peningkatan
sistem perakaran yang dapat meningkatkao kontak dengan suatu hara, terutama hara yang kurang mobil seperti P; (b) peningkatan serapao per unit akar dengan meningkatkao kinetika serapao; dan/atau ( c) kemampuan untuk menggunakan
bentuk-bentuk hara yang relatif tidal< atau kunmg tersedia bagi tanaman seperti P
Morfologi akar. Berdasarkan pemodelan yang dikembangkan oleh
Silberbush dan Barber (1983), peningkatan penyerapan P paling dipengarnhi oleh
peningkatan luas permukaan dibanding perubahan peubah akar lainnya. Genotipe
Phaseo/us vulgaris yang tumbuh baik pada kondisi P rendah memiliki perakaran
dua kali lipat dibanding perakaran genotipe kurang adaptif (Gabelman 1976).
Pada kacang gude (Cajanus cajanJ. Subbarao el al. (1997) melaporkan korelasi
positif antam hobot kering akar dengan serapan P tanaman. Pentingnya luas pennukaan akar dalam penyerapan P ditunjukkan oleh lebih tingginya efisiensi
penyerapan akar pada genotipe terigu dengan panjang akar spesifik (panjang akar
per unit berat akar) yang tinggi (Settlemacher el al. cil. Rao 19%). intersepsi tanab oleh akar meningkat dengan semakin halusnya akar. Sifat inilab yang
membuat rerumputan pereniallebih adaptif dibanding tanaman annual.
Rambut akar juga berperan dalam penyerapan P. Penyerapan per unit panjang akar meningkat dengan pembentukan rambut akar. Hal ini ada1ab karena rambut akar meningkatkan area pennukaan akar sehingga volume ekplorasi tanah per panjang akar meningkat (Junk dan Claassen cil. Rao 1999). Perakaran
tanaman memiliki sifat plastisitas terhadap ketersediaan P. Hackett cit. Blum
(1988) mengamati pembentukan akar yang ekstensif pada genotipe harley
tenggang sebagai akibat defisiensi P. Panjang dan kerapatan rambut akar
Brassica napus meningkat pada kondisi defisien P (Foshe dan Junk 1993).
Pada tanarnan padi pembentukan rambut akar sangat dipengaruhi oleh tingkungan pertumbuhan (Yoshida 1981). Kondisi aerobik seperti pada
pertanaman padi gogo mendorong pembentukan rambut akar, sedangkan kondisi an aerobik seperti padi sawah menekan pembentukan rambut akar.
Di samping modifIkasi peubah akar, perluasan permukaan perakaran dapat
juga dicapai dengan adanya asosiasi tanaman dengan mikoriza. Tanaman ubikayu dengan system perakaran yang kurang baik (percabangan kurang banyak, diameter besar) haradaptasi baik pada tanab rendab P karena adanya asosiasi dengan mikoriza (Howeler dan Sieverding 1987). Asosiasi tanaman sorghum dengan mikoriza juga terbukti dapat menittgkatkan pertumbuhan pada tanab
12
Fisiologi Akar. Perubahan fisiologi tanaman seperti peningkatan daya serap akar juga merupakan mekanisme ketenggangan tanaman terhadap terbatasnya suplai P, seperti diperlibatkan oleh tanaman jagung dan kedelai (Junk
ef ai, 1990), Pada kedua tanaman ini terlihat peningkatan daya serap akar maksimum akihat penurunan kadar P tanaman pada kondisi P rendah pada medium pertumbuhan. Peningkatan daya serap maximum (2-4 kali) pada kondisi
P rendah juga ditemui McPbarlin dan Bieleski (1987) pada tumbuhan Spirodela
dan Lemma, tanpa peningkatan afinitas transporter (penurunan Km). Nielsen dan
Schjoning (1983) mendapatkan variasi kinetik serapan P akar pada genotipe barley, dan kesesuaian serapan P tanaman di lapang dengan serapan berdasarkan
dugaan menggunakan peubah kinetik: serapan (I max, Km. Cmin). Berdasarkan hal ini dimungkinkan untuk meningkatkan efisiensi penyerapan P tanah dengan
memilih danlatau mengembangkan genotipe yang memiliki
K.n
danlatau erninrendah danlatau Imax tinggi.
Eksudasi asam organik (malat, sitrat, oksalat) adalah mekanisme lain tanaman untuk meningkatkan ketersediaan P dari tanah. Hoffland ef al. (1989) menemukan peningkatan eksudasi malat dan sitrat pada Brassica napus. Pada
Medicago sativa terlihat peningkatan eksudasi sitrat hampir dua kali lipat pada kondisi kurang P (Lipton ef al. 1987). Asam organik dapat meningkatkan ketersediaan P melalui mekanisme pelarutan senyawa P sukar larut (AI-P, Fe-P)
dengan penurunan pH atau desorbsi P dari tapak jerapan dengan pertukaran anion (Gerke cit. Crowley dan Rengel 2000). Anion dari asarn organik dapat
membentuk kompleks dengan Al atau Fe sehingga dapat melepaskan ion fosfat (Jones dan Darrah 1994) atau mencegah ion fosfat bereaksi dengan ion Al atau Fe
(Bar-Yosefl991).
Kemampuan mobiJisasi P oleh berbagai asam organik tidak sarna Asam sitrat diikuti oksalat memiliki kamampuan mobilisasi tinggi, malat dan tartarat
sedang, asetat dan suksinat rendah. Kemarnpuan ini berkaitan dengan stabilitas
kompleks anion asam organik dengan ion AI atau Fe dengan urutan
sitrat>malat>aspartat (Bar-Y osef 1991).
sitrat sintase mitokondria yang mengakibatkan peningkatan produksi sitmt yang
kemudian dieksresikan (Takita et al. 1999). Koyama et aJ. (2000) telah berhasil mengisolasi gen penyandi sitrat sintase mitokondria (CS) tanaman wortel dan
mengintroduksikannya pada Arabidopsis (ha/iana. Tumbuhan A. thaliana
transgenik memperlibatkao overekspresi CS pada tanah dengan ketersediaan P rendah dan pertumbuhan lebih baik dibanding tumbuhan asal (non transgenik).
Eksudasi asam organik (dan senyawa karbon lainnya) tidak banya berpengaruh langsung terbadap peningkatan ketersediaan barn bagi tanaman, tetapi memiliki pengaruh tidak langsung melalui pertumbuhan mikroorganisme
tanah karena senyawa-senyawa tersebut merupakan substrat bagi pertumbuhan mikroorganisme tanah. Menurut Lynch dan Whipps (1990) jumlah karbon yang dilepas tumbuhan ke rizosfir dapat mencapai 30-60% dari karbon yang diasimilasi tumbuhan. Peningkstan pertumbuhan mikroorganisme dapat mempengaruhi pH dan redoles tanah, serta mineralisasi bahan organik. s・ャ。ョェオエョケセ@ mikroorganisme dapat meningkstkao eksudasi akar melalui pelepasan zat pengatur tumbuh yang dapat meningkstkao pertumbuhan akar (Arshad dan Frankerberger cit. Crowley dan Rengel 2000).
Di samping peningkatan eksudasi asam organik yang clapat memobilisasi P anorganik, beberapa tumbuhan juga meningkatkan ekskresi enzim fosfatase (Ozawa et al. 1995) atau fitase (Li et aJ. 1997) yang dapat melarutkan P organik sebinga lebih tersedia untuk tumbuhan.
Peningkatan EflSiensi Penggunsan
Efisiensi Penggunaan. Definisi efisiensi penggunaan hara pada tanaman
sangat banyak dan bervariasi. Gerloff (1976) mendefinisikan efisiensi penggunaan
barn sebagai mg bobot kering tumbuhan yang dibasilkan per mg barn yang diserap
tanaman, dan disebut dengan nisbah efisiensi. Definisi ini memungkinkan
pernbandingan efisiensi penggunaan barn diantara strain dalam spesies dan
terutama diantara spesies. Definisi ini umum digunakan untuk menggambarkan
14
(konsentrasi) P, dan dipengaruhi oleb banyak faktor di luar tumbnhan, Barrow
(cit. Caradus 1990) mengemukakan bahwa definisi ini memiliki kelemahan jika digunakan dalam seleksi. Dengan definisi ini, genotipe dengan kadar hara rendah
dinyatakan sebagai pengguna efisien, namun boleh jadi sebenarnya kadar ham
rendah tersebut hanyalah karena ketidak efisienan dalam menyerap ham. Berdasarkan hal ini maka dikemukakan definisi efisiensi penggunaan hara sebagai
pertumbnhan per unit konsentrasi ham dalam tumbnhan. Siddiqi dan Glass (Glass 1990) juga mengemukakan babwa basil (bobot kering) per unit konsentrasi
hara dalam tanaman sebagai kriteria seleksi untuk efisiensi penggunaan ham. Indeks ini tidak hanya mengukur penggunaan ham internal, tetapi juga
memasukkan pengarnh perbedaan penyerapan.
Caradus (1990) mengelompokkan efisiensi hara, terutama untuk ham P, ke dalam riga katagori besar yakni; (a) memberikan basil yang lebili besar pada P
tanah yang rendab, (b) memberi basil lebib besar per jumlab P yang diserap
diistilahkan dengan efisiensi penggunaan atau nisbab penggunaan, dan (c)
memberi hasil lebih besar per unit P yang diberikan atau tersedia. Pengelompokan
yang berbeda untuk efisiensi P dikemukakan oleb Wilson (cit. Blair 1993) yakni;
(a) Efisiensi penyerapan, (b) efisiensi inkorporasi, dan (c) efisiensi penggunaan. Beberapa variable untuk menduga efisiensi berdasarkan pengelompokan Wilson ini disimpulkan oleb Blair 1993); (a) efisiensi penyerapan; serapan atau laju
serapan per unit panjang atau berat akar, Vrnax,
Km,
dan Cmin • (b) efisiensiiukorporasi; bobot kering tajuk per unit P dalam tajuk, nisbah penggunaan, dan
kadar P pada kondisi defisien, (c) efisiensi penggunaan; bobot kering tajuk per unit P yang diberikan, hobot kering tajuk per unit total serapan,. bobot kering
tajuk pada kadar P sarna, bobot kering tanaman per unit serapan, dan nilai kritis
konsentrasi P.
Mekanisme Peningkatan EflSiensi Penggunaan. Beberapa mekanisme
yang dapat meningkatkan efisiensi penggunaan P pada tumbnhan telab dilaporkan. Secara garis besar, efisiensi penggunaan P dapat meningkat melalui
mekanisme partisi P di dalam tumbuhan dan efisiensi penggunaan pada level
dalam tumbuhan dan kadar P pada organ yang dipanen. Efisiensi pada level
selular terdiri atas kompartementasi P intraselular dan kebutuhan metabolis P.
Remobilisasi P daJam tumbuhan merupakan salah satu mekanisme yang penting dalam meningkatkan efisiensi penggunaan P. Fosfor dari organ atau jaringan yang kurang atau tidak aktif dimobilisasi ke jaringan atau organ yang
aktif sehingga P yang telab diserap tumbuhan dapa! digunakan kembali dalam
proses fisiologi. Salah satu contoh remohilisasi P yang penting adalah mobilisasi
P dan daun yang menua (senescing) ke titik tumbuh (Chapin dan Kedrowski
1983). Kadar P rendah pada organ yang dipanen dapat dianggap meningkatkan
efisiensi (agronomis) penggunaan P karena mengurangi kebutuhan P untuk
menghasilkan satuan produksi ataupun mengurangi jumlah P yang dibawa ke luar sistem produksi. Penurunan laju kematian daun juga dapat meningkatkan efisiensi penggunaan P, karena dengan ini P dapa! digunakan lebih lama (Caradus
1990).
Translokasi ham yang diserap dari akar ke tajuk juga menentukan efisiensi
penggunaan P. Penyerapan ham yang tinggi belurn teotu memecahkan masalah
defisiensi p. tergantung pada apakah penyerapan yang tinggi tersebut disertai oleh translokasi ke tajuk. Pada Arabidopsis thaliana tipe liar misalnya ditemukan
bahwa penyerapan yang tinggi tidak disertai oleh translokasi yang tinggi. Hanya
35% dari P yang diserap ditranslokasikan ke tajuk, dibandingkan 90% pada tipe
mutannya Hasil penelitian menunjukkan bahwa penyerapan dan translokasi ke tajuk diatur oleh mekanisme terpisah yang dikendalikan secara genetik (Poirer et
al. 1991).
Kompartementasi P intraselular juga dapat mempengaruhi efisiensi
penggunaan P tumbuhan. Penelitian Mimura et al. (1996) menunjukkan babwa
terjadi pergerakan P dari vakuola ke sitoplasma pada kondisi defisien P. Adanya
pergerakan ini memungkinkan kadar P dalam sitoplasma dapat dipertahankan,
sehingga proses fisiologis tetap betjalan nonna1. Proporsi fraksi P anorganik
yang lebib rendah pada perlakuan P rendah dibanding perlakuan P tinggi pada tanaman padi (Swasti 2004) mengindikasikan adanya transfer P (anorganik) dan
16
Efisiensi metabolis adalah salah satu mekanisme adaptasi yang
dikemukakan oleh Gerloff dan Gabelman (Blum 1988). Indikasi adanya
metabolisme yang efisien adalah produksi bahan kering yang lebih tinggi pada
konsentrasi ham reDdall. Bukti adanya metabolisme yang efisien P dikemukakan oleh Murley et al. (1998). Beberapa tanaman yang dapat bertenggang pada
kondisi P rendah menunjukkan aktivitas PFP (Pyrophosphat-dependent
phospho-fruktokinase) yang tinggi. Enzim ini mengkatalisis reaksi yang memotong reaksi ATP-dependent fruktokinase (PFK). Modifikasi ini dapat mendaur ulang Pi dan
menghemat penggunaan ATP (Murley et al. 1998).
Ketenggangan dan Efisiensi. Ketenggangan terhadap stress ham rendah
adalah kemampuan tanaman untuk mempertahaukan hasil pada kondisi hara
terbatas (Caradus 1990). Sifat tenggang ini tidak dapat dipisahkan dengan
efisiensi seperti terlihat pada mekanisme yang mendasarinya (Rao et al. 1999).
Namun, ketenggangan lebih menggambarkan dinamika atau respon tanaman
terhadap perobahao lingkungan (polle dan Konzak 1990) dan mengambarkan kemampuan adaptasi (Wissua dan Ae 2001). Wissua dan Ae (2001) menggunakan nilai serapan yang tinggi pada kondisi minus P yang disertai nilai relatif terhadap kondisi plus P yang tinggi sebagai kriteria tanaroan padi yang
tenggang defisien harn P.
Keragaman dan Genetik Ketenggangan Padi terbadap Fosfor Rendah
Variasi antar varietas dalam penyerapan P, pertumbuhan, dan basil tanaman padi pada tanah delisien P te1ah lama dilaporkan (Yoshida 1981). Fageria et al.
(1988) mendapatkan variasi antar varietas yang signifikan pada tinggi tanaman, jumlah anakan, konsentrasi P pada tajuk. berat kering akar, konsentrasi P tajuk, sempan P akar. dan nisbah efisiensi P tajuk pada kondisi cukup dan kahat P.
Bohot kering tajuk dan hohot kering akar disusul jurnlah anakan merupakan peubah pertumbuhan yang paling sensitif terbadap delisiensi P sehingga
disaraukao sebagai kriteria penyaringan. IRRI (1996) menggunakan jurnlah
anakan relatif sebagai kriteria evaluasi ketenggangan padi tehadap defisiensi P.
Menggunakan kriteria jurnlah anakan relatif, Chaubey et al. (1994)
oleh efek gen aditif dan dominan dengan heritabilitas dalam arti sempit bemilai sedang (0,50). Swasti (2004) mend'p.tkan pengaruh gen-gen aditif, dominan,
serta interaksinya terhadap sifat efisien P pada padi dalam keadaan tercekam
aluminium, dengan nilai heritabilitas dalam arti sempit yang kecil. Wissua dan
Ae (200 I) mengidentifikasi emp.t lokus kuantitatif yang mengendalikan serapan
P padi pada tanah kurang P, dengan satu lokus utama yang terpaut dengan gen penanda pada kromosom 12.
Kompilasi hasil penelitian terhadap genetik ketenggangan tumbuhan atau
tanaman terhadap P rendah menunjukkan bahwa sifat ini dikendalikan gen secara
kuantitatif (Blum 1988). Hal ini dapat dimengerti karen. ketenggangan terhadap P rendah berhubungan dengan karakter kuantitatif yakni pertumbuhan akar.
Pernanfaatan sifat ketenggangan defisiensi P dalam pemuliaan padi juga
telah menunjukkan basil. Wisuw. dan Ae (200\), melalui seleksi yang dibantu
penand. molekular yang terpaut lokus kuantitatif penyerapan P yang tinggi pada
tanah defisien P, mendapatkan peningkstan penyerapan P sebesar 170% dan basil
EV ALUASI KETENGGANGAN GENOTIPE PADI
TERHADAPPRENDAHDITANAHSAWAH
Ab,trak
Suatu percobaan lapang telah dilakukan untuk mengetahui keragaman ketenggangan genotipe-genotipe padi terhadap P rendah serta mengidentifikasi genotipe yang tenggang. Percobaan dilaksanakan pada tanah sawah podzolik merah kekuningan dengan P tersedia dan P potensiai rendah. Rancangan percobaan yang digunakan adalah Petak Berbaris (Strip Plot) tiga ulangan, dengan pemupukan P (tanpa dan dengan pemupukan ) sebagai faktor horizontal dan plasmanutfah (120 genotipe) sebagai faktor vertikal. Pengamatan dilakukan terhadap tinggi tanaman, jumlah anakao, dan bobot keting tajuk. Ketenggangan genotipe ditetapkan berdasarkan nilai relatif yakni nilai pengamatan pada perlakuan tanpa pupuk dibagi dengan nilai pengamatan pada perlakuan dangan pupuk, dengan interval mengacu pada interval ketenggangan terhadap defisiensi P dari IRRl. Di samping ini juga diarnati serapan P tajuk pada masing-masing lima genotipe tenggang dan peka pada perlakuan tanpa pupuk P.
Hasil percobaan menunjukkan terdapat kemgaman fenotipik dan genotipik
ketenggangan di antara genotipe yang diuji. Ketenggangan berdasarkan jumlah anakan relatif menunjukkan keragaman genotipik tertinggi (28,7%). disusul oleh bobot kering relatif(22,6%) dan tinggi tanaman relatif(7,6%). Heritabilitas daiam arti luas tertinggi juga ditunjukkan oleh peubah jumlah anakan (24,1 %), dususul oleh tinggi tanaman (16,6%), dan bobot kering tajuk 15,0%). Teridentifikasi 14 genotipe sangat tenggang berdasarkan bobot keting tajuk relatif dan 28 genotipe berdasarkan jumlah anakan relatif. Ketenggaogan di lapang disehabkan oleh tingginya serapan P dan efisiensi penggtmaan internal pada kondisi rendah P.
Abstract
Pendahuluan
Defisiensi fosfor merupakan faktor pembatas basil yang paling penting dari
pertanaman padi pada tanah Ultisois, Oxisois, tanah sulfat masam, Andosols, dan bebernpa Vertisols (Ponnamperuma 1976). Tanah-tanah ini tidak hanya memiliki
fosfor tersedia イ・ョセ@ tetapi juga memfiksasi sebagian besar fosfor dari pupuk P
yang diberikan. sehingga dibutuhkan banyak pupuk untuk mendapatkan respon.
Delisiensi fosfor dapat dikendalikan dengan berbagai jalan. Salah satu
pendakatan yang efisien dan ekonomis adalah melalui pemuliaan atau perakitan
varietas yang dapat tumbub dan mengbasi1kan pada tanah delisien P (Ahloowalia
el al. 1994). Penapisan plasmanutfah adalah kegiatan untuk mengevaluasi
keragaman sifat yang dikehendaki mengidentilikasi genotipe yang adaptif pada
tanah defisien P. Kegiatan ini adalah langkah pertama dalam perakitan varietas padi yang elisien P.
Evaluasi ataupun seleksi untuk ketenggangan terhadap def'isiensi harn mineml biasanya dllakukan di lapang di mana tanahnya telah diketahui defisien
terhadap ham tertentu (Blum 1981). Untuk memisahkan antara potensi suatu
genotipe dengan pengaruh ketenggangan atau ketenggangan maka evaluasi hams
menggunakan percobaan faktorial dengan kondisi tercekam dan tidak. Hal ini
dilakukan dengan menanam pada tanah defisien dan menciptakan kondisi tidak tercekam melalui koreksi defisiensi dengan pemupukan.
Koleksi plasmanutfah yang banyak dan luas tidak banyak gunanya bagi
pemulia jika tidak dievaluasi seeara cukup (Hawkes 1981). Evaluasi koleksi plasmanutfah padi (terutama varietas lokal Indonesia) untuk. ketenggangan terhadap defisiensi fosfor dirasakan masih terbatas. Percobaan ini bertujuan untuk melihat keragaman ketenggangan plasmanutfah padi terutama varietas lokal terhadap defisiensi fosfoT pada tanah sawah dan mengidentifikasi genotipe-genotipe yang membawa sifat-sifat tenggang.
Bahan dan Metode
Percobaan dilaksanakan pada tanah sawah delisien P di Desa Sipak, Kecamatan Jasinga (Bogor) dari bulan Juli sampai September 2002 dengan jenis
20
adalah kadar P <200 ppm P,O, dengan pengekstrak 25% HCI (Nurjaya ef al. 1993) atau 5 ppm P (Olsen) (Doberrnann dan Fairhust 2000).
Rancangan pen:obaan yang digunakan adalah Petak Berbaris (Strip Plot)
dengan tiga ulangan. Petak horizontal terdiri dari dua tingkat pemupukan P dan petak vertikalnya terdiri dari 120 genotipe padi varielas lokal dari berbagai daerah, varielas unggul, galur padi tipe barn, dan kerabat liar padi (Lampiran I).
Tingkat pemupukan P yang digunakan adalah tanpa pupuk dan pupuk P
berdasarkan kebutuhan untuk mencapai kadar P larutan optimal bagi tanaman padi
(kebutoban P ekstemal). Kebutoban P ekstemal diduga melalui kurva erapan P yang didapatkan menurut prosedur Fox dan Kamprath (Wijaya-Adhi dan Sudjadi
1987).
Kurva erapan P didapatkan dengan menginkubasi 3 g tanah kering oven yang !elah ditambahkan 0, 50, 100, 200, 300, 400, 500 ppm P di dalarn 30 ml larutan 0,oJ M CaCh selarna enam hari. Selarna inkubasi dilakukan peugocokan 2 kali 30 menit setiap
hari.
Pada hari ke enam, P terlarut daIam supernatansuspensi diukur dengan metode spektrofotometri. Fosfor tererap adalah se1isih P
yang ditambahkan dengan P terlarut dalam supernatan. Kurva erapan didapatkan
dengan memplotkan logaritma P terlarut pada sumbu X dan P tererap pada sumbu Y.
Pemupukan P untuk mencapai kebutuhan P ekstema1 ditentukan dengan
memasnkkan nilai クセ@ 0,025 ke dalarn persarnaan (kurva) erapan. Nilai 0,025 adalah rataan kebutoban P ekstemal padi pada beberapa tanah sawah di Indonesia (Sri MuIyarti ef aJ. (993).
Bibit berumur 14 hari ditanam pada plot yang terdiri dari satu baris tanaman sepanjang 1 m dengan jarak tanarn 25x20 em, satu bibitJdapur. Pupuk dasar yang diberikan adalah 90 kg Nlha (200 kg Urealha) dan 120 kg K,OIha (200 kg KCllha). Pupuk Urea diberikan dua kali yakni umur satu minggu dan tiga minggu, masing-masiug seteugah bagian. Pupuk P dan K diberikan seluruhnya satu
miuggn setelah tanarn. Pengendalian terhadap hama, penyakit dan gulma dilakukan menurut rekomendasi.
Pengarnatan dilakukan terhadap jumlah anakan, tinggi tanaman, dan bobot
tanam. Kriteria ketenggangan genotipe terhadap defisiensi P yang digunakan
adalah nilai relatif dari jumlah anakan dan bobot kering tajuk dan tinggi tanaman,
yakni perbandingan nilai pengarnatan pada perlakuan tanpa pupuk P dengan nilai
pengarnatan pada perlakuan dengan pupuk P. Pengelompokan ketenggangan
mengacu pada kriteria yang digunakan IRRI (1996) yakni; sangat tenggang
(80-100%), tenggang (60-79'10), agak peka (40-59%), peka (20-39%), dan sanga! peka (0-19%). Anakan yang dihitung adalah anakan yang mempunyai minimal dna
daun yang telah berkembang sempurna. Tinggi tanaman diukur dan permukaan
tanah sampai ujung daun tertinggi. Bobot kering ditentukan dengan menimbang tajuk tanaman yang dipanen mulai dan dasar tanarnan (pennukaan tanah) dan dikeringkan dengan oven (70°C) selama 48 jam.
Hasil dan Pembahasan
Sifat Tanah Lokasi Percobaan
Ketepatan pemilihan lokasi pada kegiatan penapisan genotipe untuk
ketenggangan terhadap cekaman merupakan suatu yang penting untuk memberi basil tepat. Lokasi penapisan harus memiliki cekaman sesuai dengan tujuan penapisan. Hasil analisis tanah menunjukkan bahwa tanah lokasi percobaan di
lapang (Percobaan I) adalah sesuai untuk penapisan genotipe untuk ketenggangan
terhadap defisiensi P, terlihat dari kadar P tersedia (Bray I) maupun P potensial yang rendah (Tabell).
Fraksionasi P memperlihatkan bahwa fraksi P anorganik didominasi oleh Fe-P yang jaub lebih tinggi dibanding Ca-P dan AI-P. Dominannya fraksi P pada
tanah sawah adalah karena tetjadinya transformasi kalsium dan aluminium fosfat
menjadi besi fosfat (De Datta 1981). Fe-P adalah fraksi P tanah yang tergolong
dapat larut a1kali (alkali-soluble) sedangkan Ca-P tergolong dapat larut asam (acid
soluble) (Hedley el al. 1994).
Erapan P Tanah dan Kebutuban P Eksternal
Tanah lokasi pereobaan menunjukkan daya erap P yang cukup tinggi
(erapan maksimum Langmuir セ@ 520.185 ppm) dengan kurva erapan seperti pada
22
besi yang ditunjukkan oleh kadar Fe20, dan pH yang rendah (Tabel I). Hasil penelitian Singh dan Gilkes (1991) juga menunjukkan bahwa tanah-tanah dengan
kandungan oksida besi yang tinggi memiliki daya erap P yang tinggi.
Tabel 1. Ciri kimia dan fisik contoh tanah pada lokasi percobaan. eiri Kimia dan Fisik
PH H20 pHKCI C total (%) N total (%)
CIN
P Bray I (ppm) P HC125% (ppm) P total (ppm) P larutan (ppm) Ca-P (ppm) AI-P (ppm) Fe-P (ppm) K (me/IOO g) Ca (me/IOO g) Mg (me/IOO g) Na (mellOO g)
AI'+ (KCIIN)(meIlOOg)
W
(KCIIN) (meIlOOg) KTK (me/I 00 g) KB(%)Fe,O, (%) Pasir (%) Debu(%) Liat (%)
·Pusat Penelitian Tanah (1983)
Nilai Penetapan 4,6 3,9 1,77 0,14 12 0,13 32,31 175,08 0,019 2,62 3,49 36,68 0,13 14,65 2,19 tu 5,15 0,62 21,44 79 0,403 24 44 32
··kriteria tidak tersedia
Kriteria* Masam Sangat masam Rendah Rendah Sedang Sangat rendah Sangat rendah
••
Rendah Tinggi Tinggi Sangat rendah Sedang TinggiPendugaan kebutuhan P ekstemal melalui kurva semi log erapan (Gambar 3)
memberikan nilai 191,04 ppm, yang dibulatkan menjadi 200 ppm P atau 2.544 kg SP-36/ha. Dosis pemupukan P ini sangat tinggi dibandingkan dosis rekomendasi umum untuk tanah sawah. Pemberian pupuk P berdasarkan kebutuban eksternal memang membutuhkan dosis yang tinggi. Hasil penelitian Kasno e/ al. (1999)
[image:38.596.103.519.152.520.2]600
500
-E 400
セ@
"
セ@ 300•
セ@ 200"
100 -D-0
-100 0
0
0
o
- 0
0.05 0.1 0.15
P terlarut (ppm)
0.2 0.25
Gambar 2. Kurva erapan P contoh tanah lokasi percobaan
600
o
o
0 - - --...
---0.01
'J = 183.94ln(x) + 836.04
R2 '" 0.8155
0.1
Log P tenarut (ppm)
Gambar 3. Kurva semi log erapan P contoh tanah lokasi percobaan
Pengaruh Pemupukan P terhadap Pertumbuhan Tanaman
Kondisi Japang yang digunakan kondusif untuk penapisan genotipe, terlihat
dari adanya pengaruh nyata pemberian P dan genotipe pada tiga peubah yang diamati yakni tinggi taoaman, jumlah aoakan, bobot kering tajuk (Tabel 2).
Tabel 2. Kuadrat tengah pengaruh dosis P terhadap pertumbuban 120 genotipe
padi dengan dua dosis pemupukan di tanah sawah.
Kuadrat Tengah
Sumber Keragaman Tinggi Tanaman Anakan BK Tajuk
Dosis P (P) 75522,5* 7847,4' 55807,8**
Genotipe (G) 829,9*' 224,9** 661,6*'
GxP 129,9** 26,5* 114,8'"
m • • • tidak nyata, nyam, sangat nyata dengan uji F
24
Pengaruh pemberian P untuk masing-masing genotipe dapat dilihat pada
Lampiran 3-5. Perbedaan nyata pada pertumbuhan tanaman padi akibat pemberian
P pada tanah sawah berstatus P rendah adalah suatu gejaJa yang umum dijumpai (Srimulyani dan Wijaya-adhi 1993; Nurjaya et al. 1993). Namun demikian.
tingkat pengarub P tidal< sama pada ketiga peubab yang diamati. Berdasarkan
perbedaan nilai relatif pengamatan pada perlakuan dengan pemupukan dan tanpa pemupukan terlihat pengarub pemberian P lebih besar pada bobot kering tajuk.
disusul oleh jumlah anakan dan tinggi tanaman dengan penurunan berturut-turut
47,6,38,3, dan 21,7% (Gambar 4). Fageria e/ al. (1988) juga menemukan bahwa
bobot kering tajuk merupakan peubah pertumbuhan tanaman padi yang paling
peka terhadap defisiensi P dan menyarankan penggunaannya sebagai kriteria untuk seleksi ketenggangan terhadap defisiensi P.
120
100 100 100
100
セ@
セ@
i
セ@
400
P2 P1 p, P1 P2 P1
Bobct Kering Jumlah Anakan Tinggi T ancwnan
Gambar 4. Pengarub pemberian P terhadap pertumbuhan tanaman padi pada tanah sawah, 8 MST. (Rataan dan 120 genotipe, tiga ulangan; piセ@ tanpa pemberian P; P2= pemberian sesuai kebutuhan external
*
nilai pada PIIP2, pRセQPPEI@Sebaran Ketenggangan Genotipe
Terdapat perbedaan ketenggangan yang oyata di antara genotipe yang diuji terhadap defisiensi P yang dicenninkan oleh nilai relatif bobot kering, jumlah
anakan, dan tinggi tanaman (Lampiran 3-5). Narnun demikian, sebaran genotipe
menurut tingkat ketenggangan menunjukkan pola yang berbeda berdasarkan
peubah pertumbuhan yang diamati. (Gambar 5).
Pada Gambar 5 terlihat bahw. sebaran genotipe menurut ketenggangan
berdasarkan jumlah anakan lebih merata dibanding sebaran berdasarkan dua
kelompok agak peka, sedangkan sebaran menurut tinggi tanaman relatif adaJah
yang paling sempit dan terpusat pada kelompok sangat tenggang dan tenggang. Sempitnya sebaran genotipe berdasarkan tinggi tanaman relatif disebabkan kurang
pekanya peubah ini terbadap defisiensi P (Gambar 4).
60-, A 60- B 60
50j
4'
50
501l.40 ; セ@ 40- 1l.40
'5 '5 32 32 '5
c c c
&30
•
セ@ 30-28•
'"
30セ@ セ@ セ@
•
..
..
セRP@
E E..; 20
"
20,
,
10 10 10
0 0 0
[image:41.596.117.510.163.385.2]ST T AP P SP ST T AP P SP ST T AP P SP
Gambar 5. Sebaran 120 genotipe padi menurut tingkat ketenggangan terhadap defisiensi P pada tanab sawab berdasarkan nilai relatif BK tajuk (A), jumlab anakan (8), dan tinggi tanarnan (C). Hstセ@ sangat tengang,
80-100%; tセ@ tenggang, 60-80%; apセ@ agak peka, TPMVPE[pセ@ peka, 20-TPE[sセ@ sangat peka, 0-20%)
Koefisien ragarn fenotipik dan genotipik (Tabe! 3) juga menunjukkan nilai yang rendah untuk peubah tinggi tanaman, dan nilai yang cukup tinggi untuk. peubah jumlab anakan dan bobot kering tajuk. Koefisien ragarn genotipik yang
tinggi pada ketenggangan berdasarkan jumlab anakan dan bobot kering tajuk
relatif menunjukkan adanya peluang pemanfaatan plasmanutfah dalam perbaikan
genetik tanaman padj untuk ketenggangan terhadap P rendah di tanah sawah. Tabel3.
Peubah
Tinggi
Anakan
Ragam dan koefisien ragam fenotipik dan genotipik ketenggangan 120 genotipe padi terhad.p P rendab di tanab sawab berdasarkan nilai rel.fif tiga peubab pertumbuban
Ragam Ragam
Fenotipik Genotipik
202,7 33,7
1221,3 294,3
KoefRagam
Fenotipik (%)
18, I
Koef. Ragam Genotipik (%)
7,6
28,7
16,6
24,1
B.K. Tajuk 921,3 138,3
58,4
58,4 22,6 15,0
26
Semua genotipe sangat tenggang berdasarkan hobot kering tajuk relatif
tergolong pada kultivar lokal dan kerabat liar (Tabel 4). Hal ini diduga adalalt
sebagai hasil dari proses seleksi yang dialami oteh genotipe-genotipe tersebut